
Placental AA/EPA Ratio Is Associated with Obesity Risk Parameters in the Offspring at 6 Years of Age

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Descriptive Characteristics of the Mother-Infant Pairs
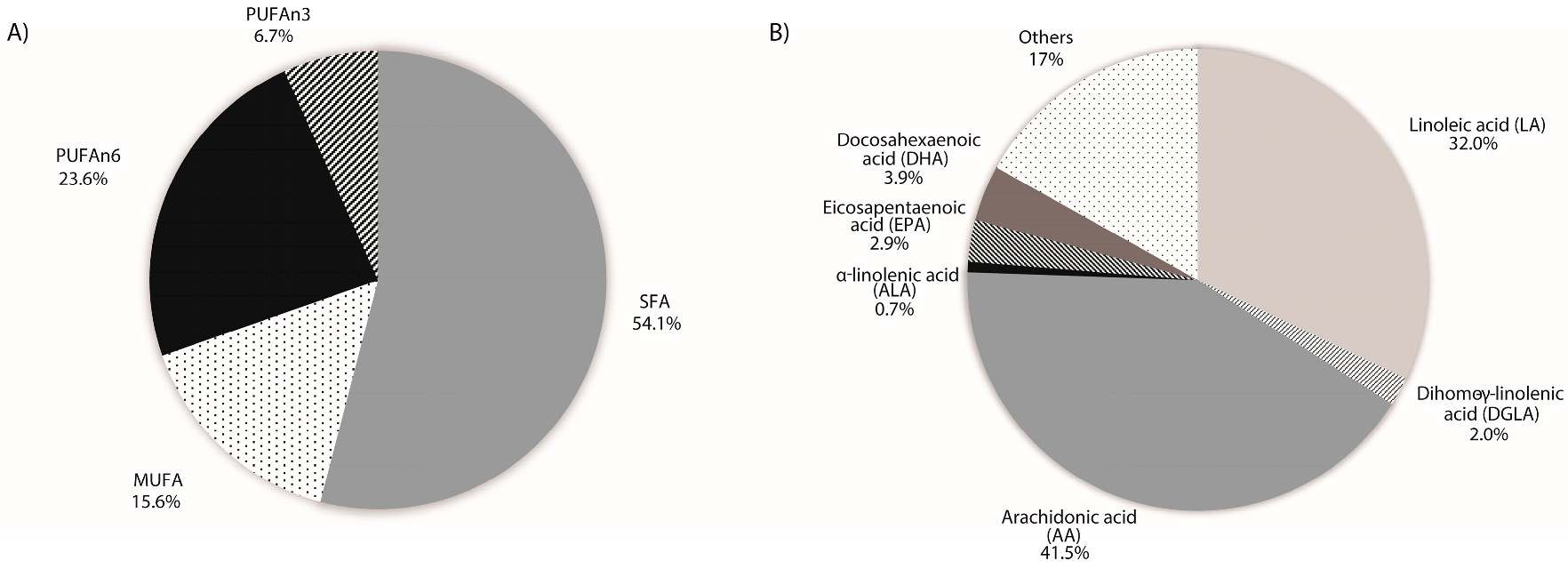
2.2. Placental Fatty Acid Profile
2.3. Placental Long Chain Polyunsaturated Fatty Acid (LC-PUFA) Abundance and Offspring Anthropometric and Metabolic Traits
2.4. Placental Fatty Acid Transporter Relative Expression
2.5. Placental Ʃ FATP Relative Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Study Population and Ethics
4.2. Maternal Anthropometric Assessments
4.3. Infant Anthropometric and Clinical Assessments at Delivery
4.4. Infant Anthropometric and Clinical Assessments at Six Year Follow-Up
4.5. Placental Sample Collection
4.6. Placental Fatty Acid Profile
4.7. Gene Expression Analysis
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EFA | essential fatty acid |
| FATP | fatty acid transporter |
| GWG | gestational weight gain |
| SDS | standard deviation score |
References
- Tvrzicka, E.; Kremmyda, L.S.; Stankova, B.; Zak, A. Fatty acids as biocompounds: Their role in human metabolism, health and disease—A review. Part 1: Classification, dietary sources and biological functions. Biomed. Pap. Med. Fac. Univ. Palacky. Olomouc. Czech. Repub. 2011, 155, 117–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumida, C.; Graber, R.; Nunez, E. Role of fatty acids in signal transduction: Modulators and messengers. Prostaglandins Leukot. Essent. Fat. Acids 1993, 48, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Marion-Letellier, R.; Savoye, G.; Ghosh, S. Polyunsaturated fatty acids and inflammation. IUBMB Life 2015, 67, 659–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Politano, C.A.; López-Berroa, J. Omega-3 Fatty Acids and Fecundation, Pregnancy and Breastfeeding. Rev. Bras. Ginecol. Obstet. 2020, 42, 160–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, R.; El Marroun, H.; Voortman, T.; Hillegers, M.; White, T.; Tiemeier, H. Maternal polyunsaturated fatty acids during pregnancy and offspring brain development in childhood. Am. J. Clin. Nutr. 2021, 114, 124–133. [Google Scholar] [CrossRef]
- Kapoor, B.; Kapoor, D.; Gautam, S.; Singh, R.; Bhardwaj, S. Dietary Polyunsaturated Fatty Acids (PUFAs): Uses and Potential Health Benefits. Curr. Nutr. Rep. 2021, 10, 232–242. [Google Scholar] [CrossRef]
- Wada, M.; DeLong, C.J.; Hong, Y.H.; Rieke, C.J.; Song, I.; Sidhu, R.S.; Yuan, C.; Warnock, M.; Schmaier, A.H.; Yokoyama, C.; et al. Enzymes and Receptors of Prostaglandin Pathways with Arachidonic Acid-derived versus Eicosapentaenoic Acid-derived Substrates and Products*. J. Biol. Chem. 2007, 282, 22254–22266. [Google Scholar] [CrossRef] [Green Version]
- Wiktorowska-Owczarek, A.; Berezińska, M.; Nowak, J.Z. PUFAs: Structures, Metabolism and Functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef]
