
Histone H3 and H4 Modifications Point to Transcriptional Suppression as a Component of Winter Freeze Tolerance in the Gall Fly Eurosta solidaginis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
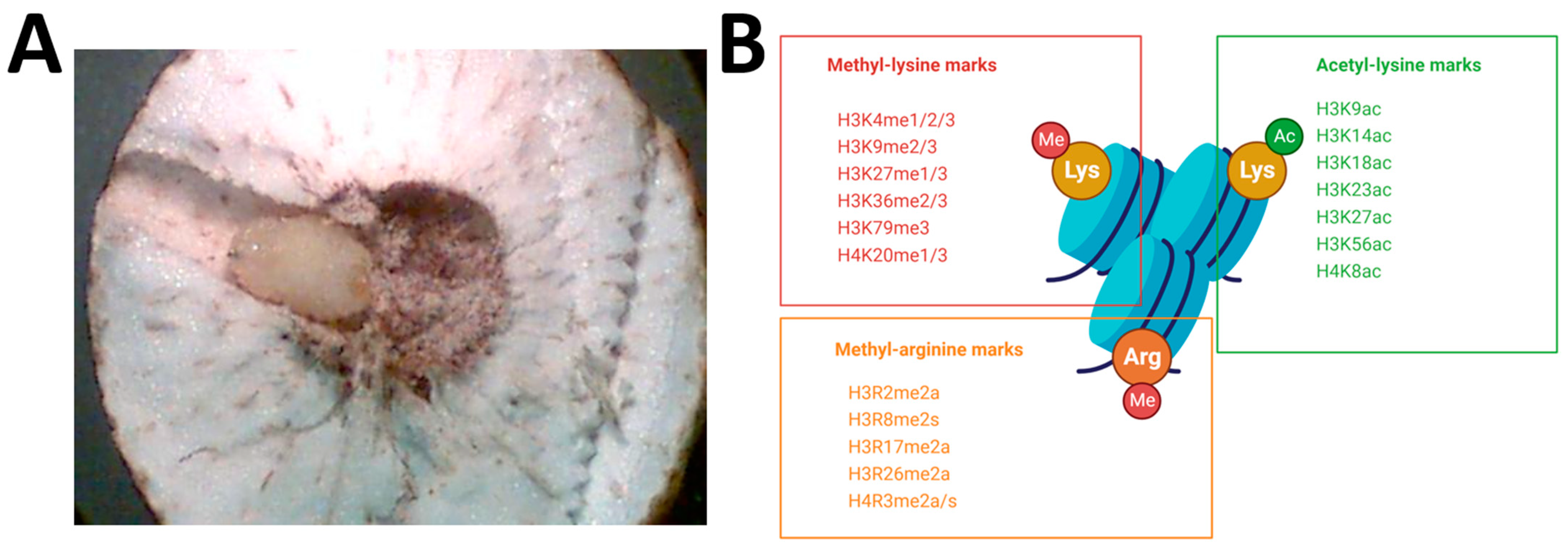
:1. Introduction
2. Results
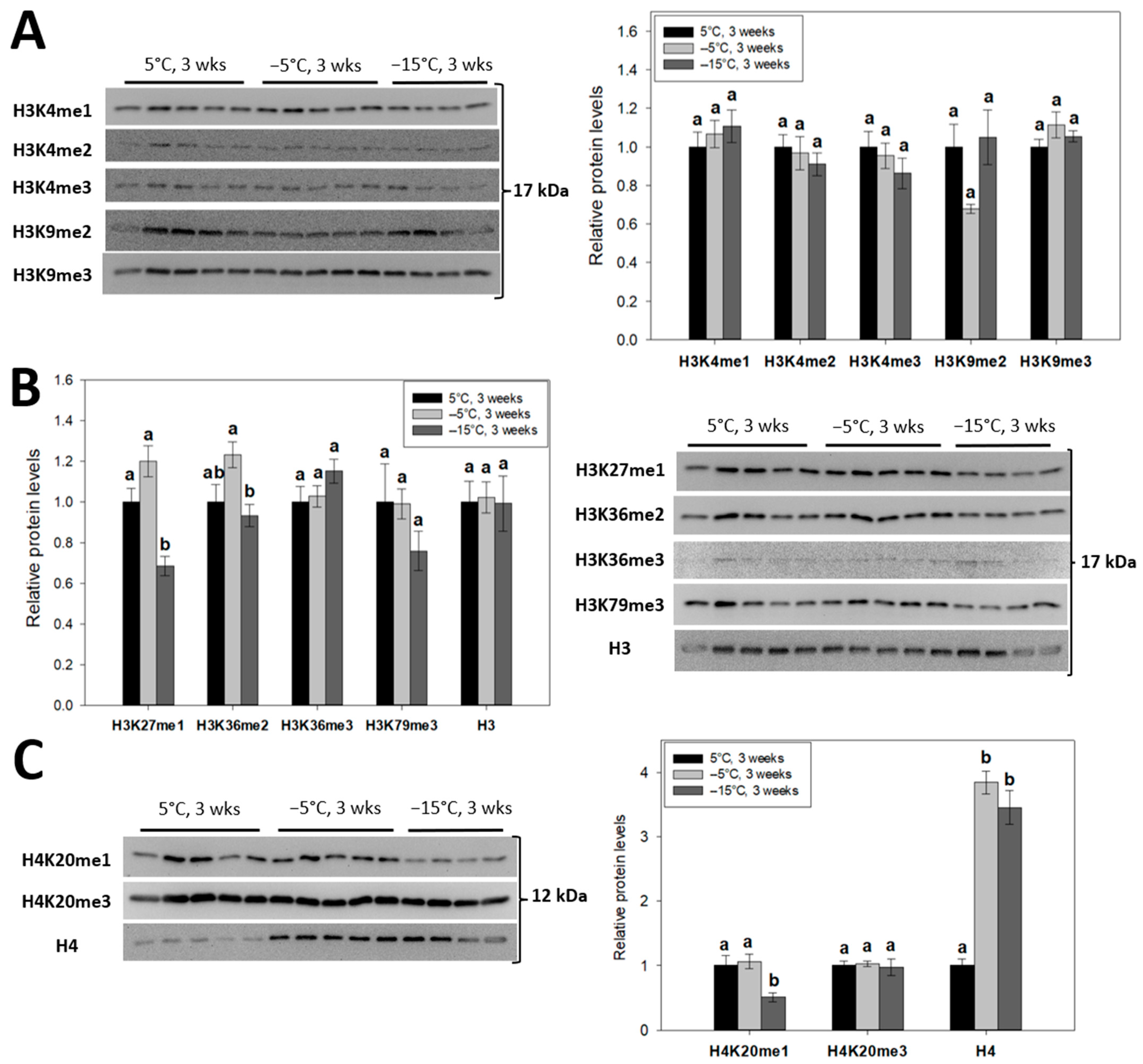
2.1. Permissive Monomethylations of H3K27 and H4K20 Are Significantly Reduced in Frozen Goldenrod Gall Fly Larvae
2.2. Cold and Freezing Stresses Upregulate the Nuclear Levels of Histone H4, but Not H3
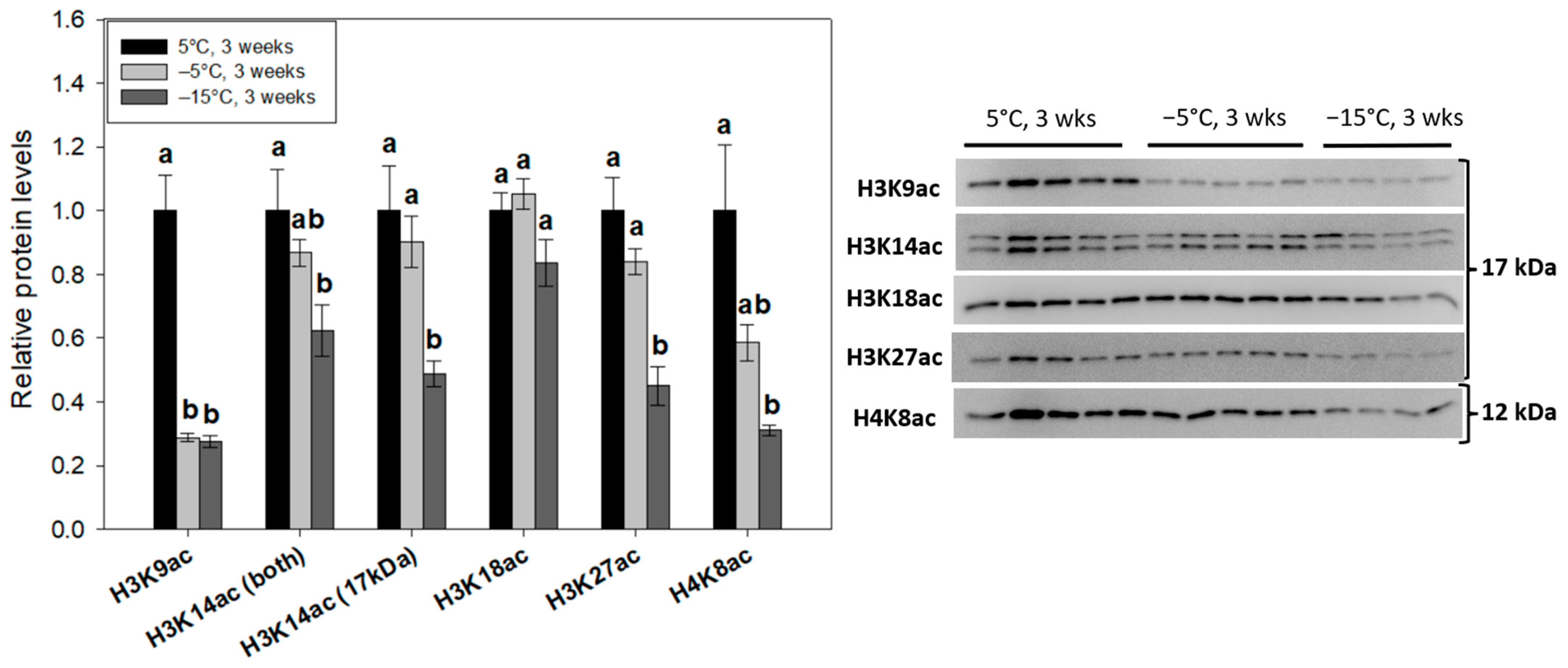
2.3. Decreased Acetylation of Prominent Histone Lysine Residues Occurs during Freezing
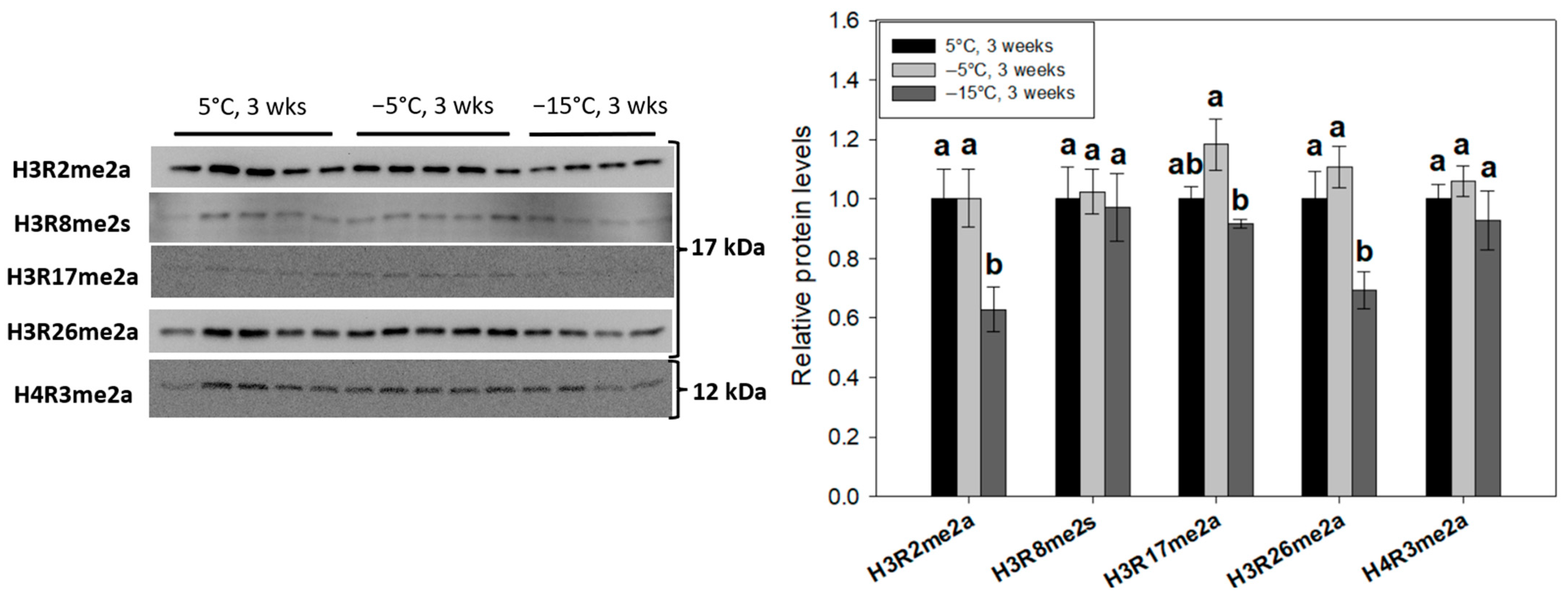
2.4. H3R2me2a, H3R17me2a and H3R26me2a Are Cold-Responsive Methyl-Arginine Histone Modifications
3. Discussion
3.1. Freeze Exposure Triggers Reductions in Many Transcriptionally Permissive Histone Modifications
3.2. Elevated Levels of Histone H4 May Play a Role in Transcriptional Regulation and Cell Protection
3.3. Conclusions
4. Materials and Methods
4.1. Animal Experiments
4.2. Nuclear Protein Isolation
4.3. Western Immunoblotting
4.4. Quantification and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Storey, K.B.; Storey, J.M. Metabolic rate depression in animals: Transcriptional and translational controls. Biol. Rev. Camb. Philos. Soc. 2004, 79, 207–233. [Google Scholar]
- Storey, K.B.; Storey, J.M. Metabolic rate depression and biochemical adaptation in anaerobiosis, hibernation and estivation. Q. Rev. Biol. 1990, 65, 145–174. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B. Regulation of hypometabolism: Insights into epigenetic controls. J. Exp. Biol. 2015, 218, 150–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolfe, D.F.S.; Brown, G.C. Cellular energy utilization and molecular origin of standard metabolic rate in mammals. Physiol. Rev. 1997, 77, 731–758. [Google Scholar] [CrossRef] [Green Version]
- Hochachka, P.W.; Buck, L.T.; Doll, C.J.; Land, S.C. Unifying theory of hypoxia tolerance: Molecular/metabolic defense and rescue mechanisms for surviving oxygen lack. Proc. Natl. Acad. Sci. USA 1996, 93, 9493. [Google Scholar] [CrossRef] [Green Version]
