
Effects of Cobra Cardiotoxins on Intracellular Calcium and the Contracture of Rat Cardiomyocytes Depend on Their Structural Types
 , , and
, , and 
Abstract
:1. Introduction
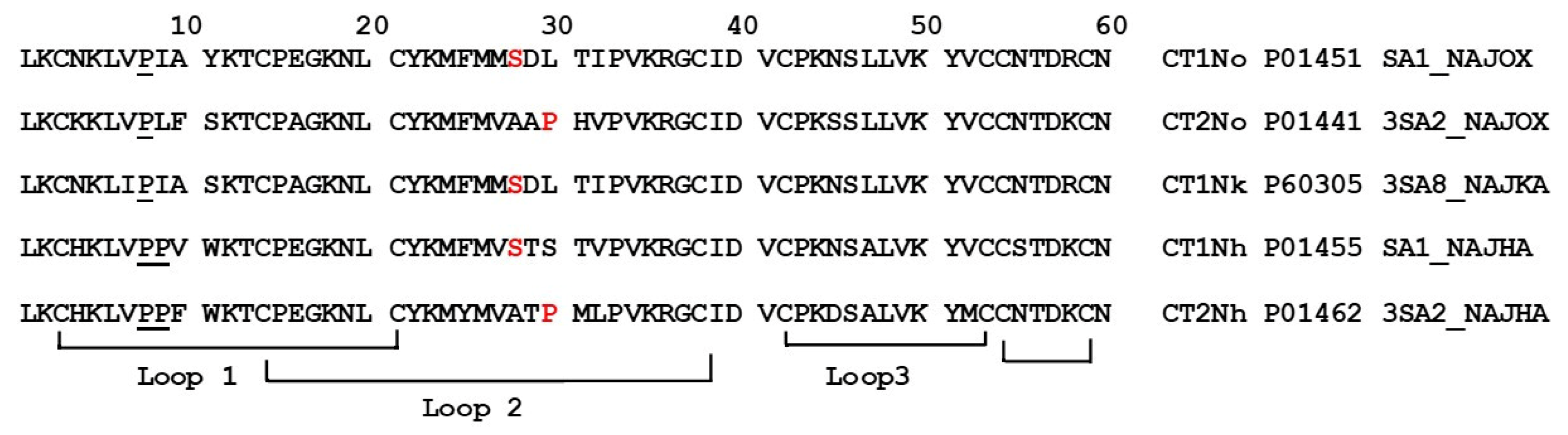
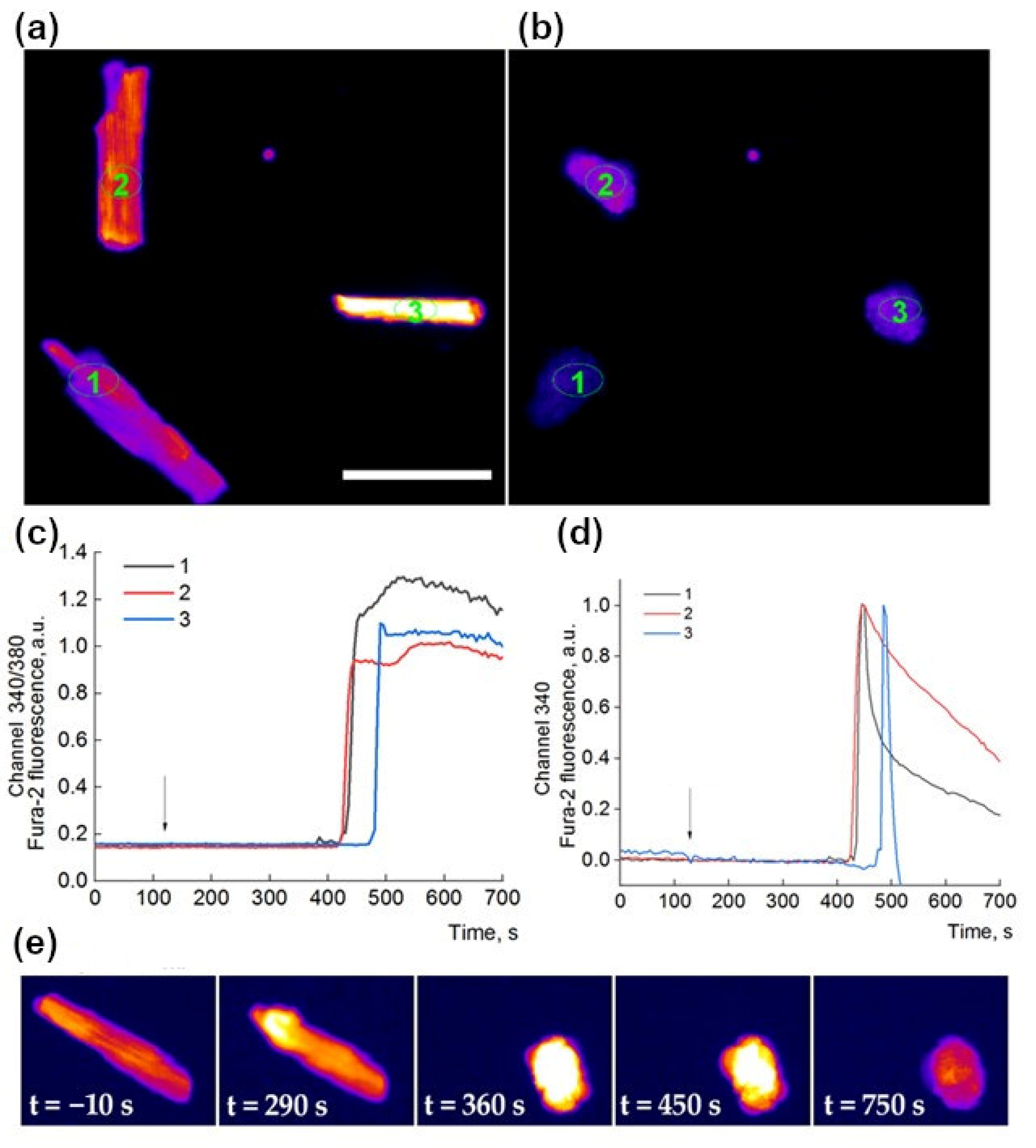