- Blasbalg, T.L.; Hibbeln, J.R.; Ramsden, C.E.; Majchrzak, S.F.; Rawlings, R.R. Changes in consumption of omega-3 and omega-6 fatty acids in the United States during the 20th century. Am. J. Clin. Nutr. 2011, 93, 950–962. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Mark, P.J.; Waddell, B.J. Maternal dietary omega-3 fatty acids and placental function. Reproduction 2014, 147, R143–R152. [Google Scholar] [CrossRef] [Green Version]
- Akerele, O.A.; Cheema, S.K. A balance of omega-3 and omega-6 polyunsaturated fatty acids is important in pregnancy. J. Nutr. Intermed. Metab. 2016, 5, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Gude, N.M.; Roberts, C.T.; Kalionis, B.; King, R.G. Growth and function of the normal human placenta. Thromb. Res. 2004, 114, 397–407. [Google Scholar] [CrossRef]
- Haggarty, P. Effect of placental function on fatty acid requirements during pregnancy. Eur. J. Clin. Nutr. 2004, 58, 1559–1570. [Google Scholar] [CrossRef] [Green Version]
- Haggarty, P. Fatty Acid Supply to the Human Fetus. Annu. Rev. Nutr. 2010, 30, 237–255. [Google Scholar] [CrossRef]
- Hay, W. Metabolic interrelationships of placenta and fetus. Placenta 1995, 16, 19–30. [Google Scholar] [CrossRef]
- Jeppesen, J.; Jordy, A.B.; Sjøberg, K.A.; Füllekrug, J.; Stahl, A.; Nybo, L.; Kiens, B. Enhanced Fatty Acid Oxidation and FATP4 Protein Expression after Endurance Exercise Training in Human Skeletal Muscle. PLoS ONE 2012, 7, e29391. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Wang, N.; Peng, Y.; Sun, B.; Cui, W. Placental lipid transport and content in response to maternal overweight and gestational diabetes mellitus in human term placenta. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 692–702. [Google Scholar] [CrossRef]
- Donahue, S.M.; Rifas-Shiman, S.L.; Gold, D.R.; Jouni, Z.E.; Gillman, M.W.; Oken, E. Prenatal fatty acid status and child adiposity at age 3 y: Results from a US pregnancy cohort. Am. J. Clin. Nutr. 2011, 93, 780–788. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, M.C.; Young, B.E.; Lemas, D.J.; Palmer, C.E.; Hernandez, T.L.; Barbour, L.A.; Friedman, J.E.; Krebs, N.F.; MacLean, P.S. Early infant adipose deposition is positively associated with the n-6 to n-3 fatty acid ratio in human milk independent of maternal BMI. Int. J. Obes. 2017, 41, 510–517. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Maekawa, R.; Patterson, N.E.; Reynolds, D.M.; Calder, B.R.; Reznik, S.E.; Heo, H.J.; Einstein, F.H.; Greally, J.M. Amnion as a surrogate tissue reporter of the effects of maternal preeclampsia on the fetus. Clin. Epigenetics 2016, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Tsugane, S. Why has Japan become the world’s most long-lived country: Insights from a food and nutrition perspective. Eur. J. Clin. Nutr. 2021, 75, 921–928. [Google Scholar] [CrossRef]
