
Anti-Inflammatory Effects of Cannabigerol in Rheumatoid Arthritis Synovial Fibroblasts and Peripheral Blood Mononuclear Cell Cultures Are Partly Mediated by TRPA1

Abstract
:1. Introduction
2. Results
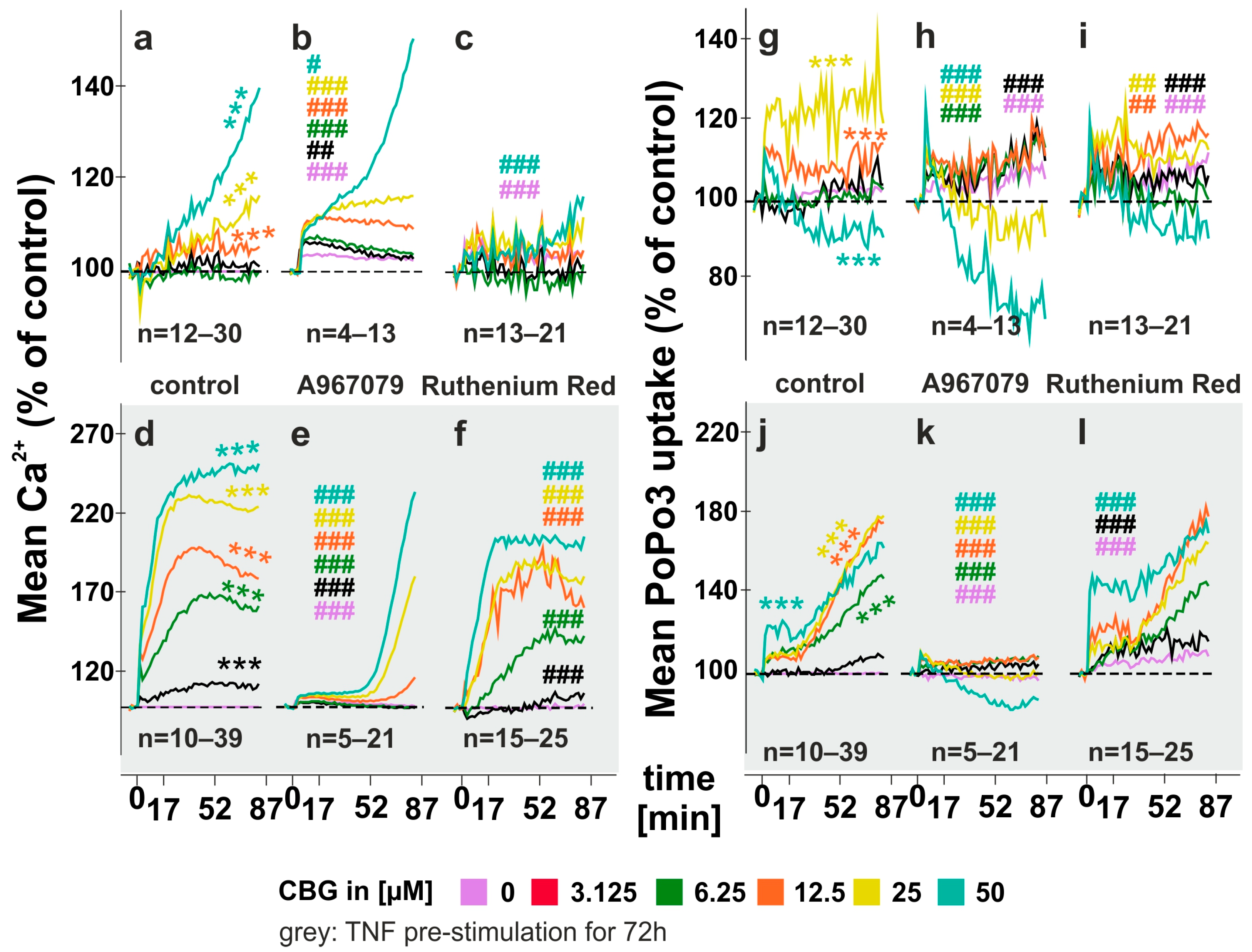
2.1. CBG Increases Intracellular Calcium Levels and Compound Uptake
2.2. Modulation of CBG-Induced Calcium Levels and PoPo3 Uptake in TNF Stimulated RASF
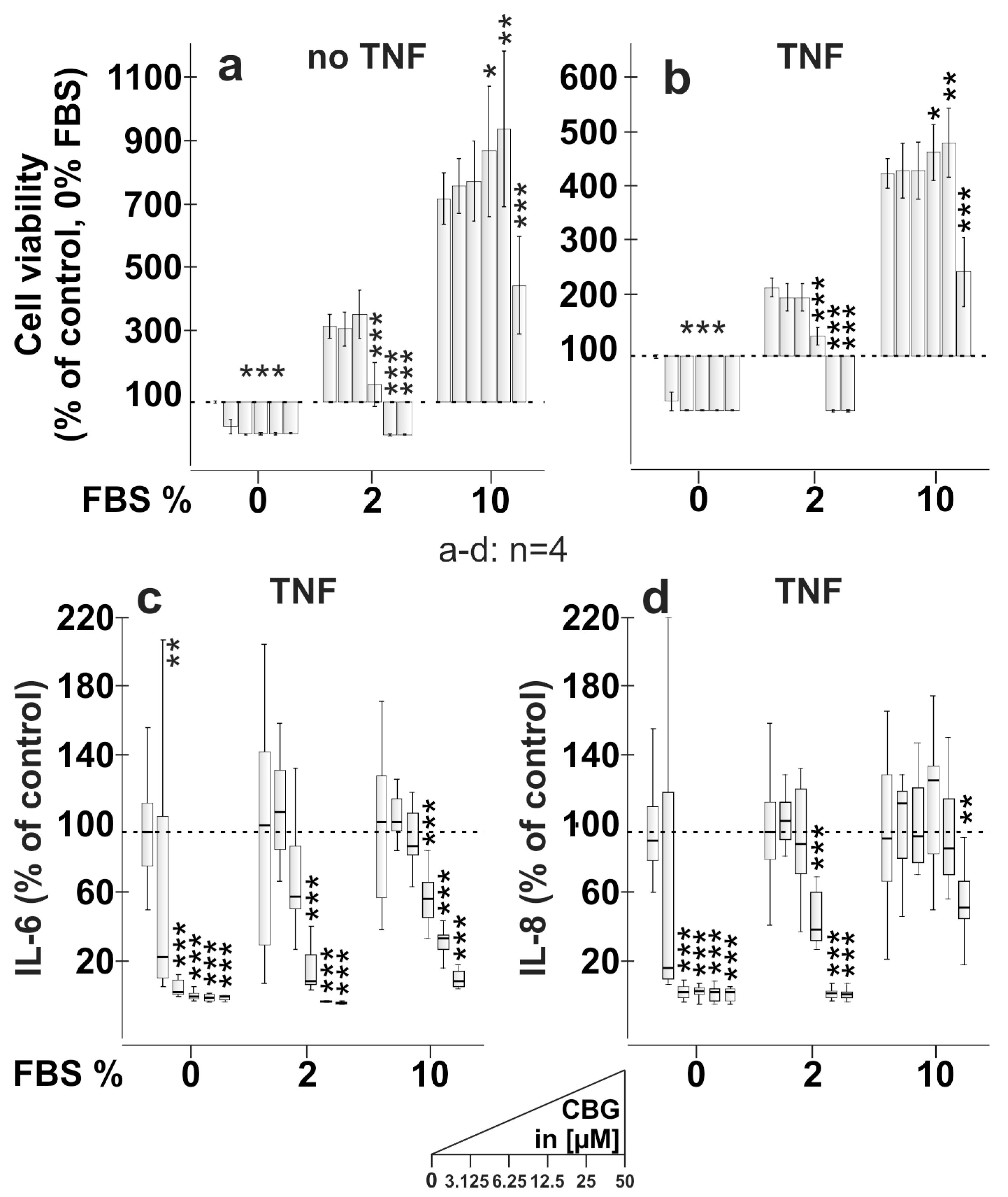
2.3. CBG Reduces Cell Viability, IL-6 and IL-8 Production Dependent on FBS Content
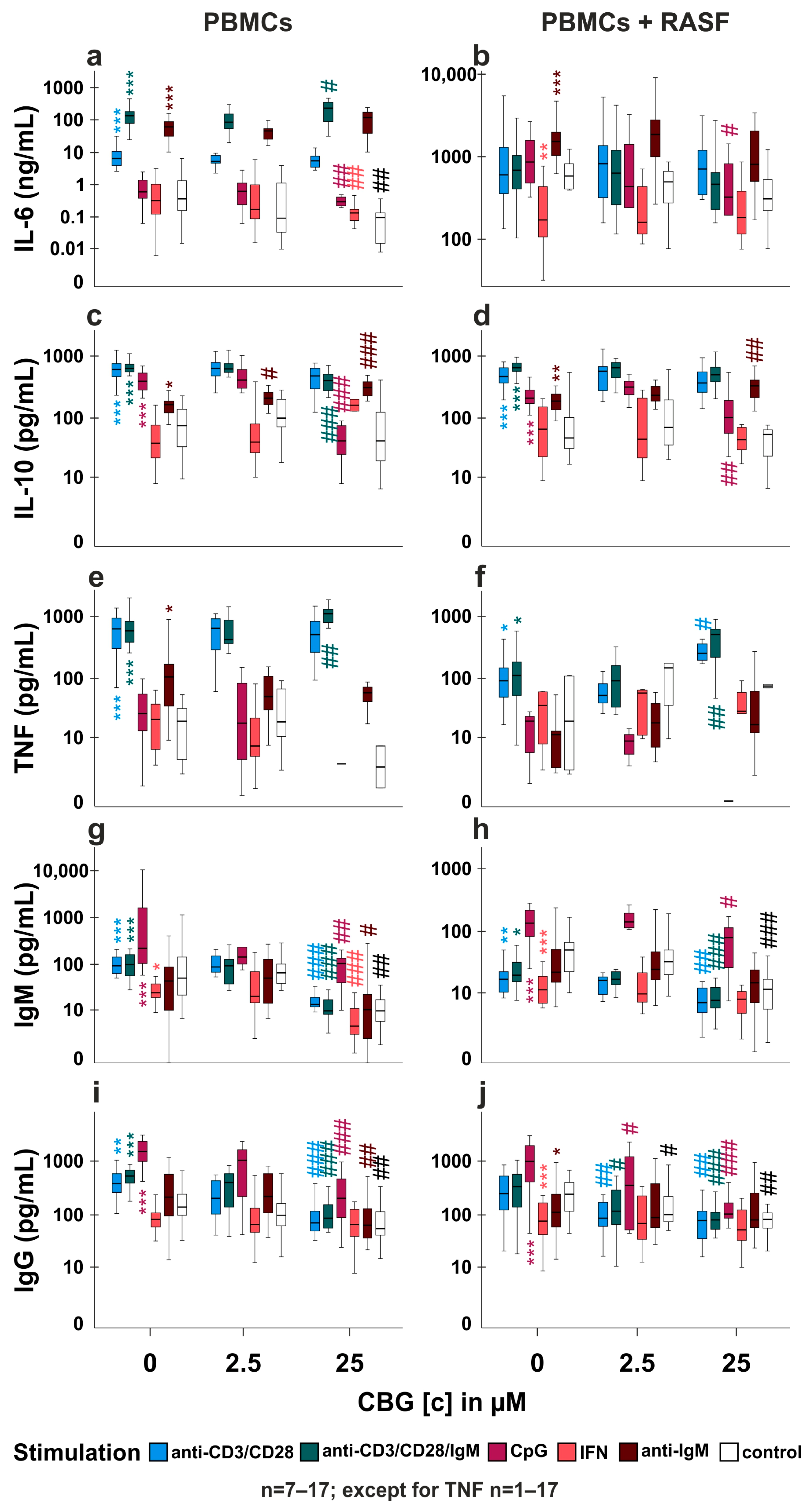
2.4. CBG Modulates Cytokine and Immunoglobulin Production in PBMC and RASF/PBMC Co-Cultures
2.5. Is TRPA1 Involved in the Regulation of Cytokine and Immunoglobulin Production by CBG?
3. Discussion
3.1. CBG Modulates Intracellular Calcium and PoPo3 Uptake via TRPA1 (Figure 1 and Figure 2)
3.2. CBG Reduces Cell Viability and Cytokine Production in an FBS-Dependent Fashion (Figure 3)
3.3. Influence of CBG on PBMC and PBMC/RASF Co-Culture (Figure 4a–f)
3.4. Does CBG Act on TRPA1 in PBMCs? (Figure 5a,b,d,e,f,h)
3.5. TRPA1 Influences Antibody Production (Figure 4g–j, Figure 5c,g and Figure S2)
4. Materials and Methods
4.1. Patients
4.2. Compounds and Antibodies
4.3. Synovial Tissue Preparation and SFs Culture
4.4. Isolation of PBMCs from Peripheral Blood