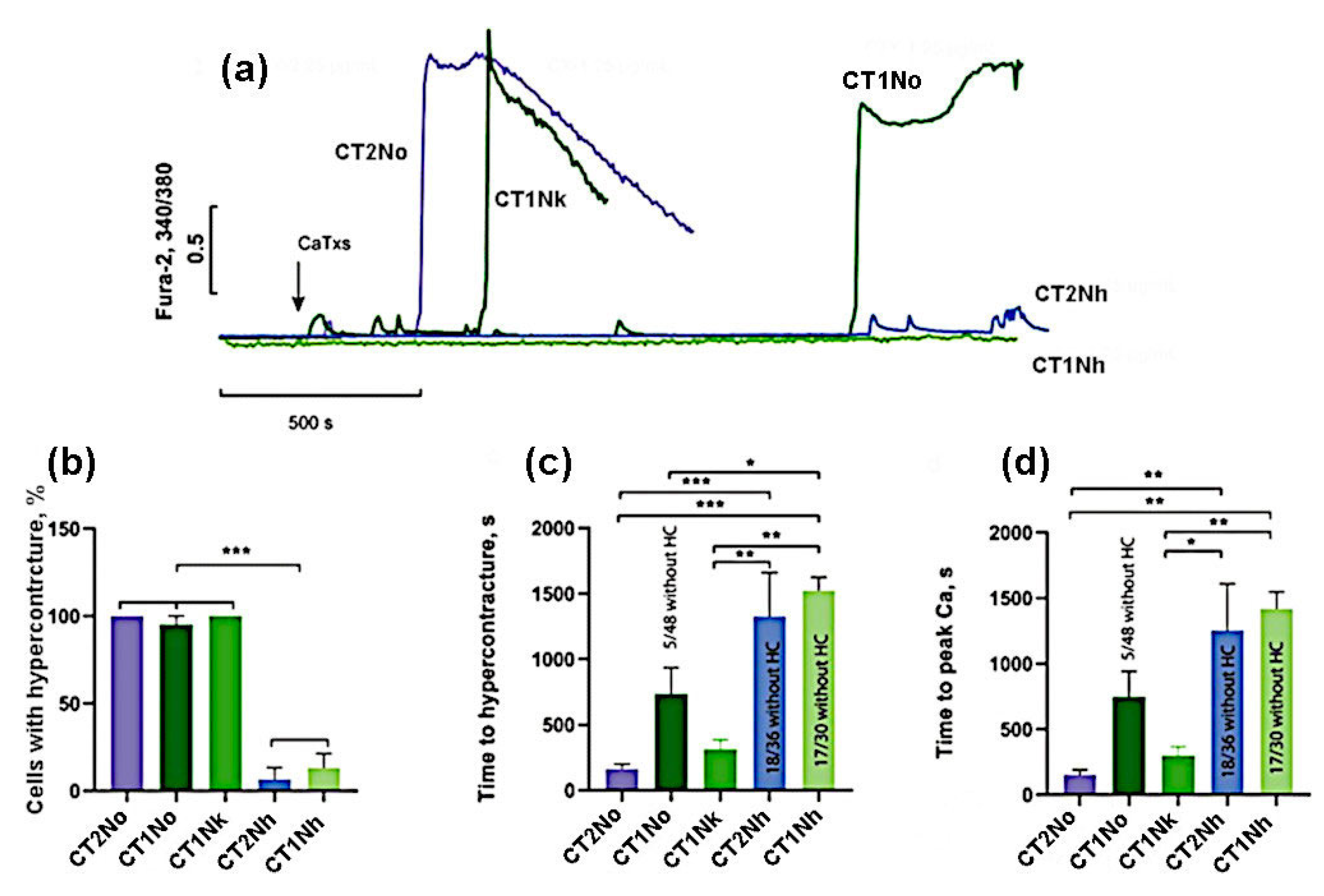
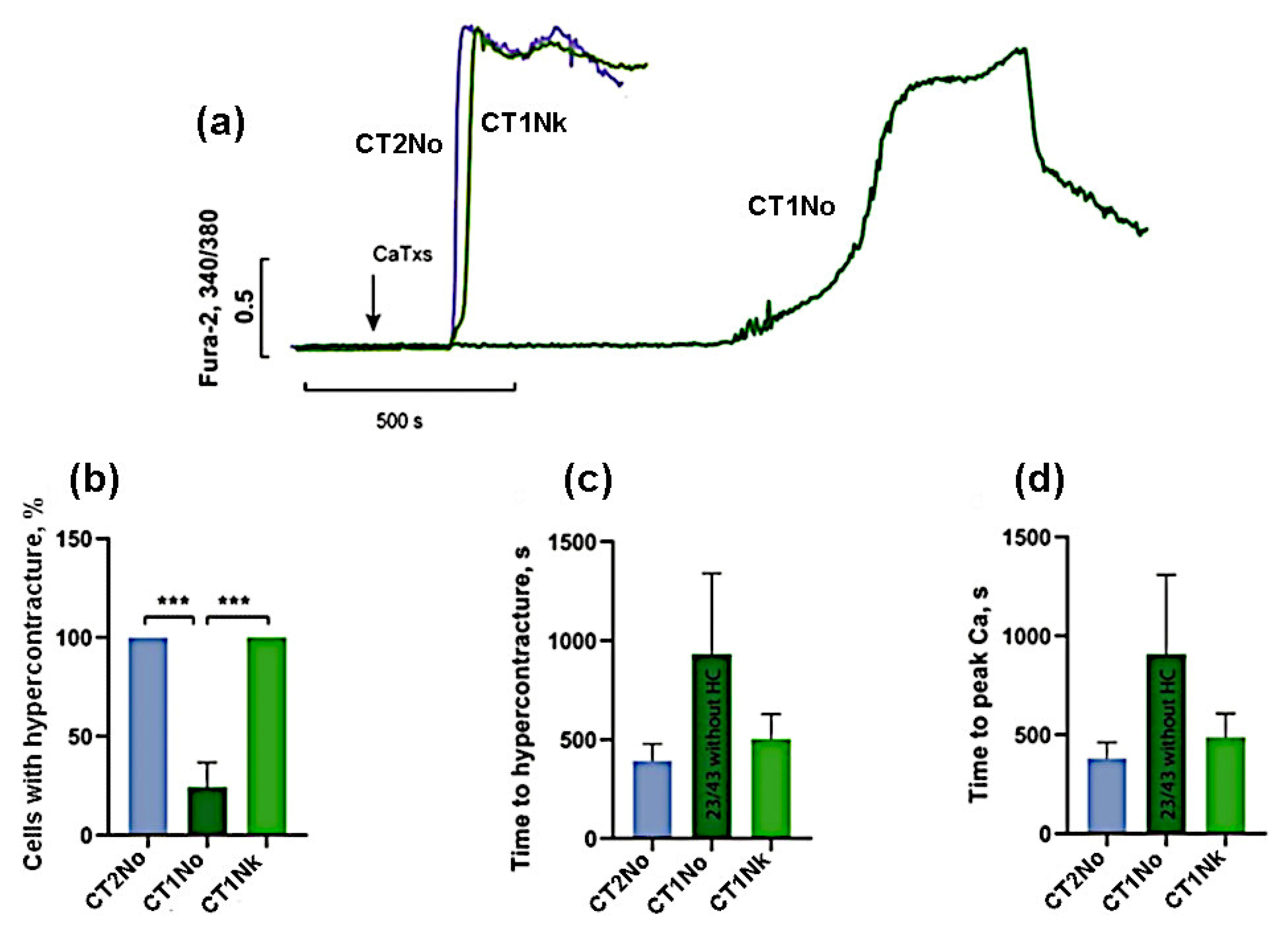
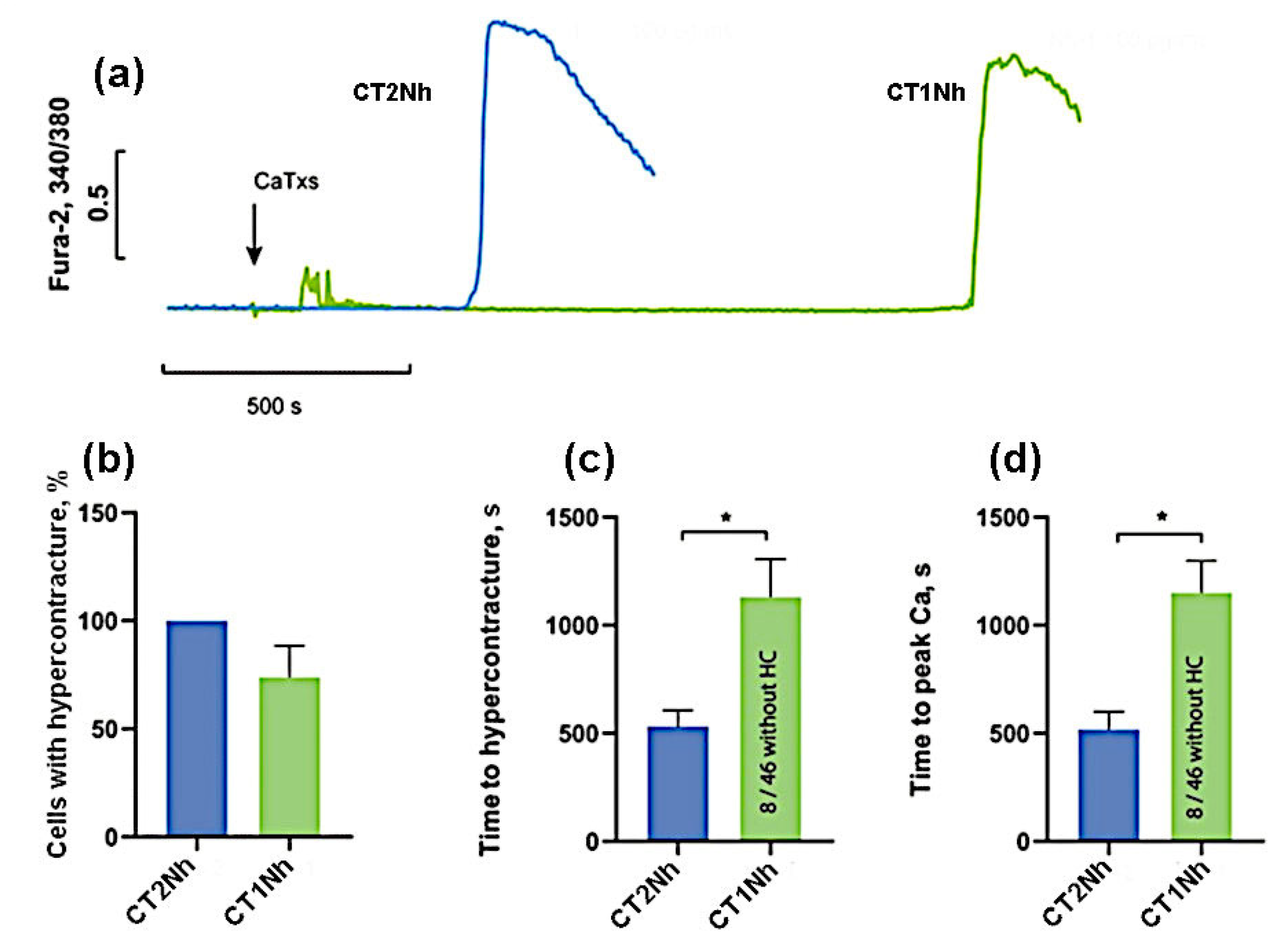
2. Results
- (1)
- the group with high activity, which included CT2No, CT1Nk, and CT1No;
- (2)
- the group with low activity: CT2Nh and CT1Nh.

