

Genome-Wide Identification of the YABBY Gene Family in Dendrobium Orchids and Its Expression Patterns in Dendrobium chrysotoxum

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
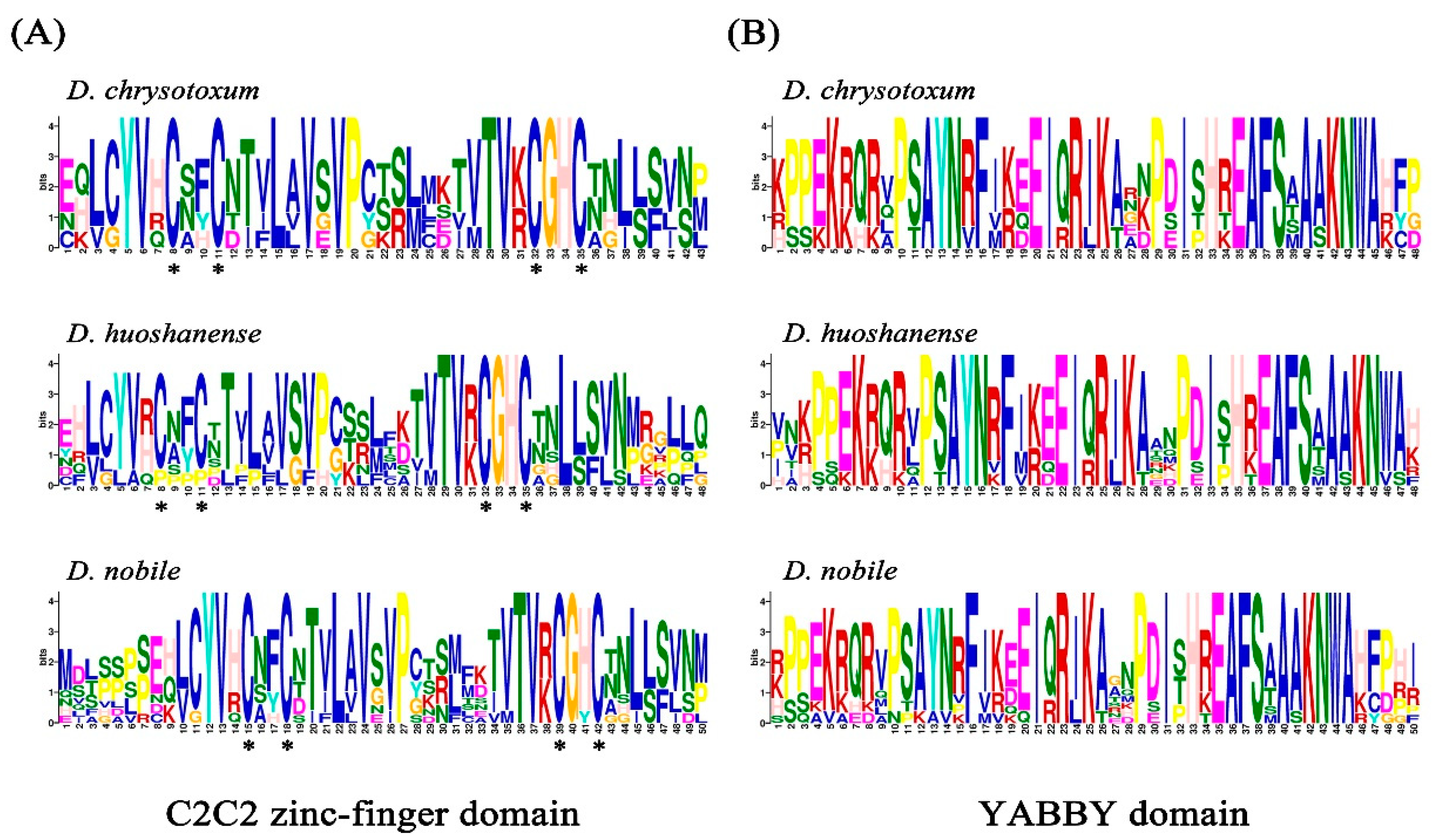
2.1. Screening of DchYABBY, DhuYABBY, and DnoYABBY Transcription Factors
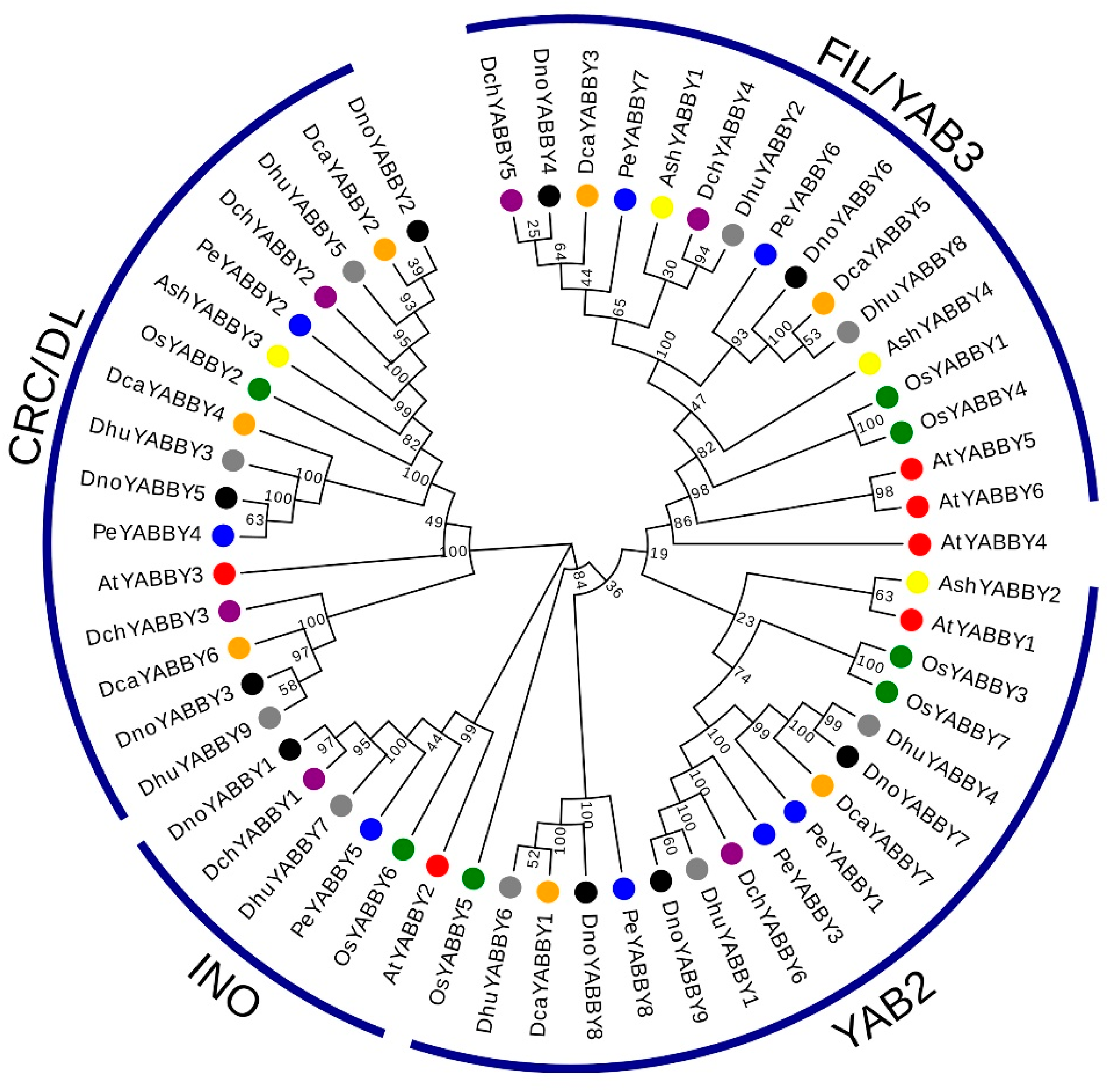
2.2. Phylogeny and Classification of YABBY Genes
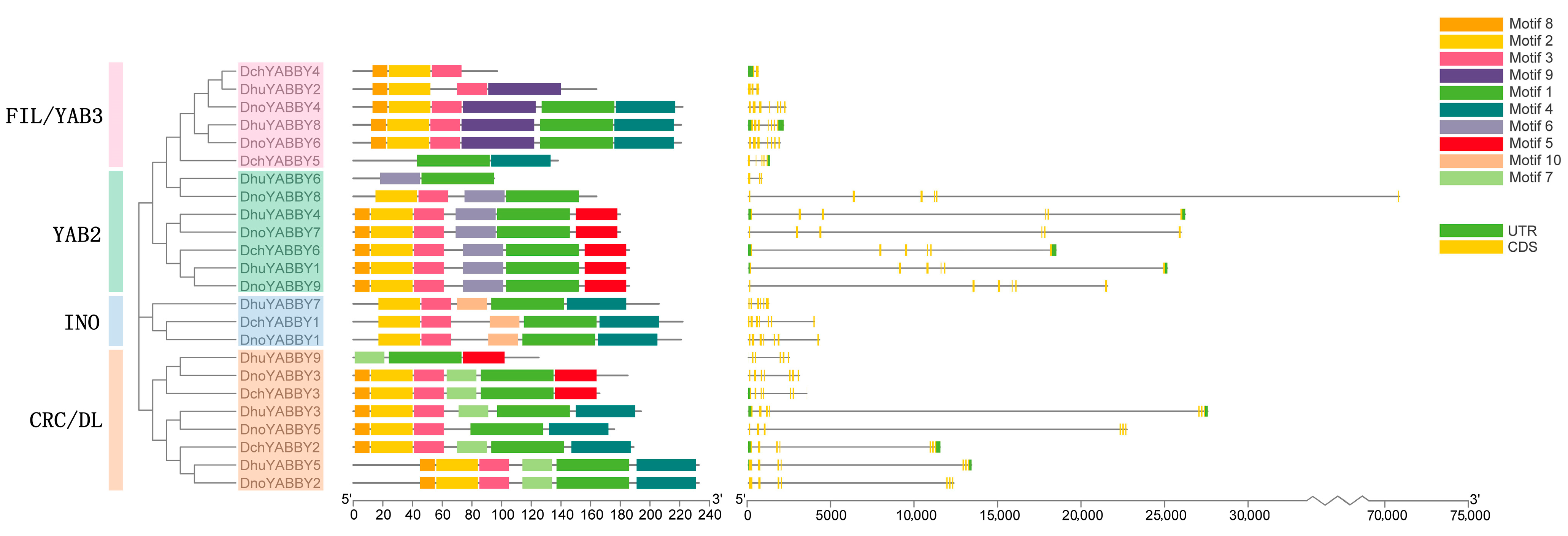
2.3. Gene Structure and Motif Analysis of YABBY Genes
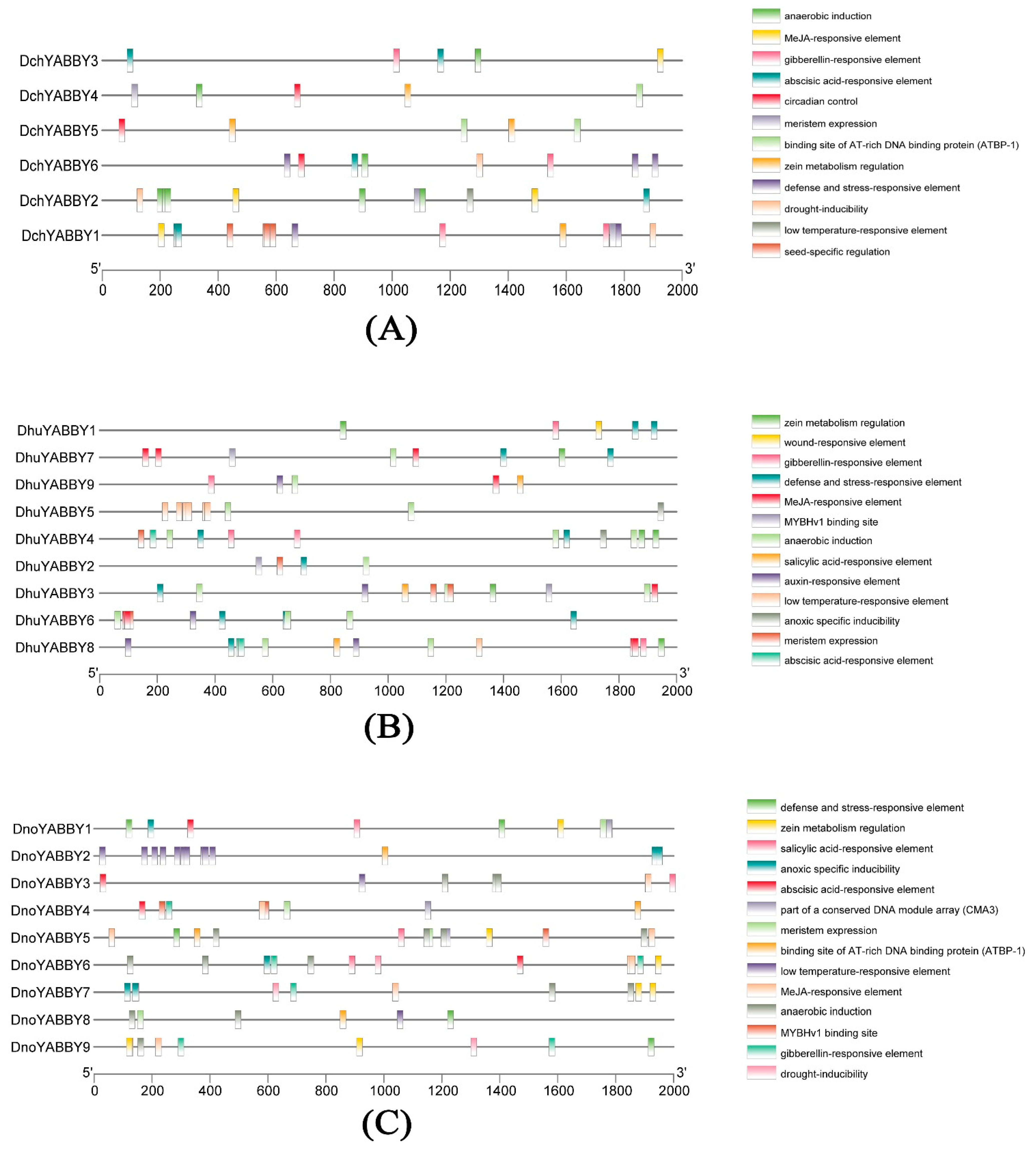
2.4. Promoter Analysis of YABBY Genes
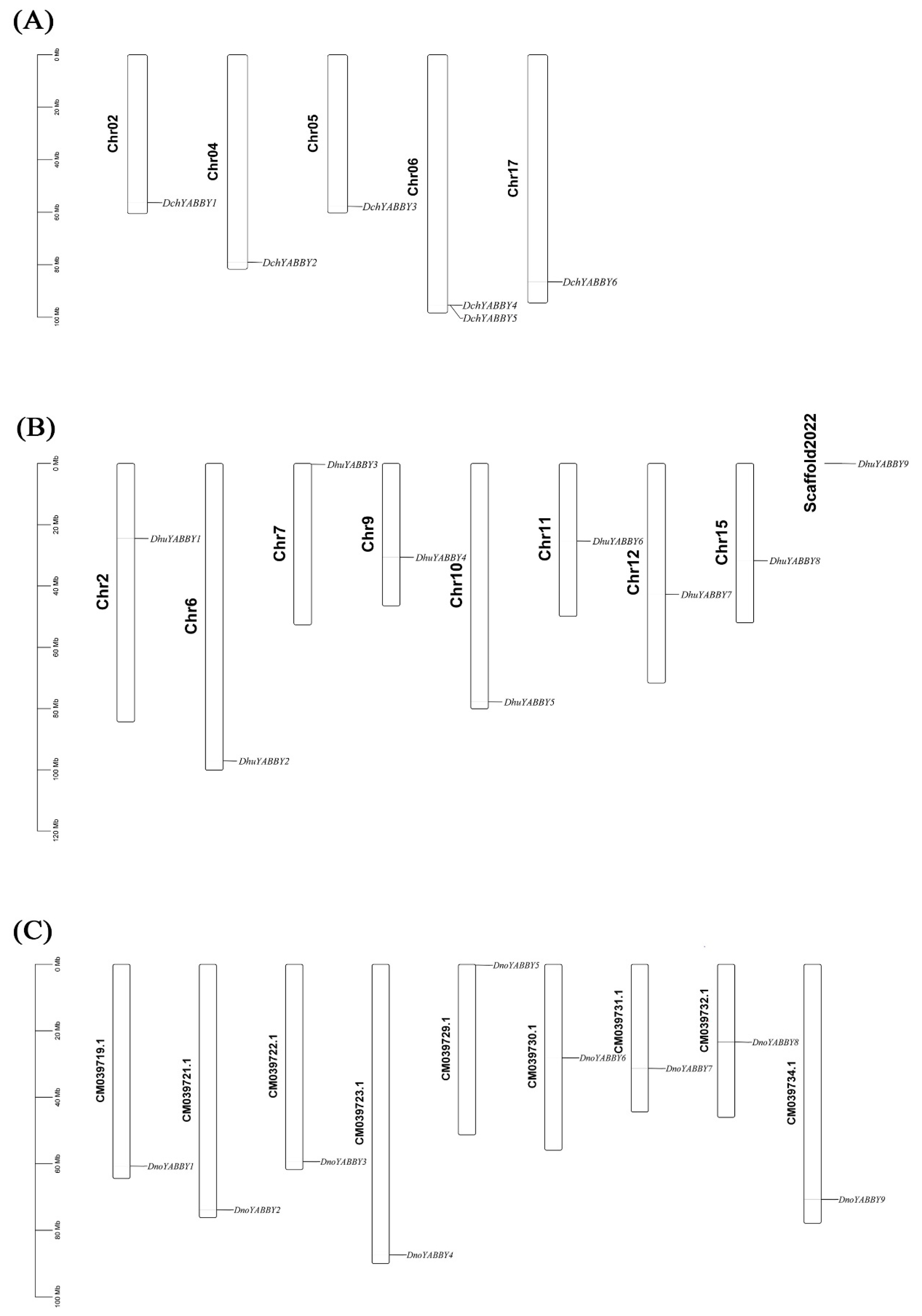
2.5. Chromosomal Localization of YABBY Genes
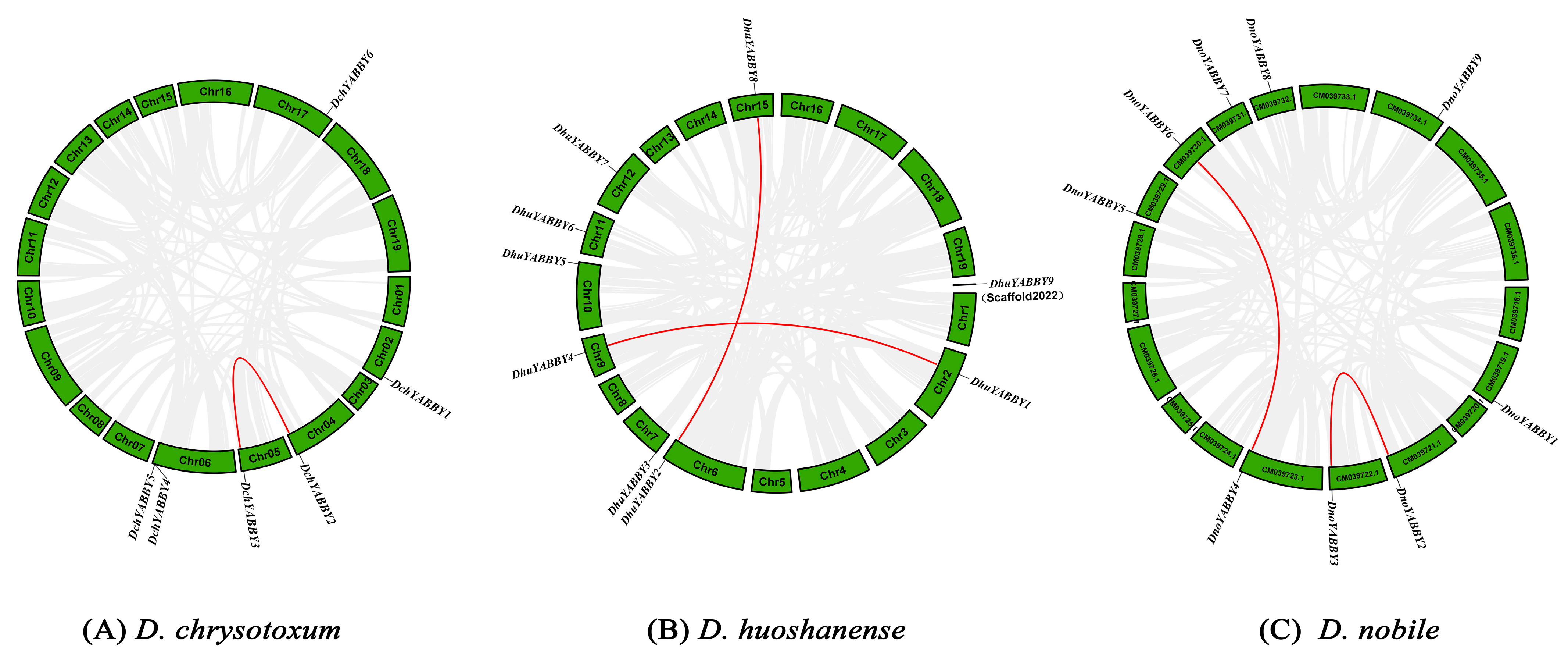
2.6. Collinearity Analysis and Ka/Ks Value of YABBY Genes
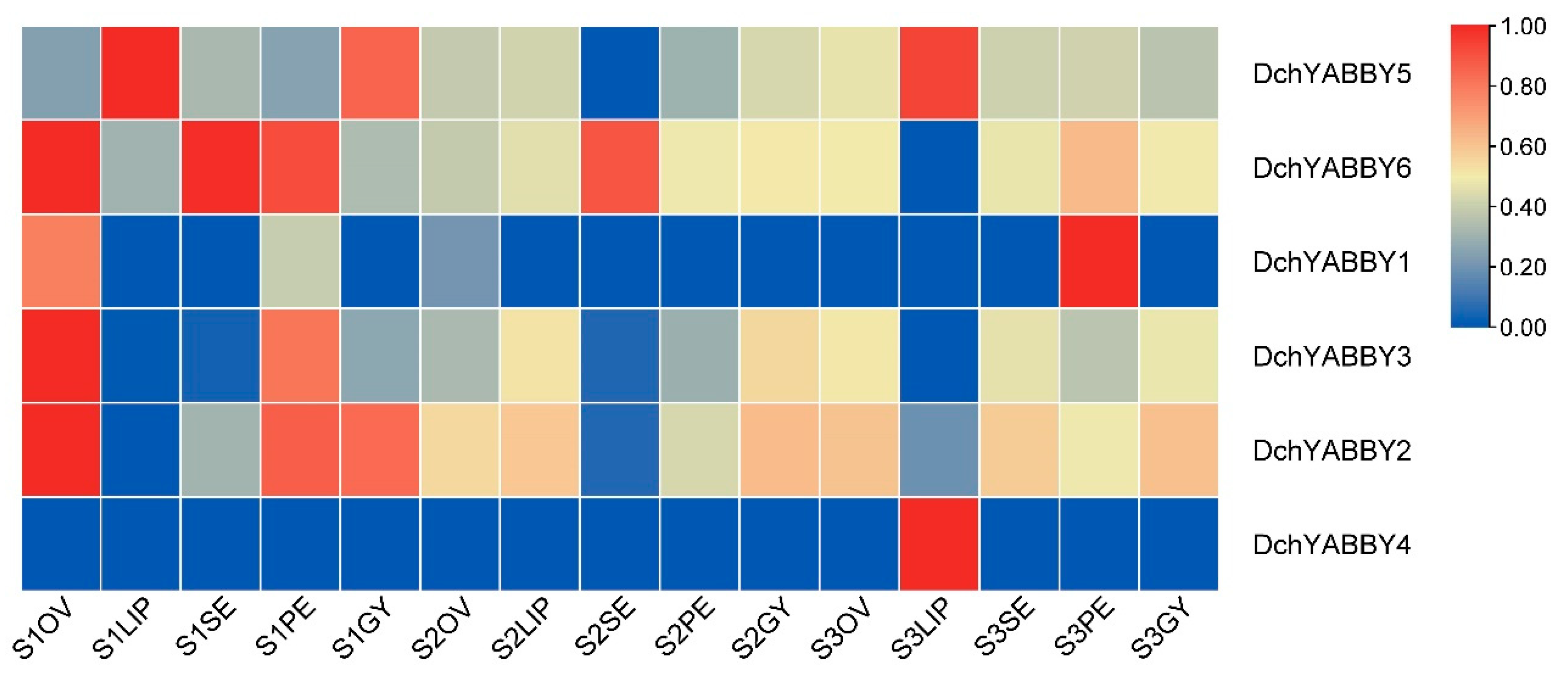
2.7. Expression Patterns of YABBY Genes in D. chrysotoxum
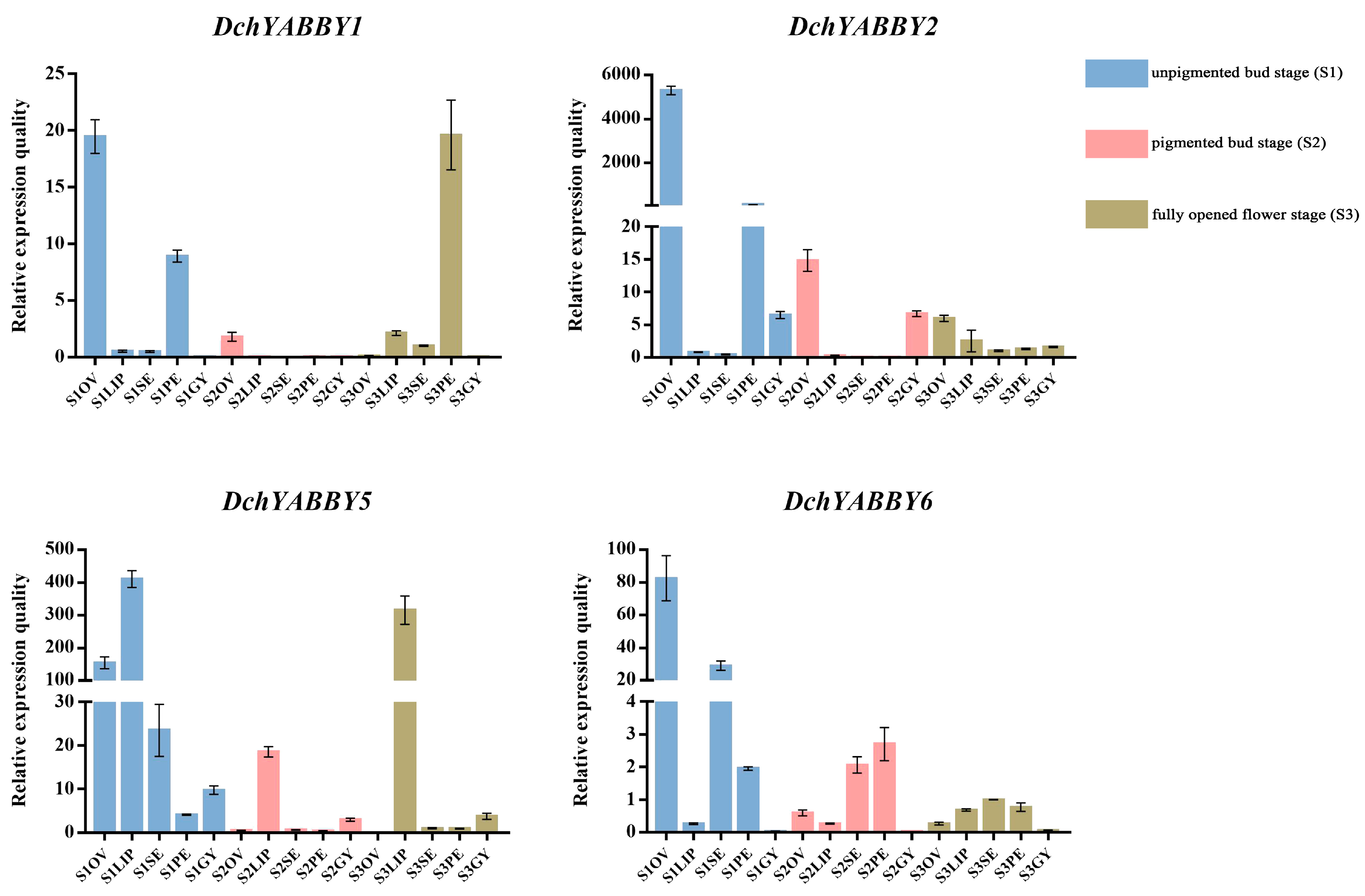
2.8. The qRT-PCR Analysis of YABBY Genes in D. chrysotoxum
3. Discussion
4. Materials and Methods
4.1. Data Sources
4.2. Identification and Physicochemical Properties of the YABBY Gene Family
4.3. Phylogenetic Analysis of YABBY Genes
4.4. Gene Structure and Conserved Motif Analysis of YABBY Genes
4.5. Analysis of YABBY Genes Promoter Sequences
4.6. Chromosomal Localization and Synteny Analysis of YABBY Genes
4.7. Expression Analysis and RT‒qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bowman, J.L.; Smyth, D.R.; Meyerowitz, E.M. Genes directing flower development in Arabidopsis. Plant Cell 1989, 1, 37–52. [Google Scholar]
