

Endogenous Biological Drivers in Diabetic Lower Limb Wounds Recurrence: Hypothetical Reflections
Abstract
:1. Introduction
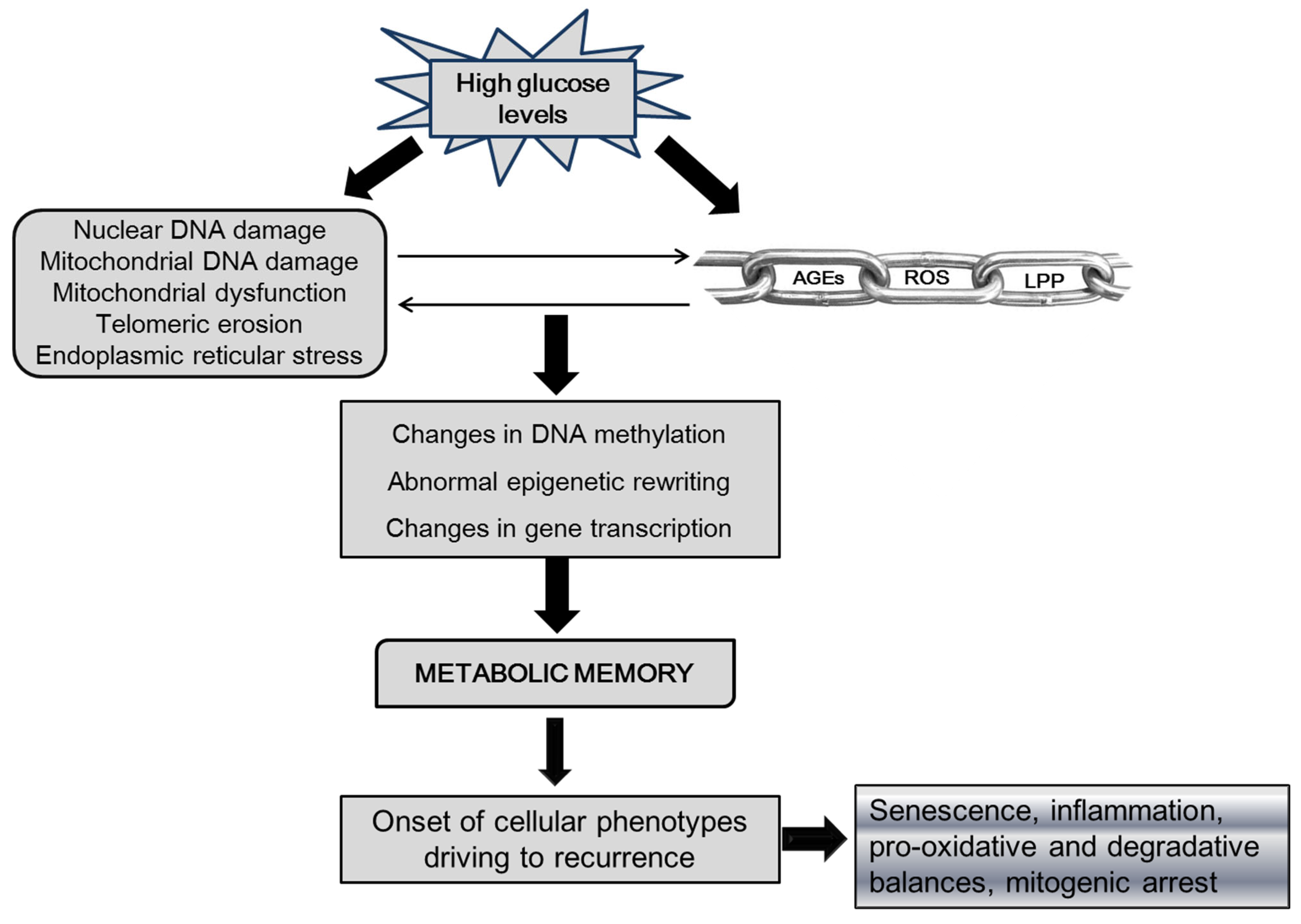
1.1. Potential Epigenetic Drivers in Ulcer Recurrence
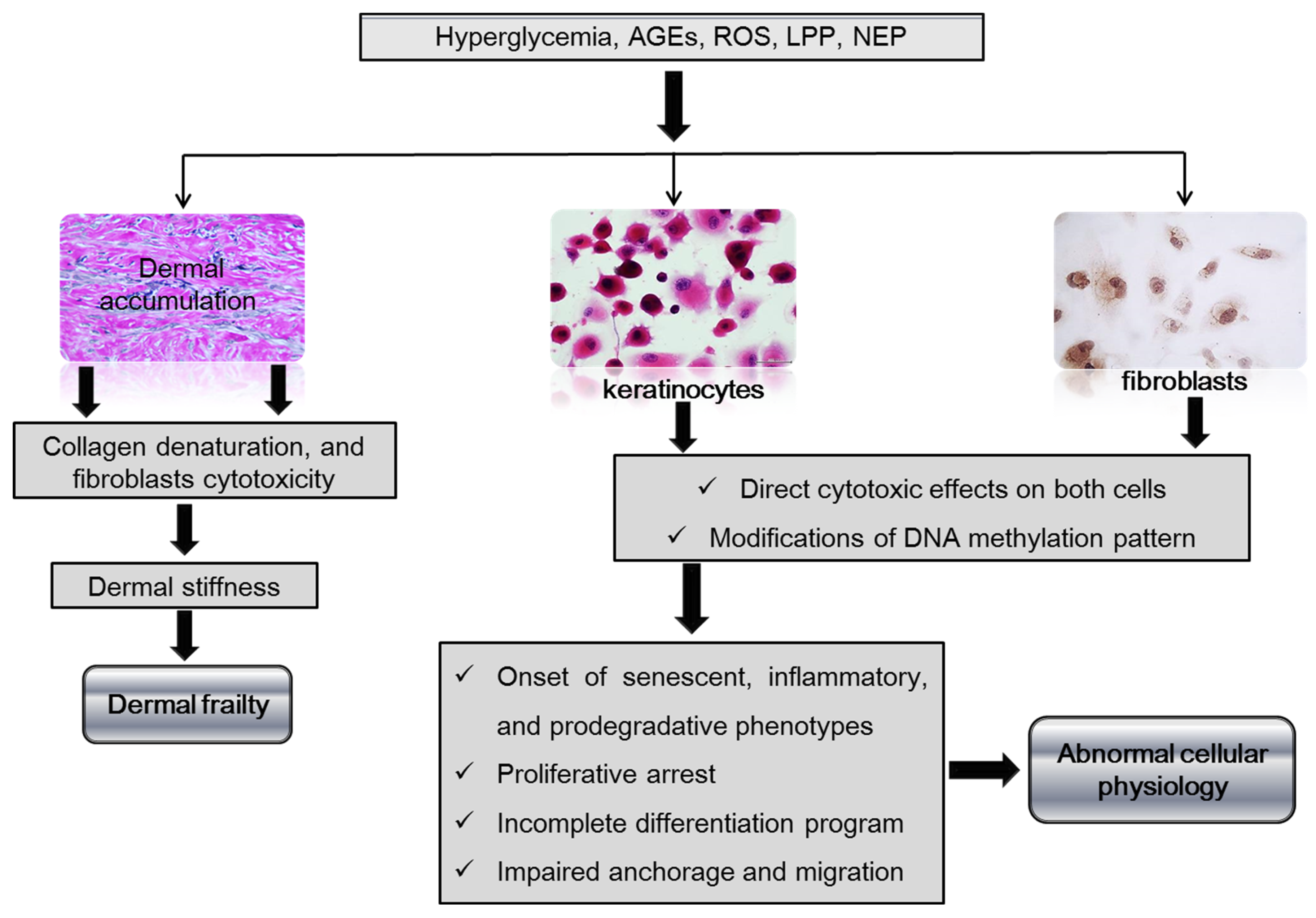
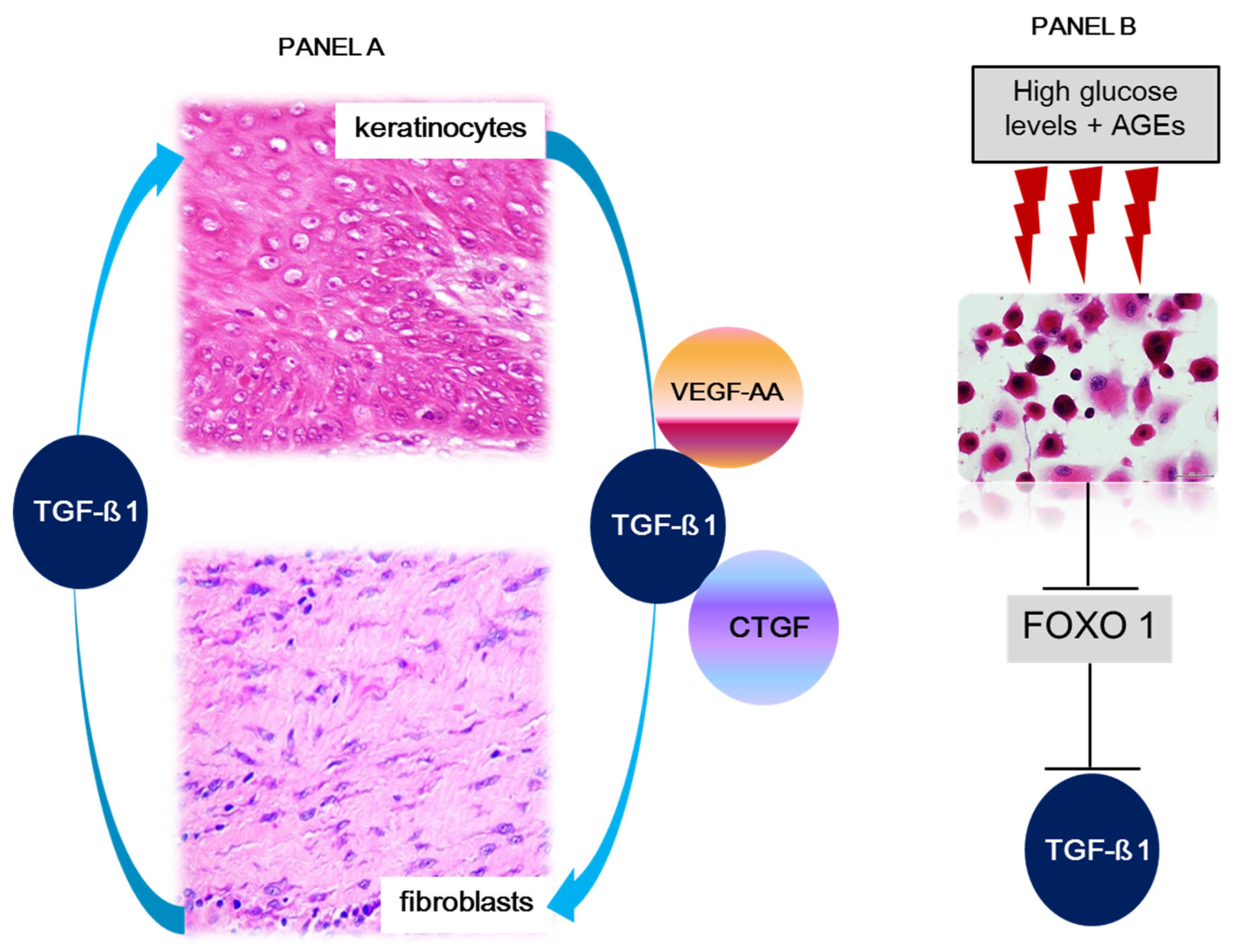
1.2. Dermal Matrix, Fibroblasts, and Keratinocytes in Ulcer Recurrence
2. Ulcer Recurrences in the Clinical Arena
3. Concluding Remarks and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cheng, H.T.; Xu, X.; Lim, P.S.; Hung, K.Y. Worldwide Epidemiology of Diabetes-Related End-Stage Renal Disease, 2000–2015. Diabetes Care 2021, 44, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, Z.H.; Qiang, H.; Wu, J.; Shen, M.; Zhang, L.; Lyu, J. Trends in the incidence of diabetes mellitus: Results from the Global Burden of Disease Study 2017 and implications for diabetes mellitus prevention. BMC Public Health 2020, 20, 1415. [Google Scholar] [CrossRef] [PubMed]
- Mobasseri, M.; Shirmohammadi, M.; Amiri, T.; Vahed, N.; Hosseini, F.H.; Ghojazadeh, M. Prevalence and incidence of type 1 diabetes in the world: A systematic review and meta-analysis. Health Promot. Perspect. 2020, 10, 98–115. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martin, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Magliano, D.J.; Sacre, J.W.; Harding, J.L.; Gregg, E.W.; Zimmet, P.Z.; Shaw, J.E. Young-onset type 2 diabetes mellitus—Implications for morbidity and mortality. Nat. Rev. Endocrinol. 2020, 16, 321–331. [Google Scholar] [CrossRef]
- Rosengren, A.; Dikaiou, P. Cardiovascular outcomes in type 1 and type 2 diabetes. Diabetologia 2023, 66, 425–437. [Google Scholar] [CrossRef]
- Mauricio, D.; Alonso, N.; Gratacòs, M. Chronic Diabetes Complications: The Need to Move beyond Classical Concepts. Trends Endocrinol. Metab. 2020, 31, 287–295. [Google Scholar] [CrossRef]
- Natarajan, R. Epigenetic Mechanisms in Diabetic Vascular Complications and Metabolic Memory: The 2020 Edwin Bierman Award Lecture. Diabetes 2021, 70, 328–337. [Google Scholar] [CrossRef]
- Deng, J.-Y.; Wu, X.-Q.; He, W.-J.; Liao, X.; Tang, M.; Nie, X.-Q. Targeting DNA methylation and demethylation in diabetic foot ulcers. J. Adv. Res. 2023. [Google Scholar] [CrossRef]
