
“ThermoTRP” Channel Expression in Cancers: Implications for Diagnosis and Prognosis (Practical Approach by a Pathologist)

Abstract
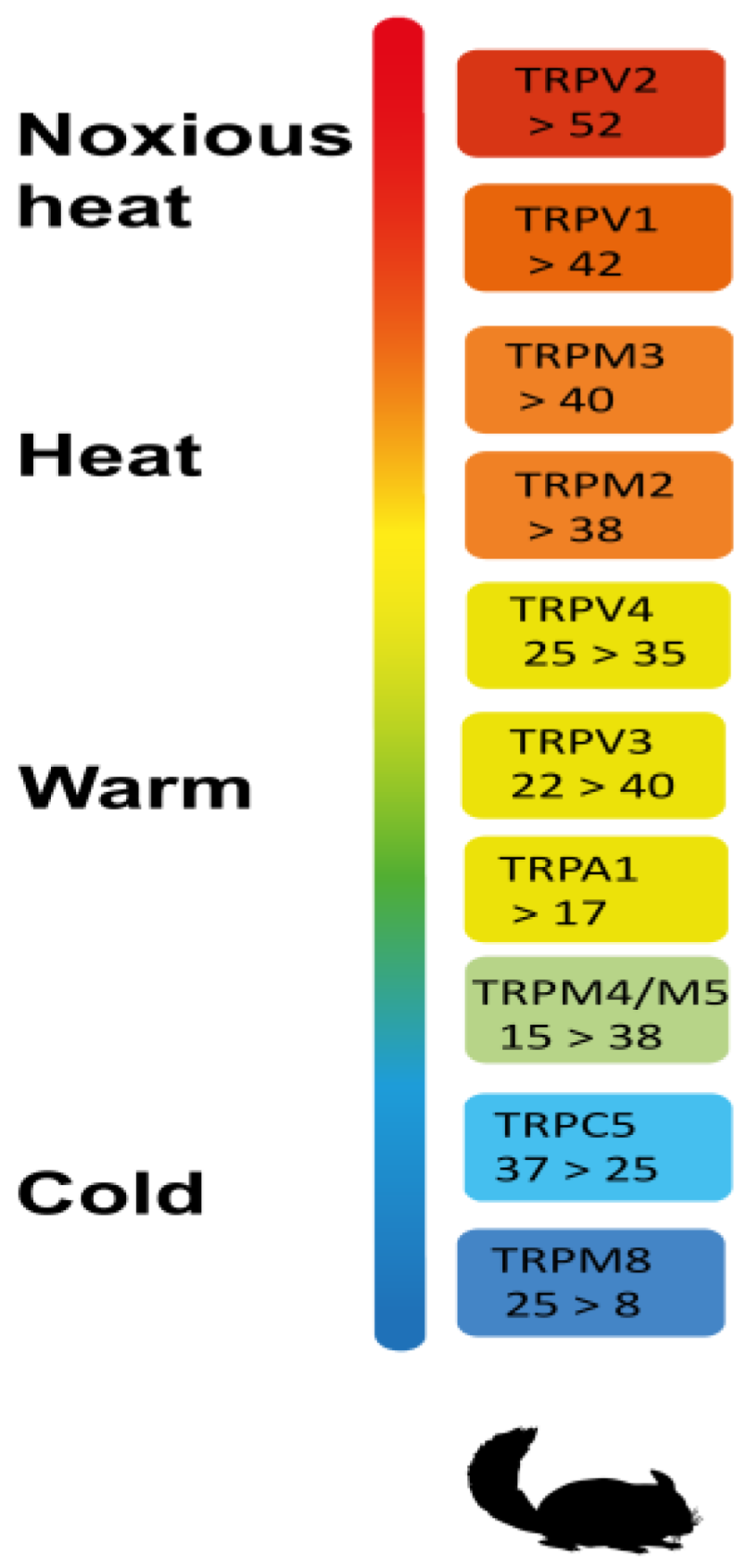
:1. TRPs and “ThermoTRPs”: A Brief Introduction to Terminology and Biology
2. The Expression Landscape and Function of “ThermoTRPs” in Normal Tissues
2.1. TRPA1
2.2. TRPC5
2.3. TRPM2
2.4. TRPM3
2.5. TRPM4
2.6. TRPM5
2.7. TRPM8
2.8. TRPV1
2.9. TRPV2
2.10. TRPV3
2.11. TRPV4
3. Aberrant “ThermoTRP” Expression in Cancers: Implications for Diagnosis and Prognostication
- a.
- Squamous cell carcinoma of the skin and the head-and-neck
- b.
- Pulmonary small cell carcinoma and adenocarcinoma
- c.
- Prostate cancer
- d.
- Bladder cancer
- e.
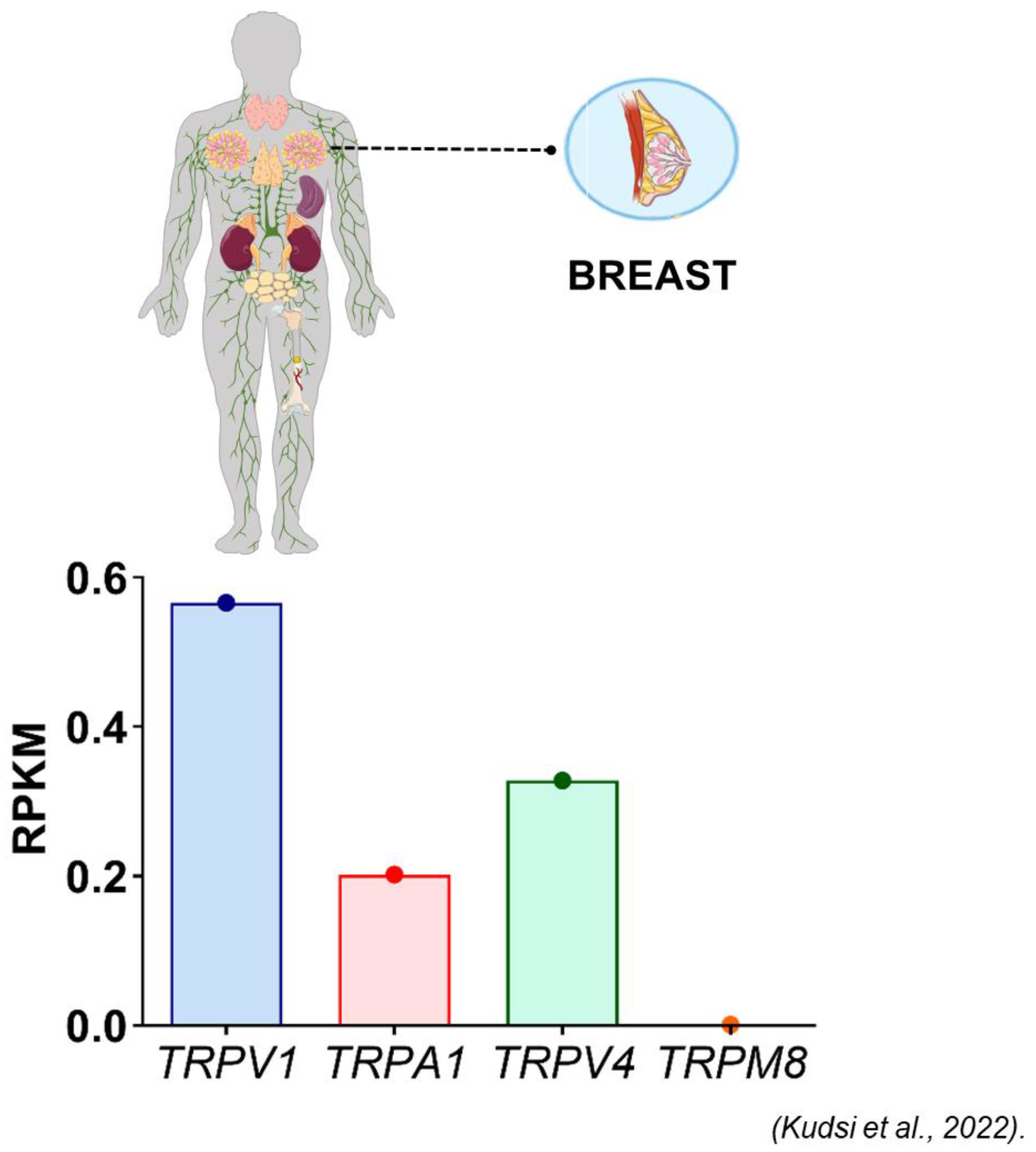
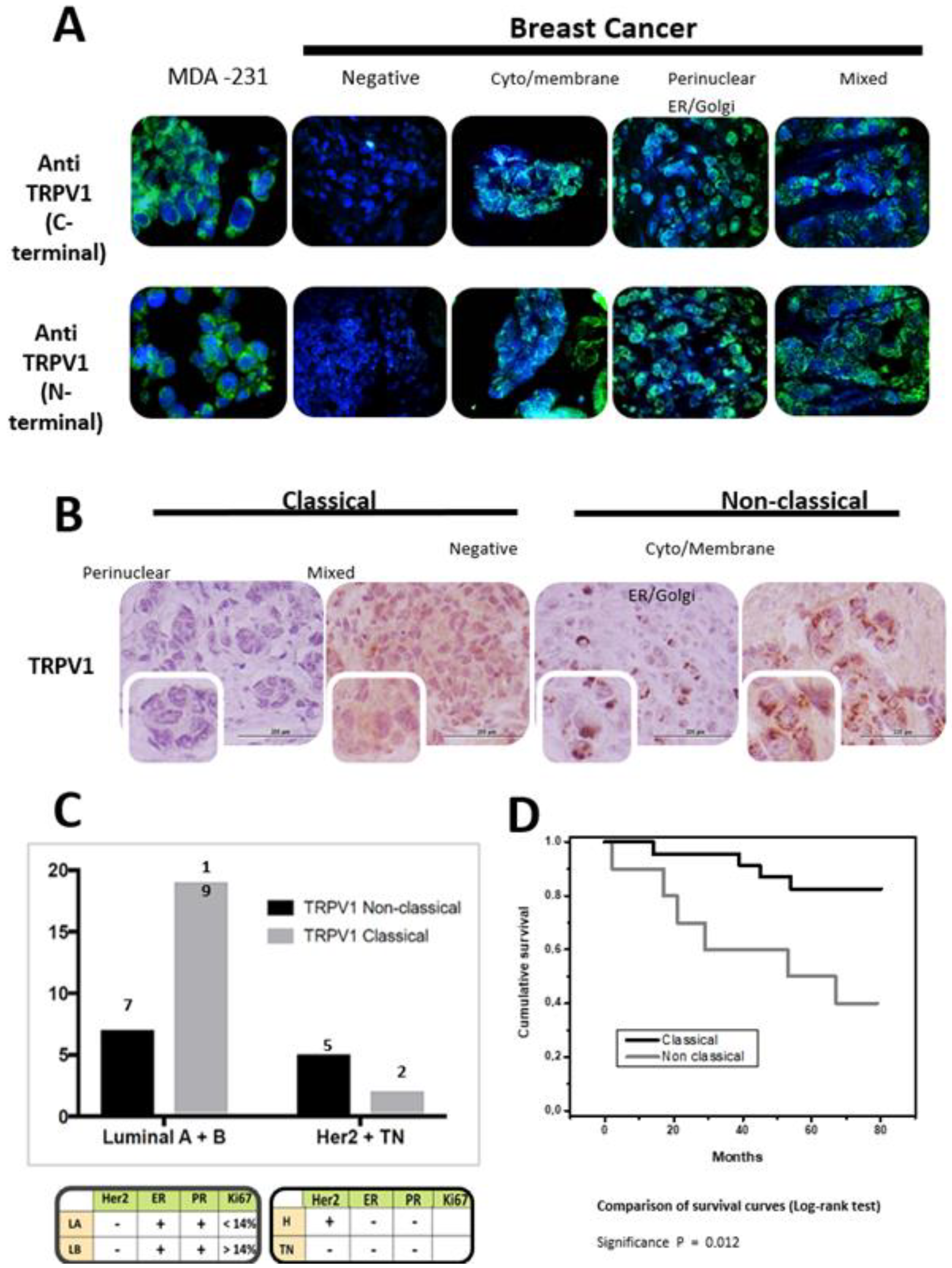
- Breast cancer
- f.
- Gastric adenocarcinoma
- g.
- Colorectal adenocarcinoma
- h.
- Pancreatic ductal adenocarcinoma
- i.
- Endometrial and ovarian carcinoma
- j.
- Renal cell carcinoma
- k.
- Hepatocellular carcinoma
- l.
- Melanoma
- m.
- Gliomas, including glioblastoma multiforme (GBM)
- n.
