
Further Insights into the Gut Microbiota of Cow’s Milk Allergic Infants: Analysis of Microbial Functionality and Its Correlation with Three Fecal Biomarkers

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
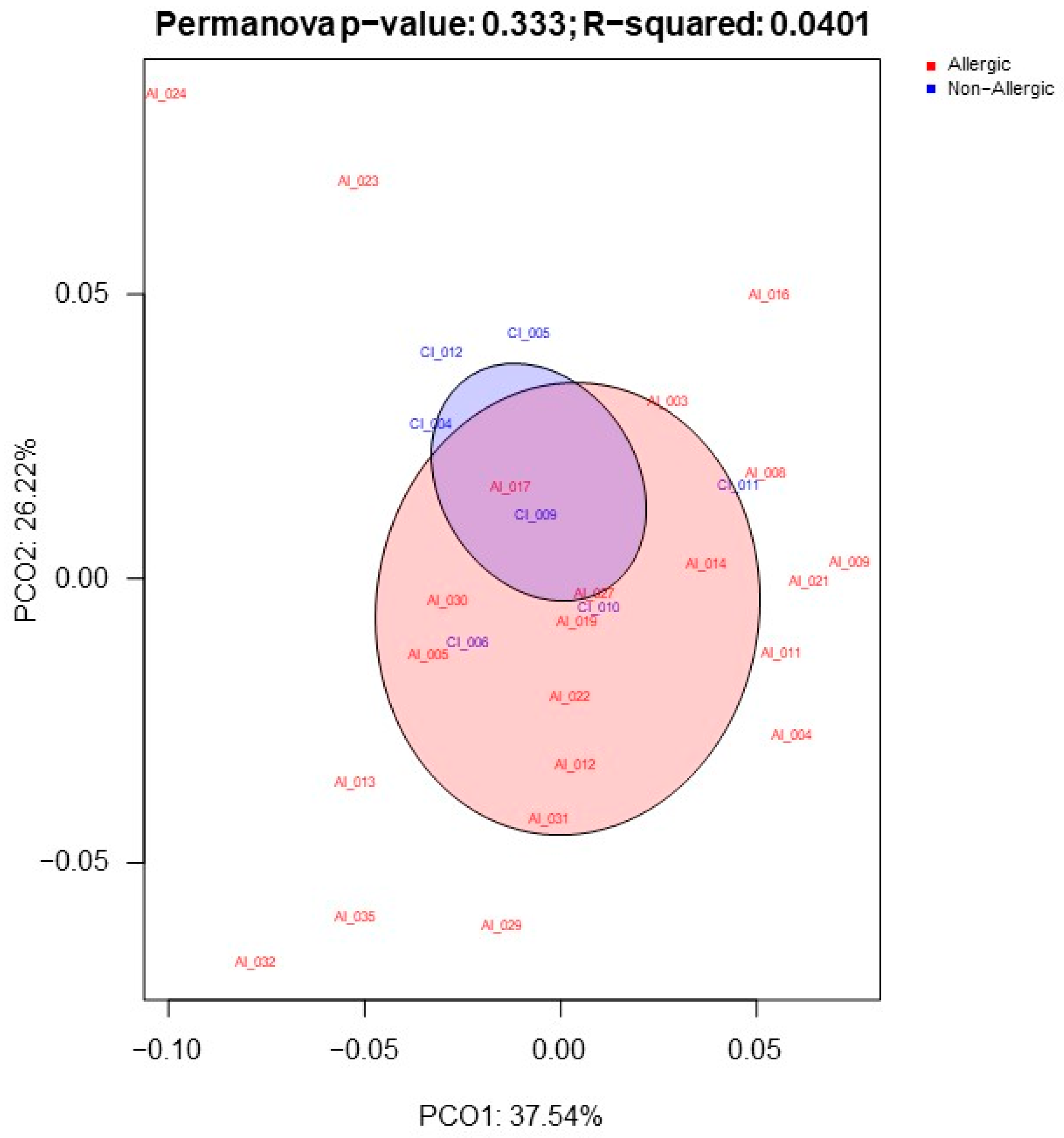
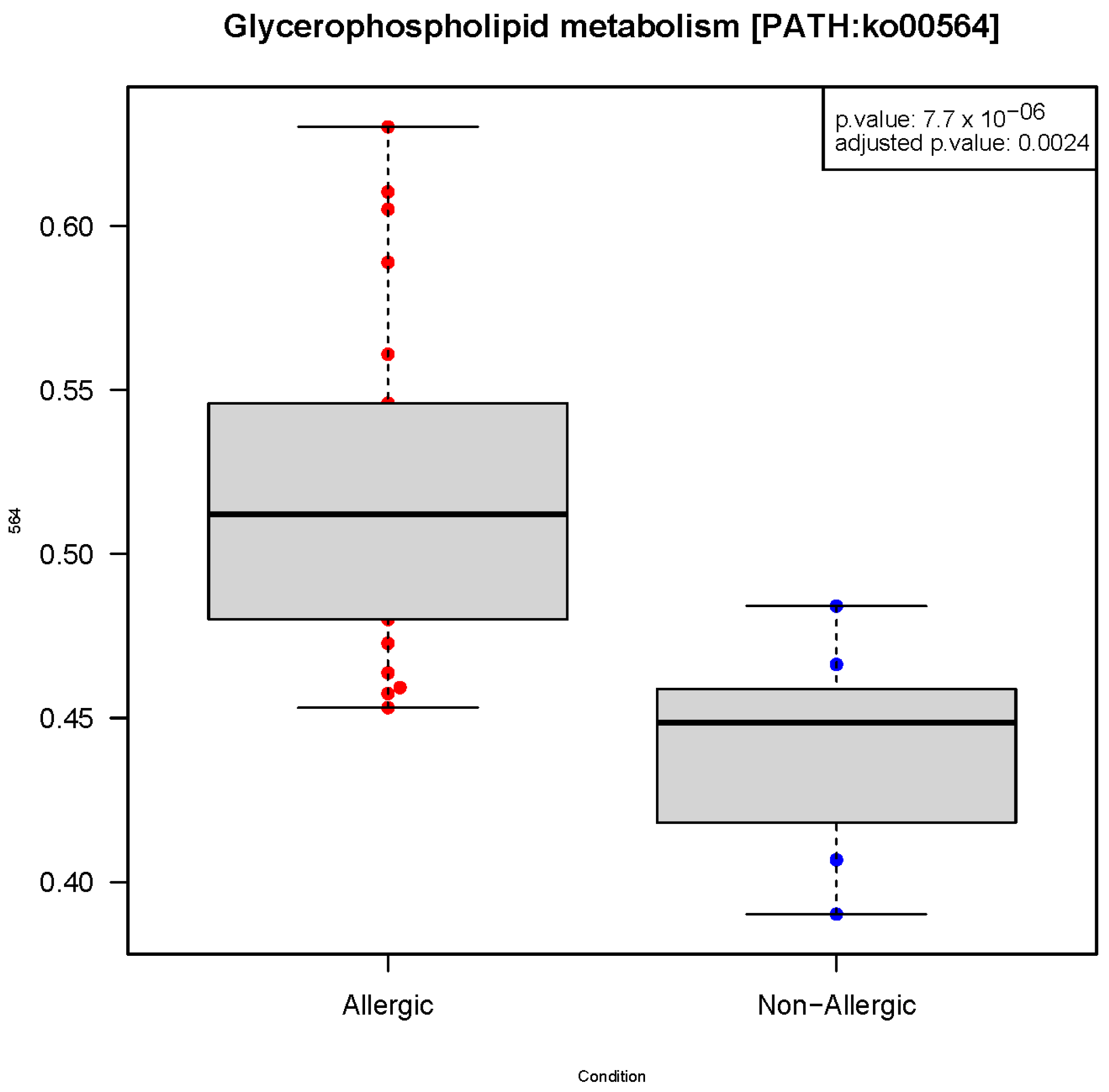
2.1. Gut Microbiota Functionality Profile Associated with CMA
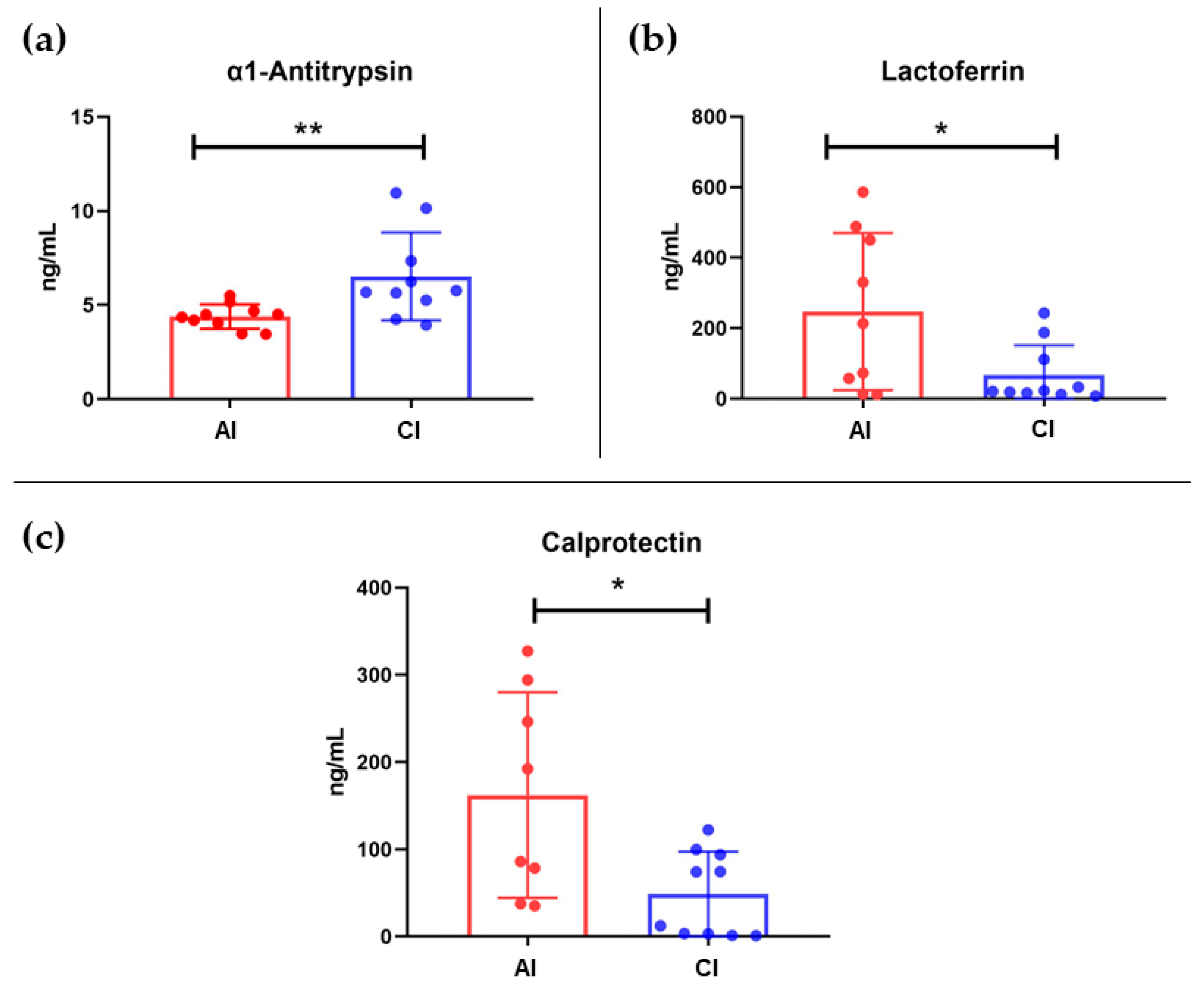
2.2. Fecal Protein Immunodetection
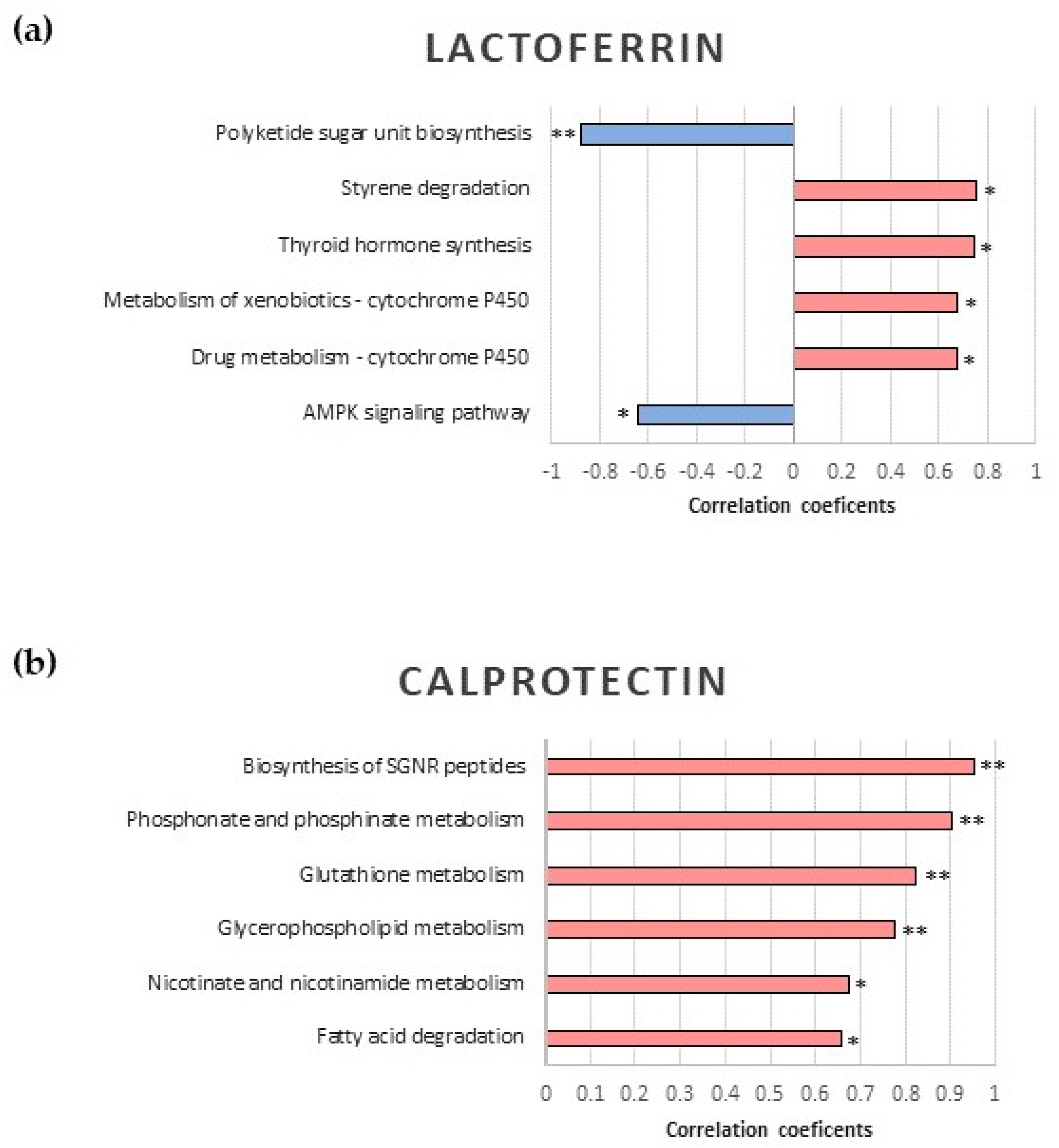
2.3. Correlation Analysis between Fecal Proteins and Microbiota Functionality
3. Discussion
4. Materials and Methods
4.1. Study Design and Population
4.2. Inclusion and Exclusion Criteria
4.3. Sample Collection and Processing
4.4. Shotgun Metagenomics
4.4.1. Sample Preparation
4.4.2. Shotgun Sequencing
4.4.3. Bioinformatics and Statistical Analysis
4.5. Fecal Protein Immunodetection (ELISA)
4.5.1. Sample Preparation
4.5.2. ELISA Protocol
4.5.3. Data Processing and Statistical Analysis
4.6. Correlation Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Power, S.E.; O’Toole, P.W.; Stanton, C.; Ross, R.P.; Fitzgerald, G.F. Intestinal Microbiota, Diet and Health. Br. J. Nutr. 2014, 111, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Sartor, R. Microbial Influences in Inflammatory Bowel Diseases. Gastroenterology 2008, 134, 577–594. [Google Scholar] [CrossRef] [PubMed]
- Burz, S.D.; Abraham, A.-L.; Fonseca, F.; David, O.; Chapron, A.; Béguet-Crespel, F.; Cénard, S.; le Roux, K.; Patrascu, O.; Levenez, F.; et al. A Guide for Ex Vivo Handling and Storage of Stool Samples Intended for Fecal Microbiota Transplantation. Sci. Rep. 2019, 9, 8897. [Google Scholar] [CrossRef] [PubMed]
- Zubeldia-Varela, E.; Barber, D.; Barbas, C.; Perez-Gordo, M.; Rojo, D. Sample Pre-Treatment Procedures for the Omics Analysis of Human Gut Microbiota: Turning Points, Tips and Tricks for Gene Sequencing and Metabolomics. J. Pharm. Biomed Anal. 2020, 191, 113592. [Google Scholar] [CrossRef]
- Zubeldia-Varela, E.; Raczkowska, B.A.; Ferrer, M.; Perez-Gordo, M.; Rojo, D. Techniques for Phenotyping the Gut Microbiota Metabolome. In Microbiome and Metabolome in Diagnosis, Therapy, and Other Strategic Applications; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–41. [Google Scholar]
- Gomez de Agüero, M.; Ganal-Vonarburg, S.C.; Fuhrer, T.; Rupp, S.; Uchimura, Y.; Li, H.; Steinert, A.; Heikenwalder, M.; Hapfelmeier, S.; Sauer, U.; et al. The Maternal Microbiota Drives Early Postnatal Innate Immune Development. Science 2016, 351, 1296–1302. [Google Scholar] [CrossRef]
- Liu, L.; Guo, Q.; Cui, M.; Liu, J.; Yang, C.; Li, X.; Liu, P.; Wang, L. Impact of Maternal Nutrition during Early Pregnancy and Diet during Lactation on Lactoferrin in Mature Breast Milk. Nutrition 2022, 93, 111500. [Google Scholar] [CrossRef]
- Wopereis, H.; Oozeer, R.; Knipping, K.; Belzer, C.; Knol, J. The First Thousand Days—Intestinal Microbiology of Early Life: Establishing a Symbiosis. Pediatr. Allergy Immunol. 2014, 25, 428–438. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Palacio, S.D.; Montes, S.A.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, S.; Sanchez-Carrillo, S.; Ciordia, S.; Mena, M.C.; Méndez-García, C.; Rojo, D.; Bargiela, R.; Zubeldia-Varela, E.; Martínez-Martínez, M.; Barbas, C.; et al. Functional Microbiome Deficits Associated with Ageing: Chronological Age Threshold. Aging Cell 2020, 19, e13063. [Google Scholar] [CrossRef]
