

Global DNA Methylation and Hydroxymethylation Levels in PBMCs Are Altered in RRMS Patients Treated with IFN-β and GA—A Preliminary Study
 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Data for the Patients
2.2. The Quantitative Variables Correlated with Each Other in a Logical Manner
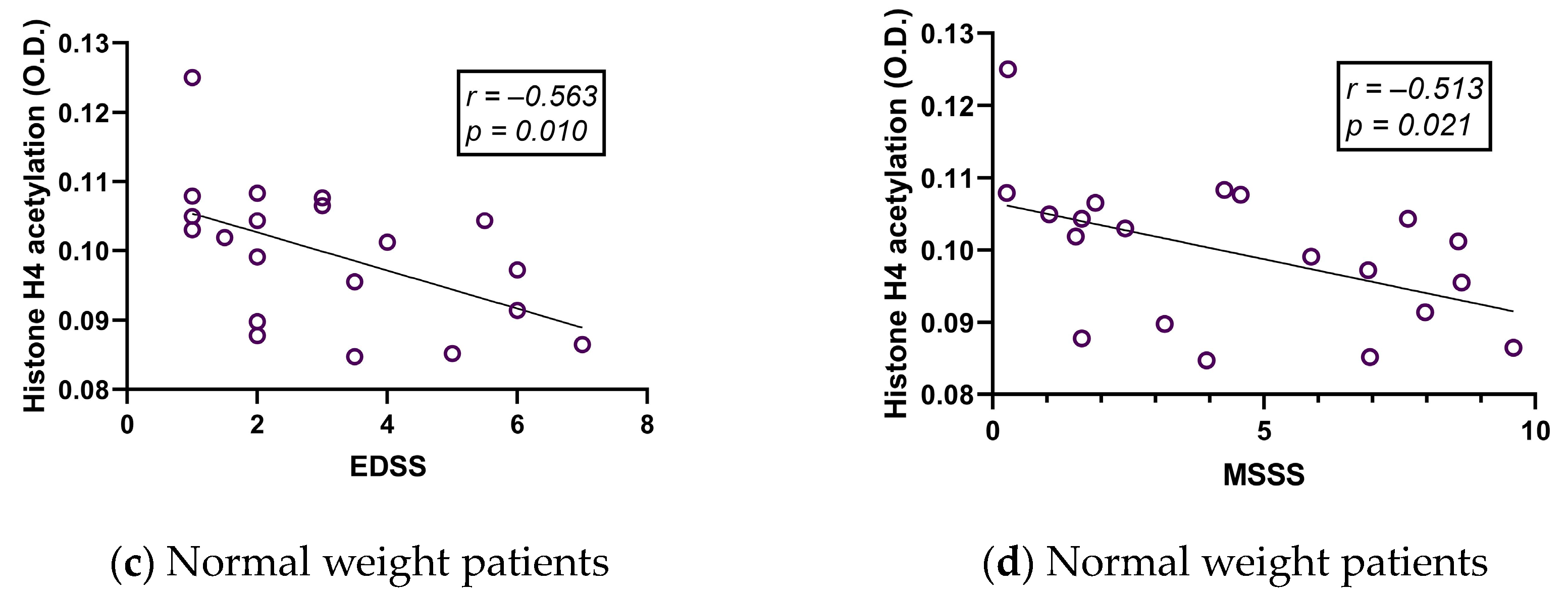
2.3. Histone Acetylation Correlates with Age but Hardly with Clinical Variables in MS
2.4. The Correlation between DNA Hydroxymethylation (5-hmC) and Body Mass Index (BMI) in Healthy Controls Is No Longer Present in RRMS Patients
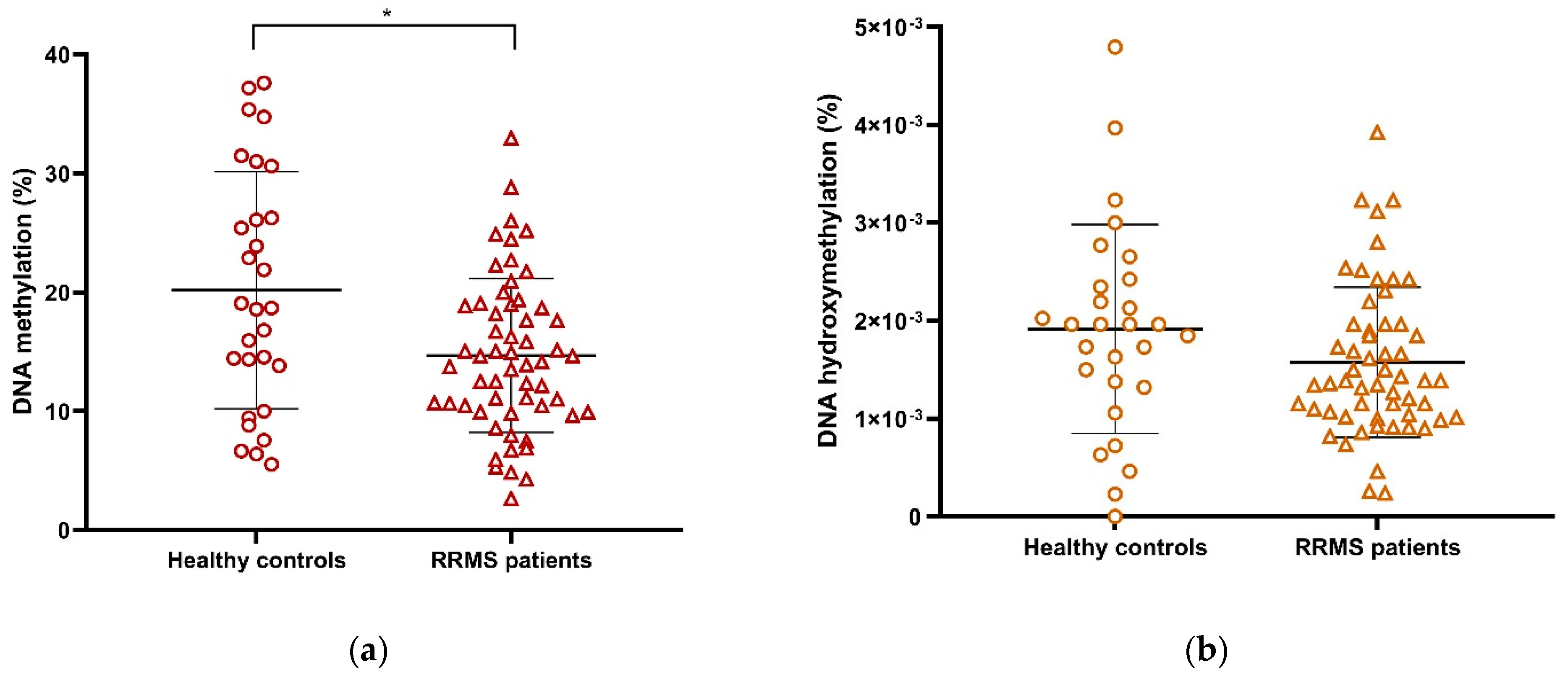
2.5. Global DNA Methylation (5-mC) Is Decreased in RRMS Patients Compared with Healthy Controls
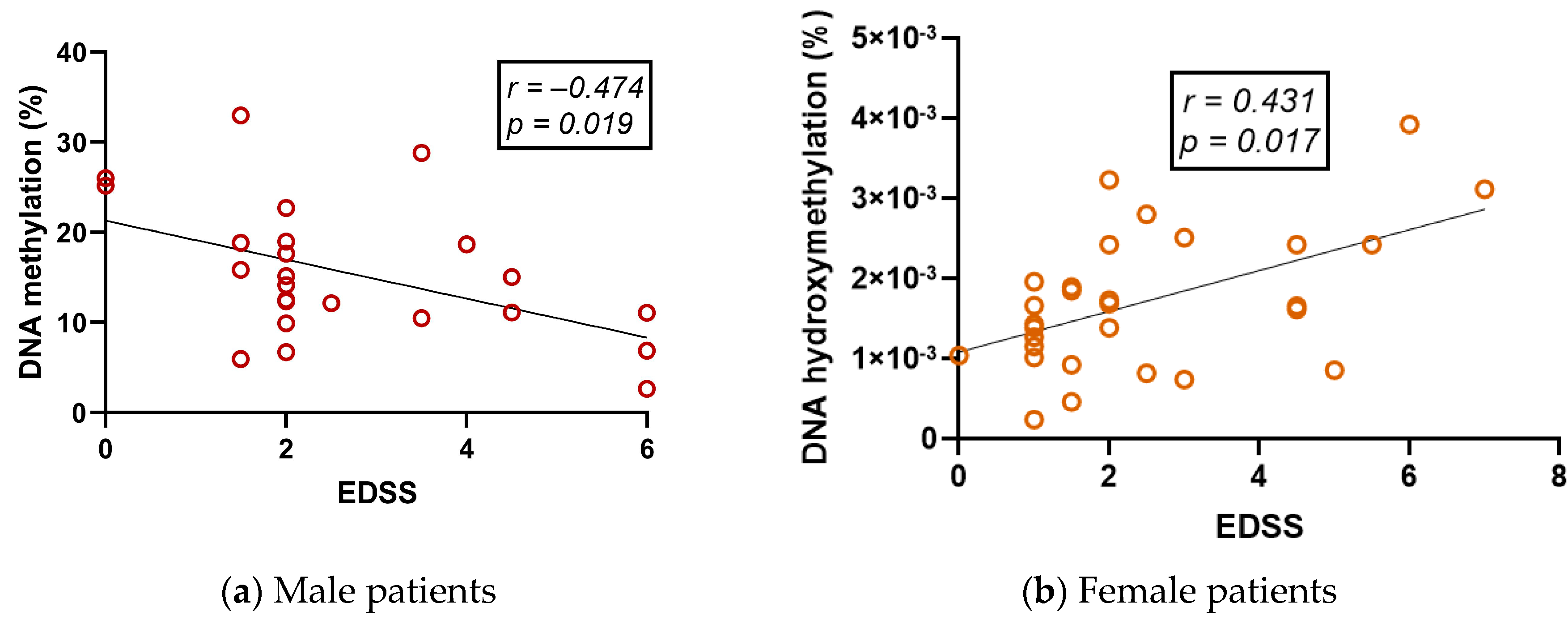
2.6. Global DNA Methylation (5-mC) and DNA Hydroxymethylation (5-hmC) Vary Differently in RRMS Patients and Are Correlated with Clinical Variables
2.7. Global Level of DNA Methylation in Peripheral Blood Mononuclear Cells (PBMCs) Is Decreased in Interferon Beta (IFN-β)- and Glatiramer Acetate (GA)-Treated Patients but Increased in Untreated Patients
2.8. Participant Type (Patient vs. Control) and Age Are Variables Related to Low or High DNA Methylation Levels in Peripheral Blood Mononuclear Cells (PBMCs)
3. Discussion
4. Materials and Methods
4.1. Patients and Healthy Controls
4.2. Blood Sample Collection and Peripheral Blood Mononuclear Cells (PBMC) Isolation
4.3. Quantification of Global DNA Methylation and Hydroxymethylation
4.4. Quantification of Global Histone H3 and H4 Acetylation
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Attfield, K.E.; Jensen, L.T.; Kaufmann, M.; Friese, M.A.; Fugger, L. The immunology of multiple sclerosis. Nat. Rev. Immunol. 2022, 22, 734–750. [Google Scholar] [CrossRef] [PubMed]
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sørensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Duncan, I.D.; Radcliff, A.B. Inherited and acquired disorders of myelin: The underlying myelin pathology. Exp. Neurol. 2016, 283, 452–475. [Google Scholar] [CrossRef] [PubMed]
- Ysrraelit, M.C.; Correale, J. Impact of sex hormones on immune function and multiple sclerosis development. Immunology 2019, 156, 9–22. [Google Scholar] [CrossRef]
- Bertado-Cortés, B.; Venzor-Mendoza, C.; Rubio-Ordoñez, D.; Pérez-Pérez, J.R.; Novelo-Manzano, L.A.; Villamil-Osorio, L.V.; de Jiménez-Ortega, M.J.; de la Villalpando-Gueich, M.L.; Sánchez-Rosales, N.A.; García-Talavera, V. Demographic and clinical characterization of multiple sclerosis in Mexico: The REMEMBer study. Mult. Scler. Relat. Disord. 2020, 46, 102575. [Google Scholar] [CrossRef]
- Nociti, V.; Romozzi, M.; Mirabella, M. Update on Multiple Sclerosis Molecular Biomarkers to Monitor Treatment Effects. J. Pers. Med. 2022, 12, 549. [Google Scholar] [CrossRef]
- Ayrignac, X.; Bilodeau, P.-A.; Prat, A.; Girard, M.; Labauge, P.; Le Lorier, J.; Larochelle, C.; Duquette, P. Assessing the risk of multiple sclerosis disease-modifying therapies. Expert Rev. Neurother. 2019, 19, 695–706. [Google Scholar] [CrossRef]