- Hematolymphoid malignancies
4. Conclusions and Future Research Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J. How do you feel? A warm and touching 2021 Nobel tribute. J. Clin. Investig. 2021, 131, e156587. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A.; Blumberg, P.M. Vanilloid (capsaicin) receptors and mechanisms. Pharmacol. Rev. 1999, 51, 159–212. [Google Scholar]
- Cosens, D.J.; Manning, A. Abnormal electroretinogram from a Drosophila mutant. Nature 1969, 224, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Minke, B. Drosophila mutant with a transducer defect. Biophys. Struct. Mech. 1977, 3, 59–64. [Google Scholar] [CrossRef]
- Montell, C.; Birnbaumer, L.; Flockerzi, V.; Bindels, R.J.; Bruford, E.A.; Caterina, M.J.; Clapham, D.E.; Harteneck, C.; Heller, S.; Julius, D.; et al. A unified nomenclature for the superfamily of TRP cation channels. Moll. Cell 2002, 9, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E.; Montell, C.; Schultz, G.; Julius, D. International Union of Pharmacology. XLIII. Compendium of voltage-gated ion channels: Transient receptor potential channels. Pharmacol. Rev. 2003, 55, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.J.; Sweet, T.B.; Clapham, D.E. International Union of Basic and Clinical Pharmacology. LXXVI. Current progress in the mammalian TRP ion channel family. Pharmacol. Rev. 2010, 62, 381–404. [Google Scholar] [CrossRef]
- Nilius, B.; Owsianik, G.; Voets, T.; Peters, J. Transient receptor potential cation channels in disease. Physiol. Rev. 2007, 87, 165–217. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. Transient receptor potential channelopathies. Pflügers Arch. 2010, 460, 437–450. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. TRP channels: A TR(i)P through a world of multifunctional cation channels. Pflügers Arch. 2005, 451, 1–10. [Google Scholar] [CrossRef]
- Woodward, G.E.; Sage, C.O.; Rosado, J.A. Transient receptor potential channels and intracellular signaling. Int. Rev. Cytol. 2007, 35, 133–135. [Google Scholar]
- Zhang, K.; Julius, D.; Cheng, Y. Structural snapshots of TRPV1 reveal mechanism of polymodal functionality. Cell 2021, 184, 5138–5150. [Google Scholar] [CrossRef]
- Chard, P.S.; Bleakman, D.; Savidge, J.R.; Miller, R.J. Capsaicin-induced neurotoxicity in cultured dorsal root ganglion neurons: Involvement of calcium-activated proteases. Neuroscience 1995, 65, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- Jancsó, G.; Karcsú, S.; Király, E.; Szebeni, A.; Tóth, L.; Bácsy, E.; Joó, F.; Párducz, Á. Neurotoxin induced nerve degeneration: Possible involvement of calcium. Brain Res. 1984, 295, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Miyake, T.; Shirakawa, H.; Nakagawa, T.; Kaneko, S. Activation of mitochondrial transient receptor potential vanilloid 1 channel contributes to microglial migration. Glia 2015, 63, 1870–1881. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Morelli, M.B.; Aguzzi, C.; Zeppa, L.; Nabissi, M.; Polidori, C.; Santoni, G.; Amantini, C. Calcium influx, oxidative stress, and apoptosis induced by TRPV1 in chronic myeloid leukemia. Front. Mol. Biosci. 2023, 10, 1129202. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ávila, M.; Islas, L.D. What is new about mild temperature sensing? A review of recent findings. Temperature 2019, 2, 132–141. [Google Scholar] [CrossRef]
- Laursen, W.J.; Schneider, E.R.; Merriman, D.K.; Gracheva, E.O. Low-cost functional spasticity of TRPV1 supports heat tolerance in squirrels and camels. Proc. Natl. Acad. Sci. USA 2016, 113, 11342–11347. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Babaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Manitpisitkul, P.; Flores, C.M.; Moyer, J.A.; Romano, G.; Shalayda, K.; Tatikola, K.; Hutchison, J.S.; Mayorga, A.J. A multiple-dose, double-blind randomized study to evaluate the safety, pharmacokinetics, pharmacodynamics and analgesic efficacy of the TRPV1 antagonist, JNJ-39439335 (mavatrep). Scand. J. Pain 2018, 18, 151–164. [Google Scholar] [CrossRef] [PubMed]
- De Petrocellis, L.; Moriello, A.S. Modulation of the TRPV1 channel: Current clinical trials and recent patents with focus on neurological conditions. Recent Pat. CNS Drug Discov. 2013, 8, 180–204. [Google Scholar] [CrossRef]
- Yarmolinsky, D.A.; Peng, Y.; Pogorzala, L.A.; Rutlin, M.; Hoon, M.A.; Zuker, C.S. Coding and plasticity in the mammalian thermosensory system. Neuron 2016, 92, 1079–1092. [Google Scholar] [CrossRef] [PubMed]
- Karashima, Y.; Talavera, K.; Everaerts, W.; Janssens, A.; Kwan, K.Y.; Vennekens, R.; Nilius, B.; Voets, T. TRPA1 acts as a cold sensor in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1273–1278. [Google Scholar] [CrossRef]
- Story, G.M.; Peier, A.M.; Reeve, A.J. ANKTM1, a TRP-like channel expressed in nociceptive neurons, is activated by cold temperatures. Cell 2003, 112, 819–829. [Google Scholar] [CrossRef]
- Saito, S.; Nakatsuka, K.; Takahashi, K.; Fukuta, N.; Imagawa, T.; Ohta, T.; Tominaga, M. Analysis of transient receptor potential ankyrin 1 (TRPA1) in frogs and lizards illuminates both nociceptive heat and chemical sensitivities and coexpression with TRP Vanilloid 1 (TRPV1) in ancestral vertebrates. J. Biol. Chem. 2012, 287, 30743–30754. [Google Scholar] [CrossRef]
- Saito, S.; Banzawa, N.; Fukuta, N.; Saito, C.T.; Takahashi, K.; Imagawa, T.; Ohta, T.; Tominaga, M. Heat and noxious chemical sensor, chicken TRPA1, as a target for bird repellents and identification of its structural determinants by multispecies functional comparison. Mar. Mol. Biol. Evol. 2014, 313, 708–722. [Google Scholar] [CrossRef] [PubMed]
- Moparthi, L.; Kichko, T.I.; Eberhardt, M.; Högestätt, E.D.; Kjellbom, P.; Johanson, U.; Reeh, P.W.; Leffler, A.; Filipovic, M.R.; Zygmunt, P.M. Human TRPA1 is a heat sensor displaying intrinsic U-shaped thermosensitivity. Sci. Rep. 2016, 6, 28763. [Google Scholar] [CrossRef]
- Moparthi, L.; Sinica, V.; Moparthi, V.K.; Kreir, M.; Vignane, T.; Filipovic, M.R.; Vlachova, V.; Zygmunt, P.M. The human TRPA1 intrinsic cold and heat sensitivity involves separate channel structures beyond the N-ARD domain. Nat. Commun. 2022, 13, 6113. [Google Scholar] [CrossRef]
- Chen, J.; Kang, D.; Xu, J.; Lake, M.; Hogan, J.O.; Sun, C.; Walter, K.; Yao, B.; Kim, D. Species differences and molecular determinant of TRPA1 cold sensitivity. Nat. Commun. 2013, 4, 2501. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Nabhan, G.P. Seed dispersal. Directed deterrence by capsaicin in chilies. Nature 2001, 412, 403–404. [Google Scholar] [CrossRef]
- Jordt, S.E.; Julius, D. Molecular basis for species-specific sensitivity to “hot” chili peppers. Cell 2002, 108, 421–430. [Google Scholar] [CrossRef]
- Denda, M.; Tsutsumi, M. Roles of transient receptor potential proteins (TRPs) in epidermal keratinocytes. Adv. Exp. Med. Biol. 2011, 704, 847–860. [Google Scholar] [PubMed]
- Santoni, G.; Farfariello, V.; Amantini, C. TRPV channels in tumor growth and metastasis. Adv. Exp. Med. Biol. 2011, 704, 947–967. [Google Scholar]
- Shapovalov, G.; Ritaine, A.; Skryma, R.; Prevarskaya, N. Role of TRP channels in cancer and tumorigenesis. Semin. Immunopathol. 2016, 38, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.K.; Banham, A.H.; Yaacob, N.S.; Nur Husna, S.M. The oncogenic roles of TRPM ion channels in cancer. J. Cell. Physiol. 2019, 234, 14556–14573. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Wei, Y.; Chen, Y.; Ma, W.; Wang, M.; Chen, L.; Luo, Y.; Du, J. The role of transient receptor potential channels in metastasis. Biomed. Pharmacother. 2023, 158, 114074. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Gao, Y.; Xu, G.; Zhou, P.; Li, S.; Guo, J.; Zhou, H.; Xu, Q.; Huang, X.; Xu, J.; et al. Pan-cancer analyses reveal the genetic and pharmacogenomics landscape of transient receptor potential channels. NPJ Genom. Med. 2022, 7, 32. [Google Scholar] [CrossRef]
- Park, Y.R.; Chun, J.N.; So, I.; Kim, H.J.; Baek, S.; Jeon, J.-H.; Shin, S.-Y. Data-driven analysis of TRP channels in cancer: Linking variation in gene expression to clinical significance. Cancer Genom. Proteom. 2016, 13, 83–90. [Google Scholar]
- Bautista, D.M.; Movahed, P.; Hinman, A.; Axelsson, H.E.; Sterner, O.; Högestätt, E.D.; Julius, D.; Jordt, S.E.; Zygmunt, P.M. Pungent products form garlic activate the sensory ion channel TRPA1. Proc. Natl. Acad. Sci. USA 2005, 102, 12248–12252. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 mediates the inflammatory actions of environmental irritants and proalgesic agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef]
- Andrè, E.; Campi, B.; Materazzi, S.; Trevisani, M.; Amadesi, S.; Massi, D.; Creminon, C.; Vaksman, N.; Nassini, R.; Civelli, M.; et al. Cigarette smoke-inducedm neurogenic inflammation is mediated by α,β-unsaturated aldehydes and the TRPA1 receptors in rodents. J. Clin. Investig. 2008, 118, 2574–2582. [Google Scholar] [PubMed]