- Christian, L.M.; Blair, L.M.; Porter, K.; Lower, M.; Cole, R.M.; Belury, M.A. Polyunsaturated Fatty Acid (PUFA) Status in Pregnant Women: Associations with Sleep Quality, Inflammation, and Length of Gestation. PLoS ONE 2016, 11, e0148752. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, M.H.; Nielsen, P.K.; Michaelsen, K.F.; Lund, P.; Lauritzen, L. The composition of polyunsaturated fatty acids in erythrocytes of lactating mothers and their infants. Matern. Child Nutr. 2006, 2, 29–39. [Google Scholar] [CrossRef]
- Dutta-Roy, A.K. Transport mechanisms for long-chain polyunsaturated fatty acids in the human placenta. Am. J. Clin. Nutr. 2000, 71, 315S–322S. [Google Scholar] [CrossRef] [Green Version]
- Haggarty, P. Placental Regulation of Fatty Acid Delivery and its Effect on Fetal Growth—A Review. Placenta 2002, 23, S28–S38. [Google Scholar] [CrossRef]
- Muhlhausler, B.S.; Ailhaud, G.P. Omega-6 polyunsaturated fatty acids and the early origins of obesity. Curr. Opin. Endocrinol. Diabetes 2013, 20, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Naughton, S.S.; Mathai, M.L.; Hryciw, D.H.; McAinch, A.J. Linoleic acid and the pathogenesis of obesity. Prostaglandins Other Lipid Mediat. 2016, 125, 90–99. [Google Scholar] [CrossRef]
- Vioque, J.; Navarrete-Muñoz, E.-M.; Gimenez-Monzó, D.; García-De-La-Hera, M.; Granado, F.; Young, I.S.; Ramón, R.; Ballester, F.; Murcia, M.; Rebagliato, M.; et al. Reproducibility and validity of a food frequency questionnaire among pregnant women in a Mediterranean area. Nutr. J. 2013, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, H.; Cheema, S.K. Breastmilk with a high omega-6 to omega-3 fatty acid ratio induced cellular events similar to insulin resistance and obesity in 3T3-L1 adipocytes. Pediatr. Obes. 2018, 13, 285–291. [Google Scholar] [CrossRef]
- Ailhaud, G.; Guesnet, P.; Cunnane, S.C. An emerging risk factor for obesity: Does disequilibrium of polyunsaturated fatty acid metabolism contribute to excessive adipose tissue development? Br. J. Nutr. 2008, 100, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Kothapalli, K.S.; Brenna, J.T. Desaturase and elongase-limiting endogenous long-chain polyunsaturated fatty acid biosynthesis. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 103–110. [Google Scholar] [CrossRef]
- Takic, M.; Pokimica, B.; Petrovic-Oggiano, G.; Popovic, T. Effects of Dietary α-Linolenic Acid Treatment and the Efficiency of Its Conversion to Eicosapentaenoic and Docosahexaenoic Acids in Obesity and Related Diseases. Molecules 2022, 27, 4471. [Google Scholar] [CrossRef]
- Aparicio, E.; Martín-Grau, C.; Bedmar, C.; Orus, N.S.; Basora, J.; Arija, V. Maternal factors associated with levels of fatty acids, specifically n-3 pufa during pregnancy: Eclipses study. Nutrients 2021, 13, 317. [Google Scholar] [CrossRef]