- Bloskie, T.; Storey, K.B. Epigenetics of the frozen brain: Roles for lysine methylation in hypometabolism. FEBS Lett. 2022, 596, 2007–2020. [Google Scholar]
- Krivoruchko, A.; Storey, K.B. Epigenetics in anoxia tolerance: A role for histone deacetylases. Mol. Cell. Biochem. 2010, 342, 151–161. [Google Scholar] [CrossRef]
- Morin, P.; Storey, K.B. Evidence for a reduced transcriptional state during hibernation in ground squirrels. Cryobiology 2006, 53, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Wijenayake, S.; Storey, K.B. The role of DNA methylation during anoxia tolerance in a freshwater turtle (Trachemys scripta elegans). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 333–342. [Google Scholar] [CrossRef]
- Alvarado, S.; Mak, T.; Liu, S.; Storey, K.B.; Szyf, M. Dynamic changes in global and gene-specific DNA methylation during hibernation in adult thirteen-lined ground squirrels, Ictidomys tridecemlineatus. J. Exp. Biol. 2015, 218, 1787–1795. [Google Scholar] [CrossRef] [Green Version]
- Storey, K.B.; Storey, J.M. Insect cold hardiness: Metabolic, gene, and protein adaptation. Can. J. Zool. 2012, 90, 456–475. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Energetics of insect diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Hoback, W.; Stanley, D. Insects in hypoxia. J. Insect Physiol. 2001, 47, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- Costanzo, J.P.; Lee, R.E. Avoidance and tolerance of freezing in ectothermic vertebrates. J. Exp. Biol. 2013, 216, 1961–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B.; Storey, J.M. Freeze tolerance in animals. Physiol. Rev. 1988, 68, 27–84. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, D.R.; Storey, K.B. Oxidative stress and antioxidants in overwintering larvae of cold-hardy goldenrod gall insects. J. Exp. Biol. 1996, 199, 1483–1491. [Google Scholar] [CrossRef]
- Rider, M.H.; Hussain, N.; Dilworth, S.M.; Storey, J.M.; Storey, K.B. AMP-activated protein kinase and metabolic regulation in cold-hardy insects. J. Insect Physiol. 2011, 57, 1453–1462. [Google Scholar] [CrossRef]
- Smolinski, M.B.; Green, S.R.; Storey, K.B. Glucose-6-phosphate dehydrogenase is posttranslationally regulated in the larvae of the freeze-tolerant gall fly, Eurosta solidaginis, in response to freezing. Arch. Insect Biochem. Physiol. 2019, 102, e21618. [Google Scholar] [CrossRef]