- Deering-Rice, C.E.; Romero, E.G.; Shapiro, D.; Hughen, R.W.; Light, A.R.; Yost, G.S.; Veranth, J.M.; Reilly, C.A. Electrophilic components of diesel exhaust particles (DEP) activate transient receptor potential ankyrin-1 (TRPA1): A probable mechanism of acute pulmonary toxicity for DEP. Chem. Res. Toxicol. 2011, 24, 950–959. [Google Scholar] [CrossRef]
- Kang, K.; Pulver, S.R.; Panzano, V.C.; Chang, E.C.; Griffith, L.C.; Theobald, D.L.; Garrity, P.A. Analysis of Drosophila TRPA1 reveals an ancient origin for human chemical nociception. Nature 2010, 464, 597–600. [Google Scholar] [CrossRef] [PubMed]
- Melo, N.; Capek, M.; Arenas, O.M.; Afify, A.; Yilmaz, A.; Potter, C.J.; Laminette, P.J.; Para, A.; Gallio, M.; Stensmyr, M.C. The irritant receptor TRPA1 mediates the mosquito repellant effect of catnip. Curr. Biol. 2021, 31, 1988–1994. [Google Scholar] [CrossRef]
- Inocente, E.A.; Shaya, M.; Acosta, M.; Rakotondraibe, L.H.; Piermarini, P.M. A natural agonist of mosquito TRPA1 from the medicinal plant Cinnamosma fragrans that is toxic, antifeedant, and repellent to the yellow fever mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006265. [Google Scholar] [CrossRef] [PubMed]
- Du, E.J.; Ahn, T.J.; Choi, M.S.; Kwon, I.; Kim, H.W.; Kwon, J.Y.; Kang, K. The mosquito repellent citronellal directly potentiates Drosophila TRPA1m facilitating feeding suppression. Mol. Cells. 2015, 38, 911–917. [Google Scholar] [PubMed]
- Kwon, Y.; Kim, A.H.; Ronderos, D.S.; Lee, Y.; Akitake, B.; Woodward, O.M.; Guggino, W.B.; Smith, D.P.; Montell, C. Drosophila TRPA1 channel is required to avoid the naturally occurring insect repellent citronella. Curr. Biol. 2010, 20, 1672–1678. [Google Scholar] [CrossRef]
- Kremeyer, B.; Lopera, F.; Cox, J.J.; Momin, A.; Rugiero, F.; Marsh, S.; Woods, C.G.; Jones, N.G.; Paterson, K.J.; Fricker, F.R.; et al. A gain-of-function mutation in TRPA1 causes familial episodic pain syndrome. Neuron 2010, 66, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.-P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug. Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Gratzke, C.; Strengm, T.; Waldkirch, E.; Sigl, K.; Stief, C.; Andersson, K.E.; Hedlund, P. Transient receptor potential A1 (TRPA1) activity in the human urethra—Evidence for a functional role for TRPA1 in the outflow region. Eur. Urol. 2009, 55, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.H.; Jhang, J.F.; Birder, L.A.; Kuo, H.C. Sensory receptor, inflammatory and apoptotic protein expression in the bladder urothelium of patients with different subtypes of interstitital cystitis/bladder pain syndrome. Int. J. Mol. Sci. 2023, 24, 820. [Google Scholar] [CrossRef] [PubMed]
- Thakore, P.; Ali, S.; Earley, S. Regulation of vascular tone by transient receptor potential ankyrin 1 channels. Curr. Top. Membr. 2020, 85, 119–150. [Google Scholar] [PubMed]
- Atoyan, R.; Shander, D.; Botchkareva, N.V. Non-neuronal expression of transient receptor potential type A1 (TRPA1) in human skin. J. Investig. Dermatol. 2009, 129, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Bohonyi, N.; Pohóczky, K.; Szalontai, B.; Perkecz, A.; Kovács, K.; Kajtár, B.; Orbán, L.; Varga, T.; Szegedi, S.; Bódis, J.; et al. Local upregulation of transient receptor potential ankyrin 1 and transient receptor potential vanilloid 1 ion channels in rectosigmoid deep infiltrating endometriosis. Mol. Pain 2017, 13, 1744806917705564. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, H.K.; Kwon, T.K.; Kim, C.S.; Cho, J.H.; Ahn, D.K.; Bae, Y.C. Expression of transient receptor potential ankyrin 1 in human dental pulp. J. Endod. 2012, 38, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, L.J.; Paukkeri, E.-L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient receptor potential ankyrin 1 (TRPA1) is functionally expressed in primary human osteoarthritic chondrocytes. Arthritis Res. Ther. 2016, 18, 185. [Google Scholar] [CrossRef]
- Okata, T.; Shimizu, S.; Wakamori, M.; Maeda, A.; Kurosaki, T.; Takada, N.; Imoto, K.; Mori, Y. Molecular cloning and functional characterization of a novel receptor-activated TRP Ca2+ channel from mouse brain. J. Biol. Chem. 1998, 273, 10279–10287. [Google Scholar] [CrossRef]
- Philipp, S.; Hambrechtm, J.; Braslavski, L.; Schroth, G.; Freichel, M.; Murakami, M.; Cavaliè, A.; Flockerzi, V. A novel capacitative calcium entry channel expressed in excitable cells. EMBO J. 1998, 17, 4274–4282. [Google Scholar] [CrossRef]
- Sossey-Alaoui, K.; Lyon, J.A.; Jones, L.; Abidi, F.E.; Hartung, A.J.; Hane, B.; Schwartz, C.E.; Stevenson, R.E.; Srivastava, A.K. Molecular cloning and characterization of TRPC5 (HTRP5), then human homologue of mouse brain receptor-activated capacitative Ca2+ entry channel. Genomics 1999, 60, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Clarson, L.H.; Roberts, V.H.; Hamark, B.; Elliott, A.C.; Powell, T. Store-operated Ca2+ entry in first trimester and term human placenta. J. Physiol. 2003, 550, 515–528. [Google Scholar] [CrossRef]
- Cai, S.; Fatherazi, S.; Presland, R.B.; Belton, C.M.; Izutsu, K.T. TRPC channel expression during calcium-induced differentiation of human gingival keratinocytes. J. Dermatol. Sci. 2005, 40, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bernal, L.; Sotelo-Hitschfeld, P.; König, C.; Sinica, V.; Wyatt, A.; Winter, Z.; Hein, A.; Reinhard, S.; Tragl, A.; Kusuda, R.; et al. Odontoblast TRPC5 channels signal cold pain in teeth. Sci. Adv. 2021, 7, eabf5567. [Google Scholar] [CrossRef]
- Yip, H.; Chan, W.Y.; Leung, P.C.; Kwan, H.Y.; Liu, C.; Huang, Y.; Michel, V.; Yew, D.T.; Yao, X. Expression of TRPC homologs in endothelial cells and smooth muscle layers of human arteries. Histochem. Cell Biol. 2004, 122, 553–561. [Google Scholar] [CrossRef]
- Tian, D.; Jacobo, S.M.P.; Billing, D.; Rozkalne, A.; Gage, S.D.; Anagnostou, T.; Pavenstädt, H.; Hsu, H.-H.; Schlondorff, J.; Ramos, A.; et al. Antagonistic regulation of actin dynamics and cell motility by TRPC5 and TRPC6 channels. Sci. Signal. 2010, 3, ra77. [Google Scholar] [CrossRef] [PubMed]
- Greka, A.; Mundel, P. Balancing calcium signals through TRPC5 and TRPC6 in podocytes. J. Am. Soc. Nephrol. 2011, 22, 1969–1980. [Google Scholar] [CrossRef]
- Zhou, Y.; Castonguay, P.; Sidhom, E.-H.; Clark, A.R.; Dvela-Levitt, M.; Kim, S.; Sieber, J.; Wieder, N.; Jung, J.Y.; Andreeva, S.; et al. A small molecule inhibitor of TRPC5 ion channels suppresses progressive kidney disease in animal models. Science 2017, 358, 1332–1336. [Google Scholar] [CrossRef]
- Liu, D.; Scholze, A.; Zhu, Z.; Krueger, K.; Thilo, F.; Burkert, A.; Strefferm, K.; Holz, S.; Harteneck, C.; Zidek, W.; et al. Transient receptor potential channels in essential hypertension. J. Hypertens. 2006, 24, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Greka, A.; Navarro, B.; Oancea, E.; Duggan, A.; Clapham, D.E. TRPC5 is a regulator of hippocampal neurote length and growth cone morphology. Nat. Neurosci. 2003, 6, 837–845. [Google Scholar] [CrossRef]
- Zimmermann, K.; Lennerz, J.K.; Hein, A.; Link, A.S.; Kaczmarek, J.S.; Delling, M.; Uysal, S.; Riccio, A.; Clapham, D.E. Transient receptor potential cation channel, subfamily C, member 5 (TRPC5) is a cold-transducer in the peripheral nervous system. Proc. Natl. Acad. Sci. USA 2011, 108, 18114–18119. [Google Scholar] [CrossRef]
- Ptakova, A.; Mitro, M.; Zimova, L.; Vlachova, V. Cellular context determines primary characteristics of human TRPC5 as a cold-activated channel. J. Cell Physiol. 2022, 237, 3614–3626. [Google Scholar] [CrossRef]
- Nagamine, K.; Kudoh, J.; Minoshima, S.; Kawasaki, K.; Asakawa, S.; Ito, F.; Shimizu, N. Molecular cloning of a novel putative Ca2+ channel protein (TRPC7) highly expressed in brain. Genomics 1998, 54, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Vannier, B.; Peyton, M.; Boulay, G.; Brown, D.; Qin, N.; Jiang, M.; Zhu, X.; Birnbaumer, L. Mouse trp2, the homologue of the human trpc2 pseudogene, encodes mTrp2, a store depletion-activated capacitative Ca2+ channel. Proc. Natl. Acad. Sci. USA 1999, 96, 2060–2064. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Inoue, R.; Yamazaki, K.; Maeda, A.; Kurosaki, T.; Yamakuni, T.; Tanaka, I.; Shimizu, S.; Ikenada, K.; Imoto, K.; et al. Molecular and functional characterization of a novel mouse brain transient receptor potential protein homologue TRP7. Ca2+-permeable cation channel that is constitutively activated and enhanced by stimulation of G protein-coupled receptor. J. Biol. Chem. 1999, 274, 27359–27370. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.H.; Yang, W.; Zou, J.; Beech, D. TRPM2 channel properties, functions and therapeutic potentials. Expert Opin. Ther. Targets 2010, 14, 973–988. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, N.; Ni, Y.S.; Yang, J.M.; Ma, L.; Lan, X.B.; Wu, J.; Niu, J.G.; Yu, J.Q. TRPM2 in ischemic stroke: Structure, molecular mechanisms, and drug intervention. Channels 2021, 15, 136–154. [Google Scholar] [CrossRef]
- Hiroi, T.; Wajima, T.; Negoro, T.; Ishii, M.; Nakano, Y.; Kiuchi, Y.; Mori, Y.; Shimizu, S. Neutrophil TRPM2 channels are implicated in the exacerbation of myocardial ischemia/reperfusion injury. Cardiovasc. Dis. 2013, 97, 271–278. [Google Scholar]
