

Açai Berry Administration Promotes Wound Healing through Wnt/β-Catenin Pathway

 , ,
, ,  ,
,  ,
,  ,
,  and
and 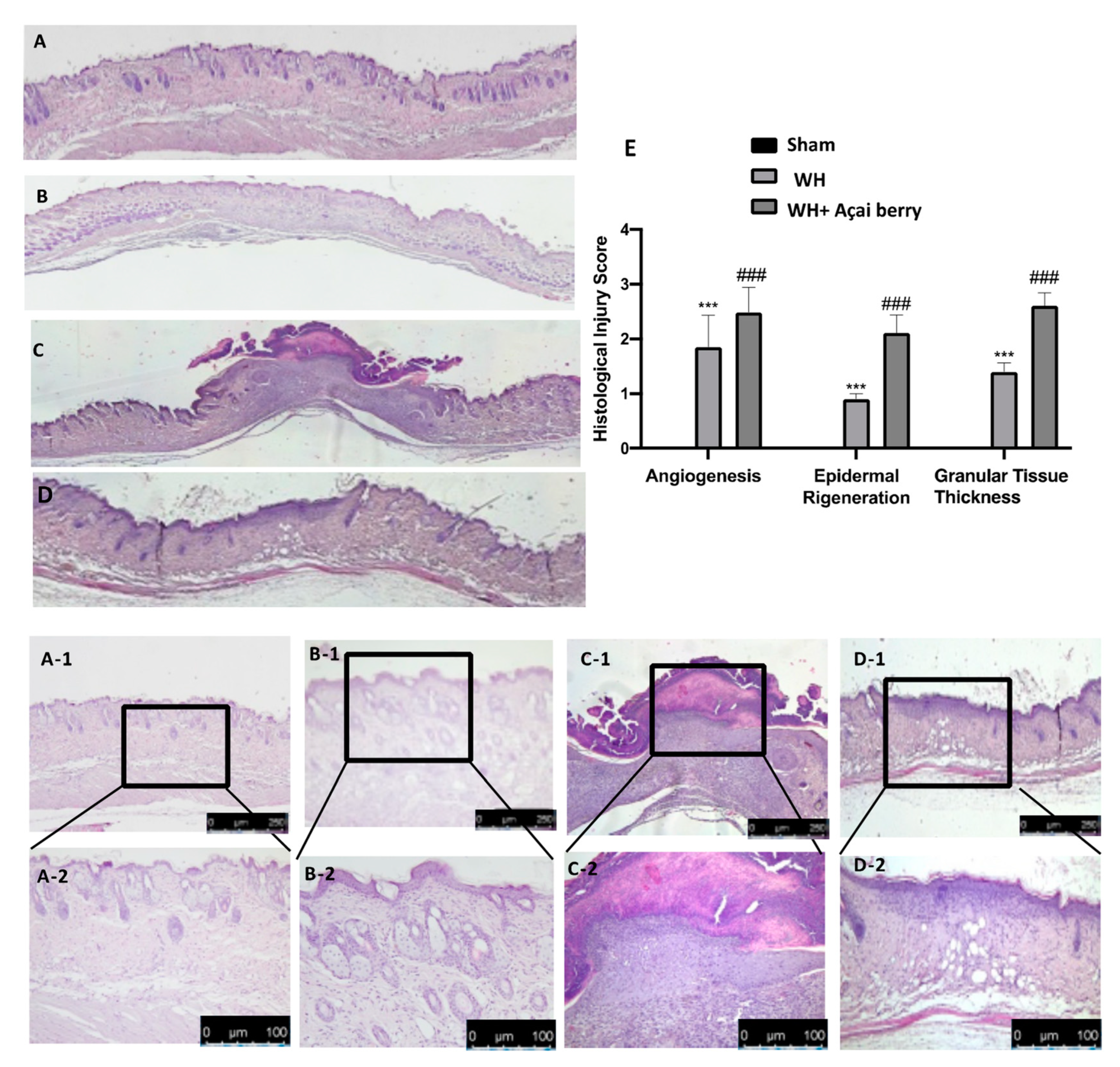{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Açai Berry on Tissue Repair
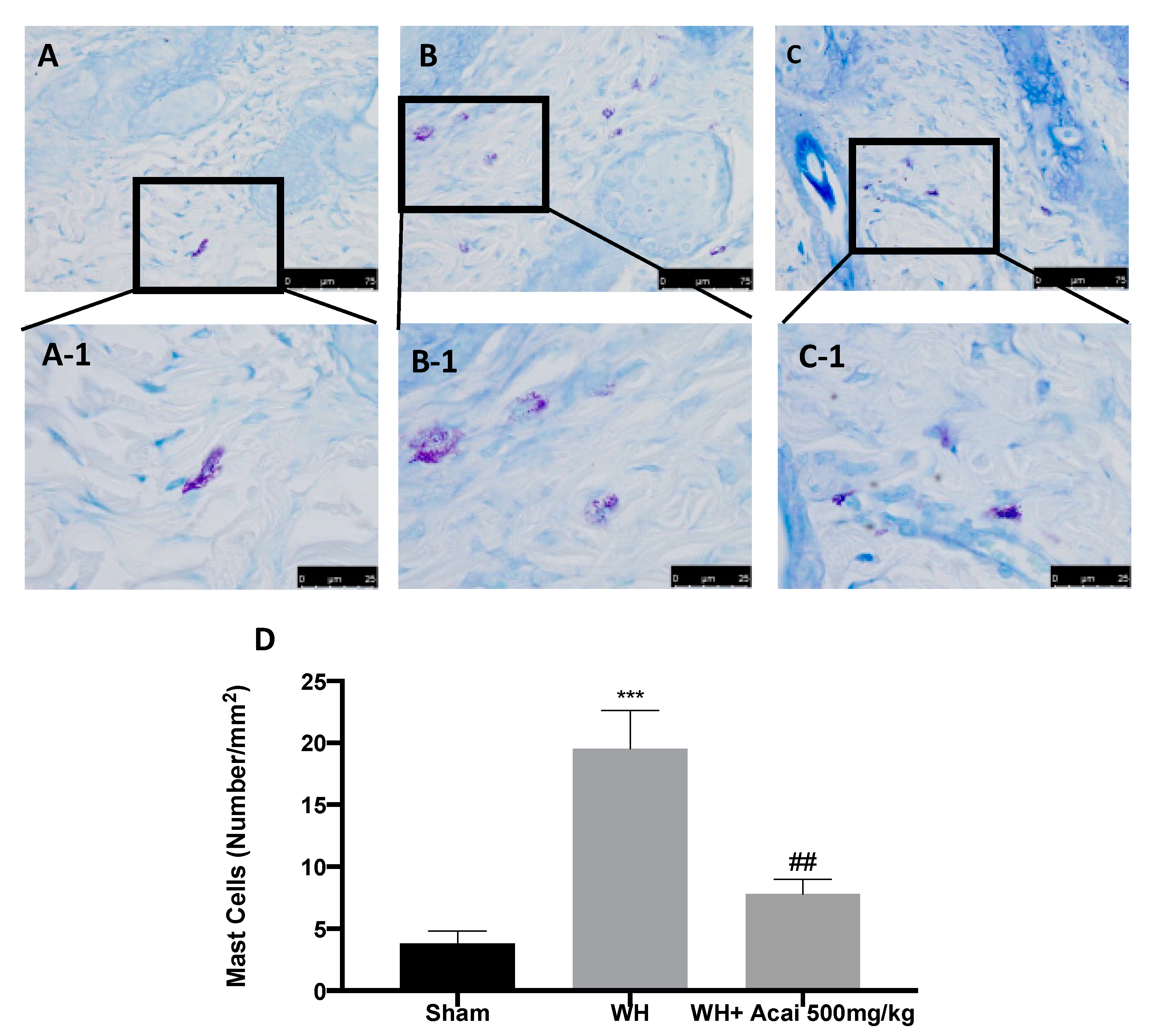
2.2. Açai Berry Effect on Fibrosis Condition
2.3. Effect of Acai on ICAM-1 and P-Selectin Expression