3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cardiomyocyte Preparation
4.3. Dye Loading
4.4. Fluorescence Microscopy
4.5. Reagents Application
4.6. Image Processing
4.7. Data Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kalita, B.; Utkin, Y.N.; Mukherjee, A.K. Current Insights in the Mechanisms of Cobra Venom Cytotoxins and Their Complexes in Inducing Toxicity: Implications in Antivenom Therapy. Toxins 2022, 14, 839. [Google Scholar] [CrossRef] [PubMed]
- Chien, K.Y.; Chiang, C.M.; Hseu, Y.C.; Vyas, A.A.; Rule, G.S.; Wu, W. Two distinct types of cardiotoxin as revealed by the structure and activity relationship of their interaction with zwitterionic phospholipid dispersions. J. Biol. Chem. 1994, 269, 14473–14483. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-S.; Chung, F.-Y.; Tjong, S.-C.; Goh, K.-S.; Huang, W.-N.; Chien, K.-Y.; Wu, P.-L.; Lin, H.-C.; Chen, C.-J.; Wu, W. Structural difference between group I and group II cobra cardiotoxins: X-ray, NMR, and CD analysis of the effect of cis-proline conformation on three-fingered toxins. Biochemistry 2005, 44, 7414–7426. [Google Scholar] [CrossRef] [PubMed]
- Grognet, J.M.; Ménez, A.; Drake, A.; Hayashi, K.; Morrison, I.E.; Hider, R.C. Circular dichroic spectra of elapid cardiotoxins. Eur. J. Biochem. 1988, 172, 383–388. [Google Scholar] [CrossRef]
- Dubovskii, P.V.; Lesovoy, D.M.; Dubinnyi, M.A.; Konshina, A.G.; Utkin, Y.N.; Efremov, R.G.; Arseniev, A.S. Interaction of three-finger toxins with phospholipid membranes: Comparison of S- and P-type cytotoxins. Biochem. J. 2005, 387, 807–815. [Google Scholar] [CrossRef]
- Dubovskii, P.V.; Ignatova, A.A.; Alekseeva, A.S.; Starkov, V.G.; Boldyrev, I.A.; Feofanov, A.V.; Utkin, Y.N. Membrane-Disrupting Activity of Cobra Cytotoxins Is Determined by Configuration of the N-Terminal Loop. Toxins 2022, 15, 6. [Google Scholar] [CrossRef]
- Averin, A.S.; Astashev, M.E.; Andreeva, T.V.; Tsetlin, V.I.; Utkin, Y.N. Cardiotoxins from Cobra Naja oxiana Change the Force of Contraction and the Character of Rhythmoinotropic Phenomena in the Rat Myocardium. Dokl. Biochem. Biophys. 2019, 487, 282–286. [Google Scholar] [CrossRef]
- Averin, A.S.; Nenov, M.N.; Starkov, V.G.; Tsetlin, V.I.; Utkin, Y.N. Effects of Cardiotoxins from Naja oxiana Cobra Venom on Rat Heart Muscle and Aorta: A Comparative Study of Toxin-Induced Contraction Mechanisms. Toxins 2022, 14, 88. [Google Scholar] [CrossRef]
- Averin, A.S.; Goltyaev, M.V.; Andreeva, T.V.; Starkov, V.G.; Tsetlin, V.I.; Utkin, Y.N. S- and P-type cobra venom cardiotoxins differ in their action on isolated rat heart. J. Venom. Anim. Toxins Incl. Trop. Dis. 2022, 28, e20210110. [Google Scholar] [CrossRef]
- Tzeng, W.F.; Chen, Y.H. Suppression of snake-venom cardiotoxin-induced cardiomyocyte degeneration by blockage of Ca2+ influx or inhibition of non-lysosomal proteinases. Biochem. J. 1988, 256, 89–95. [Google Scholar] [CrossRef]
- Wang, H.X.; Lau, S.Y.; Huang, S.J.; Kwan, C.Y.; Wong, T.M. Cobra venom cardiotoxin induces perturbations of cytosolic calcium homeostasis and hypercontracture in adult rat ventricular myocytes. J. Mol. Cell. Cardiol. 1997, 29, 2759–2770. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Wu, W.-g. Amphiphilic beta-sheet cobra cardiotoxin targets mitochondria and disrupts its network. FEBS Lett. 2005, 579, 3169–3174. [Google Scholar] [CrossRef] [PubMed]