4.5. RASF Co-Culture with PBMCs
4.6. IL-6, IL-8, IL-10 and TNF ELISA and Stimulation of SF
4.7. IgM and IgG ELISA
4.8. Intracellular Calcium and PoPo3 Uptake
4.9. Cell Viability Assay
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lowin, T.; Schneider, M.; Pongratz, G. Joints for joints: Cannabinoids in the treatment of rheumatoid arthritis. Curr. Opin. Rheumatol. 2019, 31, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Vimal, D.; D′Souza, L.C.; Rai, V.; Lal, S.; Sharma, A.; Gupta, S.C. Efficacy of cannabis and its constituents in disease management: Insights from clinical studies. Curr. Med. Chem. 2022. [Google Scholar] [CrossRef]
- Jastrzab, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. The Origin and Biomedical Relevance of Cannabigerol. Int. J. Mol. Sci. 2022, 23, 7929. [Google Scholar] [CrossRef] [PubMed]
- Walsh, K.B.; McKinney, A.E.; Holmes, A.E. Minor Cannabinoids: Biosynthesis, Molecular Pharmacology and Potential Therapeutic Uses. Front. Pharmacol. 2021, 12, 777804. [Google Scholar] [CrossRef] [PubMed]
- Almoallim, H.; Al Saleh, J.; Badsha, H.; Ahmed, H.M.; Habjoka, S.; Menassa, J.A.; El-Garf, A. A Review of the Prevalence and Unmet Needs in the Management of Rheumatoid Arthritis in Africa and the Middle East. Rheumatol. Ther. 2021, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Knab, K.; Chambers, D.; Kronke, G. Synovial Macrophage and Fibroblast Heterogeneity in Joint Homeostasis and Inflammation. Front. Med. 2022, 9, 862161. [Google Scholar] [CrossRef]