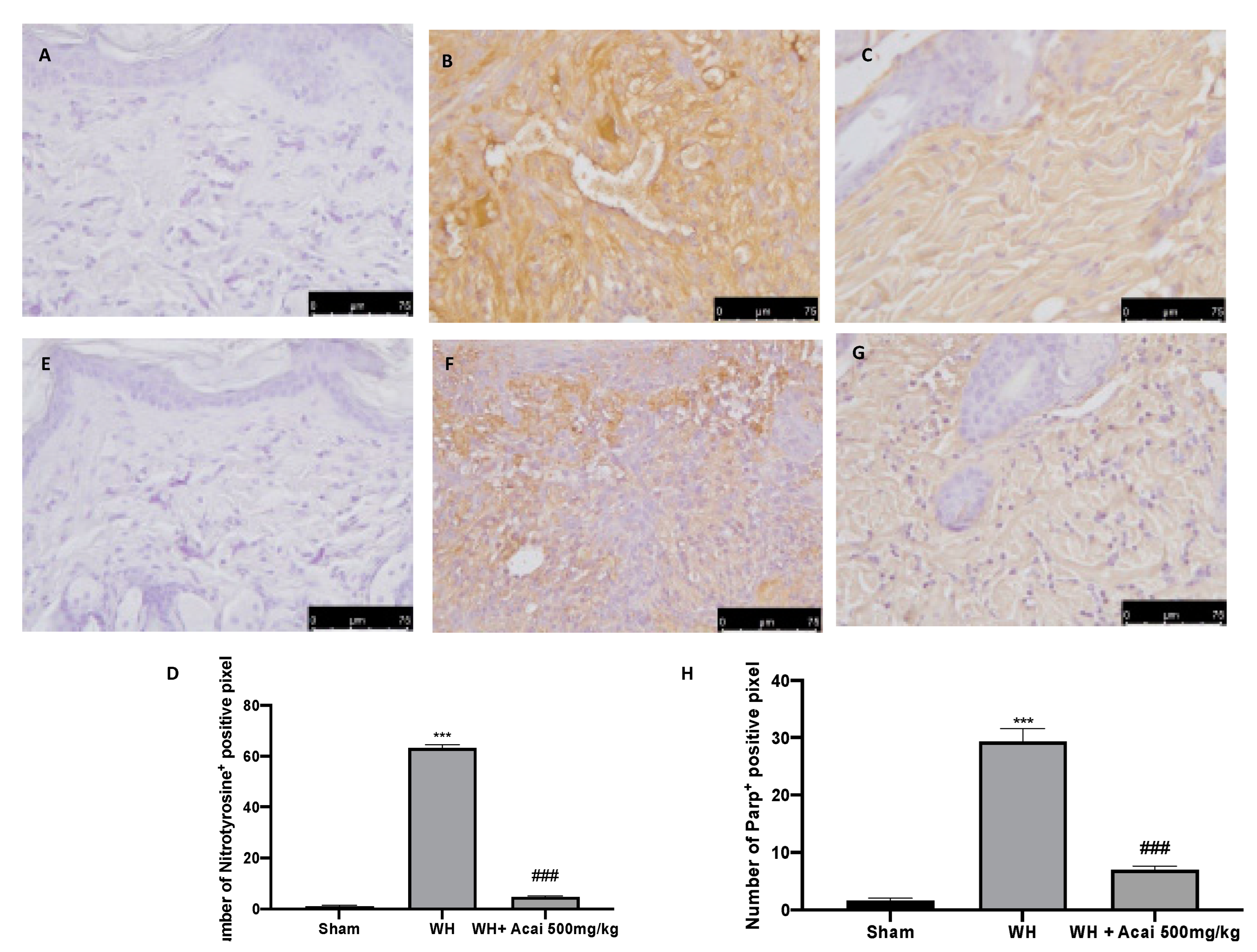
2.4. Effect of Acai on Nitrotyrosine and Parp-1 Expression
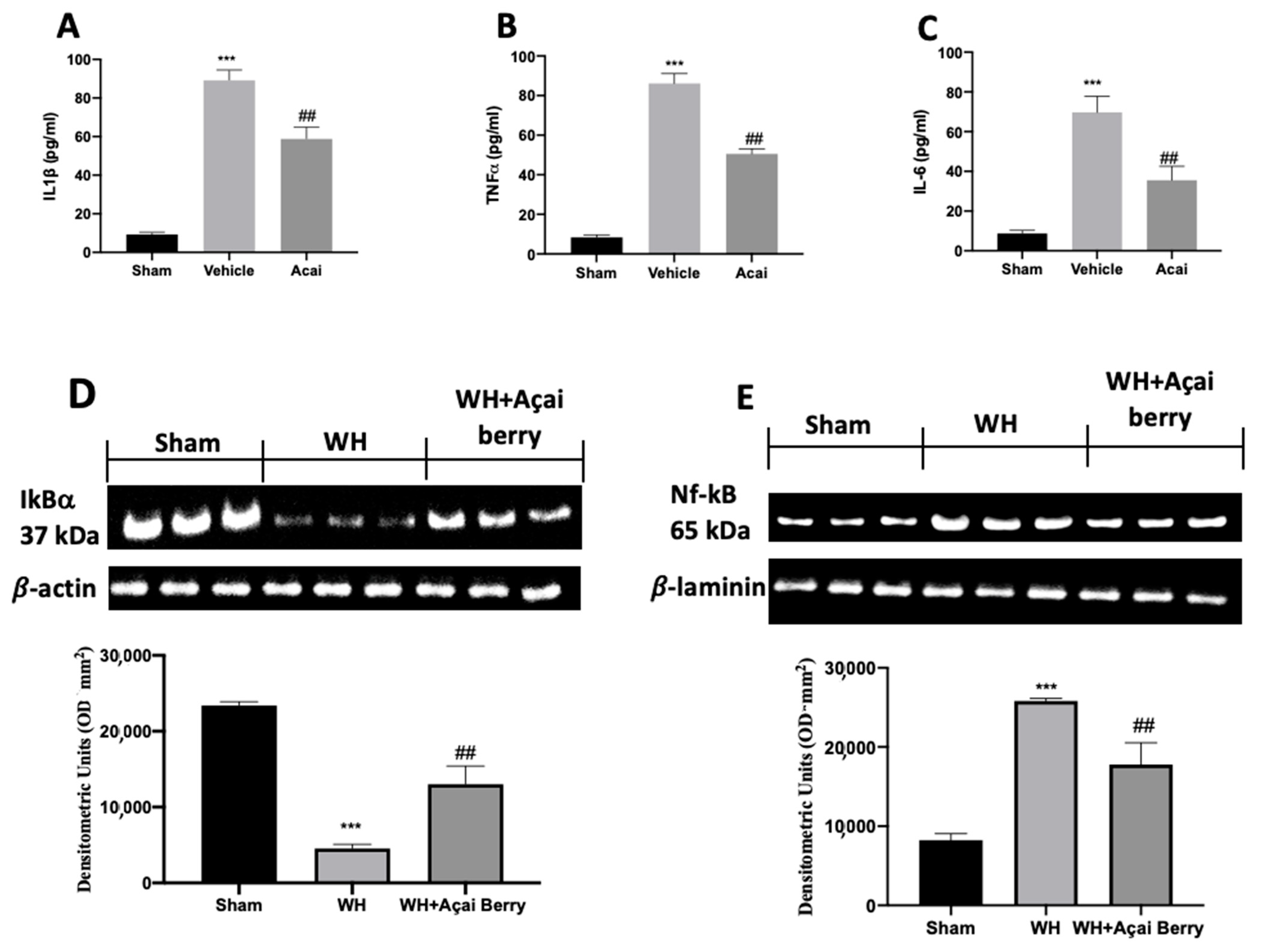
2.5. Effect of Açai Berry on Inflammation
2.6. Effect of Açai Berry Administration on Wnt/FZ/β-Catenin Pathway
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of Experimental Wound Healing
4.3. Experimental Groups
- -
- WH group: mice were subjected to full-thickness excisional wounds as described above.
- -
- WH + açai berry group: mice received oral administration of açai berry (500 mg/kg dissolved in saline) for 6 days after induction of the wound.
- -
- Sham group: mice were subjected to all procedures described above, except that the full-thickness excisional wounds were not applied and saline was administered for 6 days after.
- -
- Sham + açai berry group: mice were subjected to all procedures described above, except that the full-thickness excisional wounds were not applied and saline with açai berry was administered (500 mg/kg dissolved in saline) for 6 days after.