- Malko, P.; Syed Mortadza, S.A.; McWilliam, J.; Jiang, L.-H. TRPM2 channels in microglia as anew player in neuroinflammation associated with a spectrum of central nervous system pathologies. Front. Pharmacol. 2019, 10, 239. [Google Scholar] [CrossRef]
- Tan, C.-H.; McNaughton, P.A. The TRPM2 ion channel is required for sensitivity to warmth. Nature 2016, 536, 460–463. [Google Scholar] [CrossRef]
- Paracio-Montesinos, R.; Schwaller, F.; Udhayachandran, A.; Rau, F.; Walcher, J.; Evangelista, R.; Vriens, J.; Voets, T.; Poulet, J.F.A.; Lewin, G.J. The sensory coding of warm perception. Neuron 2020, 106, 830–841. [Google Scholar] [CrossRef]
- Kamm, G.B.; Boffi, J.C.; Zuza, K.; Nencini, S.; Campos, J.; Schrenk-Siemens, K.; Sonntag, I.; Kabaoglu, B.; Abd Al Hay, M.Y.; Schwartz, Y.; et al. A synaptic temperature sensor for body cooling. Neuron 2021, 109, 3283–3297. [Google Scholar] [CrossRef]
- Grimm, C.; Kraft, R.; Sauerbruch, S.; Schultz, G.; Harteneck, C. Molecular and functional characterization of the melastatin-related cation channel TRPM3. J. Biol. Chem. 2003, 278, 21493–21501. [Google Scholar] [CrossRef]
- Lee, N.; Chen, J.; Sun, L.; Wu, S.; Gray, K.R.; Rich, A.; Huang, M.; Lin, J.-H.; Feder, J.N.; Janovitz, E.B.; et al. Expression and characterization of human transient receptor potential melastatin 3 (hTRPM3). J. Biol. Chem. 2003, 278, 20890–20897. [Google Scholar] [CrossRef]
- Fonfria, E.; Murdock, P.R.; Cusdin, F.S.; Benham, C.D.; Kelsell, R.E.; McNultym, S. Issue distribution profiles of the human TRPM cation channel family. J. Recept. Signal Transduct. 2006, 26, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Wagner, T.F.J.; Loch, S.; Lambert, S.; Straub, I.; Mannebach, S.; Mathar, I.; Düfer, M.; Lis, A.; Flockerzi, V.; Philipp, S.E.; et al. Transient receptor potential M3 channels are ionotropic steroid receptors in pancreatic beta cells. Nat. Cell. Biol. 2008, 10, 1421–1430. [Google Scholar] [CrossRef]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Bennett, T.M.; Mackay, D.S.; Siegfried, C.J.; Shiels, A. Mutation in the melastatin-related cation channels, TRPM3, underlies inherited cataract and glaucoma. PLoS ONE 2014, 9, e104000. [Google Scholar] [CrossRef] [PubMed]
- Burglen, L.; van Hoeymissen, E.; Qebibo, L.; Barth, M.; Belnap, N.; Boschann, F.; Depienne, C.; De Clercq, K.; Douglas, A.G.L.; Fitzgerald, M.P.; et al. Gain-of-function variants in the ion channel gene TRPM3 underline a spectrum of neurodevelopmental disorders. eLife 2023, 12, e81032. [Google Scholar] [CrossRef] [PubMed]
- Shawn, X.-Z.; Moebius, F.; Gill, D.L.; Montell, C. Regulation of melastatin, a TRP-related protein, through interaction with a cytoplasmic isoform. Proc. Natl. Acad. Sci. USA 2001, 98, 10692–10697. [Google Scholar]
- Nilius, B.; Prenen, J.; Droogmans, G.; Voets, T.; Vennekens, R.; Freichel, M.; Wissenbach, U.; Flockerzi, V. Voltage dependence of the Ca2+-activated cation channel TRPM4. J. Biol. Chem. 2003, 278, 30813–30820. [Google Scholar] [CrossRef] [PubMed]
- Launay, P.; Cheng, H.; Srivatsan, S.; Penner, R.; Fleig, A.; Kinet, J.-P. TRPM4 regulates calcium oscillations after T-cell activation. Science 2004, 306, 1374–1377. [Google Scholar] [CrossRef]
- Earley, S.; Waldron, B.J.; Brayden, J.E. Critical role for transient receptor potential channel TRPM4 in myogenic constriction of cerebral arteries. Circ. Res. 2004, 96, 922–929. [Google Scholar] [CrossRef]
- Ali, S.; Solano, A.S.; Gonzales, A.L.; Thakore, P.; Krishnan, V.; Yamasaki, E.; Earley, S. Nitric oxide signals through IRAG to inhibit TRPM4 channels and dilate cerebral arteries. Function 2021, 2, yqab051. [Google Scholar] [CrossRef]
- Mehta, R.I.; Tosun, C.; Ivanova, S.; Tsymbalyuk, N.; Famakin, B.M.; Kwon, M.S.; Castellani, R.J.; Gerzanichm, V.; Simard, J.M. Sur1-Trpm4 cation channel expression in human cerebral infarcts. J. Neuropathol. Exp. Neurol. 2015, 74, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Stallmeyer, B.; Zumhagen, S.; Denjoy, I.; Duthoit, G.; Hebert, J.L.; Ferrer, X.; Maugenre, S.; Schmitz, W.; Kirchhefer, U.; Schulze-Bahn, E.; et al. Mutational spectrum in the Ca2+-activated cation channel gene TRPM4 in patients with cardiac conductance disturbances. Hum. Mutat. 2012, 33, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chatel, S.; Simard, C.; Syam, N.; Salle, L.; Probst, V.; Morel, J.; Millat, G.; Lopez, M.; Abriel, H.; et al. Molecular genetics and functional anomalies in a series of 248 Brugada cases with 11 mutations in the TRPM4 channel. PLoS ONE 2013, 8, e54131. [Google Scholar] [CrossRef]
- Talevara, K.; Yasumatsu, K.; Voets, T.; Droogmans, G.; Shigemura, N.; Ninomiya, Y.; Margolskee, R.F.; Nilius, B. Heat activation of TRPM5 underlies thermal sensitivity of taste. Nature 2005, 438, 1022–1025. [Google Scholar] [CrossRef] [PubMed]
- Pèrez, C.A.; Huang, L.; Rong, M.; Kozak, J.A.; Preuss, A.K.; Zhang, H.; Max, M.; Margolskee, R.F. A transient receptor potential channel expressed in taste receptor cells. Nat. Neurosci. 2002, 5, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hoon, M.A.; Chabndrashekar, J.; Mueller, K.L.; Cook, B.; Wu, D.; Zuker, C.S.; Ryba, N.J.P. Coding of sweet, bitter and unami tastes: Different receptor cells sharing similar signaling pathways. Cell 2003, 112, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Mardaryev, A.; van Lessen, M.; Alam, M.; Acosta, F.J.; Bíró, T.; Paus, R. Hair follicle chemosensation: TRPM5 signaling is required for anagen maintenance. J. Investig. Dermatol. 2021, 141, 2300–2303. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.J.; Glockner, M.; Schicht, M.; Bräuer, L.; Paulsen, F. Detection of intrinsic cholinergic system in human lacrimal drainage system: Evidence and potential implications. Graefes Arch. Clin. Exp. Ophthalmol. 2018, 256, 2097–2101. [Google Scholar] [CrossRef]
- Kyriazis, G.A.; Soundarapandian, M.M.; Tyrberg, B. Sweet taste receptor signaling in beta cells mediates fructose-induced potentiation of glucose-stimulated insulin secretion. Proc. Natl. Acad. Sci. USA 2012, 109, E524–E532. [Google Scholar] [CrossRef]
- Barham, H.P.; Cooper, S.E.; Anderson, C.B.; Tizzano, M.; Kingdom, T.T.; Finger, T.E.; Kinnamon, S.C.; Ramakrishnan, V.R. Solitary chemosensory cells and bitter taste receptor signaling in human sinonasal mucosa. Int. Forum Allergy Rhinol. 2013, 3, 450–457. [Google Scholar] [CrossRef]
- Bezencon, C.; le Coutre, J.; Damak, S. Taste-signaling proteins are coexpressed in solitary intestinal epithelial cells. Chem. Senses 2007, 32, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, S.; Nogueira, C.; Cantero-Recasens, G.; Kiefer, K.; Fernandez-Fernandez, J.M.; Popoff, J.F.; Casano, L.; Bard, F.A.; Gomez, R.; Valverde, M.A.; et al. TRPM5-mediated calcium uptake regulates mucin secretion from human colon goblet cells. eLife 2013, 2, e00658. [Google Scholar] [CrossRef]
- Colsoul, B.; Schraenen, A.; Lemaire, K.; Quintens, R.; Van Lommel, L.; Segal, A.; Owsianik, G.; Talavera, K.; Voets, T.; Margolskee, R.F.; et al. Loss of high-frequency glucose-induced Ca2+ oscillations in pancreatic islets correlates with impaired glucose tolerance in Trpm5-/- mice. Proc. Natl. Acad. Sci. USA 2010, 107, 5208–5213. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Stein, R.J.; Santos, S.; Nagatomi, J.; Hayashi, Y.; Minnery, B.S.; Xavier, M.; Patel, A.S.; Nelson, J.B.; Futrell, W.J.; Yoshimura, N.; et al. Cool (TRPM8) and hot (TRPV1) receptors in the bladder and male genital tract. J. Urol. 2004, 172, 1175–1178. [Google Scholar] [CrossRef]
- Kim, J.H.; Jang, Y.S.; Kim, H.I.; Park, J.Y.; Park, S.H.; Hwang, Y.I.; Jang, S.H.; Jung, K.S.; Park, H.S.; Park, C.S. Activation of transient receptor potential melastatin family member 8 (TRPM8) receptors induces proinflammatory cytokine expression in bronchial epithelial cells. Allergy Asthma Immunol. Res. 2020, 12, 684–700. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, K.; Ikeda, H.; Kawashima, N.; Okiji, T. Transient receptor potential melastatin (TRPM) 8 is expressed in freshly isolated native human odontoblasts. Arch. Oral Biol. 2017, 75, 55–61. [Google Scholar] [CrossRef]
- Rossato, M.; Granzotto, M.; Macchi, V.; Porzionato, A.; Petrelli, L.; Calcagno, A.; Vencato, J.; De Stefani, D.; Silvestrin, V.; Rizzuto, R.; et al. Human whitge adipocytes express the cold receptor TRPM8 which activation induces UCP1 expression, mitochrondrial activation and heat production. Mol. Cell. Endocrinol. 2014, 383, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Parra, A.; Madrid, R.; Eschevarria, D.; del Olmo, S.; Morenilla-Palao, C.; Acosta, M.C.; Gallar, J.; Dhaka, A.; Viana, F.; Belmonte, C. Ocular wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 2010, 16, 1396–1399. [Google Scholar] [CrossRef]
- Yang, J.M.; Liu, Q.; Rüedi, M.; Wei, E.T.; Lentsman, M.; Lee, H.S.; Choi, W.; Kim, S.J.; Yoon, K.C. A novel TRPM8 agonist relieves dry eye discomfort. BMC Ophthalmol. 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed]
