
Investigations on Primary Cilia of Nthy-ori 3-1 Cells upon Cysteine Cathepsin Inhibition or Thyrotropin Stimulation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
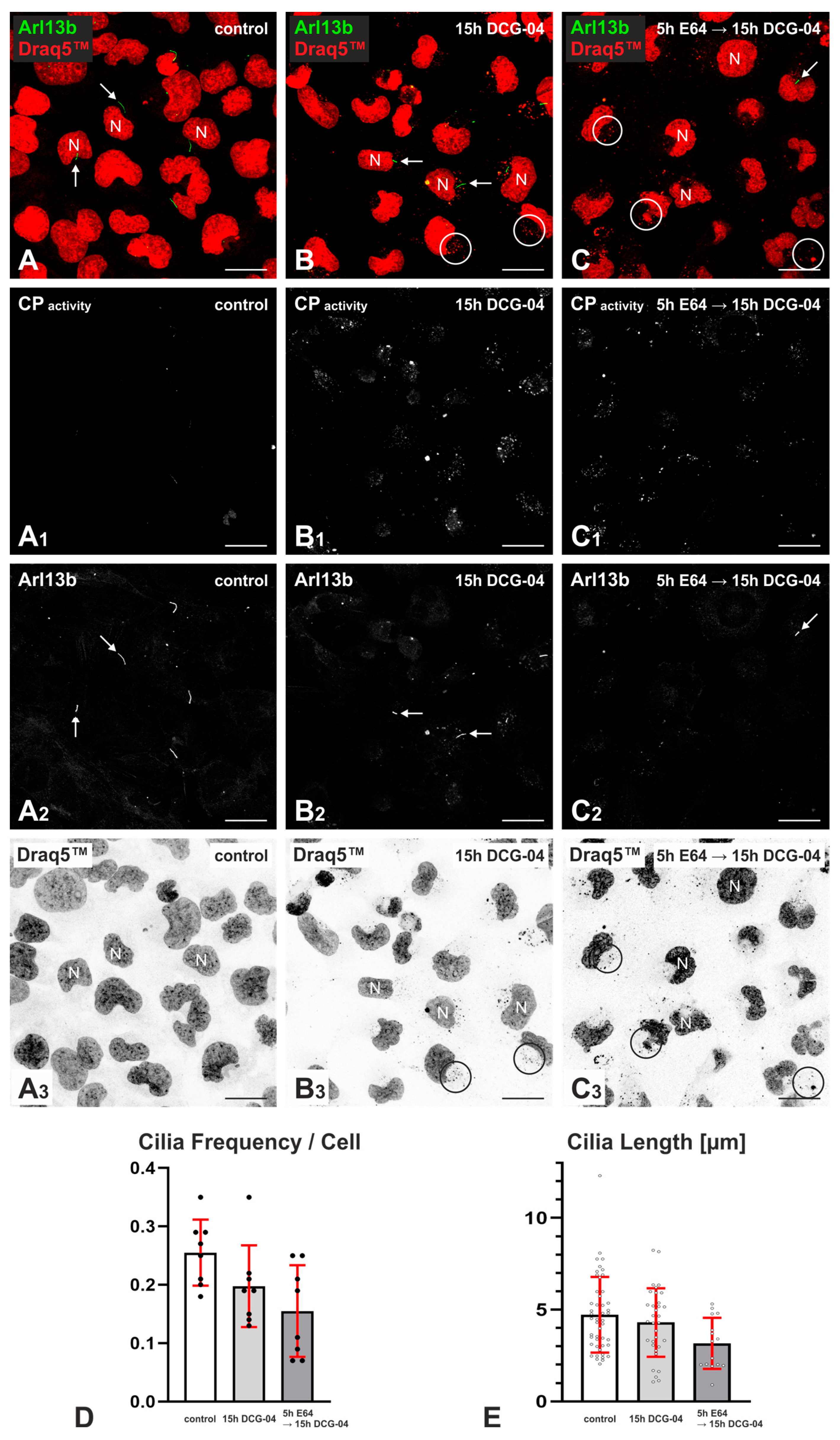
2.1. DCG-04 Labels Cysteine Peptidases and Verifies Successful Cysteine Peptidase Inhibition upon E64 Treatment of Nthy-ori 3-1 Cells
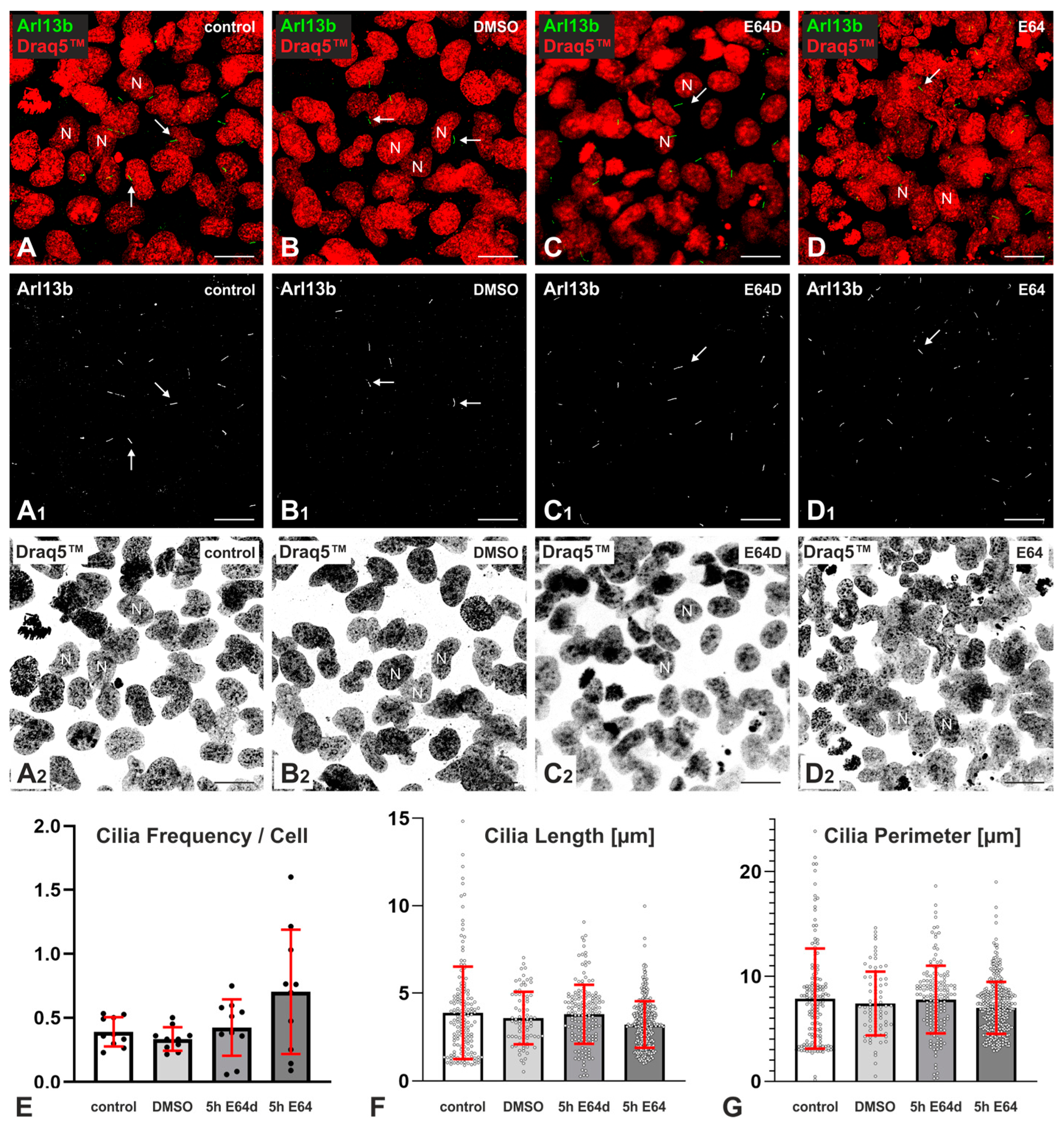
2.2. Effects of Cysteine Peptidase Inhibition on Frequency and Length of Cilia in Nthy-ori 3-1 Cells
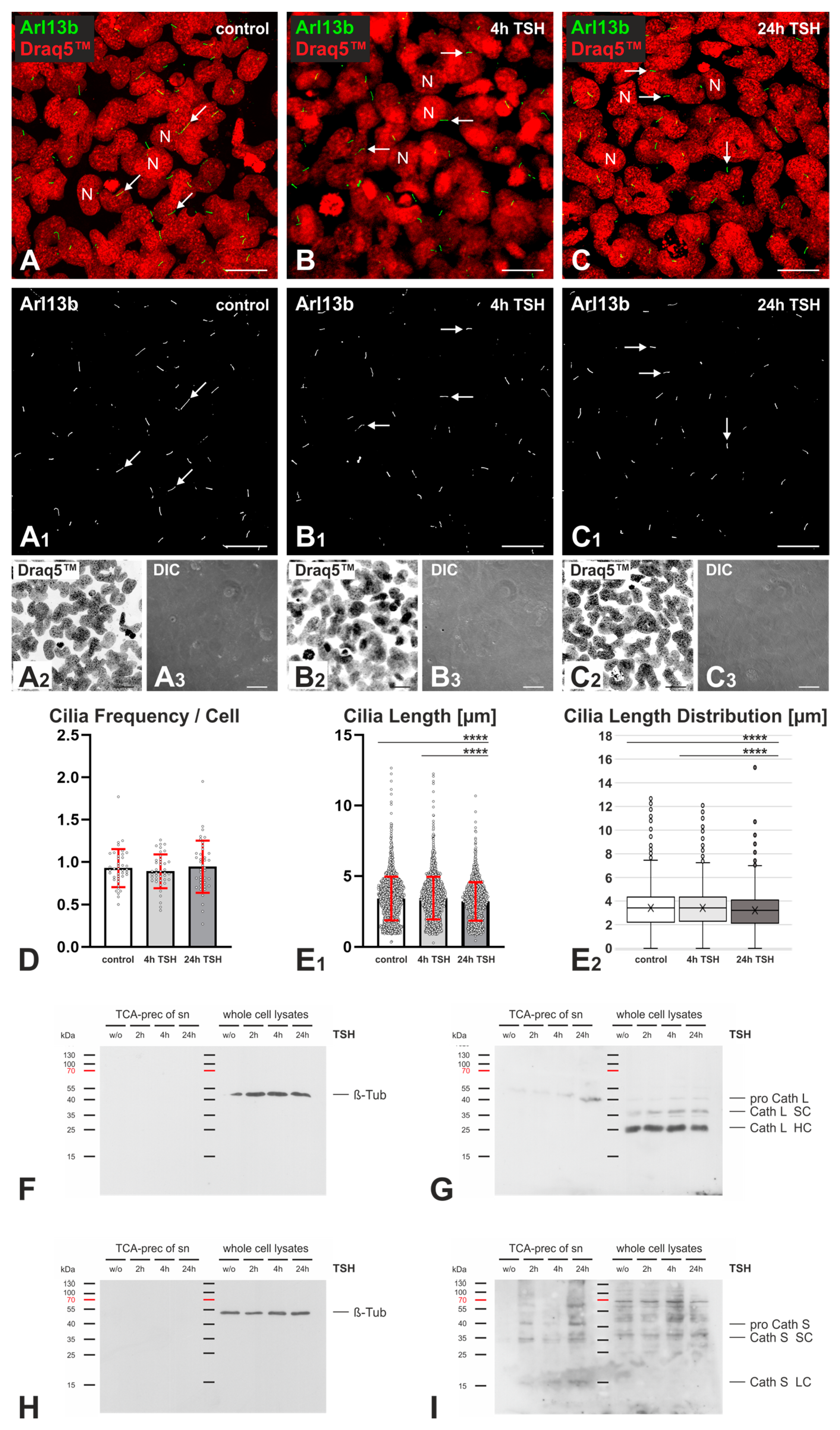
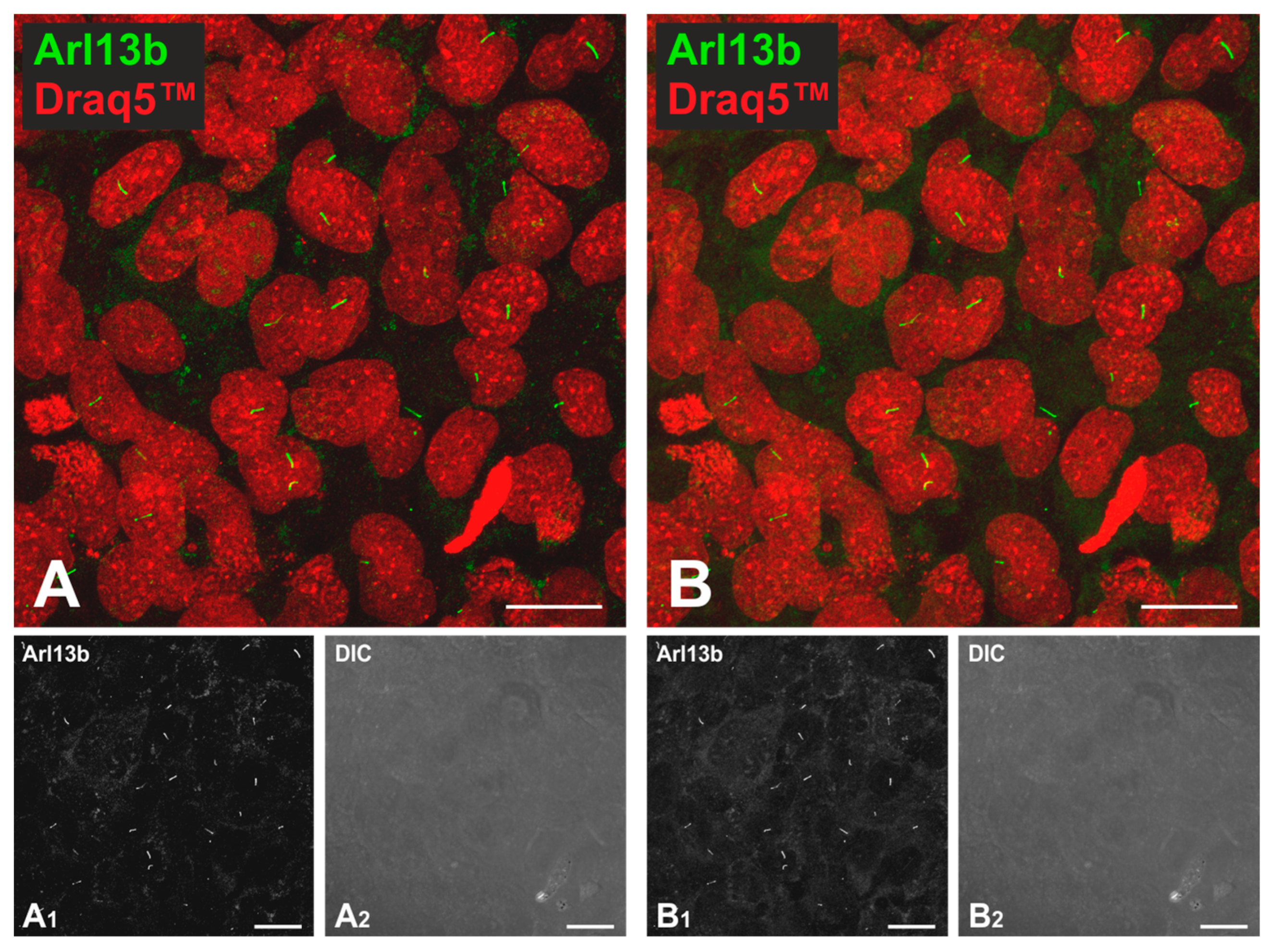
2.3. Effects of TSH Stimulation on Frequency and Length of Cilia in Nthy-ori 3-1 Cells
3. Discussion
3.1. Inhibition of Cysteine Cathepsins Affects Cilia Frequency and Length
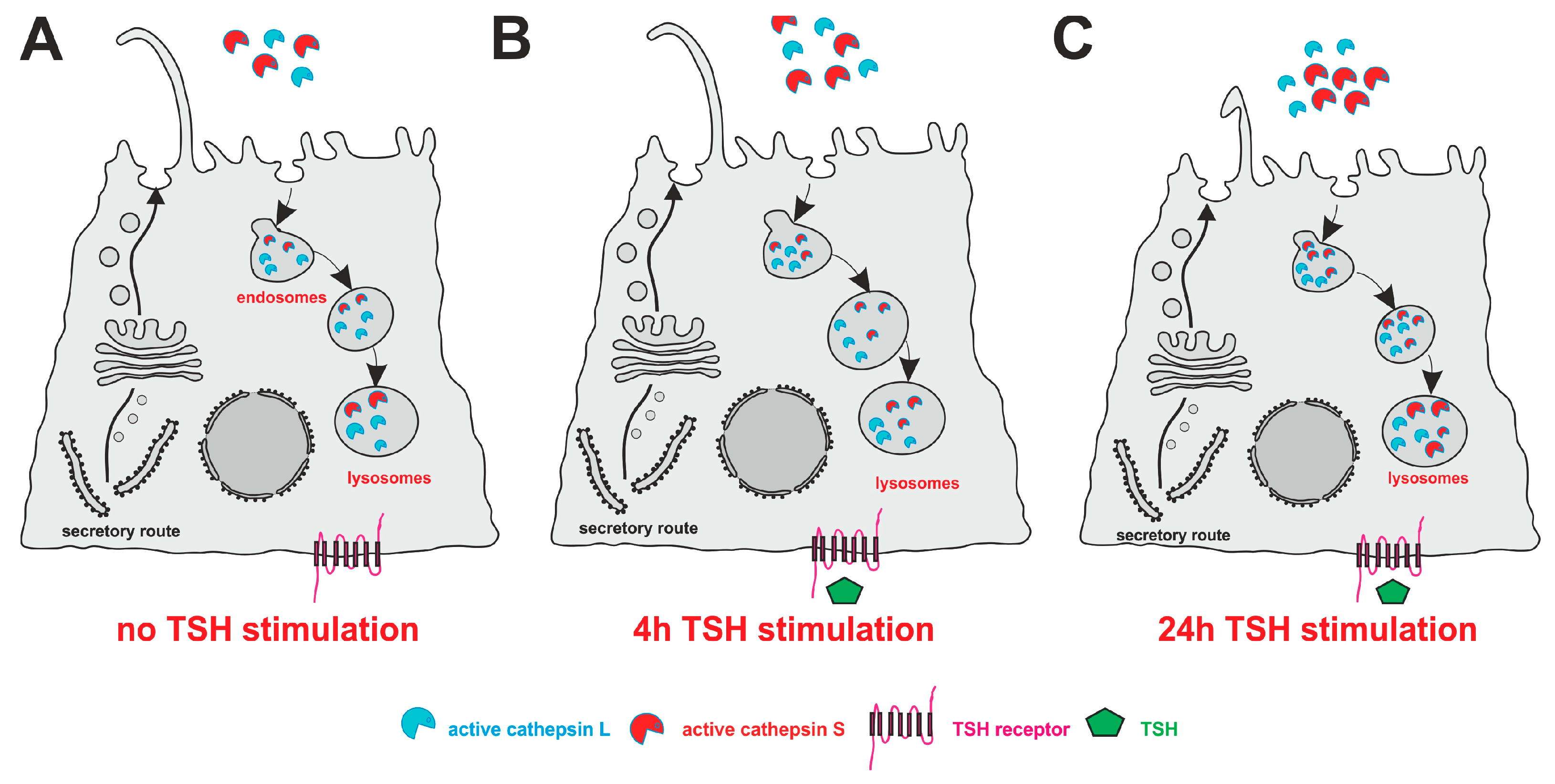
3.2. TSH Stimulation Affects Cilia Frequency and Length as Well as Cathepsin L and S Protein Amounts in Nthy-ori 3-1 Cell Cultures
4. Materials and Methods
4.1. Cell Culture
4.2. Protease Activity Inhibition Experiments
4.3. Activity-Based Probes
4.4. TCA Precipitation from Conditioned Media and Cell Lysate Preparation
4.5. SDS PAGE, Semi-Dry Transfer, and Immunoblotting
4.6. Indirect Immunofluorescence
4.7. Image Acquisition
4.8. Image Analysis with CellProfiler