4.4. Histological Analysis
4.5. Immunohistochemical Localization of VEGF, ICAM-1, P-Selectin, Nitro, and Par
4.6. Western Blot
4.7. Elisa
4.8. Materials
4.9. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gugliandolo, E.; Macrì, F.; Fusco, R.; Siracusa, R.; D’Amico, R.; Cordaro, M.; Peritore, A.F.; Impellizzeri, D.; Genovese, T.; Cuzzocrea, S. The Protective Effect of Snail Secretion Filtrate in an Experimental Model of Excisional Wounds in Mice. Vet. Sci. 2021, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, J.; Kirsner, R. Pathophysiology of acute wound healing. Clin. Dermatol. 2007, 25, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Siracusa, R.; Monaco, F.; D’Amico, R.; Genovese, T.; Cordaro, M.; Interdonato, L.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Cuzzocrea, S. Epigallocatechin-3-gallate modulates postoperative pain by regulating biochemical and molecular pathways. Int. J. Mol. Sci. 2021, 22, 6879. [Google Scholar] [CrossRef] [PubMed]
- Bejsovec, A. Wnt pathway activation: New relations and locations. Cell 2005, 120, 11–14. [Google Scholar] [PubMed] [Green Version]
- Li, Y.; Baccouche, B.; Olayinka, O.; Serikbaeva, A.; Kazlauskas, A. The Role of the Wnt Pathway in VEGF/Anti-VEGF-Dependent Control of the Endothelial Cell Barrier. Investig. Ophthalmol. Vis. Sci. 2021, 62, 17. [Google Scholar] [CrossRef]
- Burgy, O.; Königshoff, M. The WNT signaling pathways in wound healing and fibrosis. Matrix Biol. 2018, 68, 67–80. [Google Scholar] [CrossRef]
- Whyte, J.L.; Smith, A.A.; Liu, B.; Manzano, W.R.; Evans, N.D.; Dhamdhere, G.R.; Fang, M.Y.; Chang, H.Y.; Oro, A.E.; Helms, J.A. Augmenting endogenous Wnt signaling improves skin wound healing. PLoS ONE 2013, 8, e76883. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sang, A.; Zhu, M.; Zhang, G.; Guan, H.; Ji, M.; Chen, H. Tissue factor induces VEGF expression via activation of the Wnt/β-catenin signaling pathway in ARPE-19 cells. Mol. Vis. 2016, 22, 886. [Google Scholar]
- Mi, Y.; Zhong, L.; Lu, S.; Hu, P.; Pan, Y.; Ma, X.; Yan, B.; Wei, Z.; Yang, G. Quercetin promotes cutaneous wound healing in mice through Wnt/β-catenin signaling pathway. J. Ethnopharmacol. 2022, 290, 115066. [Google Scholar] [CrossRef]
- Lichtman, M.K.; Otero-Vinas, M.; Falanga, V. Transforming growth factor beta (TGF-β) isoforms in wound healing and fibrosis. Wound Repair Regen. 2016, 24, 215–222. [Google Scholar] [CrossRef]
- Akhmetshina, A.; Palumbo, K.; Dees, C.; Bergmann, C.; Venalis, P.; Zerr, P.; Horn, A.; Kireva, T.; Beyer, C.; Zwerina, J. Activation of canonical Wnt signalling is required for TGF-β-mediated fibrosis. Nat. Commun. 2012, 3, 735. [Google Scholar] [CrossRef] [PubMed]
- Edara, V.V.; Nooka, S.; Proulx, J.; Stacy, S.; Ghorpade, A.; Borgmann, K. β-Catenin regulates wound healing and IL-6 expression in activated human astrocytes. Biomedicines 2020, 8, 479. [Google Scholar] [CrossRef] [PubMed]
- Svechnikov, K.; Izzo, G.; Landreh, L.; Weisser, J.; Söder, O. Endocrine disruptors and Leydig cell function. J. Biomed. Biotechnol. 2010, 2010, 684504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holderness, J.; Schepetkin, I.A.; Freedman, B.; Kirpotina, L.N.; Quinn, M.T.; Hedges, J.F.; Jutila, M.A. Polysaccharides isolated from Acai fruit induce innate immune responses. PLoS ONE 2011, 6, e17301. [Google Scholar] [CrossRef]
- Kang, J.; Xie, C.; Li, Z.; Nagarajan, S.; Schauss, A.G.; Wu, T.; Wu, X. Flavonoids from acai (Euterpe oleracea Mart.) pulp and their antioxidant and anti-inflammatory activities. Food Chem. 2011, 128, 152–157. [Google Scholar] [CrossRef]