- Lowin, T.; Tingting, R.; Zurmahr, J.; Classen, T.; Schneider, M.; Pongratz, G. Cannabidiol (CBD): A killer for inflammatory rheumatoid arthritis synovial fibroblasts. Cell Death Dis. 2020, 11, 714. [Google Scholar] [CrossRef]
- Lowin, T.; Bleck, J.; Schneider, M.; Pongratz, G. Selective killing of proinflammatory synovial fibroblasts via activation of transient receptor potential ankyrin (TRPA1). Biochem. Pharmacol. 2018, 154, 293–302. [Google Scholar] [CrossRef]
- Story, G.M.; Peier, A.M.; Reeve, A.J.; Eid, S.R.; Mosbacher, J.; Hricik, T.R.; Earley, T.J.; Hergarden, A.C.; Andersson, D.A.; Hwang, S.W.; et al. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Kochukov, M.Y.; McNearney, T.A.; Fu, Y.; Westlund, K.N. Thermosensitive TRP ion channels mediate cytosolic calcium response in human synoviocytes. Am. J. Physiol. Cell Physiol. 2006, 291, C424–C432. [Google Scholar] [CrossRef]
- Takahashi, N.; Chen, H.Y.; Harris, I.S.; Stover, D.G.; Selfors, L.M.; Bronson, R.T.; Deraedt, T.; Cichowski, K.; Welm, A.L.; Mori, Y.; et al. Cancer Cells Co-opt the Neuronal Redox-Sensing Channel TRPA1 to Promote Oxidative-Stress Tolerance. Cancer Cell 2018, 33, 985–1003.