- Hao, Y.; Sun, X.; Wen, N.; Song, D.; Li, H. Effects of n-3 polyunsaturated fatty acid supplementation on pregnancy outcomes: A systematic review and meta-analysis. Arch. Med Sci. 2021, 18, 890. [Google Scholar] [CrossRef]
- Mori, T.A.; Woodman, R. The independent effects of eicosapentaenoic acid and docosahexaenoic acid on cardiovascular risk factors in humans. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 95–104. [Google Scholar] [CrossRef]
- Brinton, E.A.; Mason, R.P. Prescription omega-3 fatty acid products containing highly purified eicosapentaenoic acid (EPA). Lipids Health Dis. 2017, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, H.; Saito, Y. Eicosapentaenoic Acid (EPA) Reduces Cardiovascular Events: Relationship with the EPA/Arachidonic Acid Ratio. J. Atheroscler. Thromb. 2013, 20, 861–877. [Google Scholar] [CrossRef] [Green Version]
- Ninomiya, T.; Nagata, M.; Hata, J.; Hirakawa, Y.; Ozawa, M.; Yoshida, D.; Ohara, T.; Kishimoto, H.; Mukai, N.; Fukuhara, M.; et al. Association between ratio of serum eicosapentaenoic acid to arachidonic acid and risk of cardiovascular disease: The Hisayama Study. Atherosclerosis 2013, 231, 261–267. [Google Scholar] [CrossRef]
- Shojima, Y.; Ueno, Y.; Tanaka, R.; Yamashiro, K.; Miyamoto, N.; Hira, K.; Kurita, N.; Nakajima, S.; Urabe, T.; Hattori, N. Eicosapentaenoic-to-Arachidonic Acid Ratio Predicts Mortality and Recurrent Vascular Events in Ischemic Stroke Patients. J. Atheroscler. Thromb. 2020, 27, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Ailhaud, G.; Massiera, F.; Weill, P.; Legrand, P.; Alessandri, J.; Guesnet, P. Temporal changes in dietary fats: Role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog. Lipid Res. 2006, 45, 203–236. [Google Scholar] [CrossRef]
- Woodard, V.; Thoene, M.; Van Ormer, M.; Thompson, M.; Hanson, C.; Natarajan, S.K.; Mukherjee, M.; Yuil-Valdes, A.; Nordgren, T.M.; Ulu, A.; et al. Intrauterine Transfer of Polyunsaturated Fatty Acids in Mother–Infant Dyads as Analyzed at Time of Delivery. Nutrients 2021, 13, 996. [Google Scholar] [CrossRef]
- Garcia-Santillan, J.A.; Rodriguez-Saldaña, G.C.; Solis-Barbosa, M.A.; Corona-Figueroa, M.A.; Gonzalez-Dominguez, M.I.; Gomez-Zapata, H.M.; Malacara, J.M.; Barbosa-Sabanero, G. Placental Nutrient Transporters and Maternal Fatty Acids in SGA, AGA, and LGA Newborns from Mothers with and Without Obesity. Front. Cell Dev. Biol. 2022, 10, 25. [Google Scholar] [CrossRef]
- Segura, M.T.; Demmelmair, H.; Krauss-Etschmann, S.; Nathan, P.; Dehmel, S.; Padilla, M.C.; Rueda, R.; Koletzko, B.; Campoy, C. Maternal BMI and gestational diabetes alter placental lipid transporters and fatty acid composition. Placenta 2017, 57, 144–151. [Google Scholar] [CrossRef]
- Song, Y.-P.; Chen, Y.-H.; Gao, L.; Wang, P.; Wang, X.-L.; Luo, B.; Li, J.; Xu, D.-X. Differential effects of high-fat diets before pregnancy and/or during pregnancy on fetal growth development. Life Sci. 2018, 212, 241–250. [Google Scholar] [CrossRef]