- Genovese, T.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Peritore, A.F.; Crupi, R.; Interdonato, L.; Gugliandolo, E.; Cuzzocrea, S.; Di Paola, R. Açaí (Euterpe oleraceae Mart.) Seeds Regulate NF-κB and Nrf2/ARE Pathways Protecting Lung against Acute and Chronic Inflammation. Cell. Physiol. Biochem. 2022, 56 (Suppl. S3), 1–20. [Google Scholar] [CrossRef]
- Cohen-Armon, M. PARP-1 activation in the ERK signaling pathway. Trends Pharmacol. Sci. 2007, 28, 556–560. [Google Scholar] [CrossRef]
- Siracusa, R.; Impellizzeri, D.; Cordaro, M.; Gugliandolo, E.; Peritore, A.F.; Di Paola, R.; Cuzzocrea, S. Topical application of adelmidrol+ trans-traumatic acid enhances skin wound healing in a streptozotocin-induced diabetic mouse model. Front. Pharmacol. 2018, 9, 871. [Google Scholar] [CrossRef]
- Clark, R.A. Regulation of fibroplasia in cutaneous wound repair. Am. J. Med. Sci. 1993, 306, 42–48. [Google Scholar] [CrossRef]
- Wallace, H.A.; Basehore, B.M.; Zito, P.M. Wound Healing Phases; StatPearls Publishing: Treasure Island, FL, USA, 2017. [Google Scholar]
- Kim, Y.-K.; Hyun, K.-Y.; Lee, M.-K. Anti-Inflammatory and antioxidative effects of acaiberry in formalin-induced orofacial pain in rats. J. Dent. Hyg. Sci. 2014, 14, 240–247. [Google Scholar]
- Koh, T.J.; DiPietro, L.A. Inflammation and wound healing: The role of the macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutsaers, S.E.; Bishop, J.E.; McGrouther, G.; Laurent, G.J. Mechanisms of tissue repair: From wound healing to fibrosis. Int. J. Biochem. Cell Biol. 1997, 29, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, G.S.; Horan, M.A.; Ferguson, M. Aging alters the inflammatory and endothelial cell adhesion molecule profiles during human cutaneous wound healing. Lab. Investig. A J. Tech. Methods Pathol. 1998, 78, 47–58. [Google Scholar]
- Cano Sanchez, M.; Lancel, S.; Boulanger, E.; Neviere, R. Targeting oxidative stress and mitochondrial dysfunction in the treatment of impaired wound healing: A systematic review. Antioxidants 2018, 7, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandookwala, M.; Sengupta, P. 3-Nitrotyrosine: A versatile oxidative stress biomarker for major neurodegenerative diseases. Int. J. Neurosci. 2020, 130, 1047–1062. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Patel, D.; Sen, S.; Shanmugam, V.; Sidawy, A.; Mishra, L.; Nguyen, B.-N. Poly-ADP-ribose polymerase inhibition enhances ischemic and diabetic wound healing by promoting angiogenesis. J. Vasc. Surg. 2017, 65, 1161–1169. [Google Scholar] [CrossRef] [Green Version]
- Nall, A.V.; Brownlee, R.E.; Colvin, C.P.; Schultz, G.; Fein, D.; Cassisi, N.J.; Nguyen, T.; Kalra, A. Transforming growth factor β1 improves wound healing and random flap survival in normal and irradiated rats. Arch. Otolaryngol. Head Neck Surg. 1996, 122, 171–177. [Google Scholar] [CrossRef]
- Nissen, N.N.; Polverini, P.; Koch, A.E.; Volin, M.V.; Gamelli, R.L.; DiPietro, L.A. Vascular endothelial growth factor mediates angiogenic activity during the proliferative phase of wound healing. Am. J. Pathol. 1998, 152, 1445. [Google Scholar]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Moon, R.T. Wnt/β-catenin pathway. Sci. STKE 2005, 2005, cm1. [Google Scholar] [CrossRef]
- Du, Q.; Geller, D. Cross-regulation between Wnt and NF-κB signaling pathways. Onco Ther. 2010, 1, 155–181. [Google Scholar] [CrossRef]