- Furness, J.B.; Kunze, W.A.; Clerc, N. Nutrient Tasting and Signaling Mechanisms in the Gut. II. The Intestine as a Sensory Organ: Neural, Endocrine, and Immune Responses. Am. J. Physiol. 1999, 277, G922–G928. [Google Scholar] [CrossRef]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the Immune System. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef]
- Pascal, M.; Perez-Gordo, M.; Caballero, T.; Escribese, M.M.; Lopez Longo, M.N.; Luengo, O.; Manso, L.; Matheu, V.; Seoane, E.; Zamorano, M.; et al. Microbiome and Allergic Diseases. Front. Immunol. 2018, 9, 1584. [Google Scholar] [CrossRef]
- Zubeldia-Varela, E.; Barker-Tejeda, T.C.; Obeso, D.; Villaseñor, A.; Barber, D.; Pérez-Gordo, M. Microbiome and Allergy: New Insights and Perspectives. J. Investig. Allergol. Clin. Immunol. 2022, 32, 327–344. [Google Scholar] [CrossRef]
- Yepes-Nuñez, J.J.; Fiocchi, A.; Pawankar, R.; Cuello-Garcia, C.A.; Zhang, Y.; Morgano, G.P.; Ahn, K.; Al-Hammadi, S.; Agarwal, A.; Gandhi, S.; et al. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Vitamin D. World Allergy Organ. J. 2016, 9, 17. [Google Scholar] [CrossRef]
- Ivanov, I.I.; de Llanos Frutos, R.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific Microbiota Direct the Differentiation of IL-17-Producing T-Helper Cells in the Mucosa of the Small Intestine. Cell Host Microbe 2008, 4, 337–349. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, C.; Scully, P.; O’Mahony, D.; Murphy, S.; O’Brien, F.; Lyons, A.; Sherlock, G.; MacSharry, J.; Kiely, B.; Shanahan, F.; et al. Commensal-Induced Regulatory T Cells Mediate Protection against Pathogen-Stimulated NF-ΚB Activation. PLoS Pathog. 2008, 4, e1000112. [Google Scholar] [CrossRef]
- Huang, Y.J.; Marsland, B.J.; Bunyavanich, S.; O’Mahony, L.; Leung, D.Y.M.; Muraro, A.; Fleisher, T.A. The Microbiome in Allergic Disease: Current Understanding and Future Opportunities-2017 PRACTALL Document of the American Academy of Allergy, Asthma & Immunology and the European Academy of Allergy and Clinical Immunology. J. Allergy Clin. Immunol. 2017, 139, 1099–1110. [Google Scholar] [CrossRef]
- Mera-Berriatua, L.; Zubeldia-Varela, E.; Martín-Antoniano, I.A.; López de Maturana, E.; Rojo, D.; Bazire, R.; Cabrera-Freitag, P.; Barker-Tejeda, T.C.; Ubeda, C.; Barber, D.; et al. Unravelling the Gut Microbiota of Cow’s Milk-Allergic Infants, Their Mothers, and Their Grandmothers. J. Investig. Allergol. Clin. Immunol. 2022, 32, 395–398. [Google Scholar] [CrossRef]
- Su, Y.-J.; Luo, S.-D.; Hsu, C.-Y.; Kuo, H.-C. Differences in Gut Microbiota between Allergic Rhinitis, Atopic Dermatitis, and Skin Urticaria: A Pilot Study. Medicine 2021, 100, e25091. [Google Scholar] [CrossRef]
- Delgado Dolset, M.I.; Obeso, D.; Rodriguez-Coira, J.; Villaseñor, A.; González Cuervo, H.; Arjona, A.; Barbas, C.; Barber, D.; Carrillo, T.; Escribese, M.M. Contribution of Allergy in the Acquisition of Uncontrolled Severe Asthma. Front. Med. 2022, 9, 1009324. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.F.; Ong, R.Y.; Common, J.E. Skin Microbiome of Atopic Dermatitis. Allergol. Int. 2022, 71, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, T.J. An Introduction to the Analysis of Shotgun Metagenomic Data. Front. Plant Sci. 2014, 5, 209. [Google Scholar] [CrossRef]