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowin, T.; Kok, C.; Smutny, S.; Pongratz, G. Impact of Delta(9)-Tetrahydrocannabinol on Rheumatoid Arthritis Synovial Fibroblasts Alone and in Co-Culture with Peripheral Blood Mononuclear Cells. Biomedicines 2022, 10, 1118. [Google Scholar] [CrossRef] [PubMed]
- Nachnani, R.; Raup-Konsavage, W.M.; Vrana, K.E. The Pharmacological Case for Cannabigerol. J. Pharmacol. Exp. Ther. 2021, 376, 204–212. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Vellani, V.; Schiano-Moriello, A.; Marini, P.; Magherini, P.C.; Orlando, P.; Di Marzo, V. Plant-derived cannabinoids modulate the activity of transient receptor potential channels of ankyrin type-1 and melastatin type-8. J. Pharmacol. Exp. Ther. 2008, 325, 1007–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allara, M.; Bisogno, T.; Petrosino, S.; Stott, C.G.; Di Marzo, V. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef] [Green Version]
- Atakpa, P.; van Marrewijk, L.M.; Apta-Smith, M.; Chakraborty, S.; Taylor, C.W. GPN does not release lysosomal Ca(2+) but evokes Ca(2+) release from the ER by increasing the cytosolic pH independently of cathepsin C. J. Cell Sci. 2019, 132, jcs223883. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.J.; Jacob, R. Ionomycin enhances Ca2+ influx by stimulating store-regulated cation entry and not by a direct action at the plasma membrane. Biochem. J. 1994, 300, 665–672. [Google Scholar] [CrossRef] [Green Version]