- Weyer, A.D.; Lehto, S.G. Development of TRPM8 antagonists to treat chronic pain and migraine. Pharmaceuticals 2017, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.J.; Kim, J.C.; Wei, E.T.; Selescu, T.; Chung, B.Y.; Park, C.W.; Kim, H.O. A randomized, vehicle-controlled clinical trial of a synthetic TRPM8 agonist (Cryosim-1) gel for itch. J. Am. Acad. Dermatol. 2021, 84, 869–871. [Google Scholar] [CrossRef]
- NCT04866563. Available online: https://clinicaltrials.gov (accessed on 1 April 2023).
- NCT00839631. Available online: https//clinicaltrials.gov (accessed on 1 April 2023).
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Kress, M.; Zeilhofer, H.U. Capsaicin, protons and heat; new excitement about nociceptors. Trends Pharmacol. Sci. 1999, 20, 112–118. [Google Scholar] [CrossRef]
- Siemens, J.; Zhou, S.; Piskorowski, R.; Nikai, T.; Lumpkin, E.A.; Basbaum, A.I.; King, D.; Julius, D. Spider toxins activate the capsaicin receptor to produce inflammatory pain. Nature 2006, 444, 208–212. [Google Scholar] [CrossRef]
- Cuypers, E.; Yanagihara, A.; Karlsson, E.; Tytgatm, J. Jellyfish and other cnidarian envenomations cause pain by affecting TRPV1 channels. FEBS Lett. 2006, 580, 5728–5732. [Google Scholar] [CrossRef] [PubMed]
- Meza, R.C.; Ancatén-González, C.; Chiu, C.; Chávez, A.E. Transient receptor potential vanilloid 1 function at central synapses in health and disease. Front. Cell. Neurosci. 2022, 16, 864828. [Google Scholar] [CrossRef]
- Ständer, S.; Moormann, C.; Schumacher, M.; Buddenkotte, J.; Artuc, M.; Shapacovitch, V.; Brzoska, T.; Lippert, U.; Henz, B.M.; Luger, T.A.; et al. Expression of vanilloid receptor subtype 1 in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Exp. Dermatol. 2004, 13, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Omari, S.A.; Adams, M.J.; Geraghty, D.P. TRPV1 channels in immune cells and hematological malignancies. Adv. Pharmacol. 2017, 79, 173–198. [Google Scholar]
- Phan, T.X.; Ton, H.T.; Gulyás, H.; Pórszász, R.; Tóth, A.; Russo, R.; Kay, M.W.; Sahibzada, N.; Ahern, G.P. TRPV1 expressed throughout the arterial circulation regulates vasoconstriction and blood pressure. J. Physiol. 2020, 598, 5639–5659. [Google Scholar] [CrossRef]
- Szolcsányi, J. Effect of capsaicin on thermoregulation: An update with new aspects. Temperature 2015, 2, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Zsombok, A.; Derbenev, A.V. TRP channels as therapeutic targets in diabetes and obesity. Pharmaceuticals 2016, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Christie, S.; Wittert, G.A.; Li, H.; Page, A.J. Involvement of TRPV1 channels in energy homeostasis. Front. Endocrinol. 2018, 9, 420. [Google Scholar] [CrossRef]
- Hollis, M.; Wang, D.H. Transient receptor potential vanilloid in blood pressure regulation. Curr. Opin. Nephrol. Hypertens. 2013, 22, 170–176. [Google Scholar] [CrossRef] [PubMed]
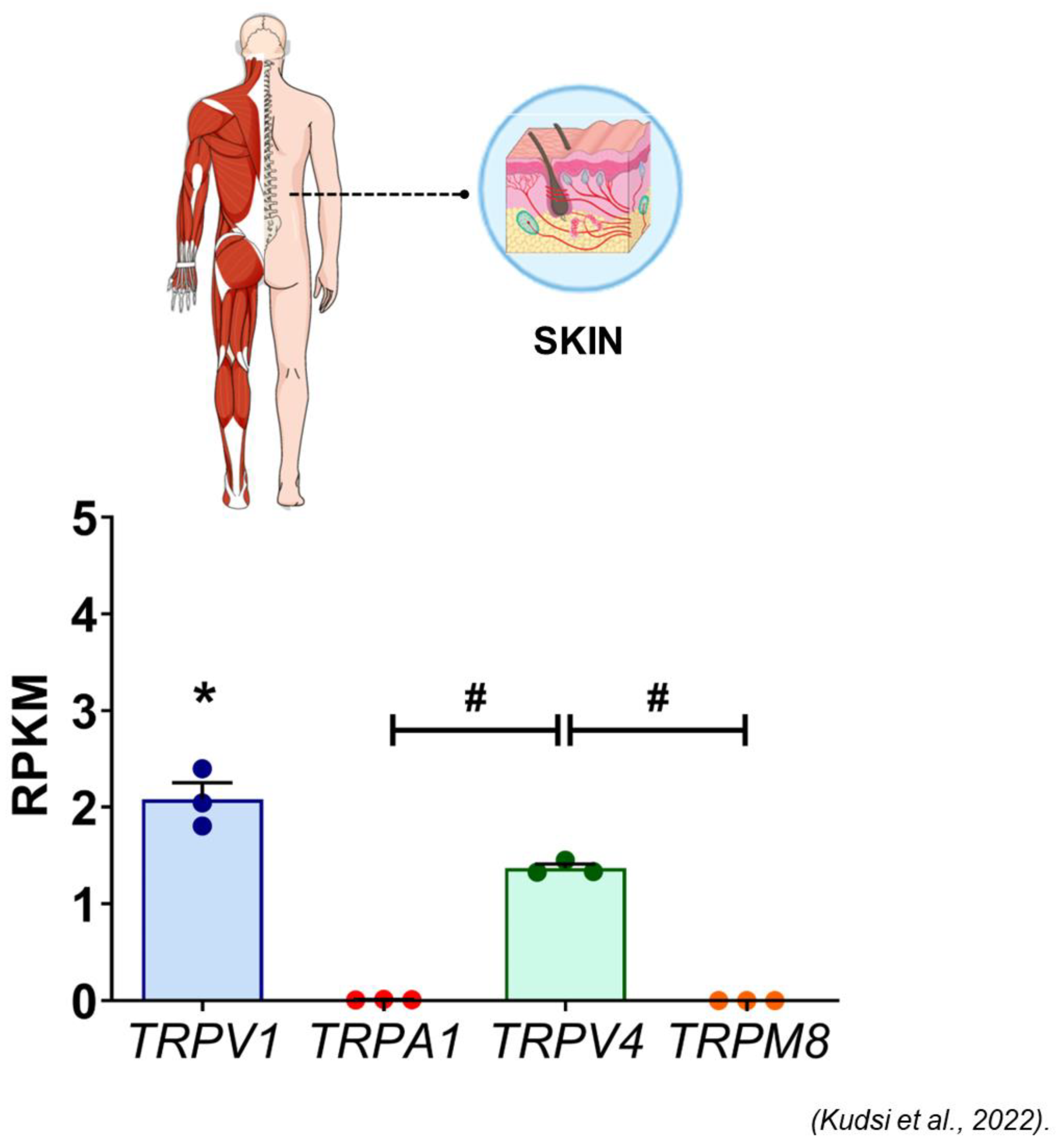
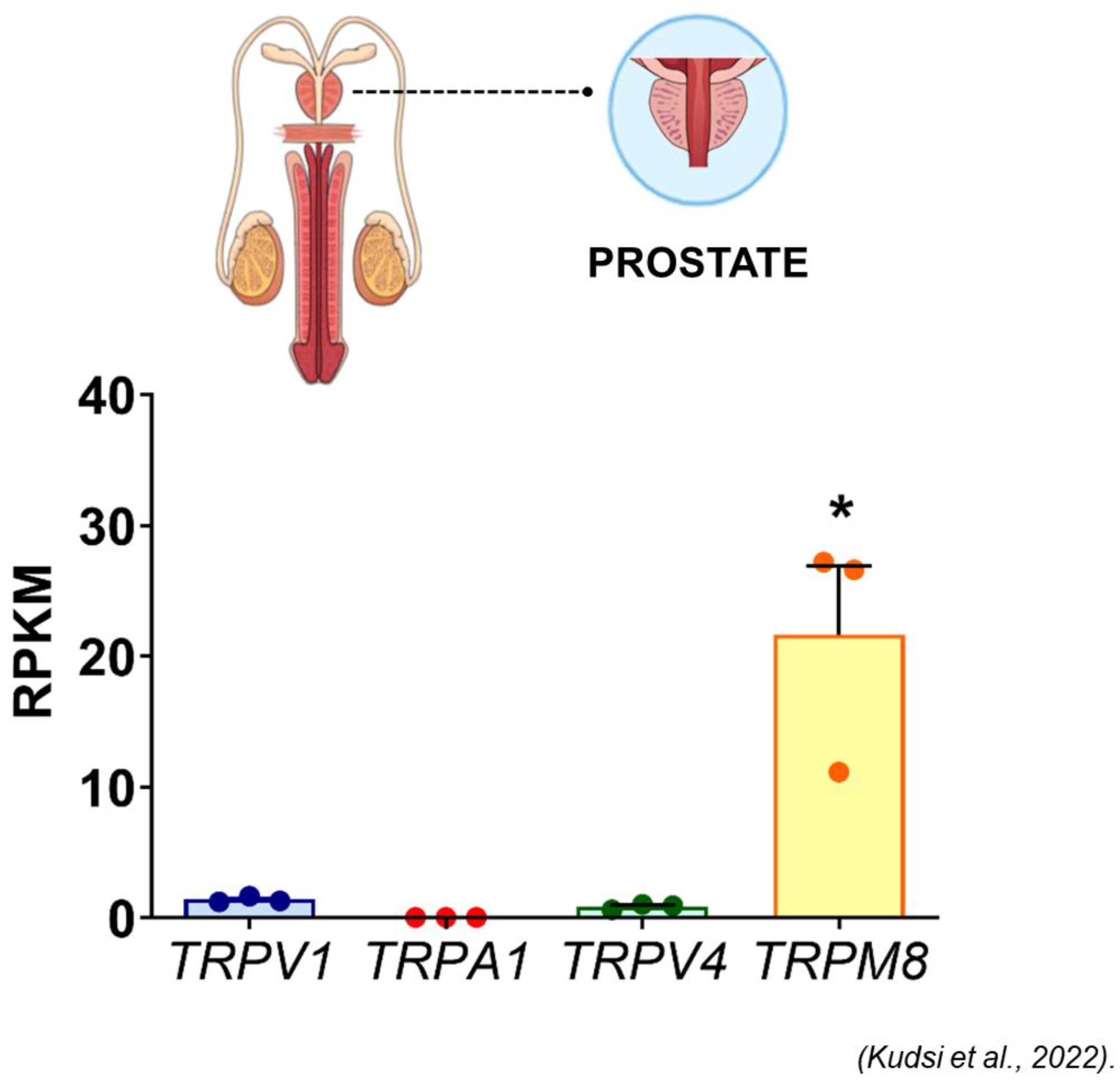
- Kudsi, S.Q.; Piccoli, B.C.; Ardisson-Araújo, D.; Trevisan, G. Transcriptional landscape of TRPV1, TRPA1, TRPV4, and TRPM8 channels throughout human tissues. Life Sci. 2022, 308, 120977. [Google Scholar] [CrossRef]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, cancer and immunity-implication of TRPV1 channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef]
- Erin, N. Role of sensory neurons, neuroimmune pathways, and transient receptor potential vanilloid 1 (TRPV1) channels in a murine model of breast cancer. Cancer Immunol. Immunother. 2020, 69, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Balood, M.; Ahmadi, M.; Eichwald, T.; Ahmadi, A.; Majdoubi, A.; Roversi, K.; Roversi, K.; Lucido, C.T.; Restaino, A.C.; Huang, S.; et al. Nociceptive neurons affect cancer immmunosurveillance. Nature 2022, 611, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Rosen, T.A.; Tominaga, M.; Brake, A.J.; Julius, D. A capsaicin-receptor homologue with a high threshold for heat. Nature 1999, 398, 436–441. [Google Scholar] [CrossRef]
- Muraki, K.; Iwata, Y.; Katanosaka, Y.; Ito, T.; Ohya, S.; Shigekawa, M.; Imaizumi, Y. TRPV2 is a component of osmotically sensitive cation channels in murine aortic myocytes. Circ. Res. 2003, 93, 829–838. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Ding, W.; Zhan, T.; Zhu, J.; Zhang, L.; Wang, H.; Shen, B.; Wang, Y. Oxidative stress-induced TRPV2 expression increase is involved in diabetic cataracts and apoptosis of lens epithelial cells in a high-glucose environment. Cells 2011, 11, 1196. [Google Scholar] [CrossRef] [PubMed]
- De Clecq, K.; Held, K.; Van Bree, R.; Meuleman, C.; Peeraer, K.; Tomassetti, C.; Voets, T.; O’Hooghe, T.; Vriens, J. Functional expression of transient receptor potential channels in human endometrial stromal cells during the luteal phase of the menstrual cycle. Hum. Reprod. 2015, 30, 1421–1436. [Google Scholar] [CrossRef]
- Muraki, K.; Shigekawa, M.; Imaizumi, Y. A new insight into the function of TRPV2 in circulatory organs. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedtke, W.B., Heller, S., Eds.; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2007; Chapter 28. [Google Scholar]
- Delany, N.S.; Hurle, M.; Facer, P.; Alnadaf, T.; Plumpton, C.; Kinghorn, I.; See, C.G.; Costigan, M.; Anand, P.; Woolf, C.J.; et al. Identification and characterization of a novel human vanilloid receptor-like protein, VRL-2. Physiol. Genom. 2001, 4, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Earley, T.J.; Hergarden, A.C.; Story, G.M.; Colley, S.; Hogenesch, J.B.; McIntyre, P.; et al. A heat-sensitive TRP channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotacha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. TRPV3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef]
- Smith, G.D.; Gunthorpe, M.J.; Kessell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Chen, Q.; Lee, M.; Cao, X.; Zhang, J.; Ma, D.; Chen, L.; Hu, X.; Wang, H.; Wang, X.; et al. Exome sequencing reveals mutations in TRPV3 as a cause of Olmsted syndrome. Am. J. Hum. Genet. 2013, 90, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Imura, K.; Yoshioka, T.; Hirasawa, T.; Sakata, T. Role of TRPV3 in immune response to development of dermatitis. J. Imflamm. 2009, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Borbíró, I.; Lisztes, E.; Tóth, B.I.; Czifra, G.; Oláh, A.; Szöllősi, A.G.; Szentandrássy, N.; Nánási, P.P.; Pèter, Z.; Paus, R.; et al. Activation of transient receptor potential vanilloid-3 inhibits human hair growth. J. Investig. Dermatol. 2011, 131, 1605–1614. [Google Scholar] [CrossRef]
- Toledo Maurino, J.J.; Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Barreto-Zuniga, R.; Martínez Benítez, B.; Granados, J.; Yamamoto-Furusho, J.K. TRPV subfamily (TRPV2, TRPV3, TRPV4, TRPV5 and TRPV6) gene and protein expression in patients with ulcerative colititis. J. Immunol. Res. 2020, 2020, 2906845. [Google Scholar] [CrossRef]
- Broad, L.M.; Moggm, A.J.; Eberle, E.; Tolley, M.; Li, D.L.; Knopp, K.L. TRPV3 in drug development. Pharmaceuticals 2016, 9, 55. [Google Scholar] [CrossRef]
- Nilius, B.; Prenen, J.; Wissenbach, U.; Bödding, M.; Droogmans, G. Differential activation of the volume-sensitive cation channel TRP12 (OTRPC4) and volume-regulated anion currents in HEK-293 cells. Pflügers Arch. 2001, 443, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Strotman, R.; Harteneck, C.; Nunnenmacher, K.; Schultz, G.; Plant, T.D. OTRPC4, a nonselective cation channel that confers sensitivity to extracellular osmolarity. Nat. Cell Biol. 2000, 2, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, W.; Friedman, J.M. Abnormal osmotic regulation in trpv4-/- mice. Proc. Natl. Acad. Sci. USA 2003, 100, 13698–13703. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Mizuno, A.; Kodaira, K.; Imai, M. Impaired pressure sensation in mice lacking TRPV4. J. Biol. Chem. 2003, 278, 22664–22668. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.K.; Lee, H.; Caterina, M.J. Warm temperatures activate TRPV4 in mouse 308 keratinocytes. J. Biol. Chem. 2003, 278, 32037–32046. [Google Scholar] [CrossRef]
- Zhou, X.; Su, Y.; Wu, S.; Wangm, H.; Jiang, R.; Jiang, X. The temperature-sensitive receptors TRPV4 and TRPM8 have important roles in the pruritus of rosacea. J. Dermatol. Sci. 2022, 108, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Mundt, N.; Spehr, M.; Lishko, P.V. TRPV4 is the temperature-sensitive ion channel of human sperm. eLife 2018, 7, e35853. [Google Scholar] [CrossRef] [PubMed]
- Plant, T.D.; Strotmann, R. TRPV4. Handb. Exp. Pharmacol. 2007, 179, 189–205. [Google Scholar]
- Dai, J.; Cho, T.J.; Unger, S.; Lausch, E.; Nishimura, G.; Kim, O.H.; Supeti-Furga, A.; Ikegawa, S. TRPV4-pathy, a novel channelopathy affecting diverse systems. J. Hum. Genet. 2010, 55, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Voets, T. The puzzle of TRPV4 channelopathies. EMBO Rep. 2013, 14, 152–163. [Google Scholar] [CrossRef]