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tedelind, S.; Poliakova, K.; Valeta, A.; Hunegnaw, R.; Yemanaberhan, E.; Heldin, N.; Kurebayashi, J.; Weber, E.; Kopitar-Jerala, N.; Turk, B.; et al. Nuclear cysteine cathepsin variants in thyroid carcinoma cells. Biol. Chem. 2010, 391, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Brix, K.; Dunkhorst, A.; Mayer, K.; Jordans, S. Cysteine cathepsins: Cellular roadmap to different functions. Biochimie 2008, 90, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Brix, K.; Szumska, J.; Weber, J.; Qatato, M.; Venugopalan, V.; Al-Hashimi, A.; Rehders, M. Auto-Regulation of the Thyroid Gland Beyond Classical Pathways. Exp. Clin. Endocrinol. Diabetes 2020, 128, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Mullur, R.; Liu, Y.; Brent, G. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed]
- Friedrichs, B.; Tepel, C.; Reinheckel, T.; Deussing, J.; von Figura, K.; Herzog, V.; Peters, C.; Saftig, P.; Brix, K. Thyroid functions of mouse cathepsins B, K, and L. J. Clin. Investig. 2003, 111, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Jordans, S.; Jenko-Kokalj, S.; Kühl, N.; Tedelind, S.; Sendt, W.; Brömme, D.; Turk, D.; Brix, K. Monitoring compartment-specific substrate cleavage by cathepsins B, K, L, and S at physiological pH and redox conditions. BMC Biochem. 2009, 10, 23. [Google Scholar] [CrossRef]