- Berezhnov, A.V.; Fedotova, E.I.; Nenov, M.N.; Kasymov, V.A.; Pimenov, O.Y.; Dynnik, V.V. Dissecting Cellular Mechanisms of Long-Chain Acylcarnitines-Driven Cardiotoxicity: Disturbance of Calcium Homeostasis, Activation of Ca2+-Dependent Phospholipases, and Mitochondrial Energetics Collapse. Int. J. Mol. Sci. 2020, 21, 7461. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Meana, M.; Abellán, A.; Miró-Casas, E.; Garcia-Dorado, D. Opening of mitochondrial permeability transition pore induces hypercontracture in Ca2+ overloaded cardiac myocytes. Basic Res. Cardiol. 2007, 102, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Antoons, G.; Mubagwa, K.; Nevelsteen, I.; Sipido, K.R. Mechanisms underlying the frequency dependence of contraction and Ca(2+)(i) transients in mouse ventricular myocytes. J. Physiol. 2002, 543, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Picht, E.; DeSantiago, J.; Huke, S.; Kaetzel, M.A.; Dedman, J.R.; Bers, D.M. CaMKII inhibition targeted to the sarcoplasmic reticulum inhibits frequency-dependent acceleration of relaxation and Ca2+ current facilitation. J. Mol. Cell. Cardiol. 2007, 42, 196–205. [Google Scholar] [CrossRef]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; León, G.; Warrell, D.A.; Theakston, R.D.G.; et al. Snake venomics of African spitting cobras: Toxin composition and assessment of congeneric cross-reactivity of the pan-African EchiTAb-Plus-ICP antivenom by antivenomics and neutralization approaches. J. Proteome Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef]
- Grishin, E.V.; Sukhikh, A.P.; Adamovich, T.B.; Ovchinnikov, Y.A. Isolation, properties, and amino acid sequence of two cytotoxins from the venom of the Central Asian cobra Naja oxiana. Bioorg. Khim. 1976, 2, 1018–1034. [Google Scholar]
- Ryabinin, V.V.; Ziganshin, R.H.; Starkov, V.G.; Tsetlin, V.I.; Utkin, Y.N. Intraspecific Variability in the Composition of the Venom from Monocled Cobra (Naja kaouthia). Russ. J. Bioorg. Chem. 2019, 45, 107–121. [Google Scholar] [CrossRef]
- Adamude, F.A.; Dingwoke, E.J.; Abubakar, M.S.; Ibrahim, S.; Mohamed, G.; Klein, A.; Sallau, A.B. Proteomic analysis of three medically important Nigerian Naja (Naja haje, Naja katiensis and Naja nigricollis) snake venoms. Toxicon 2021, 197, 24–32. [Google Scholar] [CrossRef]
- Malih, I.; Rusmili, A.M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of Moroccan cobra Naja haje legionis venom using tandem mass spectrometry. J. Proteom. 2014, 96, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Gasanov, S.E.; Shrivastava, I.H.; Israilov, F.S.; Kim, A.A.; Rylova, K.A.; Zhang, B.; Dagda, R.K. Naja naja oxiana Cobra Venom Cytotoxins CTI and CTII Disrupt Mitochondrial Membrane Integrity: Implications for Basic Three-Fingered Cytotoxins. PLoS ONE 2015, 10, e0129248. [Google Scholar] [CrossRef]