- Larqué, E.; Krauss-Etschmann, S.; Campoy, C.; Hartl, D.; Linde, J.; Klingler, M.; Demmelmair, H.; Caño, A.; Gil, A.; Bondy, B.; et al. Docosahexaenoic acid supply in pregnancy affects placental expression of fatty acid transport proteins. Am. J. Clin. Nutr. 2006, 84, 853–861. [Google Scholar] [CrossRef] [Green Version]
- Kuda, O.; Rossmeisl, M.; Kopecky, J. Omega-3 fatty acids and adipose tissue biology. Mol. Asp. Med. 2018, 64, 147–160. [Google Scholar] [CrossRef]
- Carreras-Badosa, G.; Bonmatí, A.; Ortega, F.-J.; Mercader, J.-M.; Guindo-Martínez, M.; Torrents, D.; Prats-Puig, A.; Martinez-Calcerrada, J.-M.; Platero-Gutierrez, E.; De Zegher, F.; et al. Altered Circulating miRNA Expression Profile in Pregestational and Gestational Obesity. J. Clin. Endocrinol. Metab. 2015, 100, E1446–E1456. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, A.; Fernandez, J.M.; Fernandez, C.; Ferrandez, A.; Lopez-Siguero, J.P.; Sanchez, E.; Sobradillo, B.; Yeste, D. Spanish growth studies 2008. New anthropometric standards. Endocrinol. Nutr. 2008, 55, 484–506. [Google Scholar] [CrossRef]
- Bassols, J.; Martínez-Calcerrada, J.-M.; Osiniri, I.; Díaz-Roldán, F.; Xargay-Torrent, S.; Mas-Parés, B.; Dorado-Ceballos, E.; Prats-Puig, A.; Carreras-Badosa, G.; De Zegher, F.; et al. Effects of metformin administration on endocrine-metabolic parameters, visceral adiposity and cardiovascular risk factors in children with obesity and risk markers for metabolic syndrome: A pilot study. PLoS ONE 2019, 14, e0226303. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, M.; Kumagi, T.; Kurose, K.; Nakanishi, S.; Michitaka, K.; Matsuura, B.; Horiike, N.; Onji, M. A Technique for the Measurement of Visceral Fat by Ultrasonography: Comparison of Measurements by Ultrasonography and Computed Tomography. Intern. Med. 2005, 44, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Serra-Majem, L.; Ribas, L.; Ngo, J.; Ortega, R.M.; García, A.; Pérez-Rodrigo, C.; Aranceta, J. Food, youth and the Mediterranean diet in Spain. Development of KIDMED, Mediterranean Diet Quality Index in children and adolescents. Public Health Nutr. 2004, 7, 931–935. [Google Scholar] [CrossRef] [Green Version]
- Roman, B.; Serra-Majem, L.; Ribas-Barba, L.; Pérez-Rodrigo, C.; Aranceta, J. How many children and adolescents in Spain comply with the recommendations on physical activity? J. Sport. Med. Phys. Fit. 2008, 48, 380–387. [Google Scholar]
- Jetté, M.; Sidney, K.; Blümchen, G. Metabolic equivalents (METS) in exercise testing, exercise prescription, and evaluation of functional capacity. Clin. Cardiol. 1990, 13, 555–565. [Google Scholar] [CrossRef]
- Gómez-Vilarrubla, A.; Mas-Parés, B.; Díaz, M.; Xargay-Torrent, S.; Carreras-Badosa, G.; Jové, M.; Martin-Gari, M.; Bonmatí-Santané, A.; de Zegher, F.; Ibañez, L.; et al. Fatty acids in the placenta of appropiate- versus small-for-gestational-age infants at term birth. Placenta 2021, 109, 4–10. [Google Scholar] [CrossRef]