- Falini, B.; Pileri, S.; Zinzani, P.L.; Carbone, A.; Zagonel, V.; Wolf-Peeters, C.; Verhoef, G.; Menestrina, F.; Todeschini, G.; Paulli, M.; et al. ALK+ lymphoma: Clinico-pathological findings are outcome. Blood 1999, 93, 2697–2706. [Google Scholar]
- Gouda, M.A.; Subbiah, V. Precision oncology for BRAF-mutant cancers with BRAF and MEK inhibitors: From melanoma to tissue-agnostic therapy. ESMO Open. 2023, 8, 100788. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, M.M. Trastuzumab, a recombinant DNA-derived humanized monoclonal antibody, a novel agent for the treatment of metastatic breast cancer. Clin. Ther. 1999, 21, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.D. Rituximab: A new therapeutic monoclonal antibody for non-Hodgkin lymphoma. Cancer Pract. 1998, 6, 195–197. [Google Scholar] [CrossRef]
- Foyil, K.V.; Bartlett, N.L. Anti-CD30 antibodies for Hodgkin lymphoma. Curr. Hematol. Malig. Rep. 2010, 5, 140–147. [Google Scholar] [CrossRef]
- Virk, H.S.; Rekas, M.Z.; Biddle, M.S.; Wright, A.K.A.; Sousa, J.; Weston, C.A.; Chachi, L.; Roach, K.M.; Bradding, P. Validation of antibodies for the specific detection of human TRPA1. Sci. Rep. 2019, 9, 18500. [Google Scholar] [CrossRef] [PubMed]
- Everaerts, W.; Sepulvèda, M.R.; Gevaet, T.; Roskams, T.; Nilius, B.; De Ridder, D. Where is the TRPV1 expressed in the bladder, do we see the real channel? Naunyn-Schmiedeberg’s Arch. Pharmacol. 2009, 379, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Marincsák, R.; Tóth, B.I.; Czifra, G.; Márton, I.; Rèdl, P.; Tóth, L.; Kovács, L.; Bíró, T. Increased expression of TRPV1 in squamous cell carcinoma of the human tongue. Oral Dis. 2009, 15, 328–335. [Google Scholar] [CrossRef] [PubMed]
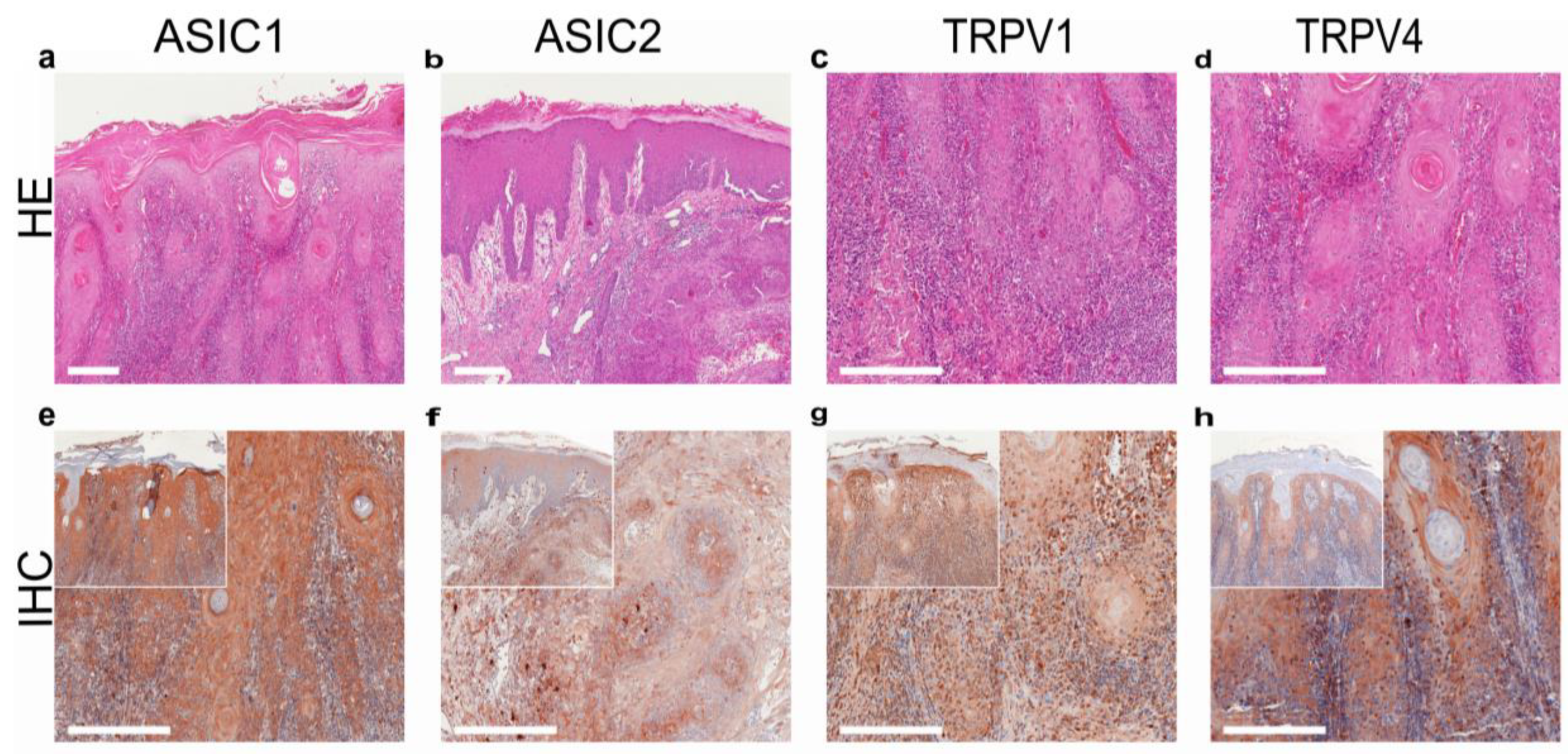
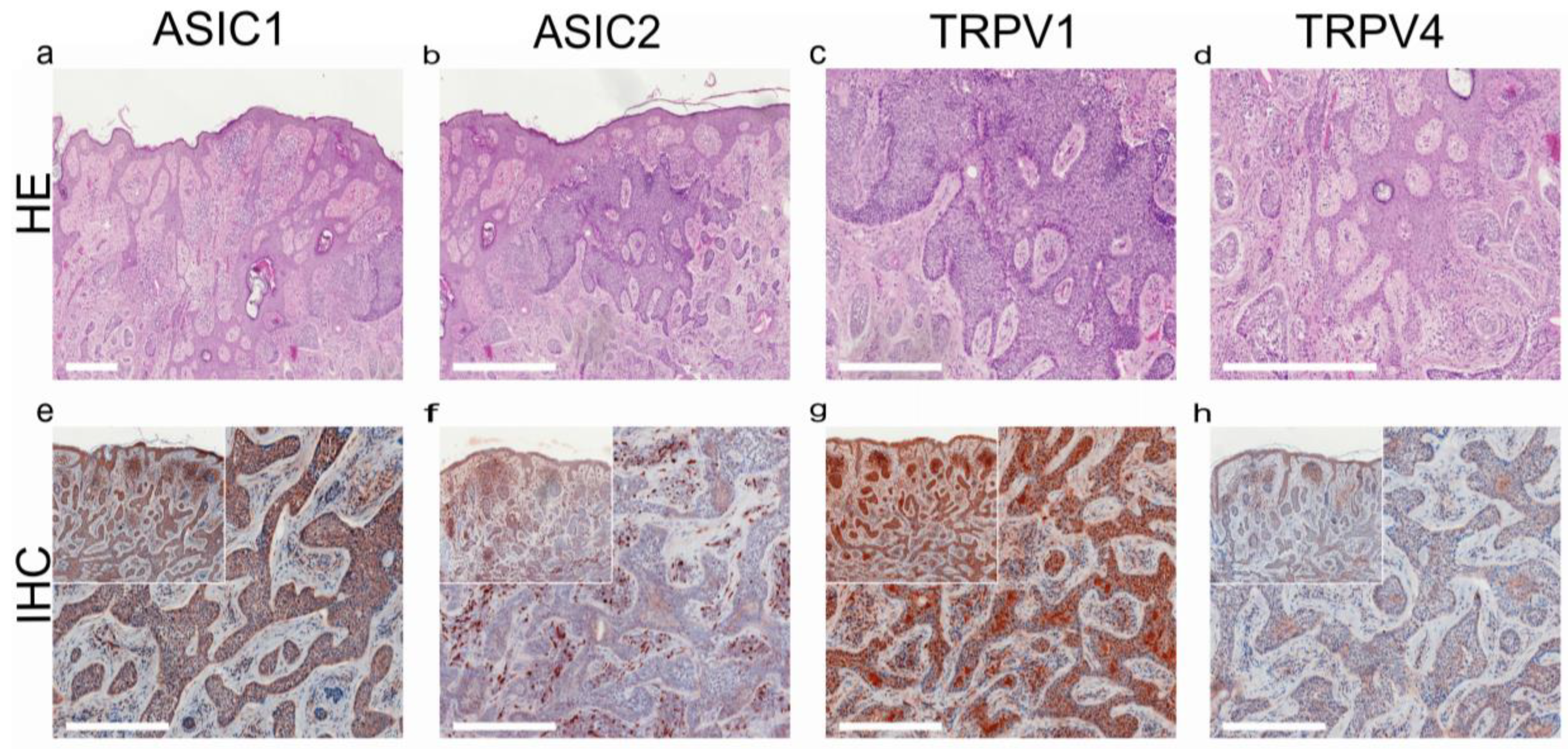
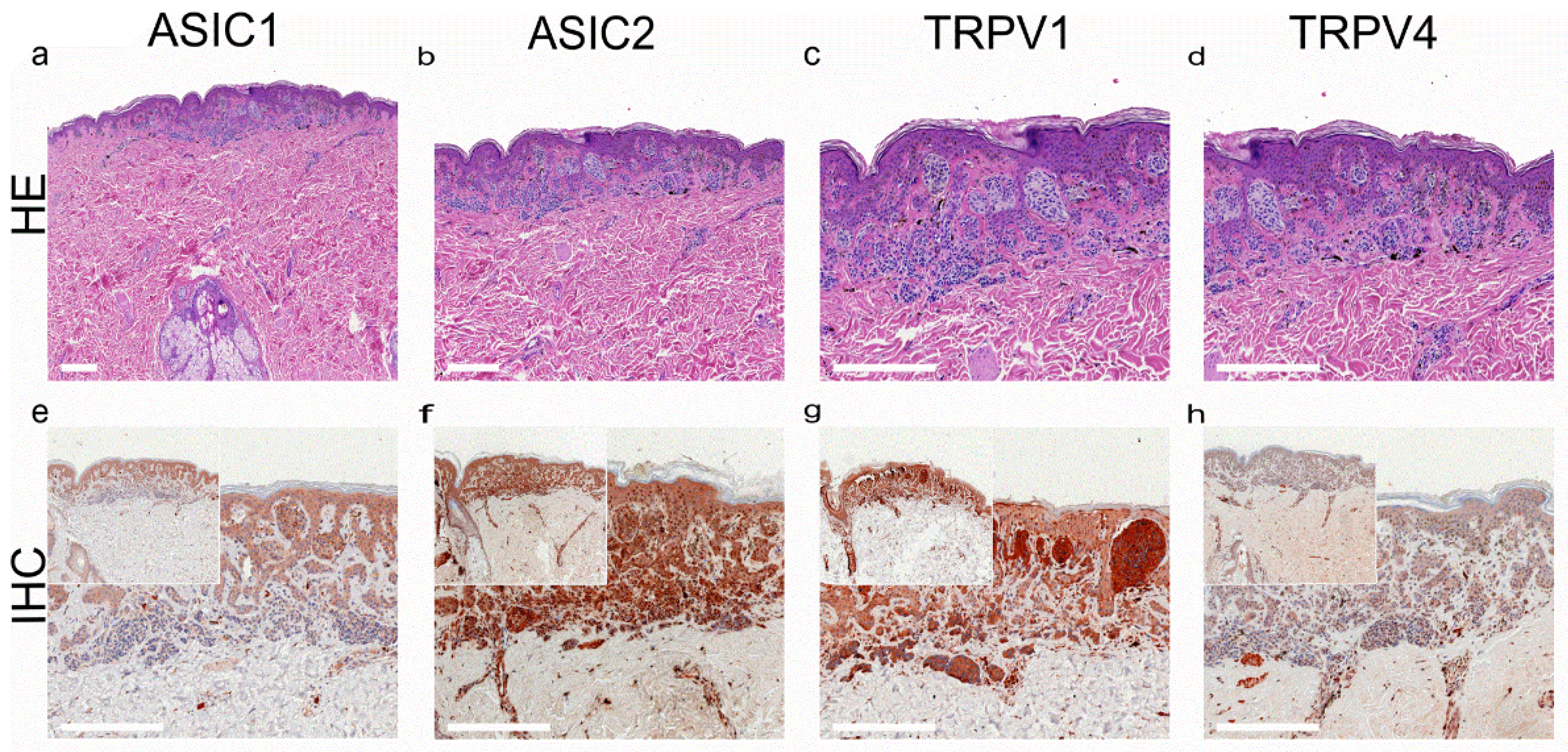
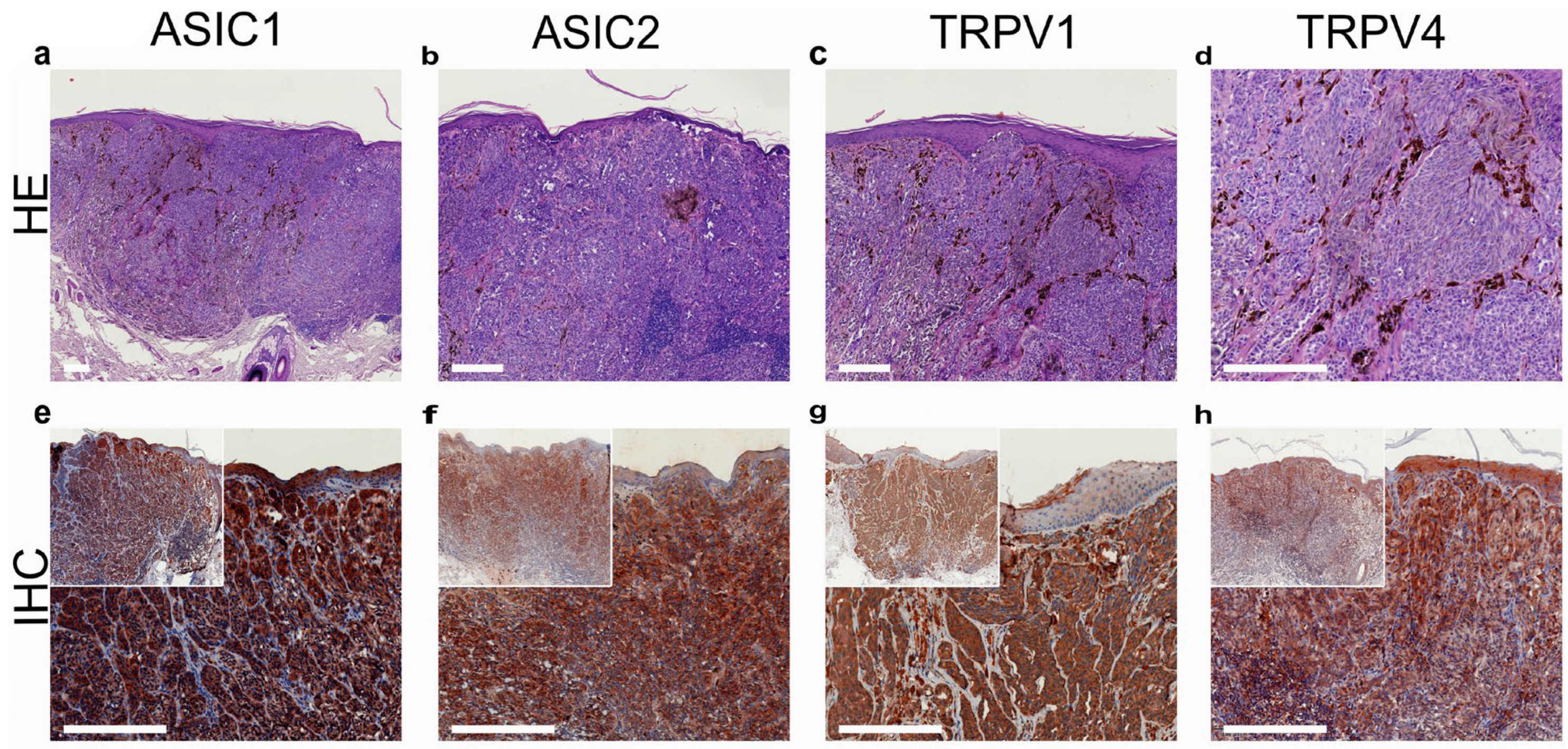
- Ackermann, K.; Wallner, S.; Brochhausen, C.; Schreml, S. Expression profiles of ASIC1/2 and TRPV1/4 in common skin tumors. Int. J. Mol. Sci. 2021, 22, 6024. [Google Scholar] [CrossRef] [PubMed]
- Kiss, F.; Kormos, V.; Szőke, E.; Kecskès, A.; Tóth, N.; Steib, A.; Szallasi, A.; Scheich, B.; Gaszner, B.; Kun, J.; et al. Functional transient receptor potential ankyrin 1 and vanilloid 1 ion channels are overexpressed in human oral squamous cell carcinoma. Int. J. Mol. Sci. 2022, 23, 1921. [Google Scholar] [CrossRef]
- Sakakibara, A.; Sakakibara, S.; Kusumoto, J.; Takeda, D.; Hasegawa, T.; Akashi, M.; MInamikawa, T.; Hashikawa, K.; Terashi, H.; Komori, T. Upregulated expression of transient receptor potential cation channel subfamily V receptors in mucosae of patients with oral squamous cell carcinoma and patients with a history of alcohol consumption and smoking. PLoS ONE 2017, 12, e0169723. [Google Scholar] [CrossRef] [PubMed]
- Fusi, C.; Materazzi, S.; Minocci, D.; Maio, V.; Oranges, T.; Massi, D.; Nassini, R. Transient receptor potential vanilloid 4 (TRPV4) is downregulated in keratinocytes in human non-melanoma skin cancer. J. Investig. Dermatol. 2014, 9, 2408–2417. [Google Scholar] [CrossRef]
- Viet, C.T.; Yu, G.; Asam, K.; Thomas, C.M.; Yoon, A.J.; Wongrorawat, Y.C.; Haghighiabyaneh, M.; Kilkuts, C.A.; McGue, C.M.; Couey, M.A.; et al. The REASON score: An epigenetic and clinicopathologic score to predict risk of poor survival in patients with early stage oral squamous cell carcinoma. Biomark. Res. 2021, 9, 42. [Google Scholar] [CrossRef]
- Okamoto, Y.; Ohkubo, T.; Ikebe, T.; Yamazaki, J. Blockade of TRPM8 activity reduces the invasion potential of oral squamous carcinoma cell lines. Int. J. Oncol. 2012, 40, 1431–1440. [Google Scholar] [PubMed]
- Zhao, L.-Y.; Xu, W.-L.; Xu, Z.-Q.; Qi, C.; Li, Y.; Chen, J.; Liu, L.-K.; Wu, Y.-N.; Gao, J.; Ye, J.-H. The overexpression of functional transient receptor potential chnnel TRPM2 in oral squamous cell carcinoma. Sci. Rep. 2016, 6, 38471. [Google Scholar] [CrossRef]
- Kurz, B.; Michael, H.P.; Förch, A.; Wallner, S.; Zeman, F.; Decking, S.-M.; Ugele, I.; Hintschich, C.; Haubner, F.; Ettl, T.; et al. Expression of pH-sensitive TRPC4 in common skin tumors. Int. J. Mol. Sci. 2023, 24, 1037. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Feng, F.; Luo, W.; Sanders, A.J.; Zhang, Y.; Liang, J.; Chen, C.; Feng, W.; Gu, W.; Liao, W.; et al. Overexpressed transient receptor potential vanilloid 1(TRPV1) in lung adenocarcinoma harbours a new opportunity for therapeutic targeting. Cancer Gene Ther. 2022, 29, 1405–1417. [Google Scholar] [CrossRef] [PubMed]
- Huang, T. TRPV1 is a potential biomarker for the prediction and treatment of multiple cancers based on a pan-cancer analysis. Math. Biosci. Eng. 2022, 19, 8361–8379. [Google Scholar] [CrossRef]
- Gao, R.; Meng, M.; Zhou, X.; Yu, M.; Li, Z.; Li, J.; Wang, X.; Song, Y.; Wang, H.; He, J. TRPV1, a novel biomarker associated with lung cancer via excluding immune infiltration. Med. Commun. 2020, 3, e139. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chen, C.; Xiang, Q.; Fan, S.; Xiao, T.; Chen, Y.; Zheng, D. Transient receptor potential cation channel subfamily V member 1 expression promotes chemoresistance in non-small-cell lung cancer. Front. Oncol. 2022, 12, 773654. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Q.; Fan, K.; Li, B.; Li, H.; Qi, H.; Guo, J.; Cao, Y.; Sun, H. Overexpression of TRPV3 correlates with tumor progression in non-small cell lung cancer. Int. J. Mol. Sci. 2016, 17, 437. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Manamiya, Y.; Watanabe, H.; Takahashi, N.; Ito, M.; Toda, H.; Konno, H.; Mitsui, M.; Motoyama, S.; Ogawa, J.-I. Expression of the transient receptor potential channel C3 correlates with a favorable prognosis in patients with adenocarcinoma of the lung. Ann. Surg. Oncol. 2011, 18, 3377–3383. [Google Scholar] [CrossRef]
- Schaefer, E.A.M.; Stohr, S.; Meister, M.; Aigner, A.; Gudermann, T.; Buech, T.R.H. Stimulation of the chemosensory TRPA1 cation channel by volatile substances promotes cell survival of small cell lung cancer cells. Biochem. Pharmacol. 2013, 85, 426–438. [Google Scholar] [CrossRef]
- Monet, M.; Lehen’kyi, V.; Gackiere, F.; Gackiere, F.; Firlej, V.; Vandenberghe, M.; Roudbaraki, M.; Gkika, D.; Pourtier, A.; Bidaux, G.; et al. Role of cation channel TRPV2 in promoting prostate cancer migration and progression to androgen resistance. Cancer Res. 2010, 70, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.R.; Pinheiro, L.C.L.; Francelino, A.L.; Miqueloto, C.A.; Losi Guembarowski, A.F.M.; de Oliveira, K.B.; Fuganti, P.E.; de Syllos Cólus, I.M.; Losi Guembarowski, R. Tissue immunostaining of candidate prognostic proteins in metastatic and non-metastatic prostate cancer. J. Cancer Res. Clin. Oncol. 2023, 149, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Ferreira De Souza, M.; Kuasne, H.; De Camargo Barros-Filho, M.; Lizotti Ciliao, H.; Albuquerque Marchi, F.; Fuganti, P.E.; Rogatto, S.R.; De Syllos Cólus, I.M. Circulating mRNA signature as a marker for high-risk prostate cancer. Carcinogenesis 2020, 41, 139–145. [Google Scholar] [CrossRef]
- Beck, B.; Bideaux, G.; Bavencoffe, A.; Lemonnier, L.; Thebault, S.; Shuba, Y.; Barrit, G.; Skryma, R.; Prevarskaya, N. Prospects for prostate cancer imaging and therapy using high-affinity TRPM8 activators. Cell Calcium 2007, 41, 285–294. [Google Scholar] [CrossRef]
- Asuthkar, S.; Velpula, K.K.; Elustondo, P.A.; Demirkhanyan, L.; Zakharian, E. TRPM8 channel as a novel molecular target in androgen-regulated prostate cancer cells. Oncotarget 2015, 6, 17221–17236. [Google Scholar] [CrossRef]
- Alaimo, A.; de Felice, D.; Genovedsi, S.; Lorenzoni, M.; Lunardi, A. Tune the channel: TRPM8 targeting in prostate cancer. Oncoscience 2021, 8, 97–100. [Google Scholar] [CrossRef]
- Fixemer, T.; Wissenbach, U.; Flockerzi, V.; Bonkhoff, H. TRPV6 in human prostate cancer: A novel prognostic marker for tumor progression. Oncogene 2003, 22, 7858–7861. [Google Scholar] [CrossRef] [PubMed]
- Sterle, I.; Zupancic, D.; Romih, R. Correlation between urothelial differentiation and sensory proteins P2X3, P2X5, TRPV1, and TRPV4 in normal urothelium and papillary carcinoma of human bladder. Biomed. Res. Int. 2014, 2014, 805236. [Google Scholar] [CrossRef]
- Kalogris, C.; Caprodossi, S.; Amantini, C.; Lambertucci, F.; Nabissi, M.; Morelli, M.B.; Farfariello, V.; Filosa, A.; Emiliozzi, M.; Mammana, G.; et al. Expression of transient receptor potential vanilloid-1 (TRPV1) in urothelial cancers of human bladder: Relation to clinicopathological and molecular parameters. Histopathology 2010, 57, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, M.; Vannucchi, M.G.; Spinelli, M.; Bizzoco, E.; Beneforti, P.; Turini, D.; Faussone-Pellegrini, M.-S. Transient receptor potential vanilloid type 1 (TRPV1) expression changes from normal urothelium to transitional cell carcinoma of human bladder. Eur. Urol. 2005, 48, 691–698. [Google Scholar] [CrossRef]
- Caprodossi, S.; Lucciarini, R.; Amantini, C.; Nabissi, M.; Canesin, G.; Balalrini, P.; Di Spilimbergo, A.; Carderelli, M.A.; Servi, L.; Mammana, G.; et al. Transient receptor potential vanilloid type 2 (TRPV2) expression in normal urothelium and in urothelial carcinoma of human bladder: Correlation with the pathologic stage. Eur. Urol. 2008, 54, 612–620. [Google Scholar] [CrossRef]