- Houschyar, K.S.; Momeni, A.; Pyles, M.N.; Maan, Z.N.; Whittam, A.J.; Siemers, F. Wnt signaling induces epithelial differentiation during cutaneous wound healing. Organogenesis 2015, 11, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaro, M.; Fusco, R.; D’Amico, R.; Siracusa, R.; Peritore, A.F.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Mandalari, G.; Cuzzocrea, S. Cashew (Anacardium occidentale L.) Nuts modulate the Nrf2 and NLRP3 pathways in pancreas and lung after induction of acute pancreatitis by cerulein. Antioxidants 2020, 9, 992. [Google Scholar] [CrossRef] [PubMed]
- Interdonato, L.; D’amico, R.; Cordaro, M.; Siracusa, R.; Fusco, R.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Coaccioli, S.; Genovese, T.; et al. Aerosol-Administered Adelmidrol Attenuates Lung Inflammation in a Murine Model of Acute Lung Injury. Biomolecules 2022, 12, 1308. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Fusco, R.; Gugliandolo, E.; Siracusa, R.; Scuto, M.; Cordaro, M.; D’Amico, R.; Evangelista, M.; Peli, A.; Peritore, A.F.; Impellizzeri, D. Formyl peptide receptor 1 signaling in acute inflammation and neural differentiation induced by traumatic brain injury. Biology 2020, 9, 238. [Google Scholar] [CrossRef]
- Fusco, R.; Cordaro, M.; Siracusa, R.; D’Amico, R.; Genovese, T.; Gugliandolo, E.; Peritore, A.F.; Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Biochemical evaluation of the antioxidant effects of hydroxytyrosol on pancreatitis-associated gut injury. Antioxidants 2020, 9, 781. [Google Scholar] [CrossRef]
- Siracusa, R.; Scuto, M.; Fusco, R.; Trovato, A.; Ontario, M.L.; Crea, R.; Di Paola, R.; Cuzzocrea, S.; Calabrese, V. Anti-inflammatory and anti-oxidant activity of Hidrox® in rotenone-induced Parkinson’s disease in mice. Antioxidants 2020, 9, 824. [Google Scholar] [CrossRef]
- D’Amico, R.; Fusco, R.; Siracusa, R.; Impellizzeri, D.; Peritore, A.F.; Gugliandolo, E.; Interdonato, L.; Sforza, A.M.; Crupi, R.; Cuzzocrea, S. Inhibition of P2X7 purinergic receptor ameliorates fibromyalgia syndrome by suppressing NLRP3 pathway. Int. J. Mol. Sci. 2021, 22, 6471. [Google Scholar] [CrossRef]
- Impellizzeri, D.; D’Amico, R.; Fusco, R.; Genovese, T.; Peritore, A.F.; Gugliandolo, E.; Crupi, R.; Interdonato, L.; Di Paola, D.; Di Paola, R. Acai Berry Mitigates Vascular Dementia-Induced Neuropathological Alterations Modulating Nrf-2/Beclin1 Pathways. Cells 2022, 11, 2616. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Interdonato, L.; Marino, Y.; Franco, G.A.; Arangia, A.; D’Amico, R.; Siracusa, R.; Cordaro, M.; Impellizzeri, D.; Fusco, R.; Cuzzocrea, S.; et al. Açai Berry Administration Promotes Wound Healing through Wnt/β-Catenin Pathway. Int. J. Mol. Sci. 2023, 24, 834. https://doi.org/10.3390/ijms24010834
Interdonato L, Marino Y, Franco GA, Arangia A, D’Amico R, Siracusa R, Cordaro M, Impellizzeri D, Fusco R, Cuzzocrea S, et al. Açai Berry Administration Promotes Wound Healing through Wnt/β-Catenin Pathway. International Journal of Molecular Sciences. 2023; 24(1):834. https://doi.org/10.3390/ijms24010834
Chicago/Turabian StyleInterdonato, Livia, Ylenia Marino, Gianluca Antonio Franco, Alessia Arangia, Ramona D’Amico, Rosalba Siracusa, Marika Cordaro, Daniela Impellizzeri, Roberta Fusco, Salvatore Cuzzocrea, and et al. 2023. "Açai Berry Administration Promotes Wound Healing through Wnt/β-Catenin Pathway" International Journal of Molecular Sciences 24, no. 1: 834. https://doi.org/10.3390/ijms24010834
APA StyleInterdonato, L., Marino, Y., Franco, G. A., Arangia, A., D’Amico, R., Siracusa, R., Cordaro, M., Impellizzeri, D., Fusco, R., Cuzzocrea, S., & Paola, R. D. (2023). Açai Berry Administration Promotes Wound Healing through Wnt/β-Catenin Pathway. International Journal of Molecular Sciences, 24(1), 834. https://doi.org/10.3390/ijms24010834