- Pamplona, R.; Dalfó, E.; Ayala, V.; Bellmunt, M.J.; Prat, J.; Ferrer, I.; Portero-Otín, M. Proteins in Human Brain Cortex Are Modified by Oxidation, Glycoxidation, and Lipoxidation: Effects of alzheimer disease and identification of lipoxidation targets. J. Biol. Chem. 2005, 280, 21522–21530. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Fatty Acid Profile | All Population | |
|---|---|---|
| N = 113 | ||
| Mean ± SD | ||
| SFA (mol %) | ||
| 14:0 | Myristic acid | 1.59 ± 0.46 |
| 16:0 | Palmitic acid | 33.03 ± 2.50 |
| 18:0 | Stearic acid | 17.27 ± 1.50 |
| 20:0 | Arachidic acid | 0.32 ± 0.08 |
| 22:0 | Behenic acid | 1.33 ± 0.92 |
| 24:0 | Lignoceric acid | 0.53 ± 0.47 |
| SFA | 54.07 ± 3.99 | |
| MUFA (mol %) | ||
| 16:1n-7 | Palmitoleic acid | 0.50 ± 0.15 |
| 18:1n-7 | Vaccenic acid | 2.00 ± 0.35 |
| 18:1n-9 | Oleic acid | 12.16 ± 1.55 |
| 20:1n-9 | Eicosenoic acid | 0.26 ± 0.06 |
| 22:1n-9 | Erucic acid | 0.67 ± 0.60 |
| MUFA | 15.59 ± 2.17 | |
| PUFA n-6 series (mol %) | ||
| 18:2n-6 | Linoleic acid (LA) | 9.71 ± 1.39 |
| 20:2n-6 | Eicosadienoic acid | 0.10 ± 0.07 |
| 20:3n-6 | Dihomo-γ-linolenic acid (DGLA) | 0.61 ± 0.09 |
| 20:4n-6 | Arachidonic acid (AA) | 12.59 ± 3.74 |
| 22:4n-6 | Adrenic acid | 0.51 ± 0.17 |
| 22:5n-6 | Osbond acid | 0.10 ± 0.06 |
| PUFAn-6 | 23.62 ± 4.56 | |
| PUFA n-3 series (mol %) | ||
| 18:3n-3 | α-linolenic acid (ALA) | 0.22 ± 0.07 |
| 18:4n-3 | Stearidonic acid | 0.65 ± 0.58 |
| 20:3n-3 | Eicosatrienoic acid | 3.54 ± 1.16 |
| 20:5n-3 | Eicosapentaenoic acid (EPA) | 0.89 ± 0.25 |
| 22:5n-3 | Docosapentaenoic acid (DPA) | 0.25 ± 0.08 |
| 22:6n-3 | Docosahexaenoic acid (DHA) | 1.18 ± 0.52 |
| PUFAn-3 | 6.72 ± 1.26 | |
| n-6/n-3 PUFA RATIOS | ||
| AA/EPA | 14.59 ± 4.05 | |
| AA/DHA | 11.80 ± 4.03 | |
| AA/EPA + DHA | 6.31 ± 1.58 | |
| PUFAn-6/PUFAn-3 | 3.55 ± 0.57 | |
| Follow-Up N = 82 | Weight-SDS | BMI- SDS | % Fat Mass-SDS | Visceral Fat | HOMA-IR |
|---|---|---|---|---|---|
| n-6 series | |||||
| AA (20:4n-6) | 0.110 | 0.062 | 0.126 | 0.225 * | 0.156 |
| PUFAn-6 | 0.070 | 0.028 | 0.098 | 0.203 | 0.143 |
| n-3 series | |||||
| EPA (20:5n-3) | −0.113 | −0.129 | −0.159 | −0.105 | −0.049 |
| DHA (22:6n-3) | 0.012 | −0.039 | 0.149 | 0.123 | 0.168 |
| PUFAn-3 | 0.055 | 0.039 | 0.190 | 0.153 | 0.158 |
| n-6/n-3 ratios | |||||
| AA/EPA | 0.245 * | 0.202 * | 0.306 ** | 0.383 ** | 0.239 * |
| AA/DHA | 0.100 | 0.115 | −0.065 | 0.080 | −0.053 |
| AA/EPA + DHA | 0.183 | 0.170 | 0.119 | 0.254 * | 0.085 |
| PUFAn-6/PUFAn-3 | 0.030 | −0.007 | −0.082 | 0.094 | 0.008 |
| Placental Fatty Acid Transporters (2−ΔCt) | All Population |
|---|---|
| N = 113 | |
| Mean ± SD | |
| FATP1 | 0.19 ± 0.19 |
| FATP4 | 0.33 ± 0.20 |
| Ʃ FATPs | 0.52 ± 0.32 |
| Ʃ FATP1 + FATP4 < 50th Centile N = 42 | Weight-SDS | BMI- SDS | % Fat Mass-SDS | Visceral Fat | HOMA-IR |