- Tanasescu, R.; Ionete, C.; Chou, I.-J.; Constantinescu, C.S. Advances in the treatment of relapsing-remitting multiple sclerosis. Biomed. J. 2014, 37, 41–49. [Google Scholar] [CrossRef]
- Polman, C.H.; Bertolotto, A.; Deisenhammer, F.; Giovannoni, G.; Hartung, H.-P.; Hemmer, B.; Killestein, J.; McFarland, H.F.; Oger, J.; Pachner, A.R.; et al. Recommendations for clinical use of data on neutralising antibodies to interferon-beta therapy in multiple sclerosis. Lancet Neurol. 2010, 9, 740–750. [Google Scholar] [CrossRef]
- Fernández, O.; Cuevas, C.; Núñez-Orozco, L.; Carrasco-Vargas, H.; Ordoñez-Boschetti, L.; De La Maza, J.; Sauri-Suárez, S.; Arrambide, G.; Padilla, O.; Blaisdel, C.; et al. Guía de práctica clínica. Esclerosis múltiple: Situación actual y tendencias futuras. Rev. Mex. Neurocienc. 2016, 17, 3–22. [Google Scholar]
- Ziemssen, T.; Akgün, K.; Brück, W. Molecular biomarkers in multiple sclerosis. J. Neuroinflam. 2019, 16, 272. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Hu, Z.; Xiao, H.; Zhou, F.; Yang, B. The tale of histone modifications and its role in multiple sclerosis. Hum. Genom. 2018, 12, 31. [Google Scholar] [CrossRef]
- Webb, L.M.; Guerau-de-Arellano, M. Emerging Role for Methylation in Multiple Sclerosis: Beyond DNA. Trends Mol. Med. 2017, 23, 546–562. [Google Scholar] [CrossRef] [PubMed]
- Kular, L.; Jagodic, M. Epigenetic insights into multiple sclerosis disease progression. J. Intern. Med. 2020, 288, 82–102. [Google Scholar] [CrossRef]
- Huynh, J.L.; Casaccia, P. Epigenetic mechanisms in multiple sclerosis: Implications for pathogenesis and treatment. Lancet Neurol. 2013, 12, 195–206. [Google Scholar] [CrossRef]
- Aslani, S.; Jafari, N.; Javan, M.R.; Karami, J.; Ahmadi, M.; Jafarnejad, M. Epigenetic Modifications and Therapy in Multiple Sclerosis. Neuromol. Med. 2017, 19, 11–23. [Google Scholar] [CrossRef]
- Gilli, F.; DiSano, K.D.; Pachner, A.R. SeXX Matters in Multiple Sclerosis. Front. Neurol. 2020, 11, 616. [Google Scholar] [CrossRef]
- Harbo, H.F.; Gold, R.; Tintoré, M. Sex and gender issues in multiple sclerosis. Ther. Adv. Neurol. Disord. 2013, 6, 237–248. [Google Scholar] [CrossRef]
- Küçükali, C.; Kürtüncü, M.; Çoban, A.; Çebi, M.; Tüzün, E. Epigenetics of multiple sclerosis: An updated review. Neuromol. Med. 2015, 17, 83–96. [Google Scholar] [CrossRef]
- Castro, K.; Ntranos, A.; Amatruda, M.; Petracca, M.; Kosa, P.; Chen, E.Y.; Morstein, J.; Trauner, D.; Watson, C.T.; Kiebish, M.A.; et al. Body Mass Index in Multiple Sclerosis modulates ceramide-induced DNA methylation and disease course. EBioMedicine 2019, 43, 392–410. [Google Scholar] [CrossRef] [PubMed]
- Field, J.; Fox, A.; Jordan, M.A.; Baxter, A.G.; Spelman, T.; Gresle, M.; Butzkueven, H.; Kilpatrick, T.J.; Rubio, J.P. Interleukin-2 receptor-α proximal promoter hypomethylation is associated with multiple sclerosis. Genes Immun. 2017, 18, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Janson, P.C.J.; Linton, L.B.; Bergman, E.A.; Marits, P.; Eberhardson, M.; Piehl, F.; Malmström, V.; Winqvist, O. Profiling of CD4+ T Cells with Epigenetic Immune Lineage Analysis. J. Immunol. 2011, 186, 92–102. [Google Scholar] [CrossRef]