- Zhang, G.; Storey, J.M.; Storey, K.B. Chaperone proteins and winter survival by a freeze tolerant insect. J. Insect Physiol. 2011, 57, 1115–1122. [Google Scholar] [CrossRef]
- Tzika, E.; Dreker, T.; Imhof, A. Epigenetics and metabolism in health and disease. Front. Genet. 2018, 9, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, L.J.; Storey, K.B. Advances and applications of environmental stress adaptation research. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 240, 110623. [Google Scholar] [CrossRef] [PubMed]
- McGee, S.L.; Hargreaves, M. Epigenetics and exercise. Trends Endocrinol. Metab. 2019, 30, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Bewick, A.J.; Vogel, K.J.; Moore, A.J.; Schmitz, R.J. Evolution of DNA methylation across insects. Mol. Biol. Evol. 2017, 34, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Lyko, F. DNA methylation learns to fly. Trends Genet. 2001, 17, 169–172. [Google Scholar] [CrossRef]
- Field, L.M.; Lyko, F.; Mandrioli, M.; Prantera, G. DNA methylation in insects. Insect Mol. Biol. 2004, 13, 109–115. [Google Scholar] [CrossRef] [Green Version]
- Boros, I.M. Histone modification in Drosophila. Brief. Funct. Genom. 2012, 11, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell. Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Cutter, A.R.; Hayes, J.J. A brief review of nucleosome structure. FEBS Lett. 2015, 589, 2914–2922. [Google Scholar] [CrossRef] [Green Version]
- Bedford, M.T.; Clarke, S.G. Protein arginine methylation in mammals: Who, what, and why. Mol. Cell 2009, 33, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guccione, E.; Richard, S. The regulation, functions and clinical relevance of arginine methylation. Nat. Rev. Mol. Cell. Biol. 2019, 20, 642–657. [Google Scholar] [CrossRef]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-resolution profiling of histone methylations in the human genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, T.S.; Ku, M.; Jaffe, D.B.; Issac, B.; Lieberman, E.; Giannoukos, G.; Alvarez, P.; Brockman, W.; Kim, T.K.; Koche, R.P.; et