- Al-Hashimi, A.; Venugopalan, V.; Sereesongsaeng, N.; Tedelind, S.; Pinzaru, A.; Hein, Z.; Springer, S.; Weber, E.; Führer, D.; Scott, C.; et al. Significance of nuclear cathepsin V in normal thyroid epithelial and carcinoma cells. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118846. [Google Scholar] [CrossRef]
- Wheatley, D. The primary cilium—Once a “rudimentary” organelle that is now a ubiquitous sensory cellular structure involved in many pathological disorders. J. Cell Commun. Signal. 2017, 12, 211–216. [Google Scholar] [CrossRef]
- Fernández-Santos, J.; Utrilla, J.; Vázquez-Román, V.; Villar-Rodríguez, J.; Gutiérrez-Avilés, L.; Martín-Lacave, I. Primary Cilium in the Human Thyrocyte: Changes in Frequency and Length in Relation to the Functional Pathology of the Thyroid Gland. Thyroid 2019, 29, 595–606. [Google Scholar] [CrossRef]
- Szumska, J.; Batool, Z.; Al-Hashimi, A.; Venugopalan, V.; Skripnik, V.; Schaschke, N.; Bogyo, M.; Brix, K. Treatment of rat thyrocytes in vitro with cathepsin B and L inhibitors results in disruption of primary cilia leading to redistribution of the trace amine associated receptor 1 to the endoplasmic reticulum. Biochimie 2019, 166, 270–285. [Google Scholar] [CrossRef]