- Kumagai, C.; Kalman, B.; Middleton, F.A.; Vyshkina, T.; Massa, P.T. Increased promoter methylation of the immune regulatory gene SHP-1 in leukocytes of multiple sclerosis subjects. J. Neuroimmunol. 2012, 246, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Liggett, T.; Melnikov, A.; Tilwalli, S.; Yi, Q.; Chen, H.; Replogle, C.; Feng, X.; Reder, A.; Stefoski, D.; Balabanov, R.; et al. Methylation patterns of cell-free plasma DNA in relapsing-remitting multiple sclerosis. J. Neurol. Sci. 2010, 290, 16–21. [Google Scholar] [CrossRef]
- Sokratous, M.; Dardiotis, E.; Bellou, E.; Tsouris, Z.; Michalopoulou, A.; Dardioti, M.; Siokas, V.; Rikos, D.; Tsatsakis, A.; Kovatsi, L.; et al. CpG Island Methylation Patterns in Relapsing-Remitting Multiple Sclerosis. J. Mol. Neurosci. 2018, 64, 478–484. [Google Scholar] [CrossRef]
- Bingen, J.M.; Clark, L.V.; Band, M.R.; Munzir, I.; Carrithers, M.D. Differential DNA methylation associated with multiple sclerosis and disease modifying treatments in an underrepresented minority population. Front. Genet. 2022, 13, 1058817. [Google Scholar] [CrossRef]
- Calabrese, R.; Valentini, E.; Ciccarone, F.; Guastafierro, T.; Bacalini, M.G.; Ricigliano, V.A.G.; Zampieri, M.; Annibali, V.; Mechelli, R.; Franceschi, C.; et al. TET2 gene expression and 5-hydroxymethylcytosine level in multiple sclerosis peripheral blood cells. Biochim. Biophys. Acta 2014, 1842, 1130–1136. [Google Scholar] [CrossRef]
- Gökdoğan-Edgünlü, T.; Ünal, Y.; Karakaş Çelik, S.; Genç, Ö.; Emre, U.; Kutlu, G. The effect of FOXO gene family variants and global DNA metylation on RRMS disease. Gene 2020, 726, 144172. [Google Scholar] [CrossRef]
- Pinto-Medel, M.J.; Oliver-Martos, B.; Urbaneja-Romero, P.; Hurtado-Guerrero, I.; Ortega-Pinazo, J.; Serrano-Castro, P.; Fernández, Ó.; Leyva, L. Global methylation correlates with clinical status in multiple sclerosis patients in the first year of IFNbeta treatment. Sci. Rep. 2017, 7, 8727. [Google Scholar] [CrossRef]
- Brorson, I.S.; Eriksson, A.M.; Høgestøl, E.; Leikfoss, I.S.; Harbo, H.F.; Berge, T.; Vitelli, V.; Bos, S.D. Global DNA methylation changes in treated and untreated MS patients measured over time. J. Neuroimmunol. 2022, 364, 577808. [Google Scholar] [CrossRef] [PubMed]
- Diniz, S.N.; da Silva, C.F.; de Almeida, I.T.; da Silva Costa, F.E.; de Oliveira, E.M.L. INFβ treatment affects global DNA methylation in monocytes of patients with multiple sclerosis. J. Neuroimmunol. 2021, 355, 577563. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Sun, X.; Shu, Y.; Wang, Y.; Xiao, L.; Wang, Z.; Hu, X.; Kermode, A.G.; Qiu, W. Serum CCL20 and its association with SIRT1 activity in multiple sclerosis patients. J. Neuroimmunol. 2017, 313, 56–60. [Google Scholar] [CrossRef]
- Qadir, M.I.; Anwer, F. Epigenetic Modification Related to Acetylation of Histone and Methylation of DNA as a Key Player in Immunological Disorders. Crit. Rev. Eukaryot. Gene Expr. 2019, 29, 1–15. [Google Scholar] [CrossRef]
