
Evaluation of Radiosensitization and Cytokine Modulation by Differentially PEGylated Gold Nanoparticles in Glioblastoma Cells

 , , and
, , and 
Abstract
:1. Introduction
2. Results
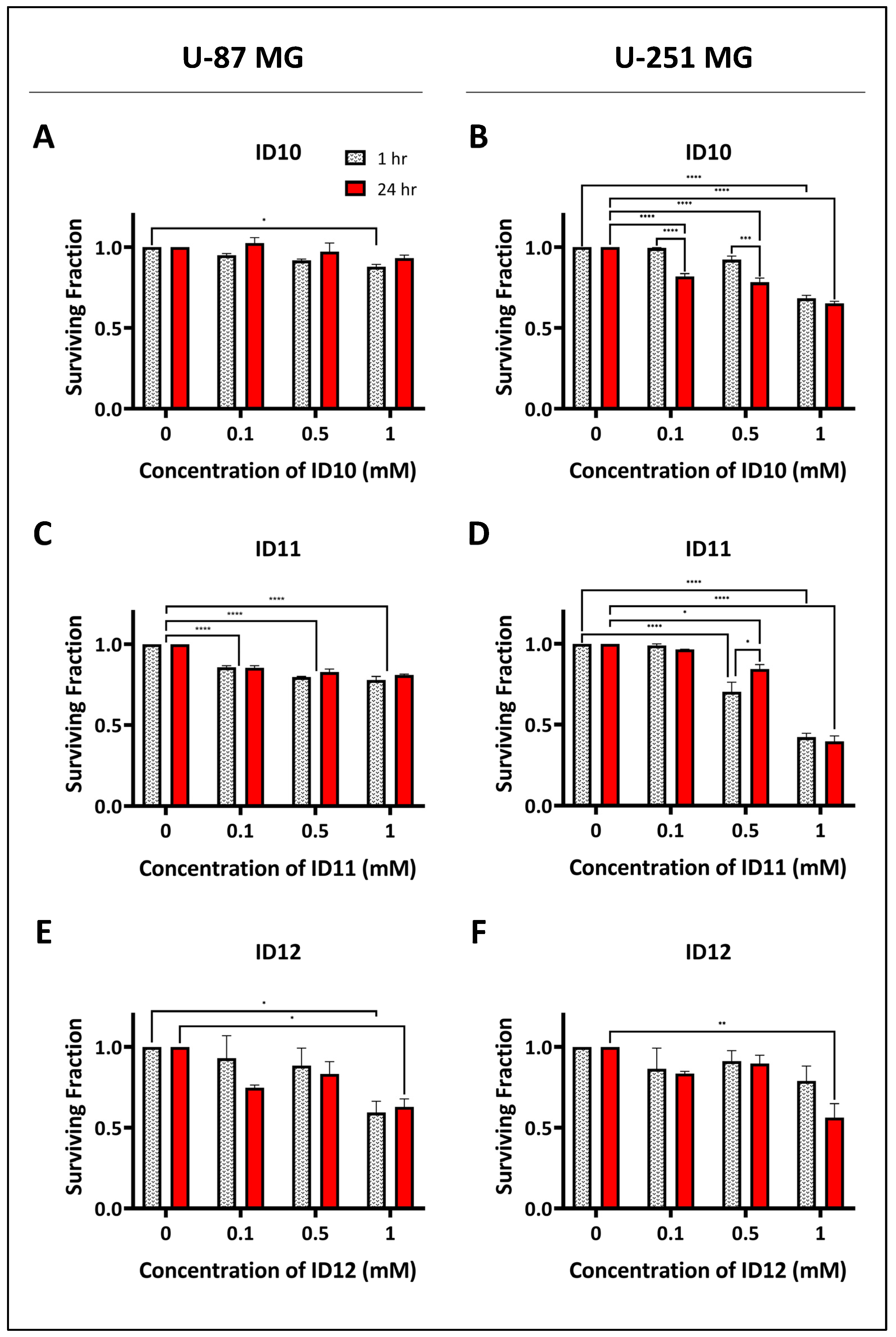
2.1. Evaluation of the Effects of GNPs on Cell Survival
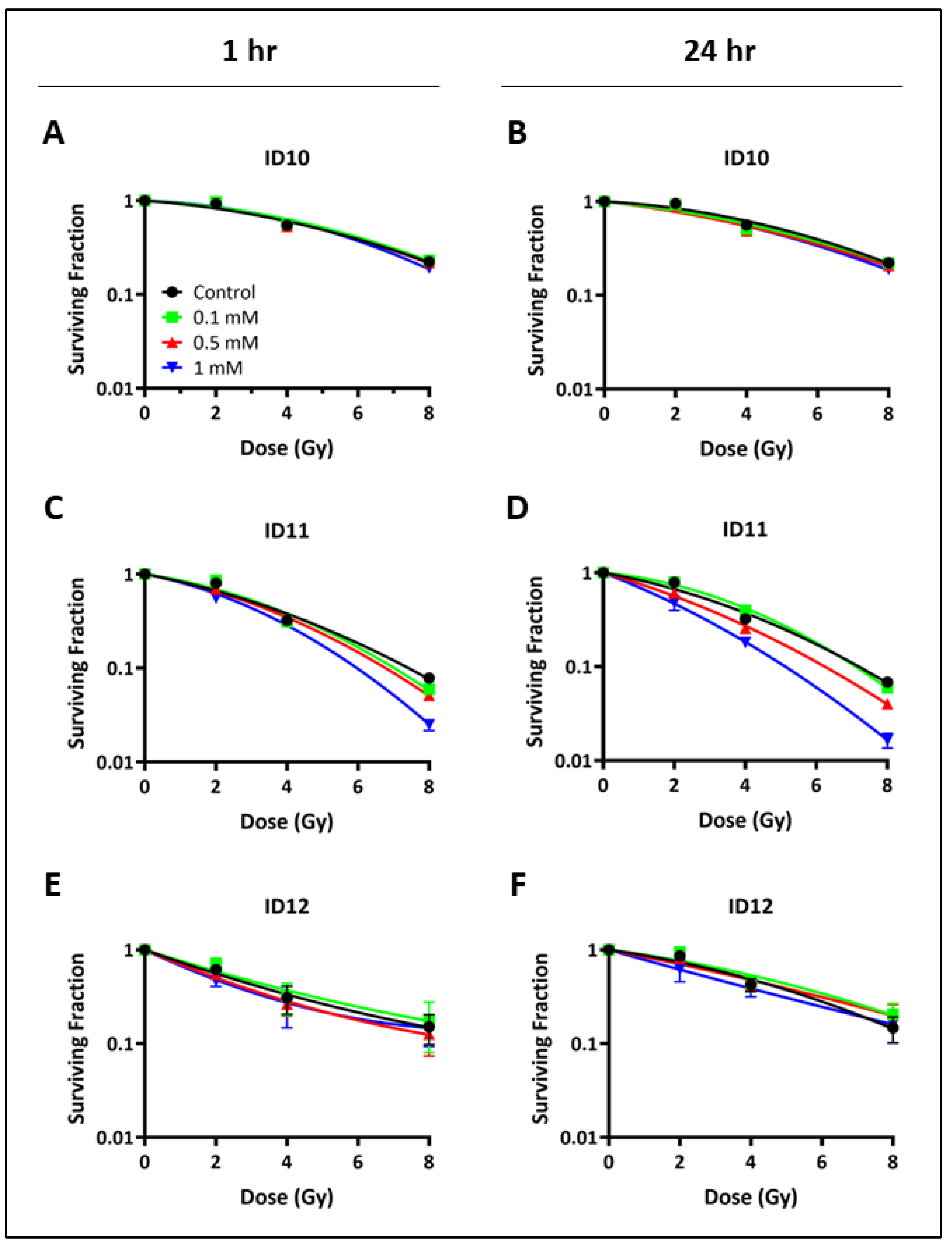
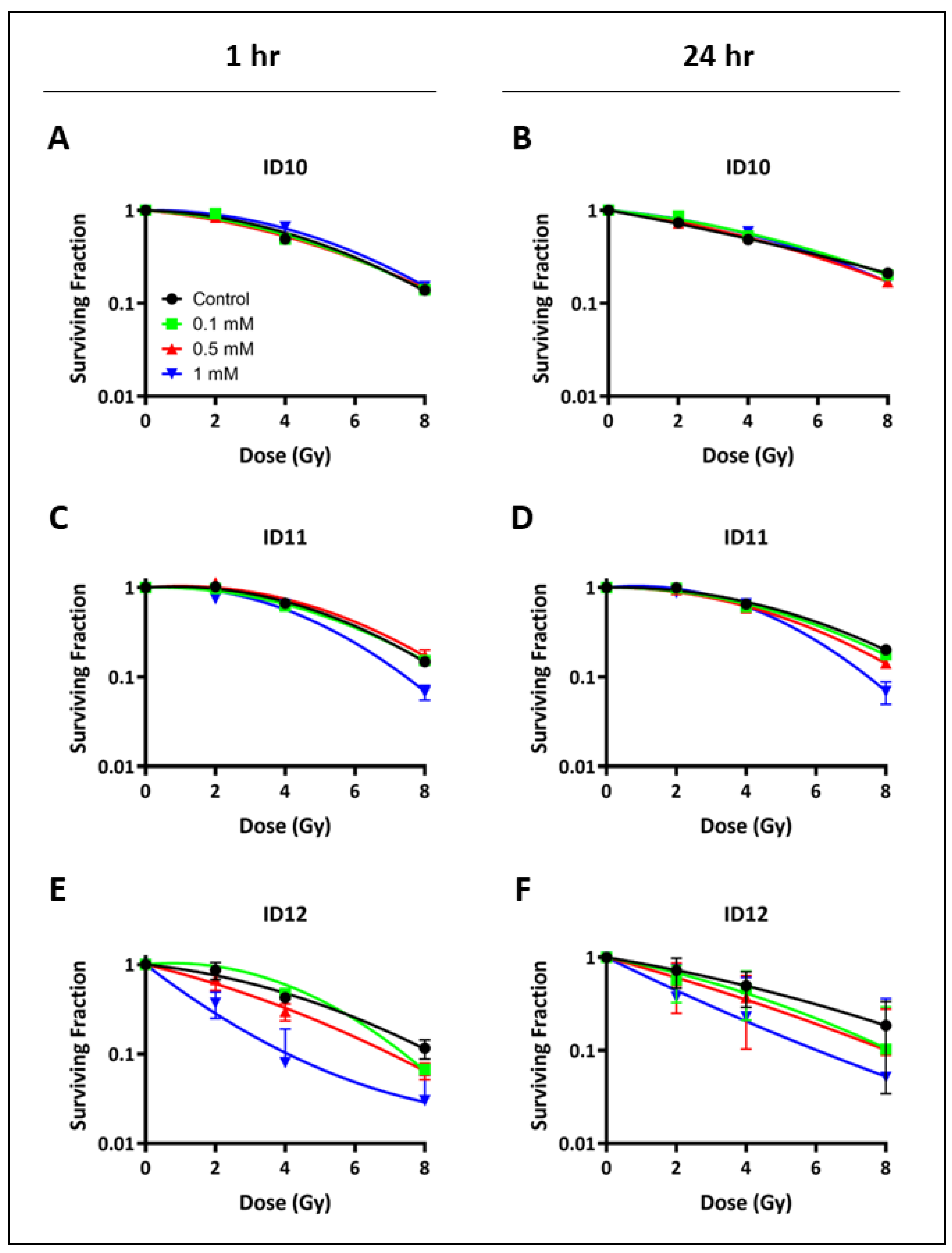
2.2. Impact on Clonogenic Cell Survival
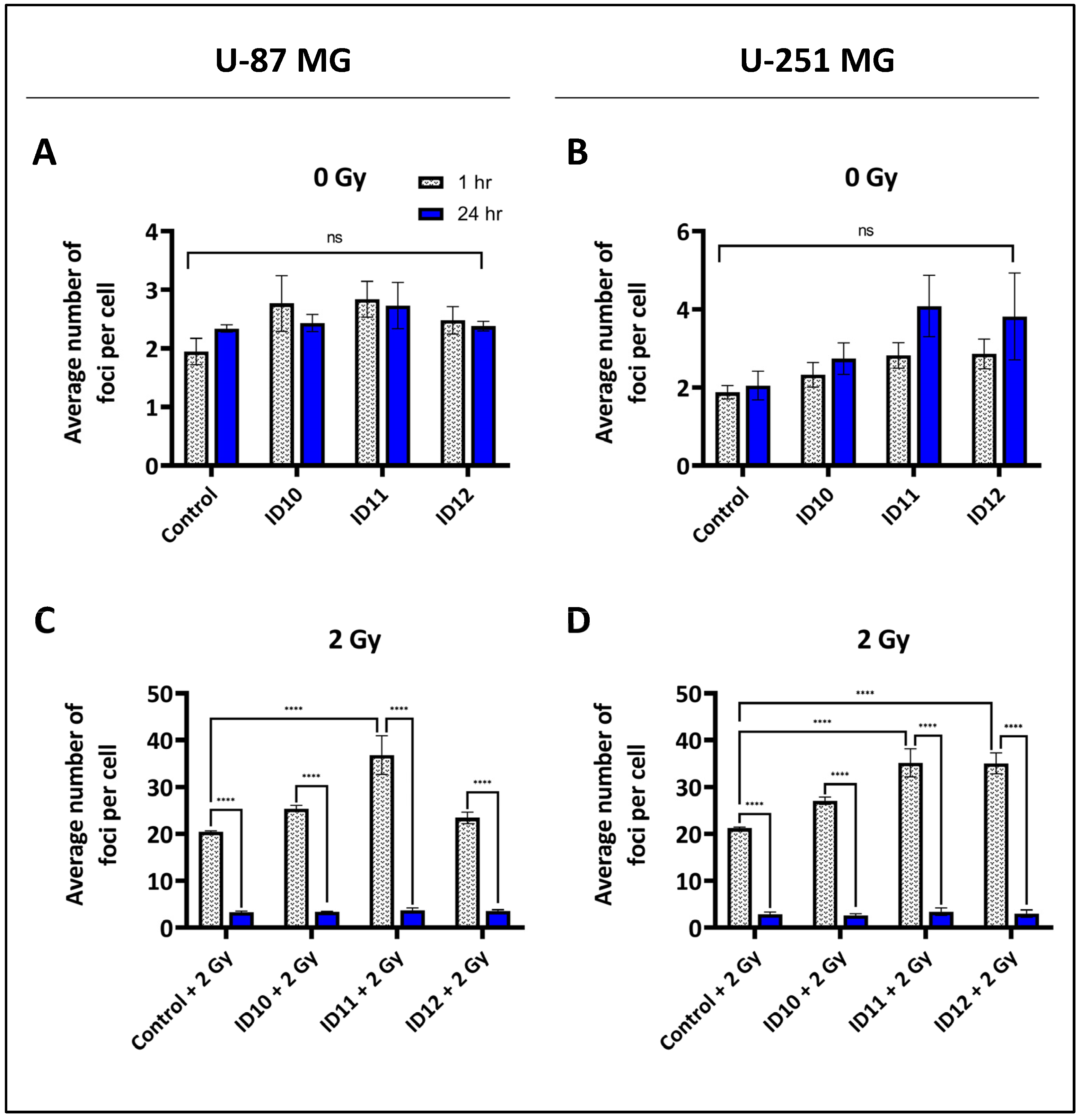
2.3. Evaluation of Double-Strand Break Induction and Repair
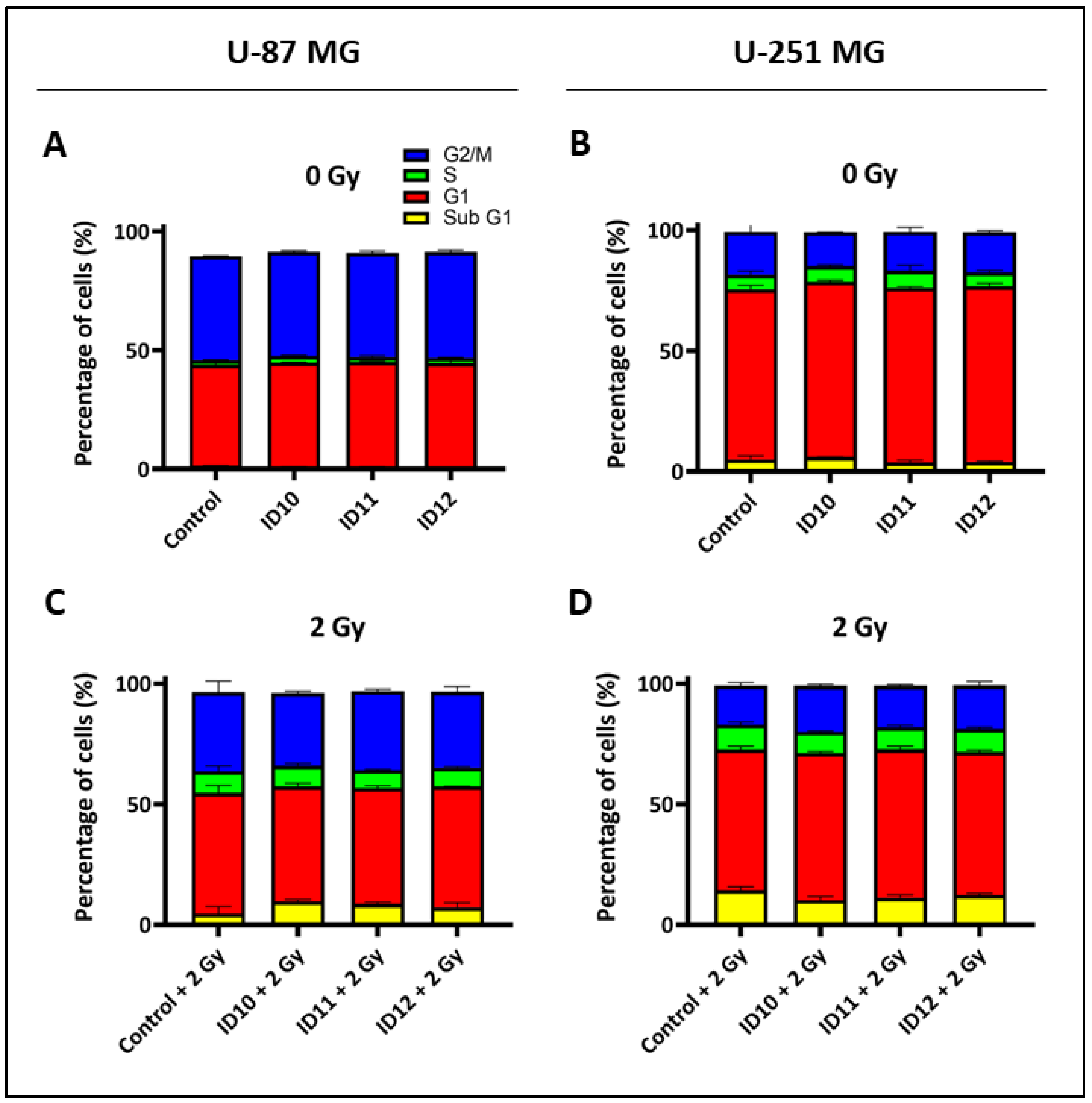
2.4. Evaluation of Cell Cycle Effects
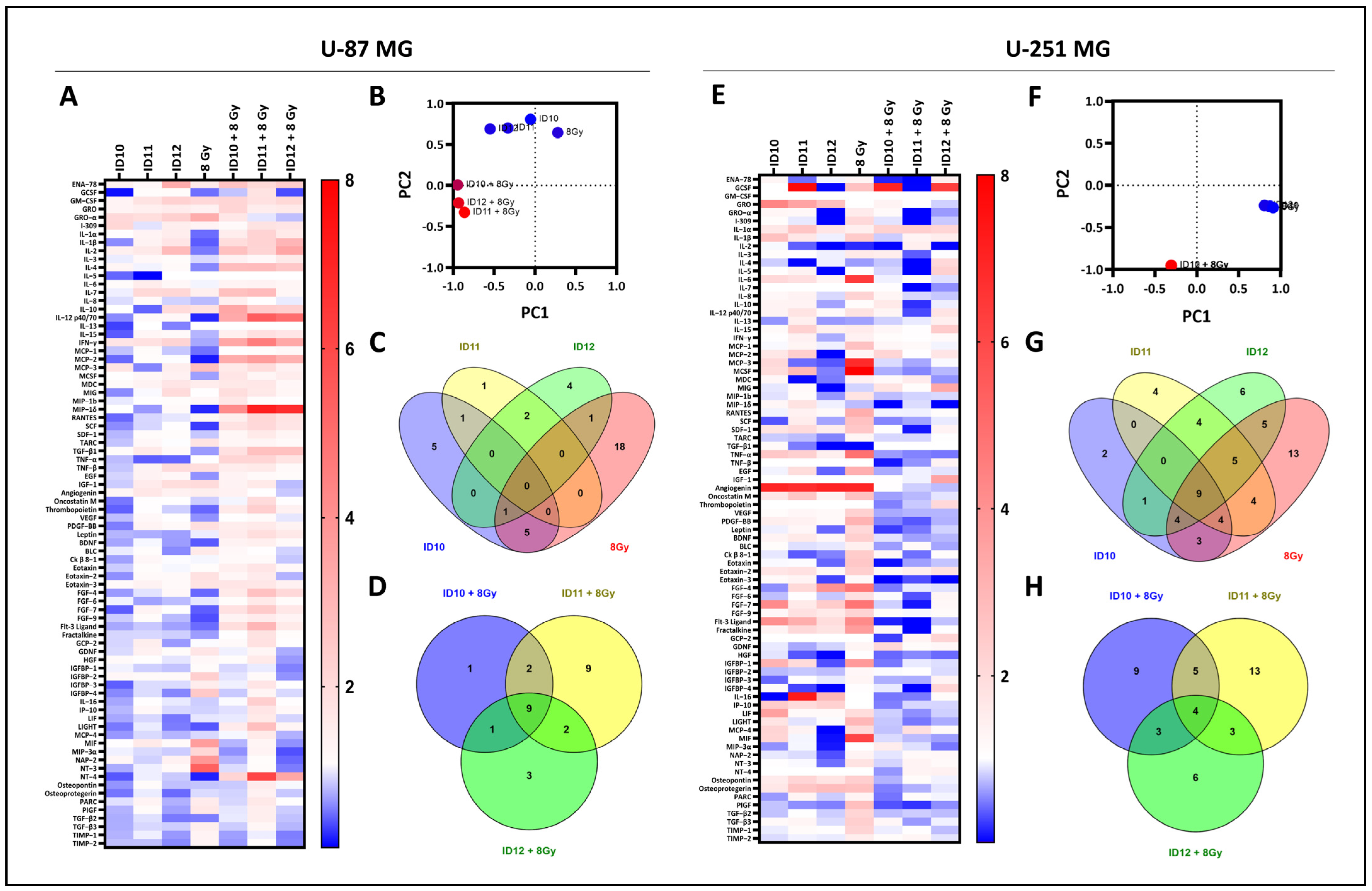
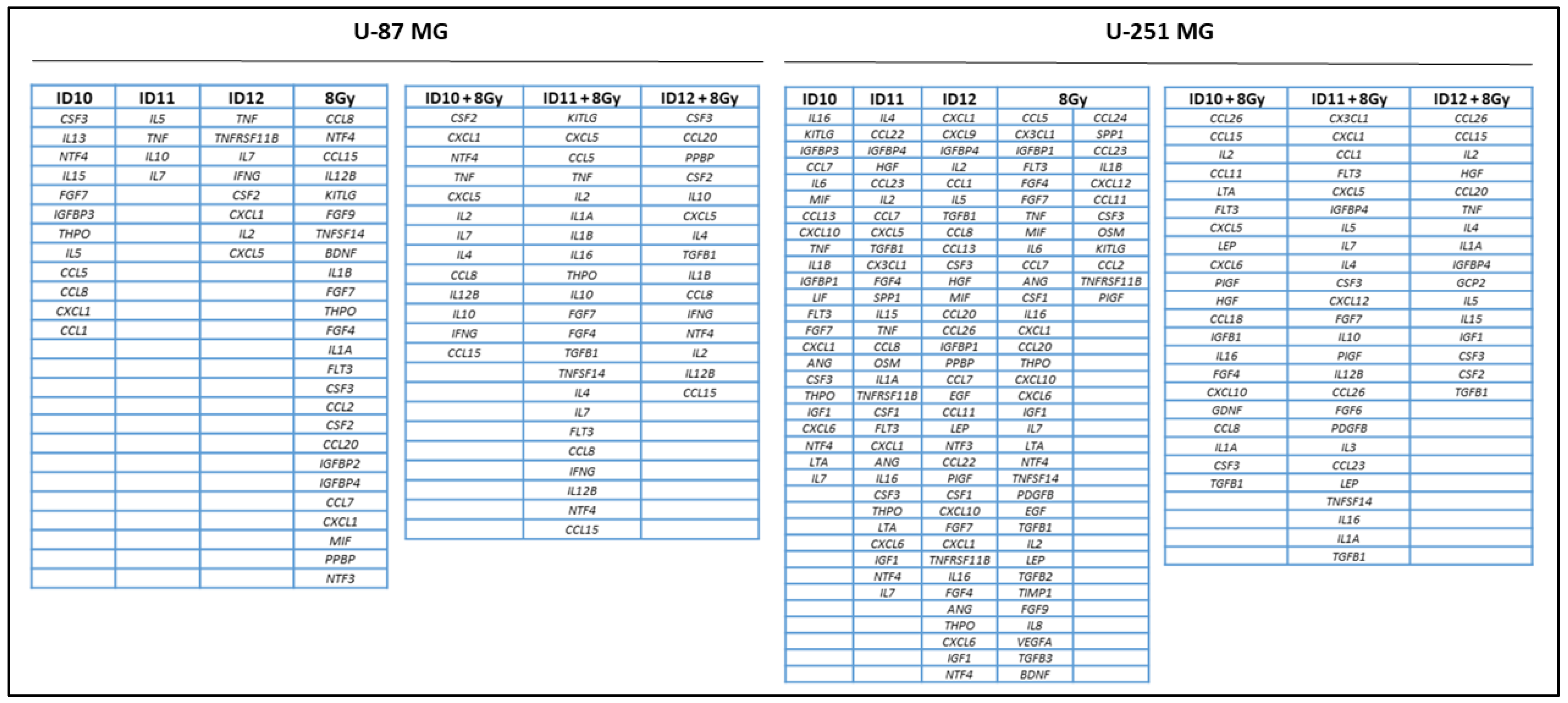
2.5. Comparison of Overall Cytokine Profiles and Differentially Expressed Cytokines
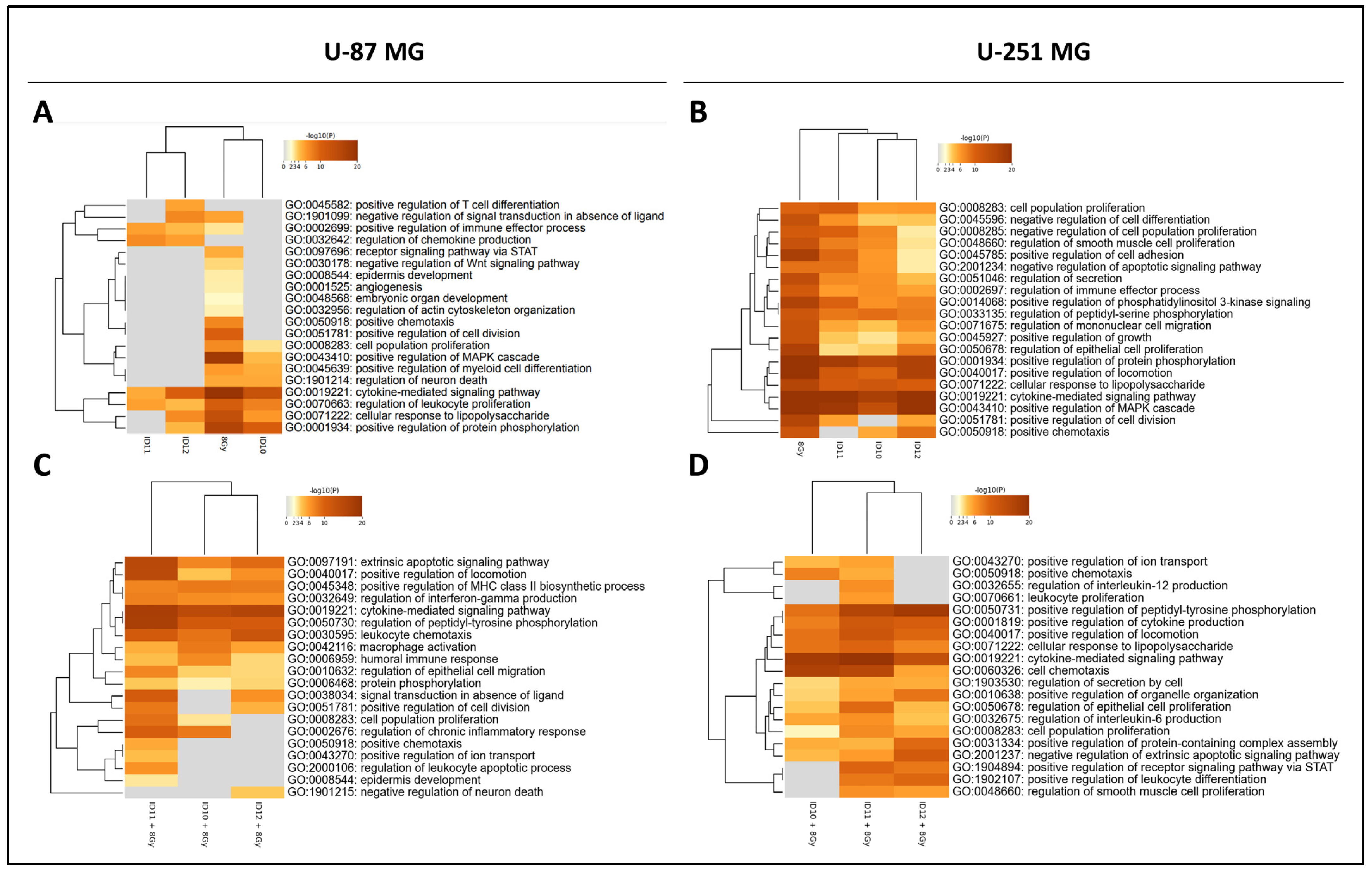
2.6. Pathway Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Synthesis and Physical Characterisation of GNPs (ID10, ID11 and ID12)
4.2. Cell Culture