- Abril, A.G.; Carrera, M.; Sánchez-Pérez, Á.; Villa, T.G. Gut Microbiome Proteomics in Food Allergies. Int. J. Mol. Sci. 2023, 24, 2234. [Google Scholar] [CrossRef]
- Wang, W.-L.; Xu, S.-Y.; Ren, Z.-G.; Tao, L.; Jiang, J.-W.; Zheng, S.-S. Application of Metagenomics in the Human Gut Microbiome. World J. Gastroenterol. 2015, 21, 803–814. [Google Scholar] [CrossRef]
- Calvani, M.; Anania, C.; Cuomo, B.; D’auria, E.; Decimo, F.; Indirli, G.C.; Marseglia, G.; Mastrorilli, V.; Sartorio, M.U.A.; Santoro, A.; et al. Non-IgE- or Mixed IgE/Non-IgE-Mediated Gastrointestinal Food Allergies in the First Years of Life: Old and New Tools for Diagnosis. Nutrients 2021, 13, 226. [Google Scholar] [CrossRef]
- Abraham, B.P.; Kane, S. Fecal Markers: Calprotectin and Lactoferrin. Gastroenterol. Clin. North Am. 2012, 41, 483–495. [Google Scholar] [CrossRef]
- Langhorst, J.; Elsenbruch, S.; Koelzer, J.; Rueffer, A.; Michalsen, A.; Dobos, G.J. Noninvasive Markers in the Assessment of Intestinal Inflammation in Inflammatory Bowel Diseases: Performance of Fecal Lactoferrin, Calprotectin, and PMN-Elastase, CRP, and Clinical Indices. Am. J. Gastroenterol. 2008, 103, 162–169. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, J.; Ren, F.; Shen, L.; Li, F. Can Fecal Calprotectin Levels Be Used to Monitor Infant Milk Protein Allergies? Allergy Asthma Clin. Immunol. 2021, 17, 132. [Google Scholar] [CrossRef]
- Ataee, P.; Zoghali, M.; Nikkhoo, B.; Ghaderi, E.; Mansouri, M.; Nasiri, R.; Eftekhari, K. Diagnostic Value of Fecal Calprotectin in Response to Mother’s Diet in Breast-Fed Infants with Cow’s Milk Allergy Colitis. Iran. J. Pediatr. 2018, 28, e66172. [Google Scholar] [CrossRef]
- Singer, A.A.M. Food Protein-Induced Enterocolitis and Alpha-1-Antitrypsin Deficiency. J. Allergy Clin. Immunol. 2020, 145, 444. [Google Scholar] [CrossRef] [PubMed]
- Carrell, R.W.; Jeppsson, J.O.; Laurell, C.B.; Brennan, S.O.; Owen, M.C.; Vaughan, L.; Boswell, D.R. Structure and Variation of Human Alpha 1-Antitrypsin. Nature 1982, 298, 329–334. [Google Scholar] [CrossRef]
- Elli, L.; Topa, M.; Rimondi, A. Protein-Losing Enteropathy. Curr. Opin. Gastroenterol. 2020, 36, 238–244. [Google Scholar] [CrossRef]
- Kanwar, J.R.; Roy, K.; Patel, Y.; Zhou, S.F.; Singh, M.R.; Singh, D.; Nasir, M.; Sehgal, R.; Sehgal, A.; Singh, R.S.; et al. Multifunctional Iron Bound Lactoferrin and Nanomedicinal Approaches to Enhance Its Bioactive Functions. Molecules 2015, 20, 9703–9731. [Google Scholar] [CrossRef]
- Johanson, B.; Virtanen, A.I.; Tweit, R.C.; Dodson, R.M. Isolation of an Iron-Containing Red Protein from Human Milk. Acta Chem. Scand. 1960, 14, 510–512. [Google Scholar] [CrossRef]
- Kell, D.B.; Heyden, E.L.; Pretorius, E. The Biology of Lactoferrin, an Iron-Binding Protein That Can Help Defend Against Viruses and Bacteria. Front. Immunol. 2020, 11, 1221. [Google Scholar] [CrossRef]
- Cao, X.; Ren, Y.; Lu, Q.; Wang, K.; Wu, Y.; Wang, Y.; Zhang, Y.; Cui, X.-S.; Yang, Z.; Chen, Z. Lactoferrin: A Glycoprotein That Plays an Active Role in Human Health. Front. Nutr. 2022, 9, 1018336. [Google Scholar] [CrossRef]
- Ward, P.P.; Uribe-Luna, S.; Conneely, O.M. Lactoferrin and Host Defense. Biochem. Cell Biol. 2002, 80, 95–102. [Google Scholar] [CrossRef]
- Conneely, O.M. Antiinflammatory Activities of Lactoferrin. J. Am. Coll. Nutr. 2001, 20, 389S–395S. [Google Scholar] [CrossRef]