- Suzuki, K.; Luo, Y. Histone Acetylation and the Regulation of Major Histocompatibility Class II Gene Expression. Adv Protein Chem. Struct. Biol. 2017, 106, 71–111. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Li, B.; Zhou, Z.; Hancock, W.W.; Zhang, H.; Greene, M.I. Histone acetyltransferase mediated regulation of FOXP3 acetylation and Treg function. Curr. Opin. Immunol. 2010, 22, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.W.Y.; Chang, C.-B.; Tung, C.-H.; Sun, J.; Suen, J.-L.; Wu, S.-F. Low-dose 5-aza-2’-deoxycytidine pretreatment inhibits experimental autoimmune encephalomyelitis by induction of regulatory T cells. Mol. Med. 2014, 20, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Mangano, K.; Fagone, P.; Bendtzen, K.; Meroni, P.L.; Quattrocchi, C.; Mammana, S.; Di Rosa, M.; Malaguarnera, L.; Coco, M.; Magro, G.; et al. Hypomethylating Agent 5-Aza-2′-deoxycytidine (DAC) Ameliorates Multiple Sclerosis in Mouse Models. J. Cell. Physiol. 2014, 229, 1918–1925. [Google Scholar] [CrossRef]
- Reyes-Mata, P.M.; Rojas-Mayorquín, A.E.; Carrera-Quintanar, L.; González-Castillo, C.; Mireles-Ramírez, M.A.; de Guerrero-García, J.J.; Ortuño-Sahagún, D. Pleiotrophin Serum Level is Increased in Relapsing-Remitting Multiple Sclerosis and Correlates With Sex, BMI and Treatment. Arch. Med. Res. 2022, 53, 59–68. [Google Scholar] [CrossRef]
- Guerrero-García, J.J.; Castañeda-Moreno, V.A.; Torres-Carrillo, N.; Muñoz-Valle, J.F.; Bitzer-Quintero, O.K.; Ponce-Regalado, M.D.; Mireles-Ramírez, M.A.; Valle, Y.; Ortuño-Sahagún, D. Interleukin-17A Levels Vary in Relapsing-Remitting Multiple Sclerosis Patients in Association with Their Age, Treatment and the Time of Evolution of the Disease. Neuroimmunomodulation 2016, 23, 8–17. [Google Scholar] [CrossRef]
- Guerrero-García, J.J.; Rojas-Mayorquín, A.E.; Valle, Y.; Padilla-Gutiérrez, J.R.; Castañeda-Moreno, V.A.; Mireles-Ramírez, M.A.; Muñoz-Valle, J.F.; Ortuño-Sahagún, D. Decreased serum levels of sCD40L and IL-31 correlate in treated patients with Relapsing-Remitting Multiple Sclerosis. Immunobiology 2018, 223, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Huynh, J.L.; Garg, P.; Thin, T.H.; Yoo, S.; Dutta, R.; Trapp, B.D.; Haroutunian, V.; Zhu, J.; Donovan, M.J.; Sharp, A.J.; et al. Epigenome-wide differences in pathology-free regions of multiple sclerosis–affected brains. Nat. Neurosci. 2014, 17, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Sospedra, M.; Martin, R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005, 23, 683–747. [Google Scholar] [CrossRef] [PubMed]
- Zheleznyakova, G.Y.; Piket, E.; Marabita, F.; Pahlevan Kakhki, M.; Ewing, E.; Ruhrmann, S.; Needhamsen, M.; Jagodic, M.; Kular, L. Epigenetic research in multiple sclerosis: Progress, challenges, and opportunities. Physiol. Genom. 2017, 49, 447–461. [Google Scholar] [CrossRef]
- Jenkins, T.G.; Aston, K.I.; Cairns, B.R.; Carrell, D.T. Paternal aging and associated intraindividual alterations of global sperm 5-methylcytosine and 5-hydroxymethylcytosine levels. Fertil. Steril. 2013, 100, 945–951. [Google Scholar] [CrossRef]