- Zhao, S.-P.; Lu, D.; Yu, T.-F.; Ji, Y.-J.; Zheng, W.-J.; Zhang, S.-X.; Chai, S.-C.; Chen, Z.-Y.; Cui, X.-Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Zhao, Y.; Ma, Q.; Hu, Y.; Hedden, P.; Zhang, Q.; Zhou, D.-X. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism. Plant Physiol. 2007, 144, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, M.K.; Bowman, J.L.; Sundaresan, V. YABBY polarity genes mediate the repression of KNOX homeobox genes in Arabidopsis. Plant Cell 2002, 14, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Finet, C.; Floyd, S.K.; Conway, S.J.; Zhong, B.; Scutt, C.P.; Bowman, J.L. Evolution of the YABBY gene family in seed plants. Evol. Dev. 2016, 18, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Golz, J.F.; Roccaro, M.; Kuzoff, R.; Hudson, A. GRAMINIFOLIA promotes growth and polarity of Antirrhinum leaves. Development 2004, 131, 3661–3670. [Google Scholar] [CrossRef] [Green Version]
- Kanaya, E.; Nakajima, N.; Okada, K. Non-sequence-specific DNA binding by the FILAMENTOUS FLOWER protein from Arabidopsis thaliana is reduced by EDTA. J. Biol. Chem. 2002, 277, 11957–11964. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Baum, S.F.; Oh, S.H.; Jiang, C.Z.; Chen, J.C.; Bowman, J.L. Recruitment of CRABS CLAW to promote nectary development within the eudicot clade. Development 2005, 132, 5021–5032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Xu, Y.Y.; Xu, Z.H.; Chong, K. A rice YABBY gene, OsYABBY4, preferentially expresses in developing vascular tissue. Dev. Genes Evol. 2007, 217, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.-Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet. Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef]
- Yamada, T.; Ito, M.; Kato, M. YABBY2-Homologue Expression in Lateral Organs of Amborella trichopoda (Amborellaceae). Int. J. Plant Sci. 2004, 165, 917–924. [Google Scholar] [CrossRef]
- Bartholmes, C.; Hidalgo, O.; Gleissberg, S. Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biol. 2012, 14, 11–23. [Google Scholar] [CrossRef]
- Soundararajan, P.; Won, S.Y.; Park, D.S.; Lee, Y.H.; Kim, J.S. Comparative Analysis of the YABBY Gene Family of Bienertia sinuspersici, a Single-Cell C4 Plant. Plants 2019, 8, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q.; et al. Genome-Wide Study of YABBY Genes in Upland Cotton and Their Expression Patterns under Different Stresses. Front. Genet. 2018, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Atkinson, A.; Otsuga, D.; Christensen, T.; Reynolds, L.; Drews, G.N. The Arabidopsis FILAMENTOUS FLOWER gene is required for flower formation. Development 1999, 126, 2715–2726. [Google Scholar] [CrossRef]
- Fourquin, C.; Vinauger-Douard, M.; Fogliani, B.; Dumas, C.; Scutt, C.P. Evidence that CRABS CLAW and TOUSLED have conserved their roles in carpel development since the ancestor of the extant angiosperms. Proc. Natl. Acad. Sci. USA 2005, 102, 4649–4654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva, J.M.; Broadhvest, J.; Hauser, B.A.; Meister, R.J.; Schneitz, K.; Gasser, C.S. INNER NO OUTER regulates abaxial-adaxial patterning in Arabidopsis ovules. Genes Dev. 1999, 13, 3160–3169. [Google Scholar] [CrossRef] [Green Version]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar] [CrossRef]
- Yang, T.; He, Y.; Niu, S.; Zhang, Y. A YABBY gene CRABS CLAW a (CRCa) negatively regulates flower and fruit sizes in tomato. Plant Sci. 2022, 320, 111285. [Google Scholar] [CrossRef]
- Jiu, S.; Zhang, Y.; Han, P.; Han, Y.; Xu, Y.; Liu, G.; Leng, X. Genome-Wide Identification and Expression Analysis of VviYABs Family Reveal Its Potential Functions in the Developmental Switch and Stresses Response During Grapevine Development. Front. Genet. 2021, 12, 762221. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef] [Green Version]
- Christenhusz, M.J.M.; Byng, J.W. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Pridgeon, A.M.; Cribb, P.J.; Chase, M.W.; Rasmussen, F.N. Genera Orchidacearum Volume 6: Epidendroideae (Part Three); Oxford University Press: Walton Street, UK, 2014. [Google Scholar]
- Zhang, Y.; Zhang, G.Q.; Zhang, D.; Liu, X.D.; Xu, X.Y.; Sun, W.H.; Yu, X.; Zhu, X.; Wang, Z.W.; Zhao, X.; et al. Chromosome-scale assembly of the Dendrobium chrysotoxum genome enhances the understanding of orchid evolution. Hortic. Res. 2021, 8, 183. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Jing, Y.; Dai, J.; Zheng, T.; Gu, F.; Zhao, Q.; Zhu, F.; Song, X.; Deng, H.; Wei, P.; et al. A Chromosome-Level Genome Assembly of Dendrobium Huoshanense Using Long Reads and Hi-C Data. Genome Biol. Evol. 2020, 12, 2486–2490. [Google Scholar] [CrossRef]
- Xu, Q.; Niu, S.-C.; Li, K.-L.; Zheng, P.-J.; Zhang, X.-J.; Jia, Y.; Liu, Y.; Niu, Y.-X.; Yu, L.-H.; Chen, D.-F.; et al. Chromosome-Scale Assembly of the Dendrobium nobile Genome Provides Insights Into the Molecular Mechanism of the Biosynthesis of the Medicinal Active Ingredient of Dendrobium. Front. Genet. 2022, 13, 844622. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Hsiao, Y.Y.; Chang, S.B.; Zhang, D.; Lan, S.R.; Liu, Z.J.; Tsai, W.C. Genome-Wide Identification of YABBY Genes in Orchidaceae and Their Expression Patterns in Phalaenopsis Orchid. Genes 2020, 11, 955. [Google Scholar] [CrossRef]
- Wang, Q.-Q.; Li, Y.-Y.; Chen, J.; Zhu, M.-J.; Liu, X.; Zhou, Z.; Zhang, D.; Liu, Z.-J.; Lan, S. Genome-wide identification of YABBY genes in three Cymbidium species and expression patterns in C. ensifolium (Orchidaceae). Front. Plant Sci. 2022, 13, 995734. [Google Scholar] [CrossRef]
- Cao, Y.; Lang, Z.; Wang, L. Characteristics and Expression Analysis of Transcription Factor YABBY Family in Maize. J. Agric. Sci. Technol. 2015, 17, 32–41. [Google Scholar]