- Doğruel, H.; Aydemir, M.; Balci, M.K. Management of diabetic foot ulcers and the challenging points: An endocrine view. World J. Diabetes 2022, 13, 27–36. [Google Scholar] [CrossRef]
- Chen, L.; Sun, S.; Gao, Y.; Ran, X. Global mortality of diabetic foot ulcer: A systematic review and meta-analysis of observational studies. Diabetes Obes. Metab. 2022, 25, 36–45. [Google Scholar] [CrossRef] [PubMed]
- McDermott, K.; Fang, M.; Boulton, A.J.; Selvin, E.; Hicks, C.W. Etiology, Epidemiology, and Disparities in the Burden of Diabetic Foot Ulcers. Diabetes Care 2023, 46, 209–221. [Google Scholar] [CrossRef]
- Pastore, D.; Deja-Simoni, A.; De Stefano, A.; Pacifici, F.; Cela, E.; Infante, M.; Coppola, A.; Di Daniele, N.; Lauro, D.; Della-Morte, D.; et al. Risk factors for diabetic foot ulcers: An Albanian retrospective study of inpatients with type 2 diabetes. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 558–572. [Google Scholar] [PubMed]
- Jiang, M.; Gan, F.; Gan, M.; Deng, H.; Chen, X.; Yuan, X.; Huang, D.; Liu, S.; Qin, B.; Wei, Y.; et al. Predicting the Risk of Diabetic Foot Ulcers from Diabetics With Dysmetabolism: A Retrospective Clinical Trial. Front. Endocrinol. 2022, 13, 929864. [Google Scholar] [CrossRef] [PubMed]
- Boulton, A.J.; Armstrong, D.D.G.; Löndahl, M.; Frykberg, D.R.G.; Game, F.L.; Edmonds, M.E.; Orgill, D.P.; Kramer, K.; Gurtner, G.C.; Januszyk, M.; et al. New Evidence-Based Therapies for Complex Diabetic Foot Wounds. ADA Clin. Compend. 2022, 2022, 1–23. [Google Scholar] [CrossRef]
- Rai, V.; Moellmer, R.; Agrawal, D.K. Clinically relevant experimental rodent models of diabetic foot ulcer. Mol. Cell. Biochem. 2022, 477, 1239–1247. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, N.; Martínez-Jiménez, I.; García-Ojalvo, A.; Mendoza-Marí, Y.; Guillén-Nieto, G.; Armstrong, D.; Berlanga-Acosta, J. Wound chronicity, impaired immunity and infection in diabetic patients. MEDICC Rev. 2022, 24, 44. [Google Scholar] [CrossRef]
- Leal, E.C.; Carvalho, E. Heme Oxygenase-1 as Therapeutic Target for Diabetic Foot Ulcers. Int. J. Mol. Sci. 2022, 23, 12043. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.A.; Guillén-Nieto, G.E.; Rodríguez-Rodríguez, N.; Mendoza-Mari, Y.; Bringas-Vega, M.L.; Berlanga-Saez, J.O.; Del Barco Herrera, D.G.; Martinez-Jimenez, I.; Hernandez-Gutierrez, S.; Valdes-Sosa, P.A. Cellular Senescence as the Pathogenic Hub of Diabetes-Related Wound Chronicity. Front. Endocrinol. 2020, 11, 573032. [Google Scholar] [CrossRef]