4.3. Clonogenic Survival Assay, GNP Treatments, and In Vitro Irradiation Procedure
4.4. Analysis of DNA DSBs by Immunofluorescence Microscopy
4.5. Cell Cycle Analysis by Flow Cytometry
4.6. Human Cytokine Arrays and Bioinformatics Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Au@TADOTAGA (ID10) | Au@TAPEG4DOTAGA (ID11) | Au@TAPEG11DOTAGA (ID12) | |
|---|---|---|---|
| dcore (nm) | 2.5 ± 0.3 | 2.5 ± 0.3 | 2.5 ± 0.6 |
| Dh (nm) | 6.4 ± 2.3 | 6.4 ± 1.9 | 8.3 ± 1.7 |
| nDOTAGA | 85 | 71 | 87 |
| ζ (mV) | −24.0 | −33.9 | −16.1 |
References
- Grech, N.; Dalli, T.; Mizzi, S.; Meilak, L.; Calleja, N.; Zrinzo, A. Rising Incidence of Glioblastoma Multiforme in a Well-Defined Population. Cureus 2020, 12, e8195. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231. [Google Scholar] [CrossRef] [PubMed]
- Soomro, S.H.; Ting, L.R.; Qing, Y.Y.; Ren, M. Molecular biology of glioblastoma: Classification and mutational locations. J. Pak. Med. Assoc. 2017, 67, 1410–1414. [Google Scholar] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcaleavey, P.G.; Walls, G.M.; Chalmers, A.J. Radiotherapy-drug combinations in the treatment of glioblastoma: A brief review. CNS Oncol. 2022, 11, CNS86. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, K.T.; McMahon, S.J.; Taggart, L.E.; Prise, K.M. Radiosensitization by gold nanoparticles: Effective at megavoltage energies and potential role of oxidative stress. Transl. Cancer Res. 2013, 2, 269–279. [Google Scholar] [CrossRef]
- Butterworth, K.T.; Nicol, J.R.; Ghita, M.; Rosa, S.; Chaudhary, P.; McGarry, C.K.; McCarthy, H.O.; Jimenez-Sanchez, G.; Bazzi, R.; Roux, S.; et al. Preclinical evaluation of gold-DTDTPA nanoparticles as theranostic agents in prostate cancer radiotherapy. Nanomedicine 2016, 11, 2035–2047. [Google Scholar] [CrossRef] [Green Version]