- Fakhri, A.; Omranipour, R.; Fakhri, S.; Mirshamsi, M.; Zangeneh, F.; Vatanpour, H.; Pourahmad, J. Naja Naja Oxiana Venom Fraction Selectively Induces ROS-Mediated Apoptosis in Human Colorectal Tumor Cells by Directly Targeting Mitochondria. Asian Pac. J. Cancer Prev. 2017, 18, 2201–2208. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.-J.; Huang, C.-H.; Lee, Y.-C.; Wang, L.-J.; Chiou, J.-T.; Chang, L.-S. Naja atra cardiotoxins enhance the protease activity of chymotrypsin. Int. J. Biol. Macromol. 2019, 136, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Marshall, R.J.; Karlsson, E. Effects of purified cardiotoxins from the Thailand cobra (Naja naja siamensis) on isolated skeletal and cardiac muscle preparations. Toxicon 1982, 20, 379–396. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Monette, R.; Lee, S.-C.; Morley, P.; Wu, W.-g. Cobra cardiotoxin-induced cell death in fetal rat cardiomyocytes and cortical neurons: Different pathway but similar cell surface target. Toxicon 2005, 46, 430–440. [Google Scholar] [CrossRef]
- Ruiz-Meana, M.; Garcia-Dorado, D.; Hofstaetter, B.; Piper, H.M.; Soler-Soler, J. Propagation of cardiomyocyte hypercontracture by passage of Na(+) through gap junctions. Circ. Res. 1999, 85, 280–287. [Google Scholar] [CrossRef]
- Kojima, A.; Kitagawa, H.; Omatsu-Kanbe, M.; Matsuura, H.; Nosaka, S. Sevoflurane protects ventricular myocytes against oxidative stress-induced cellular Ca2+ overload and hypercontracture. Anesthesiology 2013, 119, 606–620. [Google Scholar] [CrossRef]
- Garcia-Dorado, D.; Ruiz-Meana, M.; Inserte, J.; Rodriguez-Sinovas, A.; Piper, H.M. Calcium-mediated cell death during myocardial reperfusion. Cardiovasc. Res. 2012, 94, 168–180. [Google Scholar] [CrossRef]
- Solares, J.; Garcia-Dorado, D.; Oliveras, J.; González, M.A.; Ruiz-Meana, M.; Barrabés, J.A.; Gonzalez-Bravo, C.; Soler-Soler, J. Contraction band necrosis at the lateral borders of the area at risk in reperfused infarcts. Observations in a pig model of in situ coronary occlusion. Virchows Arch. 1995, 426, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, G.; Kumar, T.K.; Tsai, C.C.; Srisailam, S.; Chou, S.H.; Ho, C.L.; Yu, C. Elucidation of the solution structure of cardiotoxin analogue V from the Taiwan cobra (Naja naja atra)-identification of structural features important for the lethal action of snake venom cardiotoxins. Protein Sci. 2000, 9, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Wong, K.Y.; Huang, L.-K.; Tan, K.Y.; Tan, N.H.; Wu, W.-g. Snake Venomics and Antivenomics of Cape Cobra (Naja nivea) from South Africa: Insights into Venom Toxicity and Cross-Neutralization Activity. Toxins 2022, 14, 860. [Google Scholar] [CrossRef] [PubMed]
- Deka, A.; Gogoi, A.; Das, D.; Purkayastha, J.; Doley, R. Proteomics of Naja kaouthia venom from North East India and assessment of Indian polyvalent antivenom by third generation antivenomics. J. Proteom. 2019, 207, 103463. [Google Scholar] [CrossRef] [PubMed]
- Cher, C.D.N.; Armugam, A.; Zhu, Y.Z.; Jeyaseelan, K. Molecular basis of cardiotoxicity upon cobra envenomation. Cell. Mol. Life Sci. 2005, 62, 105–118. [Google Scholar] [CrossRef]
- Fletcher, J.E.; Yang, C.C.; Rosenberg, P. Basic phospholipase A2 from Naja nigricollis snake venom: Phospholipid hydrolysis and effects on electrical and contractile activity of the rat heart. Toxicol. Appl. Pharmacol. 1982, 66, 39–54. [Google Scholar] [CrossRef]
- Karabuva, S.; Lukšić, B.; Brizić, I.; Latinović, Z.; Leonardi, A.; Križaj, I. Ammodytin L is the main cardiotoxic component of the Vipera ammodytes ammodytes venom. Toxicon 2017, 139, 94–100. [Google Scholar] [CrossRef]
- Greene, S.C.; Osborn, L.; Bower, R.; Harding, S.A.; Takenaka, K. Monocled Cobra (Naja kaouthia) Envenomations Requiring Mechanical Ventilation. J. Emerg. Med. 2021, 60, 197–201. [Google Scholar] [CrossRef]