- Dhawan, P.; Vasishta, S.; Balakrishnan, A.; Joshi, M.B. Mechanistic insights into glucose induced vascular epigenetic reprogramming in type 2 diabetes. Life Sci. 2022, 298, 120490. [Google Scholar] [CrossRef]
- Bouly, M.; Laborne, F.-X.; Tourte, C.; Henry, E.; Penfornis, A.; Dardari, D. Post-healing follow-up study of patients in remission for diabetic foot ulcers Pied-REM study. PLoS ONE 2022, 17, e0268242. [Google Scholar] [CrossRef]
- Guo, Q.; Ying, G.; Jing, O.; Zhang, Y.; Liu, Y.; Deng, M.; Long, S. Influencing factors for the recurrence of diabetic foot ulcers: A meta-analysis. Int. Wound J. 2023, 20, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Huang, Z.; Li, S.; Kou, Y.; Huang, L.; Yu, T.; Hu, A. Risk factors for the recurrence of diabetic foot ulcers among diabetic patients: A meta-analysis. Int. Wound J. 2019, 16, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.J.; Rothenberg, G.M.; Lakhani, P.J.; Zhou, M.; Linders, D.R.; Bloom, J.D.; Wood, K.A.; Armstrong, D.G. Ulcer metastasis? Anatomical locations of recurrence for patients in diabetic foot remission. J. Foot Ankle Res. 2020, 13, 1–6. [Google Scholar] [CrossRef]
- Engberg, S.; Kirketerp-Møller, K.; Andersen, H.U.; Rasmussen, A. Incidence and predictors of recurrent and other new diabetic foot ulcers: A retrospective cohort study. Diabet. Med. 2019, 36, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Hicks, C.W.; Canner, J.K.; Mathioudakis, N.; Lippincott, C.; Sherman, R.L.; Abularrage, C.J. Incidence and Risk Factors Associated with Ulcer Recurrence Among Patients With Diabetic Foot Ulcers Treated in a Multidisciplinary Setting. J. Surg. Res. 2020, 246, 243–250. [Google Scholar] [CrossRef]
- Gazzaruso, C.; Gallotti, P.; Pujia, A.; Montalcini, T.; Giustina, A.; Coppola, A. Predictors of healing, ulcer recurrence and persistence, amputation and mortality in type 2 diabetic patients with diabetic foot: A 10-year retrospective cohort study. Endocrine 2021, 71, 59–68. [Google Scholar] [CrossRef]