- Bromma, K.; Cicon, L.; Beckham, W.; Chithrani, D.B. Gold nanoparticle mediated radiation response among key cell components of the tumour microenvironment for the advancement of cancer nanotechnology. Sci. Rep. 2020, 10, 12096. [Google Scholar] [CrossRef]
- Dorsey, J.F.; Sun, L.; Joh, D.Y.; Witztum, A.; Al Zaki, A.; Kao, G.D.; Alonso-Basanta, M.; Avery, S.; Hahn, S.M.; Al Zaki, A.; et al. Gold nanoparticles in radiation research: Potential applications for imaging and radiosensitization. Transl. Cancer Res. 2013, 2, 280–291. [Google Scholar] [CrossRef]
- Haume, K.; Rosa, S.; Grellet, S.; Śmiałek, M.A.; Butterworth, K.T.; Solov’yov, A.V.; Prise, K.M.; Golding, J.; Mason, N.J. Gold nanoparticles for cancer radiotherapy: A review. Cancer Nanotechnol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Butterworth, K.T.; McMahon, S.J.; Currell, F.J.; Prise, K.M. Physical basis and biological mechanisms of gold nanoparticle radiosensitization. Nanoscale 2012, 4, 4830–4838. [Google Scholar] [CrossRef]
- Tang, L.; Feng, Y.; Gao, S.; Mu, Q.; Liu, C. Nanotherapeutics Overcoming the Blood-Brain Barrier for Glioblastoma Treatment. Front. Pharmacol. 2021, 12, 3418. [Google Scholar] [CrossRef]
- Her, S.; Jaffray, D.A.; Allen, C. Gold nanoparticles for applications in cancer radiotherapy: Mechanisms and recent advancements. Adv. Drug Deliv. Rev. 2017, 109, 84–101. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, J.; Zhao, M.; Tang, S.; Cheng, X.; Zhang, W.; Li, W.; Liu, X.; Peng, H.; Wang, Q. Effects of polyethylene glycol on the surface of nanoparticles for targeted drug delivery. Nanoscale 2021, 13, 10748. [Google Scholar] [CrossRef]
- Pozzi, D.; Colapicchioni, V.; Caracciolo, G.; Piovesana, S.; Capriotti, A.L.; Palchetti, S.; De Grossi, S.; Riccioli, A.; Amenitsch, H.; Laganà, A. Effect of polyethyleneglycol (PEG) chain length on the bio-nano-interactions between PEGylated lipid nanoparticles and biological fluids: From nanostructure to uptake in cancer cells. Nanoscale 2014, 6, 2782–2792. [Google Scholar] [CrossRef]
- McMahon, S.J.; Paganetti, H.; Prise, K.M. Optimising element choice for nanoparticle radiosensitisers. Nanoscale 2016, 8, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.F.; Carter, T.J.; Ottaviani, D.; Mulholland, P. Harnessing the immune system in glioblastoma. Br. J. Cancer 2018, 119, 1171–1181. [Google Scholar] [CrossRef] [Green Version]
- Himes, B.T.; Geiger, P.A.; Ayasoufi, K.; Bhargav, A.G.; Brown, D.A.; Parney, I.F. Immunosuppression in Glioblastoma: Current Understanding and Therapeutic Implications. Front. Oncol. 2021, 11, 770561. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov; A Study Testing the Effect of Immunotherapy (Ipilimumab and Nivolumab) in Patients with Recurrent Glioma With Elevated Mutational Burden. Available online: https://www.clinicaltrials.gov/ct2/show/NCT04145115 (accessed on 31 August 2022).