- Brix, K.; Qatato, M.; Szumska, J.; Venugopalan, V.; Rehders, M. Thyroglobulin Storage, Processing and Degradation for Thyroid Hormone Liberation. In The Thyroid and Its Diseases; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 25–48. [Google Scholar]
- Wheway, G.; Nazlamova, L.; Hancock, J. Signaling through the Primary Cilium. Front. Cell Dev. Biol. 2018, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Luo, N.; Conwell, M.; Chen, X.; Kettenhofen, C.; Westlake, C.; Cantor, L.; Wells, C.; Weinreb, R.; Corson, T.; Spandau, D.; et al. Primary cilia signaling mediates intraocular pressure sensation. Proc. Natl. Acad. Sci. USA 2014, 111, 12871–12876. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Sul, H.; Kim, K.; Chang, J.; Shong, M. Primary Cilia Mediate TSH-Regulated Thyroglobulin Endocytic Pathways. Front. Endocrinol. 2021, 12, 700083. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, U.; Mackiewicz, U.; Czarnocka, B. TSH Signal Transduction in Thyroid Cells of Nthy-Ori 3–1 Line. Postępy Nauk. Med. 2011, 11, 924–928. [Google Scholar]
- Greenbaum, D.; Medzihradszky, K.; Burlingame, A.; Bogyo, M. Epoxide electrophiles as activity-dependent cysteine protease profiling and discovery tools. Chem. Biol. 2000, 7, 569–581. [Google Scholar] [CrossRef]