- Pang, L.; Wang, Y.; Zheng, M.; Wang, Q.; Lin, H.; Zhang, L.; Wu, L. Transcriptomic study of high–glucose effects on human skin fibroblast cells. Mol. Med. Rep. 2016, 13, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Suárez, R.; Chapela, S.P.; Álvarez-Córdova, L.; Bautista-Valarezo, E.; Sarmiento-Andrade, Y.; Verde, L.; Frias-Toral, E.; Sarmo, G. Epigenetics in Obesity and Diabetes Mellitus: New Insights. Nutrients 2023, 15, 811. [Google Scholar] [CrossRef]
- Wu, Y.-L.; Lin, Z.-J.; Li, C.-C.; Lin, X.; Shan, S.-K.; Guo, B.; Zhang, M.-H.; Li, F.; Yuan, L.-Q.; Li, Z.-h. Epigenetic regulation in metabolic diseases: Mechanisms and advances in clinical study. Signal Transduct. Target. Ther. 2023, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.A.H.; Ansari, S.A.; Mensah-Brown, E.P.K.; Emerald, B.S. The role of DNA methylation in the pathogenesis of type 2 diabetes mellitus. Clin. Epigenetics 2020, 12, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Cugalj, K.B.; Trebusak, P.K.; Kovac, J.; Sket, R.; Jenko, B.B.; Tesovnik, T.; Debelijak, M.; Batelino, T.; Bratina, N. The Role of Epigenetic Modifications in Late Complications in Type 1 Diabetes. Genes 2022, 13, 705. [Google Scholar] [CrossRef] [PubMed]
- Ramteke, P.; Deb, A.; Shepal, V.; Bhat, M.K. Hyperglycemia Associated Metabolic and Molecular Alterations in Cancer Risk, Progression, Treatment, and Mortality. Cancers 2019, 11, 1402. [Google Scholar] [CrossRef] [Green Version]
- Rehman, S.; Aatif, M.; Rafi, Z.; Khan, M.Y.; Shahab, U.; Ahmad, S.; Farhan, A. Effect of non-enzymatic glycosylation in the epigenetics of cancer. Semin. Cancer Biol. 2022, 83, 543–555. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Miao, F.; Braffett, B.H.; Lachin, J.M.; Zhang, L.; Wu, X.; Roshandel, D.; Carless, M.; Li, X.A.; Tompkins, J.D.; et al. DNA methylation mediates development of HbA1c-associated complications in type 1 diabetes. Nat. Metab. 2020, 2, 744–762. [Google Scholar] [CrossRef]
- Ye, J.; Stefan-Lifshitz, M.; Tomer, Y. Genetic and environmental factors regulate the type 1 diabetes gene CTSH via differential DNA methylation. J. Biol. Chem. 2021, 296, 100774. [Google Scholar] [CrossRef]
- Park, L.K.; Maione, A.G.; Smith, A.; Gerami-Naini, B.; Iyer, L.K.; Mooney, D.J.; Veves, A.; Garlick, J.A. Genome-wide DNA methylation analysis identifies a metabolic memory profile in patient-derived diabetic foot ulcer fibroblasts. Epigenetics 2014, 9, 1339–1349. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Luan, Y.; Feng, Q.; Chen, X.; Qin, B.; Ren, K.-D.; Luan, Y. Epigenetics and Beyond: Targeting Histone Methylation to Treat Type 2 Diabetes Mellitus. Front. Pharmacol. 2022, 12, 807413. [Google Scholar] [CrossRef]