- Prapa, M.; Chiavelli, C.; Golinelli, G.; Grisendi, G.; Bestagno, M.; Di Tinco, R.; Dall’ora, M.; Neri, G.; Candini, O.; Spano, C.; et al. GD2 CAR T cells against human glioblastoma. NPJ Precis. Oncol. 2021, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov; Vaccine Therapy in Treating Patients with Newly Diagnosed Glioblastoma Multiforme. Available online: https://clinicaltrials.gov/ct2/show/NCT00639639 (accessed on 31 August 2022).
- Tian, T.; Liang, R.; Erel-Akbaba, G.; Saad, L.; Obeid, P.J.; Gao, J.; Chiocca, E.A.; Weissleder, R.; Tannous, B.A. Immune Checkpoint Inhibition in GBM Primed with Radiation by Engineered Extracellular Vesicles. ACS Nano 2022, 16, 1940–1953. [Google Scholar] [CrossRef] [PubMed]
- Rückert, M.; Flohr, A.-S.; Hecht, M.; Gaipl, U.S. Radiotherapy and the immune system: More than just immune suppression. Stem Cells 2021, 39, 1155–1165. [Google Scholar] [CrossRef]
- Zhu, V.F.; Yang, J.; LeBrun, D.G.; Li, M. Understanding the role of cytokines in Glioblastoma Multiforme pathogenesis. Cancer Lett. 2012, 316, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Mozar, F.S.; Chowdhury, E.H. PEGylation of Carbonate Apatite Nanoparticles Prevents Opsonin Binding and Enhances Tumor Accumulation of Gemcitabine. J. Pharm. Sci. 2018, 107, 2497–2508. [Google Scholar] [CrossRef]
- Choi, K.Y.; Min, K.H.; Yoon, H.Y.; Kim, K.; Park, J.H.; Kwon, I.C.; Choi, K.; Jeong, S.Y. PEGylation of hyaluronic acid nanoparticles improves tumor targetability in vivo. Biomaterials 2011, 32, 1880–1889. [Google Scholar] [CrossRef]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a strategy for improving nanoparticle-based drug and gene delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef] [Green Version]
- El-Baz, N.; Nunn, B.M.; Bates, P.J.; O’Toole, M.G. The Impact of PEGylation on Cellular Uptake and In Vivo Biodistribution of Gold Nanoparticle MRI Contrast Agents. Bioengineering 2022, 9, 766. [Google Scholar] [CrossRef]
- Salvanou, E.A.; Stellas, D.; Tsoukalas, C.; Mavroidi, B.; Paravatou-petsotas, M.; Kalogeropoulos, N.; Xanthopoulos, S.; Denat, F.; Laurent, G.; Bazzi, R.; et al. A proof-of-concept study on the therapeutic potential of au nanoparticles radiolabeled with the alpha-emitter actinium-225. Pharmaceutics 2020, 12, 188. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Song, Z.; Zhang, T.; He, X.; Huang, K.; Zhang, Q.; Shen, J.; Pan, J. Identification of Immune Cell Infiltration and Immune-Related Genes in the Tumor Microenvironment of Glioblastomas. Front. Immunol. 2020, 11, 2708. [Google Scholar] [CrossRef]
- Basheer, A.S.; Abas, F.; Othman, I.; Naidu, R. Role of Inflammatory Mediators, Macrophages, and Neutrophils in Glioma Maintenance and Progression: Mechanistic Understanding and Potential Therapeutic Applications. Cancers 2021, 13, 4226. [Google Scholar] [CrossRef] [PubMed]
- Vauléon, E.; Tony, A.; Hamlat, A.; Etcheverry, A.; Chiforeanu, D.C.; Menei, P.; Mosser, J.; Quillien, V.; Aubry, M. Immune genes are associated with human glioblastoma pathology and patient survival. BMC Med. Genom. 2012, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrusiewicz, K.; Rodriguez, B.; Wei, J.; Hashimoto, Y.; Healy, L.M.; Maiti, S.N.; Thomas, G.; Zhou, S.; Wang, Q.; Elakkad, A.; et al. Glioblastoma-infiltrated innate immune cells resemble M0 macrophage phenotype. JCI Insight 2016, 1, 85841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.Z.; Zhang, Z.; Zheng, B.H.; Shi, Y.; Duan, M.; Ma, L.J.; Wang, Z.-C.; Dong, L.-Q.; Dong, P.-P.; Shi, J.-Y.; et al. CCL15 Recruits Suppressive Monocytes to Facilitate Immune Escape and Disease Progression in Hepatocellular Carcinoma. Hepatology 2019, 69, 143–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Chen, A.X.; Gartrell, R.D.; Silverman, A.M.; Aparicio, L.; Chu, T.; Bordbar, D.; Shan, D.; Samanamud, J.; Mahajan, A.; et al. Immune and genomic correlates of response to anti-PD-1 immunotherapy in glioblastoma. Nat. Med. 2019, 25, 462–469. [Google Scholar] [CrossRef]