- De Petrocellis, L.; Orlando, P.; Moriello, A.S.; Aviello, G.; Stott, C.; Izzo, A.A.; Di Marzo, V. Cannabinoid actions at TRPV channels: Effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol. 2012, 204, 255–266. [Google Scholar] [CrossRef]
- Lukyanenko, V.; Gyorke, I.; Subramanian, S.; Smirnov, A.; Wiesner, T.F.; Gyorke, S. Inhibition of Ca(2+) sparks by ruthenium red in permeabilized rat ventricular myocytes. Biophys. J. 2000, 79, 1273–1284. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, C.A.; Brody, J.S. Ultrastructural localization and characterization of proteoglycans in the pulmonary alveolus. Am. Rev. Respir. Dis. 1979, 120, 901–910. [Google Scholar] [CrossRef]
- Elmes, M.W.; Kaczocha, M.; Berger, W.T.; Leung, K.; Ralph, B.P.; Wang, L.; Sweeney, J.M.; Miyauchi, J.T.; Tsirka, S.E.; Ojima, I.; et al. Fatty acid-binding proteins (FABPs) are intracellular carriers for Delta9-tetrahydrocannabinol (THC) and cannabidiol (CBD). J. Biol. Chem. 2015, 290, 8711–8721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organic Cation Transporters. In Encyclopedic Reference of Molecular Pharmacology; Springer: Berlin/Heidelberg, Germany, 2004; pp. 696–701.