- Babu, M.; Durga, D.T.; Makinen, P.; Kaikkonen, M.; Lesch, H.P.; Junttila, S.; Laiho, A.; Ghimire, B.; Gyenesei, A.; Yla-Herttuala, S. Differential Promoter Methylation of Macrophage Genes Is Associated with Impaired Vascular Growth in Ischemic Muscles of Hyperlipidemic and Type 2 Diabetic Mice: Genome-Wide Promoter Methylation Study. Circ Res. 2015, 117, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Agrawal, N.K.; Gupta, S.K.; Mohan, G.; Chaturvedi, S.; Singh, K. Genetic and epigenetic alterations in Toll like receptor 2 and wound healing impairment in type 2 diabetes patients. J. Diabetes Its Complicat. 2015, 29, 222–229. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, S.; Shu, B.; Chen, L.; Yang, R.; Xu, Y.; Xie, J.; Liu, X.; Qi, S. Transient High Glucose Causes Persistent Vascular Dysfunction and Delayed Wound Healing by the DNMT1-Mediated Ang-1/NF-kappaB Pathway. J. Invest Dermatol. 2021, 141, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Bandara, N.; Gurusinghe, S.; Kong, A.; Mitchell, G.; Wang, L.; Lim, S.Y.; Strappe, P. Generation of a nitric oxide signaling pathway in mesenchymal stem cells promotes endothelial lineage commitment. J. Cell. Physiol. 2019, 234, 20392–20407. [Google Scholar] [CrossRef] [PubMed]
- Mir, A.R.; Habib, S.; Uddin, M. Recent advances in histone glycation: Emerging role in diabetes and cancer. Glycobiology 2021, 31, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Omans, N.D.; Leicher, R.; Osunsade, A.; Agustinus, A.S.; Finkin-Groner, E.; D’Ambrosio, H.; Liu, B.; Chandarlapaty, S.; Liu, S.; et al. Reversible histone glycation is associated with disease-related changes in chromatin architecture. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Rustagi, Y.; Abouhashem, A.S.; Tabasum, S.; Verma, P.; Hernandez, E.; Pal, D.; Khona, D.K.; Mohanty, S.K.; Kumar, M.; et al. Genome-wide DNA hypermethylation opposes healing in patients with chronic wounds by impairing epithelial-mesenchymal transition. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]