- Buttar, Z.A.; Yang, Y.; Sharif, R.; Wu, S.N.; Xie, Y.; Wang, C. Genome wide identification, characterization, and expression analysis of YABBY-gene family in wheat (Triticum aestivum L.). Agronomy 2020, 10, 1189. [Google Scholar] [CrossRef]
- Ma, R.; Huang, B.; Huang, Z.; Zhang, Z. Genome-wide identification and analysis of the YABBY gene family in Moso Bamboo (Phyllostachys edulis (Carrière) J. Houz). PeerJ 2021, 9, 11780. [Google Scholar] [CrossRef]
- Li, Z.; Li, G.; Cai, M.; Priyadarshani, S.V.; Aslam, M.; Zhou, Q.; Huang, X.; Wang, X.; Liu, Y.; Qin, Y. Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. Int. J. Mol. Sci. 2019, 20, 5863. [Google Scholar] [CrossRef] [Green Version]
- Hao, L.; Zhang, J.; Shi, S.; Li, P.; Li, D.; Zhang, T.; Guo, H. Identification and expression profiles of the YABBY transcription factors in wheat. PeerJ 2020, 10, 12855. [Google Scholar] [CrossRef]
- De Almeida, A.M.R.; Yockteng, R.; Schnable, J.; Alvarez-Buylla, E.R.; Freeling, M.; Specht, C.D. Co-option of the polarity gene network shapes filament morphology in angiosperms. Sci. Rep. 2014, 4, 6194. [Google Scholar] [CrossRef] [Green Version]
- Demuth, J.P.; Hahn, M.W. The life and death of gene families. BioEssays 2009, 31, 29–39. [Google Scholar] [CrossRef]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef]