- de Seabra Rodrigues Dias, I.R.; Mok, S.W.F.; Gordillo-Martinez, F.; Khan, I.; Hsiao, W.W.L.; Law, B.Y.K.; Wong, V.K.W.; Liu, L. The Calcium-Induced Regulation in the Molecular and Transcriptional Circuitry of Human Inflammatory Response and Autoimmunity. Front. Pharmacol. 2017, 8, 962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leboffe, L.; di Masi, A.; Trezza, V.; Polticelli, F.; Ascenzi, P. Human serum albumin: A modulator of cannabinoid drugs. IUBMB Life 2017, 69, 834–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viereckl, M.J.; Krutsinger, K.; Apawu, A.; Gu, J.; Cardona, B.; Barratt, D.; Han, Y. Cannabidiol and Cannabigerol Inhibit Cholangiocarcinoma Growth In Vitro via Divergent Cell Death Pathways. Biomolecules 2022, 12, 854. [Google Scholar] [CrossRef]
- Burgaz, S.; Garcia, C.; Gomez-Canas, M.; Navarrete, C.; Garcia-Martin, A.; Rolland, A.; Del Rio, C.; Casarejos, M.J.; Munoz, E.; Gonzalo-Consuegra, C.; et al. Neuroprotection with the cannabigerol quinone derivative VCE-003.2 and its analogs CBGA-Q and CBGA-Q-Salt in Parkinson′s disease using 6-hydroxydopamine-lesioned mice. Mol. Cell Neurosci. 2021, 110, 103583. [Google Scholar] [CrossRef]
- Borrelli, F.; Fasolino, I.; Romano, B.; Capasso, R.; Maiello, F.; Coppola, D.; Orlando, P.; Battista, G.; Pagano, E.; Di Marzo, V.; et al. Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease. Biochem. Pharmacol. 2013, 85, 1306–1316. [Google Scholar] [CrossRef]
- Fechtner, S.; Singh, A.K.; Ahmed, S. Role of cannabinoid receptor 2 in mediating interleukin-1beta-induced inflammation in rheumatoid arthritis synovial fibroblasts. Clin. Exp. Rheumatol. 2019, 37, 1026–1035. [Google Scholar]
- Wu, Y.Y.; Kumar, R.; Haque, M.S.; Castillejo-Lopez, C.; Alarcon-Riquelme, M.E. BANK1 controls CpG-induced IL-6 secretion via a p38 and MNK1/2/eIF4E translation initiation pathway. J. Immunol. 2013, 191, 6110–6116. [Google Scholar] [CrossRef] [Green Version]
- Kerkmann, M.; Costa, L.T.; Richter, C.; Rothenfusser, S.; Battiany, J.; Hornung, V.; Johnson, J.; Englert, S.; Ketterer, T.; Heckl, W.; et al. Spontaneous formation of nucleic acid-based nanoparticles is responsible for high interferon-alpha induction by CpG-A in plasmacytoid dendritic cells. J. Biol. Chem. 2005, 280, 8086–8093. [Google Scholar] [CrossRef] [Green Version]
- Lowin, T.; Anssar, T.M.; Bauml, M.; Classen, T.; Schneider, M.; Pongratz, G. Positive and negative cooperativity of TNF and Interferon-gamma in regulating synovial fibroblast function and B cell survival in fibroblast/B cell co-cultures. Sci. Rep. 2020, 10, 780. [Google Scholar] [CrossRef] [Green Version]
- Honke, N.; Lowin, T.; Opgenoorth, B.; Shaabani, N.; Lautwein, A.; Teijaro, J.R.; Schneider, M.; Pongratz, G. Endogenously produced catecholamines improve the regulatory function of TLR9-activated B cells. PLoS Biol. 2022, 20, e3001513. [Google Scholar] [CrossRef]
- Moseman, E.A.; Liang, X.; Dawson, A.J.; Panoskaltsis-Mortari, A.; Krieg, A.M.; Liu, Y.J.; Blazar, B.R.; Chen, W. Human plasmacytoid dendritic cells activated by CpG oligodeoxynucleotides induce the generation of CD4+CD25+ regulatory T cells. J. Immunol. 2004, 173, 4433–4442. [Google Scholar] [CrossRef] [Green Version]