- Tellez-Plaza, M.; Tang, W.; Shang, Y.; Umans, J.G.; Francesconi, K.A.; Goessler, W.; Ledesma, M.; Leon, M.; Laclaustra, M.; Pollak, J.; et al. Association of Global DNA Methylation and Global DNA Hydroxymethylation with Metals and Other Exposures in Human Blood DNA Samples. Environ. Health Perspect. 2014, 122, 946–954. [Google Scholar] [CrossRef]
- Caliri, A.W.; Caceres, A.; Tommasi, S.; Besaratinia, A. Hypomethylation of LINE-1 repeat elements and global loss of DNA hydroxymethylation in vapers and smokers. Epigenetics 2020, 15, 816–829. [Google Scholar] [CrossRef]
- Li, H.; Lu, T.; Sun, W.; Ma, R.; Zhong, H.; Wei, Y.; Chen, D.; Wen, Y.; Carlsten, C.; Wen, W. Ten-Eleven Translocation (TET) Enzymes Modulate the Activation of Dendritic Cells in Allergic Rhinitis. Front. Immunol. 2019, 10, 2271. [Google Scholar] [CrossRef]
- Gautam, P.; Sharma, A.; Bhatnagar, A. Global histone modification analysis reveals hypoacetylated H3 and H4 histones in B Cells from systemic lupus erythematosus patients. Immunol. Lett. 2021, 240, 41–45. [Google Scholar] [CrossRef]
- Lv, X.; Zhou, M.; Zhang, Q.; He, Y.; Wang, Y.; Xuan, J.; Shi, G.; Li, Y. Abnormal Histones Acetylation in Patients with Primary Sjögren’s Syndrome. Clin. Rheumatol. 2022, 41, 1465–1472. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, Q.; Liu, Y.; Xiong, Y.; Wu, S.; Yang, J.; Zhou, H.; Liu, X.; Wei, X.; Song, Y. Aberrant histone modification in CD19+ B cells of patients with chronic lymphocytic leukemia. OncoTargets Ther. 2017, 10, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Jamebozorgi, K.; Rostami, D.; Pormasoumi, H.; Taghizadeh, E.; Barreto, G.E.; Sahebkar, A. Epigenetic aspects of multiple sclerosis and future therapeutic options. Int. J. Neurosci. 2021, 131, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.-J.; Kim, K. New Insights into the Role of Histone Changes in Aging. Int. J. Mol. Sci. 2020, 21, 8241. [Google Scholar] [CrossRef] [PubMed]
- Pedre, X.; Mastronardi, F.; Bruck, W.; López-Rodas, G.; Kuhlmann, T.; Casaccia, P. Changed histone acetylation patterns in normal appearing white matter and early MS lesions. J. Neurosci. 2011, 31, 3435–3445. [Google Scholar] [CrossRef]
- Castro, K.; Casaccia, P. Epigenetic modifications in brain and immune cells of multiple sclerosis patients. Mult. Scler. 2018, 24, 69–74. [Google Scholar] [CrossRef]
- Rohde, K.; Keller, M.; Stumvoll, M.; Dietrich, A.; Blüher, M.; Böttcher, Y. DNA 5-hydroxymethylation in human adipose tissue differs between subcutaneous and visceral adipose tissue depots. Epigenomics 2015, 7, 911–920. [Google Scholar] [CrossRef]
- Mitsuya, K.; Parker, A.N.; Liu, L.; Ruan, J.; Vissers, M.C.M.; Myatt, L. Alterations in the placental methylome with maternal obesity and evidence for metabolic regulation. PLoS ONE 2017, 12, e0186115. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Nonino, C.B.; de Oliveira, B.A.P.; de Pinhel, M.A.S.; Mansego, M.L.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. DNA Methylation and Hydroxymethylation Levels in Relation to Two Weight Loss Strategies: Energy-Restricted Diet or Bariatric Surgery. Obes. Surg. 2016, 26, 603–611. [Google Scholar] [CrossRef]