- Ward, P.P.; Conneely, O.M. Lactoferrin: Role in Iron Homeostasis and Host Defense against Microbial Infection. BioMetals 2004, 17, 203–208. [Google Scholar] [CrossRef]
- Koszewicz, M.; Mulak, A.; Dziadkowiak, E.; Budrewicz, S. Is Fecal Calprotectin an Applicable Biomarker of Gut Immune System Activation in Chronic Inflammatory Demyelinating Polyneuropathy?—A Pilot Study. Front. Hum. Neurosci. 2021, 15, 733070. [Google Scholar] [CrossRef] [PubMed]
- Zubeldia-Varela, E.; Barker-Tejeda, T.C.; Blanco-Pérez, F.; Infante, S.; Zubeldia, J.M.; Pérez-Gordo, M. Non-IgE-Mediated Gastrointestinal Food Protein-Induced Allergic Disorders. Clinical Perspectives and Analytical Approaches. Foods 2021, 10, 2662. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.J.; Xie, X.L.; Li, Y.; Deng, X.Z. Current Status of Fecal Calprotectin as a Diagnostic or Monitoring Biomarker for Cow’s Milk Protein Allergy in Children: A Scoping Review. World J. Pediatr. 2021, 17, 63–70. [Google Scholar] [CrossRef]
- Murakami, M. Lipid Mediators in Life Science. Exp. Anim. 2011, 60, 7–20. [Google Scholar] [CrossRef]
- McHarg, A.S.; Leach, S. The Role of the Gut Microbiome in Paediatric Irritable Bowel Syndrome. AIMS Microbiol. 2022, 8, 454–469. [Google Scholar] [CrossRef]
- Sinclair, E.M.; Cohen, C.C.; Tran, V.; Jones, D.P.; Alvarez, J.A.; Kamaleswaran, R.; Rad, M.G.; Kruszewski, P.G.; Vos, M.B. Untargeted, High-Resolution Metabolomics in Pediatric Eosinophilic Esophagitis. J. Pediatr. Gastroenterol. Nutr. 2023, 76, 355–363. [Google Scholar] [CrossRef]
- Majamaa, H.; Aittoniemi, J.; Miettinen, A. Increased Concentration of Fecal Alpha1-Antitrypsin Is Associated with Cow’s Milk Allergy in Infants with Atopic Eczema. Clin. Exp. Allergy 2001, 31, 590–592. [Google Scholar] [CrossRef]
- Kalach, N.; Kapel, N.; Waligora-Dupriet, A.J.; Castelain, M.C.; Cousin, M.O.; Sauvage, C.; Ba, F.; Nicolis, I.; Campeotto, F.; Butel, M.J.; et al. Intestinal Permeability and Fecal Eosinophil-Derived Neurotoxin Are the Best Diagnosis Tools for Digestive Non-IgE-Mediated Cow’s Milk Allergy in Toddlers. Clin. Chem. Lab. Med. 2013, 51, 351–361. [Google Scholar] [CrossRef]
- Borkowska, A.; Liberek, A.; Łuczak, G.; Jankowska, A.; Plata-Nazar, K.; Korzon, M.; Kamińska, B. Fecal Lactoferrin, a Marker of Intestinal Inflammation in Children with Inflammatory Bowel Disease. Acta Biochim. Pol. 2015, 62, 541–545. [Google Scholar] [CrossRef]
- Mosli, M.H.; Zou, G.; Garg, S.K.; Feagan, S.G.; MacDonald, J.K.; Chande, N.; Sandborn, W.J.; Feagan, B.G. C-Reactive Protein, Fecal Calprotectin, and Stool Lactoferrin for Detection of Endoscopic Activity in Symptomatic Inflammatory Bowel Disease Patients: A Systematic Review and Meta-Analysis. Am J. Gastroenterol. 2015, 110, 802–819; quiz 820. [Google Scholar] [CrossRef]
- Berni Canani, R.; Terrin, G.; Rapacciuolo, L.; Miele, E.; Siani, M.C.; Puzone, C.; Cosenza, L.; Staiano, A.; Troncone, R. Faecal Calprotectin as Reliable Non-Invasive Marker to Assess the Severity of Mucosal Inflammation in Children with Inflammatory Bowel Disease. Dig. Liver Dis. 2008, 40, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Rapacciuolo, L.; Romano, M.T.; de Horatio, L.T.; Terrin, G.; Manguso, F.; Cirillo, P.; Paparo, F.; Troncone, R. Diagnostic Value of Faecal Calprotectin in Paediatric Gastroenterology Clinical Practice. Dig. Liver. Dis. 2004, 36, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Bunn, S.K.; Bisset, W.M.; Main, M.J.C.; Golden, B.E. Fecal Calprotectin as a Measure of Disease Activity in Childhood Inflammatory Bowel Disease. J. Pediatr. Gastroenterol. Nutr. 2001, 32, 171–177. [Google Scholar] [CrossRef]