al. Genome-wide maps of chromatin state in pluripotent and lineage-committed cells. Nature 2007, 448, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, erasing and reading histone lysine methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoaib, M.; Chen, Q.; Shi, X.; Nair, N.; Prasanna, C.; Yang, R.; Walter, D.; Frederiksen, K.S.; Einarsson, H.; Svensson, J.P.; et al. Histone H4 lysine 20 mono-methylation directly facilitates chromatin openness and promotes transcription of housekeeping genes. Nat. Commun. 2021, 12, 4800. [Google Scholar] [CrossRef]
- Kaelin, W.G.; McKnight, S.L. Influence of metabolism on epigenetics and disease. Cell 2013, 153, 56–69. [Google Scholar] [CrossRef] [Green Version]
- Shahbazian, M.D.; Grunstein, M. Functions of site-specific histone acetylation and deacetylation. Annu. Rev. Biochem. 2007, 76, 75–100. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Molecular physiology of freeze tolerance in vertebrates. Physiol. Rev. 2017, 97, 623–665. [Google Scholar] [CrossRef] [Green Version]
- Fraser, K.P.; Houlihan, D.F.; Lutz, P.L.; Leone-Kabler, S.; Manuel, L.; Brechin, J.G. Complete suppression of protein synthesis during anoxia with no post-anoxia protein synthesis debt in the red-eared slider turtle Trachemys scripta elegans. J. Exp. Biol. 2001, 204, 4353–4360. [Google Scholar] [CrossRef]
- Hawkins, L.J.; Storey, K.B. Histone methylation in the freeze-tolerant wood frog (Rana sylvatica). J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2018, 188, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Naranjo Vera, M. Histone Arginine Methylation in the Freeze-Tolerant Wood Frog, Rana sylvatica. Master’s Thesis, Carleton University, Ottawa, ON, Canada, 2022. [Google Scholar]
- Wang, L.; Joshi, P.; Miller, E.L.; Higgins, L.; Slattery, M.; Simon, J.A. A role for monomethylation of histone H3-K27 in gene activity in Drosophila. Genetics 2018, 208, 1023–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari, K.J.; Scelfo, A.; Jammula, S.; Cuomo, A.; Barozzi, I.; Stützer, A.; Fischle, W.; Bonaldi, T.; Pasini, D. Polycomb-dependent H3K27me1 and H3K27me2 regulate active transcription and enhancer fidelity. Mol. Cell 2014, 53, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Wijenayake, S.; Hawkins, L.J.; Storey, K.B. Dynamic regulation of six histone H3 lysine (K) methyltransferases in response to prolonged anoxia exposure in a freshwater turtle. Gene 2018, 649, 50–57. [Google Scholar] [CrossRef]