|---|---|---|---|---|---|
| n-6 series | |||||
| AA (20:4n-6) | 0.173 | 0.148 | 0.113 | 0.196 | 0.038 |
| PUFAn-6 | 0.097 | 0.089 | 0.038 | 0.147 | 0.041 |
| n-3 series | |||||
| EPA (20:5n-3) | −0.153 | −0.127 | −0.348 * | −0.327 * | −0.057 |
| DHA (22:6n-3) | −0.208 | −0.095 | 0.005 | −0.007 | 0.088 |
| PUFAn-3 | 0.057 | 0.063 | 0.195 | 0.102 | 0.009 |
| n-6/n-3 ratios | |||||
| AA/EPA | 0.230 | 0.195 | 0.255 | 0.321 * | 0.056 |
| AA/DHA | 0.273 | 0.181 | 0.092 | 0.165 | −0.025 |
| AA/EPA + DHA | 0.256 | 0.189 | 0.170 | 0.248 | 0.016 |
| PUFAn-6/PUFAn-3 | 0.062 | 0.049 | −0.091 | 0.078 | 0.037 |
| Ʃ FATP1 + FATP4 > 50th centile N = 40 | Weight-SDS | BMI- SDS | % Fat Mass-SDS | Visceral fat | HOMA-IR |
| n-6 series | |||||
| AA (20:4n-6) | 0.261 | 0.243 | 0.430 ** | 0.367 * | 0.355 * |
| PUFAn-6 | 0.228 | 0.208 | 0.421 ** | 0.351 * | 0.282 |
| n-3 series | |||||
| EPA (20:5n-3) | −0.070 | −0.034 | −0.007 | −0.161 | −0.242 |
| DHA (22:6n-3) | 0.223 | 0.131 | 0.409 * | 0.209 | 0.227 |
| PUFAn-3 | 0.155 | 0.183 | 0.361 * | 0.202 | 0.265 |
| n-6/n-3 ratios | |||||
| AA/EPA | 0.281 | 0.234 | 0.374 * | 0.460 ** | 0.474 ** |
| AA/DHA | −0.124 | −0.006 | −0.265 | −0.001 | −0.076 |
| AA/EPA + DHA | 0.070 | 0.126 | 0.023 | 0.317 * | 0.245 |
| PUFAn-6/PUFAn-3 | 0.030 | −0.024 | −0.030 | 0.101 | −0.066 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Vilarrubla, A.; Mas-Parés, B.; Carreras-Badosa, G.; Jové, M.; Berdún, R.; Bonmatí-Santané, A.; de Zegher, F.; Ibañez, L.; López-Bermejo, A.; Bassols, J. Placental AA/EPA Ratio Is Associated with Obesity Risk Parameters in the Offspring at 6 Years of Age. Int. J. Mol. Sci. 2023, 24, 10087. https://doi.org/10.3390/ijms241210087
Gómez-Vilarrubla A, Mas-Parés B, Carreras-Badosa G, Jové M, Berdún R, Bonmatí-Santané A, de Zegher F, Ibañez L, López-Bermejo A, Bassols J. Placental AA/EPA Ratio Is Associated with Obesity Risk Parameters in the Offspring at 6 Years of Age. International Journal of Molecular Sciences. 2023; 24(12):10087. https://doi.org/10.3390/ijms241210087
Chicago/Turabian StyleGómez-Vilarrubla, Ariadna, Berta Mas-Parés, Gemma Carreras-Badosa, Mariona Jové, Rebeca Berdún, Alexandra Bonmatí-Santané, Francis de Zegher, Lourdes Ibañez, Abel López-Bermejo, and Judit Bassols. 2023. "Placental AA/EPA Ratio Is Associated with Obesity Risk Parameters in the Offspring at 6 Years of Age" International Journal of Molecular Sciences 24, no. 12: 10087. https://doi.org/10.3390/ijms241210087
APA StyleGómez-Vilarrubla, A., Mas-Parés, B., Carreras-Badosa, G., Jové, M., Berdún, R., Bonmatí-Santané, A., de Zegher, F., Ibañez, L., López-Bermejo, A., & Bassols, J. (2023). Placental AA/EPA Ratio Is Associated with Obesity Risk Parameters in the Offspring at 6 Years of Age. International Journal of Molecular Sciences, 24(12), 10087. https://doi.org/10.3390/ijms241210087