- Guerrero-García, J.J.; Carrera-Quintanar, L.; López-Roa, R.I.; Márquez-Aguirre, A.L.; Rojas-Mayorquín, A.E.; Ortuño-Sahagún, D. Multiple Sclerosis and Obesity: Possible Roles of Adipokines. Mediat. Inflamm. 2016, 2016, 4036232. [Google Scholar] [CrossRef]
- Tang, Y.; Luo, M.; Pan, K.; Ahmad, T.; Zhou, T.; Miao, Z.; Zhou, H.; Sun, H.; Xu, X.; Namaka, M.; et al. DNA hydroxymethylation changes in response to spinal cord damage in a multiple sclerosis mouse model. Epigenomics 2019, 11, 323–335. [Google Scholar] [CrossRef]
- Kular, L.; Needhamsen, M.; Adzemovic, M.Z.; Kramarova, T.; Gomez-Cabrero, D.; Ewing, E.; Piket, E.; Tegnér, J.; Beck, S.; Piehl, F.; et al. Neuronal methylome reveals CREB-associated neuro-axonal impairment in multiple sclerosis. Clin. Epigenet. 2019, 11, 86. [Google Scholar] [CrossRef] [PubMed]
- Hedström, A.K.; Olsson, T.; Alfredsson, L. Body mass index during adolescence, rather than childhood, is critical in determining MS risk. Mult. Scler. 2016, 22, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Marrodan, M.; Farez, M.F.; Balbuena Aguirre, M.E.; Correale, J. Obesity and the risk of Multiple Sclerosis. The role of Leptin. Ann. Clin. Transl. Neurol. 2021, 8, 406–424. [Google Scholar] [CrossRef] [PubMed]
- Bassi, M.; Iezzi, E.; Buttari, F.; Gilio, L.; Simonelli, I.; Carbone, F.; Micillo, T.; De Rosa, V.; Sica, F.; Furlan, R.; et al. Obesity worsens central inflammation and disability in multiple sclerosis. Mult. Scler. 2019, 26, 1237–1246. [Google Scholar] [CrossRef]
- Mowry, E.M.; Azevedo, C.J.; McCulloch, C.E.; Okuda, D.T.; Lincoln, R.R.; Waubant, E.; Hauser, S.L.; Pelletier, D. Body mass index, but not vitamin D status, is associated with brain volume change in MS. Neurology 2018, 91, e2256–e2264. [Google Scholar] [CrossRef]
- Wesnes, K.; Myhr, K.-M.; Riise, T.; Kvistad, S.S.; Torkildsen, Ø.; Wergeland, S.; Holmøy, T.; Midgard, R.; Bru, A.; Edland, A.; et al. Low vitamin D, but not tobacco use or high BMI, is associated with long-term disability progression in multiple sclerosis. Mult. Scler. Relat. Disord. 2021, 50, 102801. [Google Scholar] [CrossRef]
- Govender, P.; Ghai, M.; Okpeku, M. Sex-specific DNA methylation: Impact on human health and development. Mol. Genet. Genom. 2022, 297, 1451–1466. [Google Scholar] [CrossRef]
- Avila, S.; de Guerrero-García, J.J.; Becerril-Villanueva, E.; Pérez-Sánchez, G.; Pavón, L.; Rojas-Mayorquín, A.E.; Mireles-Ramírez, M.A.; Muñoz-Valle, J.F.; Vallejo-Castillo, L.; Medina-Rivero, E.; et al. A differential sex-specific pattern of IgG2 and IgG4 subclasses of anti-drug antibodies (ADAs) induced by glatiramer acetate in relapsing-remitting multiple sclerosis patients. Mult. Scler. Relat. Disord. 2019, 34, 92–99. [Google Scholar] [CrossRef]
- Neven, K.Y.; Piola, M.; Angelici, L.; Cortini, F.; Fenoglio, C.; Galimberti, D.; Pesatori, A.C.; Scarpini, E.; Bollati, V. Repetitive element hypermethylation in multiple sclerosis patients. BMC Genet. 2016, 17, 84. [Google Scholar] [CrossRef]