- Abbas, T.; Shibata, E.; Park, J.; Jha, S.; Karnani, N.; Dutta, A. CRL4(Cdt2) regulates cell proliferation and histone gene expression by targeting PR-Set7/Set8 for degradation. Mol. Cell 2010, 40, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Armstrong, R.L.; Duronio, R.J.; MacAlpine, D.M. Methylation of histone H4 lysine 20 by PR-Set7 ensures the integrity of late replicating sequence domains in Drosophila. Nucleic Acids Res. 2016, 44, 7204. [Google Scholar]
- Biggar, K.K.; Storey, K.B. Perspectives in cell cycle regulation: Lessons from an anoxic vertebrate. Curr. Genom. 2009, 10, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.W.; Storey, K.B. Pattern of cellular quiescence over the hibernation cycle in liver of thirteen-lined ground squirrels. Cell Cycle 2012, 11, 1714–1726. [Google Scholar] [CrossRef] [Green Version]
- Williamson, S.M. Epigenetic Underpinnings of Freeze Tolerance in the Goldenrod Gall Fly Eurosta solidaginis and the Goldenrod Gall Moth Epiblema scudderiana. Master’s Thesis, Carleton University, Ottawa, ON, Canada, 2016. [Google Scholar]
- Seibel, B.A.; Häfker, N.S.; Trübenbach, K.; Zhang, J.; Tessier, S.N.; Pörtner, H.O.; Rosa, R.; Storey, K.B. Metabolic suppression during protracted exposure to hypoxia in the jumbo squid, Dosidicus gigas, living in an oxygen minimum zone. J. Exp. Biol. 2014, 217, 2555–2568. [Google Scholar] [CrossRef] [Green Version]
- Biggar, Y.; Storey, K.B. Global DNA modifications suppress transcription in brown adipose tissue during hibernation. Cryobiology 2014, 69, 333–338. [Google Scholar] [CrossRef]
- Wijenayake, S.; Storey, K.B. Dynamic regulation of histone H3 lysine (K) acetylation and deacetylation during prolonged oxygen deprivation in a champion anaerobe. Mol. Cell. Biochem. 