- Li, J.; Wei, M.; Liu, X.; Xiao, S.; Cai, Y.; Li, F.; Tian, J.; Qi, F.; Xu, G.; Deng, C. The progress, prospects, and challenges of the use of non-coding RNA for diabetic wounds. Mol. Ther. Nucleic Acids 2021, 24, 554–578. [Google Scholar] [CrossRef]
- El-Osta, A.; Brasacchio, D.; Yao, D.; Pocai, A.; Jones, P.L.; Roeder, R.G.; Cooper, M.E.; Brownlee, M. Transient high glucose causes persistent epigenetic changes and altered gene expression during subsequent normoglycemia. J. Exp. Med. 2008, 205, 2409–2417. [Google Scholar] [CrossRef]
- Bhamidipati, T.; Kumar, M.; Verma, S.S.; Mohanty, S.K.; Kacar, S.; Reese, D.; Martinez, M.M.; Kanocka, M.M.; Dunn, K.W.; Sen, C.K.; et al. Epigenetic basis of diabetic vasculopathy. Front. Endocrinol. 2022, 13, 989844. [Google Scholar] [CrossRef]
- Goodarzi, G.; Maniati, M.; Qujeq, D. The role of microRNAs in the healing of diabetic ulcers. Int. Wound J. 2019, 16, 621–633. [Google Scholar] [CrossRef]
- Ozdemir, D.; Feinberg, M.W. MicroRNAs in diabetic wound healing: Pathophysiology and therapeutic opportunities. Trends Cardiovasc. Med. 2018, 29, 131–137. [Google Scholar] [CrossRef]
- Lou, R.; Chen, J.; Zhou, F.; Wang, C.; Leung, C.-H.; Lin, L. Exosome-cargoed microRNAs: Potential therapeutic molecules for diabetic wound healing. Drug Discov. Today 2022, 27, 103323. [Google Scholar] [CrossRef] [PubMed]
- Ross, K. MiR equal than others: MicroRNA enhancement for cutaneous wound healing. J. Cell. Physiol. 2021, 236, 8050–8059. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, C.; Wang, X.Y.; Zhou, L.Y.; Lao, G.J.; Liu, D.; Wang, C.; Hu, M.D.; Zeng, T.T.; Yan, L.; et al. MicroRNA-129 and -335 Promote Diabetic Wound Healing by Inhibiting Sp1-Mediated MMP-9 Expression. Diabetes 2018, 67, 1627–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Wu, W.; Zhang, L.; Dorset-Martin, W.; Morris, M.W.; Mitchell, M.E.; Liechty, K.W. The role of microRNA-146a in the pathogenesis of the diabetic wound-healing impairment: Correction with mesenchymal stem cell treatment. Diabetes 2012, 61, 2906–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Li, Y.; Tang, Y.; Zhao, X.; Xie, D.; Chen, M. Increased Expression of miR-155 in Peripheral Blood and Wound Margin Tissue of Type 2 Diabetes Mellitus Patients Associated with Diabetic Foot Ulcer. Diabetes Metab. Syndr. Obes. 2022, 15, 3415–3428. [Google Scholar] [CrossRef]