- Beşer, Ö.F.; Sancak, S.; Erkan, T.; Kutlu, T.; Çokuǧraş, H.; Cokuǧraş, F.Ç. Can Fecal Calprotectin Level Be Used as a Markers of Inflammation in the Diagnosis and Follow-Up of Cow’s Milk Protein Allergy? Allergy Asthma Immunol. Res. 2014, 6, 33–38. [Google Scholar] [CrossRef]
- Díaz, M.; Guadamuro, L.; Espinosa-Martos, I.; Mancabelli, L.; Jiménez, S.; Molinos-Norniella, C.; Pérez-Solis, D.; Milani, C.; Rodríguez, J.M.; Ventura, M.; et al. Microbiota and Derived Parameters in Fecal Samples of Infants with Non-IgE Cow’s Milk Protein Allergy under a Restricted Diet. Nutrients 2018, 10, 1481. [Google Scholar] [CrossRef]
- Otten, C.M.T.; Kok, L.; Witteman, B.J.M.; Baumgarten, R.; Kampman, E.; Moons, K.G.M.; de Wit, N.J. Diagnostic Performance of Rapid Tests for Detection of Fecal Calprotectin and Lactoferrin and Their Ability to Discriminate Inflammatory from Irritable Bowel Syndrome. Clin. Chem. Lab. Med. 2008, 46, 1275–1280. [Google Scholar] [CrossRef]
- Menees, S.B.; Powell, C.; Kurlander, J.; Goel, A.; Chey, W.D. A Meta-Analysis of the Utility of C-Reactive Protein, Erythrocyte Sedimentation Rate, Fecal Calprotectin, and Fecal Lactoferrin to Exclude Inflammatory Bowel Disease in Adults with IBS. Am. J. Gastroenterol. 2015, 110, 444–454. [Google Scholar] [CrossRef]
- Barbut, F.; Gouot, C.; Lapidus, N.; Suzon, L.; Syed-Zaidi, R.; Lalande, V.; Eckert, C. Faecal Lactoferrin and Calprotectin in Patients with Clostridium Difficile Infection: A Case-Control Study. Eur. J. Clin. Microbiol. Infect Dis. 2017, 36, 2423–2430. [Google Scholar] [CrossRef]
- Drago-Serrano, M.E.; Campos-Rodríguez, R.; Carrero, J.C.; de la Garza, M. Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections. Int. J. Mol. Sci. 2017, 18, 501. [Google Scholar] [CrossRef]
- Gheibi, N.; Ghorbani, M.; Shariatifar, H.; Farasat, A. Effects of Unsaturated Fatty Acids (Arachidonic/Oleic Acids) on Stability and Structural Properties of Calprotectin Using Molecular Docking and Molecular Dynamics Simulation Approach. PLoS ONE 2020, 15, e0230780. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Michels, N.; van de Wiele, T.; de Henauw, S. Chronic Psychosocial Stress and Gut Health in Children: Associations With Calprotectin and Fecal Short-Chain Fatty Acids. Psychosom. Med. 2017, 79, 927–935. [Google Scholar] [CrossRef]
- Aho, V.T.E.; Houser, M.C.; Pereira, P.A.B.; Chang, J.; Rudi, K.; Paulin, L.; Hertzberg, V.; Auvinen, P.; Tansey, M.G.; Scheperjans, F. Relationships of Gut Microbiota, Short-Chain Fatty Acids, Inflammation, and the Gut Barrier in Parkinson’s Disease. Mol. Neurodegener. 2021, 16, 6. [Google Scholar] [CrossRef]
- Vaghari-Tabari, M.; Moein, S.; Qujeq, D.; Kashifard, M.; Hajian-Tilaki, K. Positive Correlation of Fecal Calprotectin With Serum Antioxidant Enzymes in Patients With Inflammatory Bowel Disease: Accidental Numerical Correlation or a New Finding? Am. J. Med. Sci. 2018, 355, 449–455. [Google Scholar] [CrossRef]
- Pijls, K.E.; Jonkers, D.M.A.E.; Elizalde, M.; Drittij-Reijnders, M.J.; Haenen, G.R.; Bast, A.; Masclee, A.A.M.; Koek, G.H. Is Intestinal Oxidative Stress Involved in Patients with Compensated Liver Cirrhosis? Ann. Hepatol. 2016, 15, 402–409. [Google Scholar] [CrossRef]