- Lenin, P.; Jiménez-Martínez, M.; Garcés-Alvarez, M.E. Inmunología Molecular, Celular y Traslacional, 2nd ed.; Wolters Kluwer: Barcelona, España, 2021; ISBN 978-84-17949-18-1. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Inmunología Celular y Molecular, 8th ed.; Elsevier: Philadelphia, PA, USA, 2015; ISBN 978-0-323-31614-9. [Google Scholar]
- Jones, A.P.; Kermode, A.G.; Lucas, R.M.; Carroll, W.M.; Nolan, D.; Hart, P.H. Circulating immune cells in multiple sclerosis. Clin. Exp. Immunol. 2017, 187, 193–203. [Google Scholar] [CrossRef]
- Hegen, H.; Adrianto, I.; Lessard, C.J.; Millonig, A.; Bertolotto, A.; Comabella, M.; Giovannoni, G.; Guger, M.; Hoelzl, M.; Khalil, M.; et al. Cytokine profiles show heterogeneity of interferon-β response in multiple sclerosis patients. Neurol. Neuroimmunol. Neuroinflamm. 2016, 3, e202. [Google Scholar] [CrossRef] [PubMed]
- Bărcuţean, L.I.; Romaniuc, A.; Maier, S.; Bajko, Z.; Moţăţăianu, A.; Adina, H.; Simu, I.; Andone, S.; Bălaşa, R. Clinical and Serological Biomarkers of Treatment’s Response in Multiple Sclerosis Patients Treated Continuously with Interferonβ-1b for More than a Decade. CNS Neurol. Disord. Drug Targets 2018, 17, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Celarain, N.; Tomas-Roig, J. Changes in Deoxyribonucleic Acid Methylation Contribute to the Pathophysiology of Multiple Sclerosis. Front. Genet. 2019, 10, 1138. [Google Scholar] [CrossRef] [PubMed]
- Dunaeva, M.; Derksen, M.; Pruijn, G.J.M. LINE-1 Hypermethylation in Serum Cell-Free DNA of Relapsing Remitting Multiple Sclerosis Patients. Mol. Neurobiol. 2018, 55, 4681–4688. [Google Scholar] [CrossRef]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444. [Google Scholar] [CrossRef]
- Roxburgh, R.H.S.R.; Seaman, S.R.; Masterman, T.; Hensiek, A.E.; Sawcer, S.J.; Vukusic, S.; Achiti, I.; Confavreux, C.; Coustans, M.; le Page, E.; et al. Multiple Sclerosis Severity Score: Using disability and disease duration to rate disease severity. Neurology 2005, 64, 1144–1151. [Google Scholar] [CrossRef]
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. National Multiple Sclerosis Society (USA) Advisory Committee on Clinical Trials of New Agents in Multiple Sclerosis. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef]
- Li, S.; Tollefsbol, T.O. DNA methylation methods: Global DNA methylation and methylomic analyses. Methods 2021, 187, 28–43. [Google Scholar] [CrossRef]
- Olova, N.N. ELISA-Based Quantitation of Global 5hmC Levels. Methods Mol. Biol. 2021, 2272, 45–63. [Google Scholar] [CrossRef]
- Grubbs, F.E.; Beck, G. Extension of Sample Sizes and Percentage Points for Significance Tests of Outlying Observations. Technometrics 1972, 14, 847–854. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Female | Male | Total | p |
|---|---|---|---|---|
| 33 | 24 | 57 | - | |
| (%) | 60 | 40 | 100 | - |
| Age (years) | 40.9 ± 9.7 | 39.4 ± 9.3 | 40.3 ± 9.5 | 0.567 a |
| Disease duration (years) | 10.7 ± 6.3 | 10.0 ± 7.3 | 10.4 ± 6.6 | 0.389 b |
| EDSS (n = 54) | 2.4 ± 1.8 | 2.7 ± 1.7 | 2.5 ± 1.7 | 0.511 b |
| MSSS (n = 54) | 3.0 ± 2.9 | 3.7 ± 2.4 | 3.3 ± 2.7 | 0.282 b |