2020, 474, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Hatzivassiliou, G.; Sachdeva, U.M.; Bui, T.V.; Cross, J.R.; Thompson, C.B. ATP-citrate lyase links cellular metabolism to histone acetylation. Science 2009, 324, 1076–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etchegaray, J.P.; Mostoslavsky, R. Interplay between metabolism and epigenetics: A nuclear adaptation to environmental changes. Mol. Cell 2016, 62, 695–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joanisse, D.R.; Storey, K.B. Fatty acid content and enzymes of fatty acid metabolism in overwintering cold-hardy gall insects. Physiol. Zool. 1996, 69, 1079–1095. [Google Scholar] [CrossRef]
- Nègre, N.; Brown, C.D.; Ma, L.; Bristow, C.A.; Miller, S.W.; Wagner, U.; Kheradpour, P.; Eaton, M.L.; Loriaux, P.; Sealfon, R.; et al. A cis-regulatory map of the Drosophila genome. Nature 2011, 471, 527–531. [Google Scholar] [CrossRef] [Green Version]
- Karmodiya, K.; Krebs, A.R.; Oulad-Abdelghani, M.; Kimura, H.; Tora, L. H3K9 and H3K14 acetylation co-occur at many gene regulatory elements, while H3K14ac marks a subset of inactive inducible promoters in mouse embryonic stem cells. BMC Genom. 2012, 13, 424. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; Sowpati, D.T.; Mishra, R.K. Epigenomic and genomic landscape of Drosophila melanogaster heterochromatic genes. Genomics 2019, 111, 177–185. [Google Scholar] [CrossRef]
- Tie, F.; Banerjee, R.; Stratton, C.A.; Prasad-Sinha, J.; Stepanik, V.; Zlobin, A.; Diaz, M.O.; Scacheri, P.C.; Harte, P.J. CBP-mediated acetylation of histone H3 lysine 27 antagonizes Drosophila Polycomb silencing. Development 2009, 136, 3131. [Google Scholar] [CrossRef] [Green Version]
- Agalioti, T.; Chen, G.; Thanos, D. Deciphering the transcriptional histone acetylation code for a human gene. Cell 2002, 111, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Guccione, E.; Bassi, C.; Casadio, F.; Martinato, F.; Cesaroni, M.; Schuchlautz, H.; Lüscher, B.; Amati, B. Methylation of histone H3R2 by PRMT6 and H3K4 by an MLL complex are mutually exclusive. Nature 2007, 449, 933–937. [Google Scholar] [CrossRef]