- Xiao, N.; Jiang, L.M.; Ge, B.; Zhang, T.Y.; Zhao, X.K.; Zhou, X. Overexpression of TRPM8 is associated with poor prognosis in urothelial carcinoma of the bladder. Tumour Biol. 2014, 35, 11499–11504. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.; Córdova, C.; Marchant, I.; Zúniga, R.; Ochova, P.; Ramírez-Barrantes, R.; González-Arriagada, W.A.; Rodriguez, B.; Olivero, P. Intracellular aggregated TRPV1 is associated with lower survival in breast cancer patients. Breast Cancer 2018, 10, 161–168. [Google Scholar] [CrossRef]
- Elbaz, M.; Ahirwar, D.; Xiaoli, Z.; Zhou, X.; Lustberg, M.; Nasser, M.W.; Shilo, K.; Ganju, R.K. TRPV2 is a novel biomarker and therapeutic target in triple negative breaswt cancer. Oncotarget 2016, 9, 33459–33470. [Google Scholar] [CrossRef] [PubMed]
- Dhennin-Duthille, I.; Gautier, M.; Faouzi, M.; Guilbert, A.; Brevet, M.; Vaudry, D.; Ahidouch, A.; Sevestre, H.; Ouadid-Ahidouch, H. High expression of transient receptor potential channels in human breast cancer epithelial cells and tissues: Correlation with pathological parameters. Cell Physiol. Biochem. 2011, 28, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, N.; Wang, Y.; Yu, J.; Mi, J. Calcium channel TRPV6 promotes breast cancer metastasis by NFATC2IP. Cancer Lett. 2021, 519, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Aydar, E.; Yeo, S.; Djamgoz, M.; Palmer, C. Abnormal expression, localization and interactions of canonical transient receptor potential ion channels in human breast cancer cell lines and tissues: A potential target for breast cancer diagnosis and therapy. Cancer Cell Int. 2009, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Yang, F.; Chen, S.; Wan, H.; Zhao, X.; Dong, H. The role of TRPV1 ion channel in the suppression of gastric cancer development. J. Exp. Clin. Cancer Res. 2020, 39, 206. [Google Scholar] [CrossRef]
- Zoppoli, P.; Calice, S.; Laurino, S.; Ruggieri, V.; La Rocca, F.; La Torre, G.; Ciuffi, M.; Amendola, E.; De Vita, F.; Petrillo, A.; et al. TRPV2 calcium channel gene expression and outcomes in gastric cancer patients: A clinically relevant association. J. Clin. Med. 2019, 11, 662. [Google Scholar] [CrossRef]
- Xu, Q.; Kong, N.; Zhang, J.; Bai, N.; Bi, J.; Li, W. Expression of transient receptor potential cation channel subfamily M member 8 in gastric cancer and its clinical significance. Exp. Ther. Med. 2021, 21, 377. [Google Scholar] [CrossRef]
- Laurino, S.; Mazzone, P.; Ruggieri, V.; Zoppoli, P.; Calice, G.; Lapenta, A.; Ciuffi, M.; Ignomirelli, O.; Vita, G.; Sgambato, A.; et al. Cationic channel TRPV2 overexpression promotes resistance to cisplatin-induced apoptosis in gastric cancer cells. Front. Pharmacol. 2021, 12, 746628. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, B.; Wang, X.; Mao, J.; Li, W.; Sun, Y.; Yuan, Y.; Ben, Q.; Hua, L.; Qian, A. TRPV4 overexpression promotes metastasis through epithelial-mesenchymal transient in gastric cancer and correlates with poor prognosis. OncoTargets Ther. 2020, 13, 8383–8394. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.; He, X.; Yang, Y.; Fu, J.; Zhang, W.; Guo, Z.; Hu, Y.; Liang, L.; Xie, W.; Xiong, H.; et al. TRPV1 induced apoptosis of colorectal cancer cells by activating calcineurin-NFAT2-P53 signaling pathway. Biomed. Res. Int. 2019, 2019, 6712536. [Google Scholar] [CrossRef] [PubMed]
- Sozucan, Y.; Kalender, M.E.; Sari, I.; Suner, A.; Oztuzcu, S.; Arman, K.; Yumrutas, O.; Bozgeyik, I.; Cengiz, B.; Igci, Y.Z.; et al. TRP genes family expression in colorectal cancer. Exp. Oncol. 2015, 37, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Pagano, E.; Romano, B.; Cicia, D.; Ianotti, F.A.; Venneri, T.; Lucariello, G.; Nani, M.F.; Cattaneo, F.; de Cicco, P.; D’Armiento, M.; et al. TRPM8 indicates poor prognosis in colorectal cancer patients and its pharmacological targeting reduces tumour growth in mice by inhibiting Wnt/β-catenin signaling. Br. J. Pharmacol. 2023, 180, 235–251. [Google Scholar] [CrossRef]
- Yee, N.S.; Zhou, W.M.; Lee, M. Transient receptor potential channel TRPM8 is over-expressed and required for cellular proliferation in pancreatic adenocarcinoma. Cancer Lett. 2010, 297, 49–55. [Google Scholar] [CrossRef]
- Yee, N.S.; Brown, M.S.; Lee, M.; Zhou, W.; Jensen, C.; Gerke, H.; Yee, R.K. TRPM8 ion channel is aberrantly expressed and required for preventing replicative senescence in pancreatic adenocarcinoma: Potential role for TRPM8 as a biomarker and target. Cancer Biol. Ther. 2012, 13, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Yee, N.S.; Chan, A.S.; Yee, J.D.; Yee, R.K. TRPM7 and TRPM8 ion channels in pancreatic adenocarcinoma: Potential roles as cancer biomarkers and targets. Scientifica 2012, 2012, 415158. [Google Scholar] [CrossRef]
- Chelaru, N.-R.; Chiosa, A.; Sorop, A.; Spiridon, A.; Cojocaru, F.; Domocos, D.; Cucu, D.; Popescu, I.; Dima, S.-O. The association between TRP channel expression and clinicopathological characteristics of patients with pancreatic adenocarcinoma. Int. J. Mol. Sci. 2022, 23, 9045. [Google Scholar] [CrossRef]
- Du, J.-D.; Zheng, X.; Chen, Y.-L.; Huang, Z.-Q.; Cai, S.-W.; Jiao, H.-B.; Zhu, Z.-M.; Hu, B. Elevated transient receptor potential melastatin 8 (TRPM8) expression is correlated with poor prognosis in pancreatic cancer. Med. Sci. Monit. 2018, 24, 3720–3725. [Google Scholar] [CrossRef]
- Cojocaru, F.; Selescu, T.; Domocos, D.; Marutescu, L.; Chritoiu, G.; Chelaru, N.-R.; Dima, S.; Mihailescu, D.; Babes, A.; Cucu, D. Functional expression of the transient receptor potential ankytin type 1 channel in pancreatic adenocarcinoma cells. Sci. Rep. 2021, 11, 2018. [Google Scholar] [CrossRef]
- Van den Eynde, C.; de Clerq, K.; van Bree, R.; Luyten, K.; Annibali, D.; Amant, F.; Han, S.; Van Nieuwenhuysen, E.; Baert, T.; Peeraer, K.; et al. TRP channel expression correlates with the epithelial-mesenchymal transition and high-risk endometrial carcinoma. Cell Mol. Life Sci. 2021, 79, 26. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, O.; Morelli, M.B.; Annibali, D.; Aguzzi, C.; Zeppa, L.; Tuyaerts, S.; Amantini, C.; Amant, F.; Ferretti, B.; Maggi, F.; et al. The effects of cannabidiol and prognostic role of TRPV2 in human endometrial cancer. Int. J. Mol. Sci. 2020, 21, 5409. [Google Scholar] [CrossRef] [PubMed]
- Han, G.H.; Chay, D.B.; Nam, S.; Cho, H.; Chung, J.-Y.; Kim, J.-H. Prognostic significance of transient receptor potential vanilloid type 1 (TRPV1) and phosphatase and tension homolog (PTEN) in epithelian ovarian cancer. Cancer Genom. Proteom. 2020, 17, 309–319. [Google Scholar] [CrossRef]
- Wang, K.; Feng, X.; Zheng, L.; Chai, Z.; Yu, J.; You, X.; Li, X.; Cheng, X. TRPV4 is a prognostic biomarker that correlates with the immunosuppressive microenvironment and chemoresistance of anti-cancer drugs. Front. Mol. Biosci. 2021, 8, 690500. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Z.; Zhao, H.; Qiu, M.; Wen, Y.; Yao, X.; Tang, W.H. Identification of TRPM2 as a marker associated with poor prognosis and immune infiltration in kidney renal clear cell carcinoma. Front. Mol. Biosci. 2022, 8, 774905. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Liu, X.-Y.; Zhuo, D.-X.; Huang, H.-B.; Zhang, F.-B.; Liao, S.-F. Decreased expression of TRPV1 in renal cell carcinoma: Association with tumor Fuhrman grades and histopathological subtypes. Cancer Manag. Res. 2018, 10, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Liu, G.; Xu, X.; Xie, C.; Sun, F.; Yang, Y.; Zhang, T.; Hua, S.; Fan, W.; Li, Q.; et al. High expression of vanilloid receptor-1 is associated with better prognosis of patients with hepatocellular carcinoma. Cancer Genet. Cytogenet. 2008, 186, 25–32. [Google Scholar] [CrossRef]
- Liu, G.; Xie, C.; Sun, F.; Xu, X.; Yang, Y.; Zhang, T.; Deng, Y.; Wang, D.; Huang, Z.; Yang, L.; et al. Clinical significance of transient receptor potential vanilloid 2 expression in human hepatocellular carcinoma. Cancer Genet. Cytogenet. 2010, 197, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Liu, G.; Xie, C.; Qian, K.; Lei, X.; Liu, Q.; Liu, G.; Cao, Z.; Fu, J.; Du, H.; et al. Pharmacological inhibition of TRPV4 channel suppresses malignant biological behavior of hepatocellular carcinoma via modulation of ERK signaling pathway. Biomed. Pharmacother. 2018, 101, 910–919. [Google Scholar] [CrossRef]
- Oancea, E.; Vriens, J.; Brauchi, S.; Jun, J.; Splawski, I.; Clapham, D.E. TRPM1 forms ion channels associated with melanin content in melanocytes. Sci. Signal. 2009, 2, ra21. [Google Scholar] [CrossRef]