- Carpenter, A.E.; Jones, T.R.; Lamprecht, M.R.; Clarke, C.; Kang., I.H.; Friman, O.; Guertin, D.A.; Chang, J.H.; Lindquist, R.A.; Moffat, J.; et al. CellProfiler: Image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 2006, 7, R100. [Google Scholar] [CrossRef]
- Linke, M.; Jordans, S.; Mach, L.; Herzog, V.; Brix, K. Thyroid Stimulating Hormone Upregulates Secretion of Cathepsin B from Thyroid Epithelial Cells. Biol. Chem. 2002, 383, 773–784. [Google Scholar] [CrossRef]
- Dickinson, D. Cysteine Peptidases of Mammals: Their Biological Roles and Potential Effects in the Oral Cavity and Other Tissues in Health and Disease. Crit. Rev. Oral Biol. Med. 2002, 13, 238–275. [Google Scholar] [CrossRef]
- Maher, K.; Kokelj, J.B.; Butinar, M.; Mikhaylov, G.; Manček-Keber, M.; Stoka, V.; Vasiljeva, O.; Turk, B.; Grigoryev, S.; Kopitar-Jerala, N. A Role for Stefin B (Cystatin B) in Inflammation and Endotoxemia. Biol. Chem. 2014, 289, 31736–31750. [Google Scholar] [CrossRef]
- Shirahama-Noda, K.; Yamamoto, A.; Sugihara, K.; Hashimoto, N.; Asano, M.; Nishimura, M.; Hara-Nishimura, I. Biosynthetic processing of cathepsins and lysosomal degradation are abolished in asparaginyl endopeptidase-deficient mice. Biol. Chem. 2003, 278, 33194–33199. [Google Scholar] [CrossRef]