- Kumar, T.K.; Jayaraman, G.; Lee, C.S.; Arunkumar, A.I.; Sivaraman, T.; Samuel, D.; Yu, C. Snake venom cardiotoxins-structure, dynamics, function and folding. J. Biomol. Struct. Dyn. 1997, 15, 431–463. [Google Scholar] [CrossRef]
- Ismail, A.K.; Weinstein, S.A.; Auliya, M.; Appareo, P. Ventricular bigeminy following a cobra envenomation. Clin. Toxicol. 2012, 50, 518–521. [Google Scholar] [CrossRef]
- John Binu, A.; Kumar Mishra, A.; Gunasekaran, K.; Iyadurai, R. Cardiovascular manifestations and patient outcomes following snake envenomation: A pilot study. Trop. Doct. 2019, 49, 10–13. [Google Scholar] [CrossRef]
- Le, H.Q.; Nguyen, N.T.T.; Vo, T.N.A.; van Nguyen, T.; Do, K.T.N.; Ho, T.T.C.; Nguyen, S.N.; Phan, X.T.; Nguyen, D.L.M.; Kieu, D.N.; et al. Envenoming by king cobras (Ophiophagus hannah) in Vietnam with cardiac complications and necrotizing fasciitis. Toxicon 2021, 200, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Alekseev, A.E.; Korystova, A.F.; Mavlyutova, D.A.; Kokoz, Y.M. Potential-dependent Ca2+ currents in isolated heart cells of hibernators. Biochem. Mol. Biol. Int. 1994, 33, 365–375. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Concentration | Parameter | ||
|---|---|---|---|---|
| Cells with Hypercontracture, % | Time to Hypercontracture, s | Time to Peak of Ca2+ Concentration, s | ||
| CT2No | 10 μg/mL | 100 | 393 ± 207 | 396 ± 217 |
| 25 μg/mL | 100 | 162 ± 86 | 152 ± 87 | |
| CT1Nk | 10 μg/mL | 100 | 503 ± 307 | 520 ± 318 |
| 25 μg/mL | 100 | 312 ± 148 | 299 ± 137 | |
| CT1No | 10 μg/mL | 24 ± 21 | 933 ± 707 | 909 ± 634 |
| 25 μg/mL | 89 ± 13 | 736 ± 397 | 747 ± 387 | |
| CT2Nh | 25 μg/mL | 18 ± 17 | 1328 ± 577 | 1255 ± 615 |
| 100 µg/mL | 100 | 531 ± 148 | 518 ± 163 | |
| CT1Nh | 25 μg/mL | 7 ± 12 | 1522 ± 177 | 1420 ± 223 |
| 100 µg/mL | 74 ± 29 | 1132 ± 342 | 1151 ± 295 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Averin, A.S.; Berezhnov, A.V.; Pimenov, O.Y.; Galimova, M.H.; Starkov, V.G.; Tsetlin, V.I.; Utkin, Y.N. Effects of Cobra Cardiotoxins on Intracellular Calcium and the Contracture of Rat Cardiomyocytes Depend on Their Structural Types. Int. J. Mol. Sci. 2023, 24, 9259. https://doi.org/10.3390/ijms24119259
Averin AS, Berezhnov AV, Pimenov OY, Galimova MH, Starkov VG, Tsetlin VI, Utkin YN. Effects of Cobra Cardiotoxins on Intracellular Calcium and the Contracture of Rat Cardiomyocytes Depend on Their Structural Types. International Journal of Molecular Sciences. 2023; 24(11):9259. https://doi.org/10.3390/ijms24119259
Chicago/Turabian StyleAverin, Alexey S., Alexey V. Berezhnov, Oleg Y. Pimenov, Miliausha H. Galimova, Vladislav G. Starkov, Victor I. Tsetlin, and Yuri N. Utkin. 2023. "Effects of Cobra Cardiotoxins on Intracellular Calcium and the Contracture of Rat Cardiomyocytes Depend on Their Structural Types" International Journal of Molecular Sciences 24, no. 11: 9259. https://doi.org/10.3390/ijms24119259
APA StyleAverin, A. S., Berezhnov, A. V., Pimenov, O. Y., Galimova, M. H., Starkov, V. G., Tsetlin, V. I., & Utkin, Y. N. (2023). Effects of Cobra Cardiotoxins on Intracellular Calcium and the Contracture of Rat Cardiomyocytes Depend on Their Structural Types. International Journal of Molecular Sciences, 24(11), 9259. https://doi.org/10.3390/ijms24119259