- Duncan, L.M.; Deeds, J.; Cronin, F.E.; Donovan, M.; Sober, A.J.; Kauffman, M.; McCarthy, J.J. Melastatin expression and prognosis in cutaneous malignant melanoma. J. Clin. Oncol. 2001, 19, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.; Guo, H.; Yang, S.-E.; Cornelius, L.; Linette, G.; Murphy, M.; Sheehan, C.; Ross, J.; Slominski, A.; Carlson, J.A. TRPM1 (melastatin) expression is an independent predictor of overall survival in clinical AJCC stage I and II melanoma patients. J. Cutan. Pathol. 2017, 44, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Hammock, L.; Cohen, C.; Carlson, G.; Murray, D.; Ross, J.S.; Sheehan, C.; Nazir, T.M.; Carlson, J.A. Chromogenic in situ hydridization analysis of melastatin mRNA expression in melanomas from American Joint Committee on Cancer stage I and II patients with recurrent melanoma. J. Cutan. Pathol. 2006, 33, 599–607. [Google Scholar] [CrossRef]
- Erickson, L.A.; Letts, G.A.; Shah, S.M.; Shackelton, J.B.; Duncan, L.M. TRPM1 (melastatin-1/MLSN1) mRNA expression in Spitz nevi and nodular melanomas. Mod. Pathol. 2009, 22, 969–976. [Google Scholar] [CrossRef]
- Dhingra, A.; Fina, M.E.; Neinstein, A.; Ramsey, D.J.; Xu, Y.; Fishman, G.A.; Alexander, K.R.; Qian, H.; Peachey, N.S.; Gregg, R.G.; et al. Autoantibodies in melanoma-associated retinopathy target TRPM1 cation channels in retinal ON bipolar cells. J. Neurosci. 2011, 31, 3962–3967. [Google Scholar] [CrossRef]
- Yepes, S.; Tucker, M.A.; Koka, H.; Xiao, Y.; Zhang, T.; Jones, K.; Vogt, A.; Burdette, L.; Luo, W.; Zhu, B.; et al. Integrated analysis of coexpression and exome sequencing to prioritize susceptibility genes for familial cutaneous melanoma. J. Investig. Dermatol. 2022, 142, 2464–2475. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Ugawa, S.; Ueda, T.; Morita, A.; Shimada, S. TRPM8 activation suppresses cellular viability in human melanoma. Am. J. Physiol. Cell Physiol. 2008, 295, C296–C301. [Google Scholar] [CrossRef]
- Wang, J.; Qiao, S.; Liang, S.; Qian, C.; Dong, Y.; Pei, M.; Wang, H.; Wan, G. TRPM4 and TRPV2 are two novel prognostic biomarkers and promising targeted therapy in UVM. Front. Mol. Biosci. 2022, 9, 985434. [Google Scholar] [CrossRef]
- Alptekin, M.; Eroglu, S.; Tutar, E.; Sencan, S.; Geyik, M.A.; Ulasli, M.; Demiryurek, A.T.; Camci, C. Gene expression profiling of TRP channels in glioblastoma multiforme and relation with survival. Tumour Biol. 2015, 36, 9209–9213. [Google Scholar] [CrossRef]
- Moutafidi, A.; Gatzounis, G.; Zolota, V.; Assimakopoulou, M. Heat shock factor 1 in brain tumors: A link with transient receptor potential channels TRPV1 and TRPA1. J. Mol. Histol. 2021, 52, 1233–1244. [Google Scholar] [CrossRef]
- Stock, K.; Kumar, J.; Synowitz, M.; Petrosino, S.; Imperatore, R.; Smith, E.S.J.; Wend, P.; Purfürst, B.; Nuber, U.A.; Gurok, A.; et al. Neurol precursor cells induce cell death of high-grade astrocytomas via stimulation of TRPV1. Nat. Med. 2012, 18, 1232–1238. [Google Scholar] [CrossRef]
- Morelli, M.B.; Liberati, S.; Amantini, C.; Nabissi, M.; Santoni, M.; Farfariello, V.; Santoni, G. Expression and function of the transient receptor potential ion channel family in the hematological malignancies. Curr. Mol. Pharmacol. 2013, 6, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Morelli, M.B.; Nabissi, M.; Marinelli, O.; Zeppa, L.; Aguzzi, C.; Santoni, G.; Amantini, C. Transient receptor potential (TRP) channels in haematological malignancies: An update. Biomolecules 2021, 11, 765. [Google Scholar] [CrossRef]
- Meng, L.; Gu, G.; Bi, L. Transient receptor potential channels in multiple myeloma. Oncol. Lett. 2022, 23, 108. [Google Scholar] [CrossRef]
- Bödding, M. TRP proteins and cancer. Cell Signal. 2007, 19, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Gkika, D.; Prevarskaya, N. Molecular mechanisms of TRP regulation in tumor growth and metastasis. Biochim. Biophys. Acta 2009, 1793, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Lehen’kyi, V.; Prevarskaya, N. Study of TRP channels in cancer cells. In TRP Channels; Zhu, M.X., Ed.; CRC Press, Taylor & Francis: Boca Raton, FL, USA, 2011; Chapter 17. [Google Scholar]
- Lehen’kyi, V.; Prevarskaya, N. Oncogenic TRP channels. Adv. Exp. Med. Biol. 2011, 704, 929–945. [Google Scholar]
- Perna, A.; Sellitto, C.; Komici, K.; Hay, E.; Rocca, A.; De Blasiis, P.; Lucariello, A.; Moccia, F.; Guerra, G. Transient receptor potential (TRP) channels in tumor vascularization. Int. J. Mol. Sci. 2022, 23, 14253. [Google Scholar] [CrossRef]
- Van den Eynde, C.; de Clercq, K.; Vriens, J. Transient receptor potential channels in the epithelial-to-mesenchymal transition. Int. J. Mol. Sci. 2021, 22, 8188. [Google Scholar] [CrossRef]
- Yang, D.; Kim, J. Emerging role of transient receptor potential (TRP) channels in cancer progression. BMB Rep. 2020, 53, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Wang, X.; Zhao, D.; Liu, H.; Hu, Y. Calcium homeostasis and cancer: Insights from endoplasmic reticulum-centered organelle communications. Trends Cell Biol. 2023, 33, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Zong, G.F.; Deng, R.; Yu, S.F.; Wang, A.Y.; Wei, Z.H.; Zhao, Y.; Lu, Y. Thermo-transient receptor potential channels: Therapeutic potential in gastric cancer. Int. J. Mol. Sci. 2022, 23, 15289. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.L.; Noda, M.; Yoshioka, T.; Ito, E. A novel strategy for treating cancer: Understanding the role of Ca2+ signaling from nociceptive TRP channels in regulating cancer progression. Explor. Target. Anti-Tumor Ther. 2021, 2, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Kaji, I.; Yasuoka, Y.; Karaki, S.I.; Kuwahara, A. Activation of TRPA1 by luminal stimuli induces EP4-mediated anion secretion in human and rat colon. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 302, G690–G701. [Google Scholar] [CrossRef]
- Mihara, H.; Uchida, K.; Watanabe, Y.; Nanjo, S.; Sakumura, M.; Motoo, I.; Ando, T.; Minemura, M.; Muhammad, J.S.; Yamamoto, H.; et al. Colonic TRPV4 overexpression is related to constipation severity. BMC Gastroenterol. 2023, 23, 13. [Google Scholar] [CrossRef]
- Holzer, P. Transient receptor potential (TRP) channels as drug targets for diseases of the digestive system. Pharmacol. Rev. 2011, 131, 142–170. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Kubota, K.; Ohbuchi, K.; Kaneko, A.; Ohno, N.; Mase, A.; Matsushima, H.; Yamamoto, M.; Miyano, K.; Uezono, Y.; et al. Transient receptor potential ankyrin 1 agonists improve intestinal transit in a murine model of postoperative ileus. Neurogastroenterol. Motil. 2016, 28, 1792–1805. [Google Scholar] [CrossRef]
- Buck, S.H.; Burks, T.F. The neuropharmacology of capsaicin: Review of some recent observations. Pharmacol. Rev. 1986, 38, 179–226. [Google Scholar]
- Lazzeri, M.; Vannucchi, M.G.; Zardo, C.; Spinelli, M.; Beneforti, P.; Turini, D.; Faussone-Pellegrini, M.S. Immunohistochemical evidence of vanilloid receptor 1 in normal human urinary bladder. Eur. Urol. 2004, 46, 792–798. [Google Scholar] [CrossRef]
- Hernández-Ortego, P.; Torres-Montero, R.; de la Pena, E.; Viana, F.; Fernández-Trillo, J. Validation of six commercial antibodies for detection of heterologous and endogenous TRPM8 ion channel expression. Int. J. Mol. Sci. 2022, 23, 16164. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor | Increased | Decreased |
|---|---|---|
| Oral SQCC | TRPV1, TRPA1, TRM2 | |
| BCC | TRPV1 | |
| Prostate cancer | TRPM8, TRPV2 | |
| Urothelial carcinoma | TRPV2, TRPM8 | TRPV1 |
| Breast carcinoma | TRM8, TRPV6, TRPC3, TRPC6 | |
| Gastric adenocarcinoma | TRPV1 | |
| Colorectal adenocarcinoma | TRPV1, TRPV3, TRPV4 | |
| Pancreas adenocarcinoma | TRPM7, TRPM8 | |
| Endometrial carcinoma | TRPV2 | |
| Ovarian carcinoma | TRPV1 | |
| Renal cell carcinoma | TRPM2 | TRPV1 |
| Hepatocellular carcinoma | TRPV1, TRPV2 | |
| SQCC, Squamous cell carcinoma | ||
| BCC, Basal cell carcinoma |
| Tumor | Favorable | Adverse |
|---|---|---|
| Oral SQCC | TRPA1 | |
| Lung adenocarcinoma | TRPC3 | TRPV1, TRPV3 |
| Lung small cell carcinoma | TRPA1 | |
| Prostate cancer | TRPV2, TRPV6, TRPM8 | |
| Urothelial carcinoma | TRPM8 | |
| Breast cancer | TRPV2 | |
| Gastric adenocarcinoma | TRPV2, TRPV4, TRPM8 | |
| Pancreatic adenocarcinoma | TRPM8 | |
| Endometrial adenocarcinoma | TRPV2 | |
| Ovarian carcinoma | TRPV1 | |
| Renal cell carcinoma | TRPM2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szallasi, A. “ThermoTRP” Channel Expression in Cancers: Implications for Diagnosis and Prognosis (Practical Approach by a Pathologist). Int. J. Mol. Sci. 2023, 24, 9098. https://doi.org/10.3390/ijms24109098
Szallasi A. “ThermoTRP” Channel Expression in Cancers: Implications for Diagnosis and Prognosis (Practical Approach by a Pathologist). International Journal of Molecular Sciences. 2023; 24(10):9098. https://doi.org/10.3390/ijms24109098
Chicago/Turabian StyleSzallasi, Arpad. 2023. "“ThermoTRP” Channel Expression in Cancers: Implications for Diagnosis and Prognosis (Practical Approach by a Pathologist)" International Journal of Molecular Sciences 24, no. 10: 9098. https://doi.org/10.3390/ijms24109098
APA StyleSzallasi, A. (2023). “ThermoTRP” Channel Expression in Cancers: Implications for Diagnosis and Prognosis (Practical Approach by a Pathologist). International Journal of Molecular Sciences, 24(10), 9098. https://doi.org/10.3390/ijms24109098