- Sielska, M.; Przanowski, P.; Pasierbińska, M.; Wojnicki, K.; Poleszak, K.; Wojtas, B.; Grzeganek, D.; Ellert-Miklaszewska, A.; Ku, M.-C.; Kettenmann, H.; et al. Tumour-derived CSF2/granulocyte macrophage colony stimulating factor controls myeloid cell accumulation and progression of gliomas. Br. J. Cancer 2020, 123, 438–448. [Google Scholar] [CrossRef]
- Kast, R.E.; Hill, Q.A.; Wion, D.; Mellstedt, H.; Focosi, D.; Karpel-Massler, G.; Heiland, T.; Halatsch, M.-E. Glioblastoma-synthesized G-CSF and GM-CSF contribute to growth and immunosuppression: Potential therapeutic benefit from dapsone, fenofibrate, and ribavirin. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2017, 39, 1010428317699797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.; Young, H.; Hurlstone, A.; Wellbrock, C. Differentiation of THP1 Cells into Macrophages for Transwell Co-culture Assay with Melanoma Cells. Bio-Protocol 2015, 5, e1638. [Google Scholar] [CrossRef] [Green Version]
- Puck, T.T.; Marcus, P.I. Action of x-rays on mammalian cells. J. Exp. Med. 1956, 103, 653–666. [Google Scholar] [CrossRef]







| α/β | SF2 | SER | |||||
|---|---|---|---|---|---|---|---|
| GNP | Conc | 1 h | 24 h | 1 h | 24 h | 1 h | 24 h |
| (mM) | |||||||
| U-87 MG | |||||||
| ID10 | Control | 4.00 | 2.75 | 0.92 | 0.94 | ||
| 0.1 | 2.33 | 6.30 | 0.97 | 0.92 | 0.97 | 1.04 | |
| 0.5 | 2.90 | 8.59 | 0.98 | 0.89 | 0.99 | 1.08 | |
| 1 | 1.21 | 6.28 | 1 | 0.90 | 0.98 | 1.09 | |
| ID11 | Control | 8.37 | 6.78 | 0.80 | 0.78 | ||
| 0.1 | 4.40 | 2.37 | 0.86 | 0.79 | 0.99 | 0.94 | |
| 0.5 | 5.91 | 12.79 | 0.69 | 0.60 | 1.07 | 1.20 | |
| 1 | 4.56 | 14.95 | 0.55 | 0.47 | 1.21 | 1.45 | |
| ID12 | Control | 0 | 7.91 | 0.61 | 0.86 | ||
| 0.1 | 0 | 10.00 | 0.69 | 0.94 | 0.93 | 0.93 | |
| 0.5 | 0 | 39.07 | 0.54 | 0.81 | 1.10 | 1.00 | |
| 1 | 0 | 0 | 0.46 | 0.61 | 1.13 | 1.16 | |
| U-251 MG | |||||||
| ID10 | Control | 0.94 | 33.39 | 1.01 | 0.73 | ||
| 0.1 | 2.24 | 5.69 | 0.92 | 0.86 | 1.04 | 0.92 | |
| 0.5 | 3.96 | 8.90 | 0.84 | 0.73 | 1.07 | 1.00 | |
| 1 | 0 | 3.83 | 0.85 | 0.74 | 0.96 | 0.95 | |
| ID11 | Control | 0 | 0 | 1.02 | 0.99 | ||
| 0.1 | 0 | 0 | 0.97 | 0.98 | 1.05 | 1.04 | |
| 0.5 | 0 | 0 | 1.13 | 0.94 | 0.95 | 1.08 | |
| 1 | 0 | 0 | 0.74 | 0.87 | 1.17 | 1.10 | |
| ID12 | Control | 4.42 | 16.44 | 0.86 | 0.72 | ||
| 0.1 | 0 | 11.06 | 1.2 | 0.57 | 0.85 | 1.15 | |
| 0.5 | 13.82 | 38.91 | 0.67 | 0.56 | 1.25 | 1.25 | |
| 1 | 0 | 0 | 0.37 | 0.37 | 2.08 | 1.70 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerr, B.N.; Duffy, D.; McInerney, C.E.; Hutchinson, A.; Dabaja, I.; Bazzi, R.; Roux, S.; Prise, K.M.; Butterworth, K.T. Evaluation of Radiosensitization and Cytokine Modulation by Differentially PEGylated Gold Nanoparticles in Glioblastoma Cells. Int. J. Mol. Sci. 2023, 24, 10032. https://doi.org/10.3390/ijms241210032
Kerr BN, Duffy D, McInerney CE, Hutchinson A, Dabaja I, Bazzi R, Roux S, Prise KM, Butterworth KT. Evaluation of Radiosensitization and Cytokine Modulation by Differentially PEGylated Gold Nanoparticles in Glioblastoma Cells. International Journal of Molecular Sciences. 2023; 24(12):10032. https://doi.org/10.3390/ijms241210032
Chicago/Turabian StyleKerr, Bríanna N., Daniel Duffy, Caitríona E. McInerney, Ashton Hutchinson, Inaya Dabaja, Rana Bazzi, Stéphane Roux, Kevin M. Prise, and Karl T. Butterworth. 2023. "Evaluation of Radiosensitization and Cytokine Modulation by Differentially PEGylated Gold Nanoparticles in Glioblastoma Cells" International Journal of Molecular Sciences 24, no. 12: 10032. https://doi.org/10.3390/ijms241210032
APA StyleKerr, B. N., Duffy, D., McInerney, C. E., Hutchinson, A., Dabaja, I., Bazzi, R., Roux, S., Prise, K. M., & Butterworth, K. T. (2023). Evaluation of Radiosensitization and Cytokine Modulation by Differentially PEGylated Gold Nanoparticles in Glioblastoma Cells. International Journal of Molecular Sciences, 24(12), 10032. https://doi.org/10.3390/ijms241210032