- Dall, E.; Brandstetter, H. Structure and function of legumain in health and disease. Biochimie 2016, 122, 126–150. [Google Scholar] [CrossRef] [PubMed]
- Tepel, C.; Bromme, D.; Herzog, V.; Brix, K. Cathepsin K in thyroid epithelial cells: Sequence, localization and possible function in extracellular proteolysis of thyroglobulin. J. Cell Sci. 2000, 113, 4487–4498. [Google Scholar] [CrossRef] [PubMed]
- Dauth, S.; Sîrbulescu, R.F.; Jordans, S.; Rehders, M.; Avena, M.; Oswald, J.; Lerchl, A.; Saftig, P.; Brix, K. Cathepsin K deficiency in mice induces structural and metabolic changes in the central nervous system that are associated with learning and memory deficits. BMC Neurosci. 2011, 12, 74. [Google Scholar] [CrossRef] [PubMed]
- Rapoport, B.; Jones, A. Acute Effects of Thyroid-Stimulating Hormone on Cultured Thyroid Cell Morphology. Endocrinology 1978, 102, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Dumont, J.; Lamy, F.; Roger, P.; Maenhaut, C. Physiological and pathological regulation of thyroid cell proliferation and differentiation by thyrotropin and other factors. Physiol. Rev. 1992, 72, 667–697. [Google Scholar] [CrossRef]