- Lafont, V.; Liautard, J.; Gross, A.; Liautard, J.P.; Favero, J. Tumor necrosis factor-alpha production is differently regulated in gamma delta and alpha beta human T lymphocytes. J. Biol. Chem. 2000, 275, 19282–19287. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Wen, S.; Wang, M.; Liang, W.; Li, H.H.; Long, Q.; Guo, H.P.; Liao, Y.H.; Yuan, J. TNF-alpha-secreting B cells contribute to myocardial fibrosis in dilated cardiomyopathy. J. Clin. Immunol. 2013, 33, 1002–1008. [Google Scholar] [CrossRef]
- Harris, D.P.; Haynes, L.; Sayles, P.C.; Duso, D.K.; Eaton, S.M.; Lepak, N.M.; Johnson, L.L.; Swain, S.L.; Lund, F.E. Reciprocal regulation of polarized cytokine production by effector B and T cells. Nat. Immunol. 2000, 1, 475–482. [Google Scholar] [CrossRef]
- Agrawal, S.; Gupta, S. TLR1/2, TLR7, and TLR9 signals directly activate human peripheral blood naive and memory B cell subsets to produce cytokines, chemokines, and hematopoietic growth factors. J. Clin. Immunol. 2011, 31, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Kleiveland, C.R. Peripheral Blood Mononuclear Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., Lopez-Exposito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015; pp. 161–167. [Google Scholar]
- Lowin, T.; Laaser, S.A.; Kok, C.; Bruneau, E.; Pongratz, G. Cannabidiol: Influence on B Cells, Peripheral Blood Mononuclear Cells, and Peripheral Blood Mononuclear Cell/Rheumatoid Arthritis Synovial Fibroblast Cocultures. Cannabis Cannabinoid Res. 2022. [Google Scholar] [CrossRef]
- Cascio, M.G.; Gauson, L.A.; Stevenson, L.A.; Ross, R.A.; Pertwee, R.G. Evidence that the plant cannabinoid cannabigerol is a highly potent alpha2-adrenoceptor agonist and moderately potent 5HT1A receptor antagonist. Br. J. Pharmacol. 2010, 159, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Saroz, Y.; Kho, D.T.; Glass, M.; Graham, E.S.; Grimsey, N.L. Cannabinoid Receptor 2 (CB2) Signals via G-alpha-s and Induces IL-6 and IL-10 Cytokine Secretion in Human Primary Leukocytes. ACS Pharmacol. Transl. Sci. 2019, 2, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Fazzino, F.; Castillo, M.; Mata, S.; Lima, L. Serotonin, 5-HT1A serotonin receptors and proliferation of lymphocytes in major depression patients. Neuroimmunomodulation 2007, 14, 8–15. [Google Scholar] [CrossRef]
- Straub, R.H.; Dufner, B.; Rauch, L. Proinflammatory alpha-Adrenergic Neuronal Regulation of Splenic IFN-gamma, IL-6, and TGF-beta of Mice from Day 15 onwards in Arthritis. Neuroimmunomodulation 2020, 27, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Shoyama, Y.; Sugawa, C.; Tanaka, H.; Morimoto, S. Cannabinoids act as necrosis-inducing factors in Cannabis sativa. Plant Signal Behav. 2008, 3, 1111–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutar, D.A.; Doucette, C.D.; Liwski, R.S.; Hoskin, D.W. Piperine, a Pungent Alkaloid from Black Pepper, Inhibits B Lymphocyte Activation and Effector Functions. Phytother. Res. 2017, 31, 466–474. [Google Scholar] [CrossRef]