- Larsen, S.B.; Grove, E.L.; Pareek, M.; Kristensen, S.D.; Hvas, A.M. Calprotectin and Platelet Aggregation in Patients with Stable Coronary Artery Disease. PLoS ONE 2015, 10, e0125992. [Google Scholar] [CrossRef]
- Obeso, D.; Mera-Berriatua, L.; Rodríguez-Coira, J.; Rosace, D.; Fernández, P.; Martín-Antoniano, I.A.; Santaolalla, M.; Marco Martín, G.; Chivato, T.; Fernández-Rivas, M.; et al. Multi-Omics Analysis Points to Altered Platelet Functions in Severe Food-Associated Respiratory Allergy. Allergy 2018, 73, 2137–2149. [Google Scholar] [CrossRef]
- Schwarz, J.; Vecka, M.; Stožický, F.; Pomahačová, R.; Staňková, B.; Tvrzická, E.; Kreslová, M.; Zahálková, R.; Sýkora, J. The Assessment of Plasma Fatty Acid Profiles in Newly Diagnosed Treatment-Naive Paediatric Crohn’s Disease. Physiol. Res. 2021, 70, 799–808. [Google Scholar] [CrossRef]
- Bjerrum, J.T.; Wang, Y.; Hao, F.; Coskun, M.; Ludwig, C.; Günther, U.; Nielsen, O.H. Metabonomics of Human Fecal Extracts Characterize Ulcerative Colitis, Crohn’s Disease and Healthy Individuals. Metabolomics 2015, 11, 122–133. [Google Scholar] [CrossRef]
- Sewell, G.W.; Hannun, Y.A.; Han, X.; Koster, G.; Bielawski, J.; Goss, V.; Smith, P.J.; Rahman, F.Z.; Vega, R.; Bloom, S.L.; et al. Lipidomic Profiling in Crohn’s Disease: Abnormalities in Phosphatidylinositols, with Preservation of Ceramide, Phosphatidylcholine and Phosphatidylserine Composition. Int. J. Biochem. Cell Biol. 2012, 44, 1839–1846. [Google Scholar] [CrossRef] [PubMed]
- Calcaterra, V.; de Amici, M.; Leonard, M.M.; de Silvestri, A.; Pelizzo, G.; Buttari, N.; Michev, A.; Leggio, M.; Larizza, D.; Cena, H. Serum Calprotectin Level in Children: Marker of Obesity and Its Metabolic Complications. Ann. Nutr. Metab. 2018, 73, 177–183. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Brueton, M.; Dupont, C.; Hill, D.; Isolauri, E.; Koletzko, S.; Oranje, A.P.; Staiano, A. Guidelines for the Diagnosis and Management of Cow’s Milk Protein Allergy in Infants. Arch. Dis. Child. 2007, 92, 902–908. [Google Scholar] [CrossRef]
- Kemp, A.S.; Hill, D.J.; Allen, K.J.; Anderson, K.; Davidson, G.P.; Day, A.S.; Heine, R.G.; Peake, J.E.; Prescott, S.L.; Shugg, A.W.; et al. Guidelines for the Use of Infant Formulas to Treat Cows Milk Protein Allergy: An Australian Consensus Panel Opinion. Med. J. Aust. 2008, 188, 109–112. [Google Scholar] [CrossRef]
- Lamichhane, S.; Sen, P.; Alves, M.A.; Ribeiro, H.C.; Raunioniemi, P.; Hyötyläinen, T.; Orešič, M. Linking Gut Microbiome and Lipid Metabolism: Moving beyond Associations. Metabolites 2021, 11, 55. [Google Scholar] [CrossRef]
- World Medical Association World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [CrossRef]
- Dreborg, S. Skin Testing. The Safety of Skin Tests and the Information Obtained from Using Different Methods and Concentrations of Allergen. Allergy 1993, 48, 473–475. [Google Scholar] [CrossRef]
- Thompson, J.A.; Oliveira, R.A.; Djukovic, A.; Ubeda, C.; Xavier, K.B. Manipulation of the Quorum Sensing Signal AI-2 Affects the Antibiotic-Treated Gut Microbiota. Cell Rep. 2015, 10, 1861–1871. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Tamames, J.; Puente-Sánchez, F. SqueezeMeta, A Highly Portable, Fully Automatic Metagenomic Analysis Pipeline. Front. Microbiol. 2019, 9, 3349. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “vegan” Title Community Ecology Package, Version 2.5-7; 2020. CRAN Repository. Available online: https://github.com/vegandevs/vegan (accessed on 18 May 2023).