- Ma, C.; Ma, X.; Li, Y.; Fu, S. The Role of Primary Cilia in Thyroid Cancer: From Basic Research to Clinical Applications. Front. Endocrinol. 2021, 12, 685228. [Google Scholar] [CrossRef]
- Petanceska, S.; Devi, L. Sequence analysis, tissue distribution, and expression of rat cathepsin S. J. Biol. Chem. 1992, 267, 26038–26043. [Google Scholar] [CrossRef]
- Al-Hashimi, A.; Venugopalan, V.; Rehders, M.; Sereesongsaeng, N.; Hein, Z.; Springer, S.; Weber, H.; Führer, D.; Bogyo, M.; Scott, C.; et al. Procathepsin V is Secreted in a TSH Regulated Manner from Human Thyroid Epithelial Cells and Is Accessible to an Activity-Based Probe. Int. J. Mol. Sci. 2020, 21, 9140. [Google Scholar] [CrossRef]
- Novinec, M.; Grass, R.N.; Stark, W.J.; Turk, V.; Baici, A.; Lenarčič, B. Mechanism of Elastinoysis and Inhibition by Macromolecular inhibitors. J. Biol. Chem. 2007, 282, 7893–7902. [Google Scholar] [CrossRef]
- Qatato, M.; Venugopalan, V.; Al-Hashimi, A.; Rehders, M.; Valentine, A.D.; Hein, Z.; Dallto, U.; Springer, S.; Brix, K. Trace Amine-Associated Receptor 1 Trafficking to Cilia of Thyroid Epithelial Cells. Cells 2021, 10, 1518. [Google Scholar] [CrossRef]