- Soltis, P.S.; Soltis, D.E. Ancient WGD events as drivers of key innovations in angiosperms. Curr. Opin. Plant Biol. 2016, 30, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Li, G.; Dai, J.; Deng, H. Genome-Wide Analysis of PEBP Genes in Dendrobium huoshanense: Unveiling the Antagonistic Functions of FT/TFL1 in Flowering Time. Front. Genet. 2021, 12, 687689. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, W.; Zhang, Y.; Zhang, X.; Lang, D.; Zhang, X. The roles of methyl jasmonate to stress in plants. Funct. Plant Biol. 2019, 46, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.A.; Rumpho, M.E.; Fox, T.C. Anaerobic Metabolism in Plants. Plant Physiol. 1992, 100, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Si, C.; da Silva, J.A.T.; Dai, G.; Duan, J.; He, C. Characterization of YABBY genes in Dendrobium officinale reveals their potential roles in flower development. Protoplasma 2023, 260, 483–495. [Google Scholar] [CrossRef]
- Lee, J.Y.; Baum, S.F.; Alvarez, J.; Patel, A.; Chitwood, D.H.; Bowman, J.L. Activation of CRABS CLAW in the nectaries and carpels of Arabidopsis. Plant Cell 2005, 17, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Boter, M.; Golz, J.F.; Gimenez-Ibanez, S.; Fernandez-Barbero, G.; Franco-Zorrilla, J.M.; Solano, R. FILAMENTOUS FLOWER is a direct target of JAZ3 and modulates responses to jasmonate. Plant Cell 2015, 27, 3160–3174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Wang, Z. Integrative Metabolomic and Transcriptome Analysis Reveal the Differential Mechanisms of Spot Color in the Lips of Dendrobium chrysotoxum. J. Plant Biol. 2022, 1–13. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, 216–227. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Q.; Zhao, X.; Huang, Y.; Zhang, M.-M.; He, X.; Ke, S.; Li, Y.; Zhang, C.; Ahmad, S.; Lan, S.; et al. Genome-Wide Identification of the YABBY Gene Family in Dendrobium Orchids and Its Expression Patterns in Dendrobium chrysotoxum. Int. J. Mol. Sci. 2023, 24, 10165. https://doi.org/10.3390/ijms241210165
Zheng Q, Zhao X, Huang Y, Zhang M-M, He X, Ke S, Li Y, Zhang C, Ahmad S, Lan S, et al. Genome-Wide Identification of the YABBY Gene Family in Dendrobium Orchids and Its Expression Patterns in Dendrobium chrysotoxum. International Journal of Molecular Sciences. 2023; 24(12):10165. https://doi.org/10.3390/ijms241210165
Chicago/Turabian StyleZheng, Qinyao, Xuewei Zhao, Ye Huang, Meng-Meng Zhang, Xin He, Shijie Ke, Yuanyuan Li, Cuili Zhang, Sagheer Ahmad, Siren Lan, and et al. 2023. "Genome-Wide Identification of the YABBY Gene Family in Dendrobium Orchids and Its Expression Patterns in Dendrobium chrysotoxum" International Journal of Molecular Sciences 24, no. 12: 10165. https://doi.org/10.3390/ijms241210165
APA StyleZheng, Q., Zhao, X., Huang, Y., Zhang, M. -M., He, X., Ke, S., Li, Y., Zhang, C., Ahmad, S., Lan, S., Li, M., & Liu, Z. -J. (2023). Genome-Wide Identification of the YABBY Gene Family in Dendrobium Orchids and Its Expression Patterns in Dendrobium chrysotoxum. International Journal of Molecular Sciences, 24(12), 10165. https://doi.org/10.3390/ijms241210165