- Sanchez-Solares, J.; Delgado-Dolset, M.I.; Mera-Berriatua, L.; Hormias-Martin, G.; Cumplido, J.A.; Saiz, V.; Carrillo, T.; Moreno-Aguilar, C.; Escribese, M.M.; Gomez-Casado, C.; et al. Respiratory Allergies with No Associated Food Allergy Disrupt Oral Mucosa Integrity. Allergy 2019, 74, 2261–2265. [Google Scholar] [CrossRef]
- Rosace, D.; Gomez-Casado, C.; Fernandez, P.; Perez-Gordo, M.; del Carmen Dominguez, M.; Vega, A.; Belver, M.T.; Ramos, T.; Vega, F.; Marco, G.; et al. Profilin-Mediated Food-Induced Allergic Reactions Are Associated with Oral Epithelial Remodeling. J. Allergy Clin. Immunol. 2019, 143, 681–690.e1. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Pathway | KP | p-Value | Adj. p-Value | log2FC |
|---|---|---|---|---|
| Glycerophospholipid metabolism * | ko00564 | 0.000008 | 0.002419 | 0.243 |
| Nicotinate and nicotinamide metabolism | ko00760 | 0.001577 | 0.246808 | 0.233 |
| Primary immunodeficiency | ko05340 | 0.005161 | 0.538457 | 0.328 |
| Inositol phosphate metabolism | ko00562 | 0.008359 | 0.654097 | 0.451 |
| Glycerolipid metabolism | ko00561 | 0.016640 | 0.892611 | 0.183 |
| Phosphatidylinositol signaling system | ko04070 | 0.017552 | 0.892611 | 0.203 |
| Lipoic acid metabolism | ko00785 | 0.022028 | 0.892611 | 0.328 |
| Atrazine degradation | ko00791 | 0.028442 | 0.892611 | 0.983 |
| Glycine, serine, and threonine metabolism | ko00260 | 0.028572 | 0.892611 | 0.139 |
| Chlorocyclohexane and chlorobenzene degradation | ko00361 | 0.030698 | 0.892611 | 0.404 |
| Biosynthesis of unsaturated fatty acids | ko01040 | 0.035266 | 0.892611 | 0.337 |
| Carbon fixation pathways in prokaryotes | ko00720 | 0.035961 | 0.892611 | −0.175 |
| Ubiquitin system | ko04121 | 0.046358 | 0.892611 | 0.461 |
| Limonene and pinene degradation | ko00903 | 0.047001 | 0.892611 | 0.438 |
| Sample ID | Age (Months) | Gender | Condition | Delivery Mode | Antibiotics in Labor | Feeding |
|---|---|---|---|---|---|---|
| AI_01 | 4 | M | Allergic | C-section | Yes | BM+H |
| AI_02 | 5 | F | Allergic | C-section | No | BM |
| AI_03 | 5 | M | Allergic | Vaginal | No | BM+H |
| AI_04 | 4 | M | Allergic | C-section | No | BM+H |
| AI_05 | 5 | F | Allergic | Vaginal | Yes | BM+H |
| AI_06 | 3 | F | Allergic | Vaginal | No | BM+H |
| AI_07 | 2 | M | Allergic | Vaginal | No | H |
| AI_08 | 6 | F | Allergic | Vaginal | No | H |
| AI_09 | 6 | M | Allergic | Vaginal | No | H |
| AI_10 | 3 | F | Allergic | C-section | Yes | BM+MF |
| AI_11 | 5 | M | Allergic | Vaginal | No | H |
| AI_12 | 6 | F | Allergic | Vaginal | No | BM |
| AI_13 | 5 | M | Allergic | Vaginal | No | H |
| AI_14 | 5 | F | Allergic | Vaginal | No | BM+MF |
| AI_15 | 3 | M | Allergic | Vaginal | No | BM+H |
| AI_16 | 4 | F | Allergic | Vaginal | Yes | BM |
| AI_17 | 5 | F | Allergic | Vaginal | No | H |
| AI_19 | 5 | F | Allergic | Vaginal | No | BM+H |
| AI_20 | 7 | M | Allergic | Vaginal | No | BM |
| AI_21 | 6 | F | Allergic | Vaginal | No | BM+MF |
| AI_22 | 6 | F | Allergic | C-section | No | BM |
| AI_23 | 6 | F | Allergic | Vaginal | No | H |
| AI_24 | 6 | F | Allergic | Vaginal | No | BM+H |