- Rosenfeld, J.A.; Wang, Z.; Schones, D.E.; Zhao, K.; DeSalle, R.; Zhang, M.Q. Determination of enriched histone modifications in non-genic portions of the human genome. BMC Genom. 2009, 10, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchard, C.; Sahu, P.; Meixner, M.; Nötzold, R.R.; Rust, M.B.; Kremmer, E.; Feederle, R.; Hart-Smith, G.; Finkernagel, F.; Bartkuhn, M.; et al. Genomic location of PRMT6-dependent H3R2 methylation is linked to the transcriptional outcome of associated genes. Cell Rep. 2018, 24, 3339–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, M.C.; Miranda, T.B.; Clarke, S.; Di Fruscio, M.; Suter, B.; Lasko, P.; Richard, S. Characterization of the Drosophila protein arginine methyltransferases DART1 and DART4. Biochem. J. 2004, 379, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Prado, F.; Jimeno-González, S.; Reyes, J.C. Histone availability as a strategy to control gene expression. RNA Biol. 2017, 14, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zheng, X.; Zhang, L.; Zhou, B.; Hu, H.; Li, Z.; Zhang, L.; Lin, Y.; Wang, X. Histone H4 expression is cooperatively maintained by IKKβ and Akt1 which attenuates cisplatin-induced apoptosis through the DNA-PK/RIP1/IAPs signaling cascade. Sci. Rep. 2017, 7, 41715. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bloskie, T.; Storey, K.B. Histone H3 and H4 Modifications Point to Transcriptional Suppression as a Component of Winter Freeze Tolerance in the Gall Fly Eurosta solidaginis. Int. J. Mol. Sci. 2023, 24, 10153. https://doi.org/10.3390/ijms241210153
Bloskie T, Storey KB. Histone H3 and H4 Modifications Point to Transcriptional Suppression as a Component of Winter Freeze Tolerance in the Gall Fly Eurosta solidaginis. International Journal of Molecular Sciences. 2023; 24(12):10153. https://doi.org/10.3390/ijms241210153
Chicago/Turabian StyleBloskie, Tighe, and Kenneth B. Storey. 2023. "Histone H3 and H4 Modifications Point to Transcriptional Suppression as a Component of Winter Freeze Tolerance in the Gall Fly Eurosta solidaginis" International Journal of Molecular Sciences 24, no. 12: 10153. https://doi.org/10.3390/ijms241210153
APA StyleBloskie, T., & Storey, K. B. (2023). Histone H3 and H4 Modifications Point to Transcriptional Suppression as a Component of Winter Freeze Tolerance in the Gall Fly Eurosta solidaginis. International Journal of Molecular Sciences, 24(12), 10153. https://doi.org/10.3390/ijms241210153