- Liu, J.; Xu, Y.; Shu, B.; Wang, P.; Tang, J.; Chen, L.; Qi, S.; Liu, X.; Xie, J. Quantification of the differential expression levels of microRNA-203 in different degrees of diabetic foot. Int. J. Clin. Exp. Pathol. 2015, 8, 13416–13420. [Google Scholar]
- Wu, T.; Xie, D.; Zhao, X.; Xu, M.; Luo, L.; Deng, D.; Chen, M. Enhanced Expression of miR-34c in Peripheral Plasma Associated with Diabetic Foot Ulcer in Type 2 Diabetes Patients. Diabetes Metab. Syndr. Obes. 2021, 14, 4263–4273. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Kuai, L.; Jiang, J.; Li, W.; Li, B.; Yin, S. Long non-coding RNAs in diabetic wound healing: Current research and clinical relevance. Int. Wound J. 2022, 19, 583–600. [Google Scholar] [CrossRef]
- Shen, J.; Zhao, X.; Zhong, Y.; Yang, P.; Gao, P.; Wu, X.; Wang, X.; An, W. Exosomal ncRNAs: The pivotal players in diabetic wound healing. Front. Immunol. 2022, 13, 1005307. [Google Scholar] [CrossRef] [PubMed]
- Pirola, L. Epigenetics of Diabetic Microvascular Disease. Microvasc. Dis. Diabetes 2020, 45–57. [Google Scholar]
- Legiawati, L. The Role of Oxidative Stress, Inflammation, and Advanced Glycation End Product in Skin Manifestations of Diabetes Mellitus. Curr. Diabetes Rev. 2022, 18, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.; Moellmer, R.; Agrawal, D.K. Role of fibroblast plasticity and heterogeneity in modulating angiogenesis and healing in the diabetic foot ulcer. Mol. Biol. Rep. 2022, 50, 1913–1929. [Google Scholar] [CrossRef]
- Liechty, C.; Hu, J.; Zhang, L.; Liechty, K.W.; Xu, J. Role of microRNA-21 and Its Underlying Mechanisms in Inflammatory Responses in Diabetic Wounds. Int. J. Mol. Sci. 2020, 21, 3328. [Google Scholar] [CrossRef] [PubMed]
- Choi, L.-S.; Ahmed, K.; Kim, Y.-S.; Yim, J.-E. Skin accumulation of advanced glycation end products and cardiovascular risk in Korean patients with type 2 diabetes mellitus. Heliyon 2022, 8, e09571. [Google Scholar] [CrossRef]
- Zheng, W.; Li, H.; Go, Y.; Chan, X.H.; Huang, Q.; Wu, J. Research Advances on the Damage Mechanism of Skin Glycation and Related Inhibitors. Nutrients 2022, 14, 4588. [Google Scholar] [CrossRef]
- Deng, L.; Du, C.; Song, P.; Chen, T.; Rui, S.; Armstrong, D.G.; Deng, W. The Role of Oxidative Stress and Antioxidants in Diabetic Wound Healing. Oxidative Med. Cell. Longev. 2021, 2021, 1–11. [Google Scholar] [CrossRef]