- Liu, C.; Reese, R.; Vu, S.; Rouge, L.; Shields, S.D.; Kakiuchi-Kiyota, S.; Chen, H.; Johnson, K.; Shi, Y.P.; Chernov-Rogan, T.; et al. A Non-covalent Ligand Reveals Biased Agonism of the TRPA1 Ion Channel. Neuron 2021, 109, 273–284.e4. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Arnett, F.C.; Edworthy, S.M.; Bloch, D.A.; McShane, D.J.; Fries, J.F.; Cooper, N.S.; Healey, L.A.; Kaplan, S.R.; Liang, M.H.; Luthra, H.S.; et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988, 31, 315–324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Compound/Antibody | Abbreviation (if Applicable) | Order # | Company |
|---|---|---|---|
| A967079 | A96 | 4716 | Bio-Techne/Tocris, Wiesbaden-Nordenstadt, Germany |
| Ruthenium red | RR | 1439 | Bio-Techne/Tocris, Wiesbaden-Nordenstadt, Germany |
| Cannabigerol | CBG | THC pharm, Frankfurt, Germany (discontinued) | |
| Decynium-22 | D22 | 4722 | Bio-Techne/Tocris (discontinued) |
| Ionomycin | Iono | 1704 | Bio-Techne/Tocris, Wiesbaden-Nordenstadt, Germany |
| glycyl-l-phenylalanine 2-naphthylamide | GPN | 14634 | Biomol/Cayman, Hamburg, Germany |
| anti-CD3 | 16-0039-81 | VWR International/Thermo Fisher/Life Sc., Darmstadt, Germany | |
| anti-CD28 | 16-0289-81 | VWR International/Thermo Fisher/Life Sc., Darmstadt, Germany | |
| CpG Oligonucelotide | CpG | tlrl-2006-1 | Invivogen, Toulouse, France |
| Interferon-gamma | IFN-γ | 300-02 | Peprotech/Thermo Fisher Scientific, Hamburg, Germany |
| Tumor necrosis factor | TNF | 300-01A | Peprotech/Thermo Fisher Scientific, Hamburg, Germany |
| Goat Anti-Human IgM | 109-006-129 | Dianova/Jackson ImmunoResearch, Hamburg, Germany | |
| IgG from human serum | I2511 | Sigma-Aldrich, Taufkirchen, Germany | |
| IgM from human serum | I8260 | Sigma-Aldrich, Taufkirchen, Germany | |
| Peroxidase AffiniPure Goat Anti-Human IgG (H+L) | 109-035-003 | Dianova/Jackson ImmunoResearch, Hamburg, Germany | |
| Goat anti-Human IgG, IgM, IgA (H+L) Secondary Antibody | 31128 | Invitrogen/Thermo Fisher Scientific, Schwerte, Germany |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lowin, T.; Tigges-Perez, M.S.; Constant, E.; Pongratz, G. Anti-Inflammatory Effects of Cannabigerol in Rheumatoid Arthritis Synovial Fibroblasts and Peripheral Blood Mononuclear Cell Cultures Are Partly Mediated by TRPA1. Int. J. Mol. Sci. 2023, 24, 855. https://doi.org/10.3390/ijms24010855
Lowin T, Tigges-Perez MS, Constant E, Pongratz G. Anti-Inflammatory Effects of Cannabigerol in Rheumatoid Arthritis Synovial Fibroblasts and Peripheral Blood Mononuclear Cell Cultures Are Partly Mediated by TRPA1. International Journal of Molecular Sciences. 2023; 24(1):855. https://doi.org/10.3390/ijms24010855
Chicago/Turabian StyleLowin, Torsten, Marianne Sofia Tigges-Perez, Eva Constant, and Georg Pongratz. 2023. "Anti-Inflammatory Effects of Cannabigerol in Rheumatoid Arthritis Synovial Fibroblasts and Peripheral Blood Mononuclear Cell Cultures Are Partly Mediated by TRPA1" International Journal of Molecular Sciences 24, no. 1: 855. https://doi.org/10.3390/ijms24010855
APA StyleLowin, T., Tigges-Perez, M. S., Constant, E., & Pongratz, G. (2023). Anti-Inflammatory Effects of Cannabigerol in Rheumatoid Arthritis Synovial Fibroblasts and Peripheral Blood Mononuclear Cell Cultures Are Partly Mediated by TRPA1. International Journal of Molecular Sciences, 24(1), 855. https://doi.org/10.3390/ijms24010855