- Szumska, J.; Qatato, M.; Rehders, M.; Führer, D.; Biebermann, H.; Grandy, D.K.; Köhrle, J.; Brix, K. Trace Amine-Associated Receptor 1 Localization at the Apical Plasma Membrane Domain of Fisher Rat Thyroid Epithelial Cells is Confined to Cilia. Eur. Thyroid. J. 2015, 4, 30–41. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doğru, A.G.; Rehders, M.; Brix, K. Investigations on Primary Cilia of Nthy-ori 3-1 Cells upon Cysteine Cathepsin Inhibition or Thyrotropin Stimulation. Int. J. Mol. Sci. 2023, 24, 9292. https://doi.org/10.3390/ijms24119292
Doğru AG, Rehders M, Brix K. Investigations on Primary Cilia of Nthy-ori 3-1 Cells upon Cysteine Cathepsin Inhibition or Thyrotropin Stimulation. International Journal of Molecular Sciences. 2023; 24(11):9292. https://doi.org/10.3390/ijms24119292
Chicago/Turabian StyleDoğru, Alara Gaye, Maren Rehders, and Klaudia Brix. 2023. "Investigations on Primary Cilia of Nthy-ori 3-1 Cells upon Cysteine Cathepsin Inhibition or Thyrotropin Stimulation" International Journal of Molecular Sciences 24, no. 11: 9292. https://doi.org/10.3390/ijms24119292
APA StyleDoğru, A. G., Rehders, M., & Brix, K. (2023). Investigations on Primary Cilia of Nthy-ori 3-1 Cells upon Cysteine Cathepsin Inhibition or Thyrotropin Stimulation. International Journal of Molecular Sciences, 24(11), 9292. https://doi.org/10.3390/ijms24119292