| AI_26 | 8 | F | Allergic | Vaginal | No | BM+H |
| AI_27 | 5 | M | Allergic | Vaginal | No | BM+H |
| AI_28 | 7 | M | Allergic | Vaginal | Yes | BM |
| AI_29 | 5 | F | Allergic | Vaginal | Yes | BM+H |
| AI_30 | 6 | M | Allergic | Vaginal | No | H |
| AI_31 | 4 | F | Allergic | Vaginal | No | H |
| AI_32 | 6 | M | Allergic | C-section | No | H |
| AI_33 | 4 | F | Allergic | Vaginal | No | BM+H |
| AI_34 | 3 | M | Allergic | Vaginal | No | BM+H |
| AI_35 | 6 | M | Allergic | Vaginal | No | H |
| AI_36 | 1 | F | Allergic | Vaginal | No | H |
| CI_01 | 6 | M | Non-allergic | Vaginal | No | BM |
| CI_02 | 7 | M | Non-allergic | Vaginal | No | BM+MF |
| CI_03 | 6 | F | Non-allergic | Vaginal | No | BM+MF |
| CI_04 | 6 | F | Non-allergic | Vaginal | Yes | BM+MF |
| CI_05 | 6 | M | Non-allergic | Vaginal | No | MF |
| CI_06 | 6 | M | Non-allergic | Vaginal | No | MF |
| CI_07 | 8 | M | Non-allergic | Vaginal | No | BM+MF |
| CI_08 | 5 | F | Non-allergic | Vaginal | No | MF |
| CI_09 | 5 | F | Non-allergic | C-section | No | MF |
| CI_10 | 5 | F | Non-allergic | C-section | No | MF |
| CI_11 | 5 | F | Non-allergic | Vaginal | No | BM |
| CI_12 | 4 | M | Non-allergic | Vaginal | No | MF |
| CI_13 | 3 | F | Non-allergic | Vaginal | No | MF |
| CI_14 | 4 | F | Non-allergic | Vaginal | No | BM+MF |
| CI_15 | 1 | F | Non-allergic | Vaginal | No | BM+MF |
| CI_16 | 3 | F | Non-allergic | C-section | Yes | BM+MF |
| CI_23 | 4 | F | Non-allergic | C-section | - | BM+MF |
| CI_24 | 3 | F | Non-allergic | C-section | No | BM+MF |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubeldia-Varela, E.; Barker-Tejeda, T.C.; Mera-Berriatua, L.; Bazire, R.; Cabrera-Freitag, P.; Ubeda, C.; Barber, D.; Francino, M.P.; Rojo, D.; Ibáñez-Sandín, M.D.; et al. Further Insights into the Gut Microbiota of Cow’s Milk Allergic Infants: Analysis of Microbial Functionality and Its Correlation with Three Fecal Biomarkers. Int. J. Mol. Sci. 2023, 24, 9247. https://doi.org/10.3390/ijms24119247
Zubeldia-Varela E, Barker-Tejeda TC, Mera-Berriatua L, Bazire R, Cabrera-Freitag P, Ubeda C, Barber D, Francino MP, Rojo D, Ibáñez-Sandín MD, et al. Further Insights into the Gut Microbiota of Cow’s Milk Allergic Infants: Analysis of Microbial Functionality and Its Correlation with Three Fecal Biomarkers. International Journal of Molecular Sciences. 2023; 24(11):9247. https://doi.org/10.3390/ijms24119247
Chicago/Turabian StyleZubeldia-Varela, Elisa, Tomás Clive Barker-Tejeda, Leticia Mera-Berriatua, Raphaëlle Bazire, Paula Cabrera-Freitag, Carles Ubeda, Domingo Barber, María Pilar Francino, David Rojo, María Dolores Ibáñez-Sandín, and et al. 2023. "Further Insights into the Gut Microbiota of Cow’s Milk Allergic Infants: Analysis of Microbial Functionality and Its Correlation with Three Fecal Biomarkers" International Journal of Molecular Sciences 24, no. 11: 9247. https://doi.org/10.3390/ijms24119247
APA StyleZubeldia-Varela, E., Barker-Tejeda, T. C., Mera-Berriatua, L., Bazire, R., Cabrera-Freitag, P., Ubeda, C., Barber, D., Francino, M. P., Rojo, D., Ibáñez-Sandín, M. D., & Pérez-Gordo, M. (2023). Further Insights into the Gut Microbiota of Cow’s Milk Allergic Infants: Analysis of Microbial Functionality and Its Correlation with Three Fecal Biomarkers. International Journal of Molecular Sciences, 24(11), 9247. https://doi.org/10.3390/ijms24119247