| BMI (n = 53) | 25.3 ± 4.6 | 27.2 ± 6.9 | 26.2 ± 5.7 | 0.107 b |
| Variable | Subgroups | n | M ± SD | p |
|---|---|---|---|---|
| AcH3 (O.D.) | RRMS patients | 57 | 0.113 ± 0.04 | 0.017 * |
| Healthy controls | 29 | 0.118 ± 0.04 | ||
| AcH3 (O.D.) | IFN-β patients | 25 | 0.107 ± 0.02 | 0.618 a |
| GA patients | 26 | 0.116 ± 0.04 | ||
| Non-treated patients | 4 | 0.102 ± 0.01 | ||
| AcH4 (O.D.) | RRMS patients | 57 | 0.101 ± 0.02 | 0.377 |
| Healthy controls | 30 | 0.098 ± 0.01 | ||
| AcH4 (O.D.) | IFN-β patients | 25 | 0.01 ± 0.02 | 0.418 a |
| GA patients | 26 | 0.103 ± 0.02 | ||
| Non-treated patients | 6 | 0.102 ± 0.01 |
| Variable | Subgroup | n | M ± SD | p |
|---|---|---|---|---|
| %5-mC | RRMS patients | 57 | 14.69 ± 6.47 | 0.019 * |
| Healthy controls | 29 | 20.19 ± 9.97 | ||
| %5-hmC | RRMS patients | 56 | 0.0016 ± 0.0008 | 0.075 |
| Healthy controls | 28 | 0.0018 ± 0.0011 |
| Variable | Subgroups | n | M ± SD | p |
|---|---|---|---|---|
| %5-mC | IFN-β patients | 26 | 13.63 ± 6.16 † | 0.004 ** |
| GA patients | 26 | 13.81 ± 5.67 ‡ | ||
| Non-treated patients | 5 | 24.75 ± 2.89 †‡ | ||
| %5-hmC | IFN-β patients | 25 | 0.0015 ± 0.0008 | 0.418 |
| GA patients | 26 | 0.0016 ± 0.0008 | ||
| Non-treated patients | 6 | 0.0016 ± 0.0006 |
| Dependent Variable | Independent Variable | OR | IC 95% | p |
|---|---|---|---|---|
| %5-mC (high vs. low) | Control vs. RRMS Patient | 5.189 | 1.608–16.743 | 0.006 * |
| Age | 0.899 | 0.838–0.964 | 0.003 * | |
| Gender | 1.682 | 0.513–5.513 | 0.390 | |
| BMI | 1.030 | 0.923–1.149 | 0.597 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-Mata, M.P.; Mireles-Ramírez, M.A.; Griñán-Ferré, C.; Pallàs, M.; Pavón, L.; Guerrero-García, J.d.J.; Ortuño-Sahagún, D. Global DNA Methylation and Hydroxymethylation Levels in PBMCs Are Altered in RRMS Patients Treated with IFN-β and GA—A Preliminary Study. Int. J. Mol. Sci. 2023, 24, 9074. https://doi.org/10.3390/ijms24109074
Reyes-Mata MP, Mireles-Ramírez MA, Griñán-Ferré C, Pallàs M, Pavón L, Guerrero-García JdJ, Ortuño-Sahagún D. Global DNA Methylation and Hydroxymethylation Levels in PBMCs Are Altered in RRMS Patients Treated with IFN-β and GA—A Preliminary Study. International Journal of Molecular Sciences. 2023; 24(10):9074. https://doi.org/10.3390/ijms24109074
Chicago/Turabian StyleReyes-Mata, María Paulina, Mario Alberto Mireles-Ramírez, Christian Griñán-Ferré, Mercè Pallàs, Lenin Pavón, José de Jesús Guerrero-García, and Daniel Ortuño-Sahagún. 2023. "Global DNA Methylation and Hydroxymethylation Levels in PBMCs Are Altered in RRMS Patients Treated with IFN-β and GA—A Preliminary Study" International Journal of Molecular Sciences 24, no. 10: 9074. https://doi.org/10.3390/ijms24109074
APA StyleReyes-Mata, M. P., Mireles-Ramírez, M. A., Griñán-Ferré, C., Pallàs, M., Pavón, L., Guerrero-García, J. d. J., & Ortuño-Sahagún, D. (2023). Global DNA Methylation and Hydroxymethylation Levels in PBMCs Are Altered in RRMS Patients Treated with IFN-β and GA—A Preliminary Study. International Journal of Molecular Sciences, 24(10), 9074. https://doi.org/10.3390/ijms24109074