- Al-Rikabi, A.H.A.; Tobin, D.J.; Riches-Suman, K.; Thornton, M.J. Dermal fibroblasts cultured from donors with type 2 diabetes mellitus retain an epigenetic memory associated with poor wound healing responses. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Miwa, S.; Kashyap, S.; Chini, E.; von Zglinicki, T. Mitochondrial dysfunction in cell senescence and aging. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]
- Pinti, M.V.; Fink, G.K.; Hathaway, Q.A.; Durr, A.J.; Kunovac, A.; Hollander, J.M. Mitochondrial dysfunction in type 2 diabetes mellitus: An organ-based analysis. Am. J. Physiol. Metab. 2019, 316, E268–E285. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in skin health, aging, and disease. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Kashpur, O.; Smith, A.; Gerami-Naini, B.; Maione, A.G.; Calabrese, R.; Tellechea, A.; Theocharidis, G.; Liang, L.; Pastar, I.; Tomic-Canic, M.; et al. Differentiation of diabetic foot ulcer-derived induced pluripotent stem cells reveals distinct cellular and tissue phenotypes. FASEB J. 2018, 33, 1262–1277. [Google Scholar] [CrossRef]
- Xu, S.; Li, S.; Bjorklund, M.; Xu, S. Mitochondrial fragmentation and ROS signaling in wound response and repair. Cell Regen. 2022, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Lu, X.; Yang, Y.; Yang, Y.; Li, Y.; Kuai, L.; Li, B.; Dong, H.; Shi, J. Microenvironment-Based Diabetic Foot Ulcer Nanomedicine. Adv. Sci. 2022, 10, e2203308. [Google Scholar] [CrossRef]
- Cabal, M.C.; González, D.E.; Berlanga-Acosta, J.; Zayas, D.; Herrera, L.; Lopez-Saura, P.; Guillen, G.; Baldomero, J.; Delgado, C.G.; Gonzalez, L.J. Quantitative Studies of the Evolution of Diabetic Foot Lesions under EGF Treatment by Magnetic Resonance Imaging. J. Radiol. Res. Pract. 2014, 2014. [Google Scholar]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced Glycation End Products and Diabetes Mellitus: Mechanisms and Perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef] [PubMed]
- Tatmatsu-Rocha, J.C.; Tim, C.R.; Avo, L.; Bernardes-Filho, R.; Brassolatti, P.; Kido, H.W.; Hamblin, M.R.; Parizotto, N.A. Mitochondrial dynamics (fission and fusion) and collagen production in a rat model of diabetic wound healing treated by photobiomodulation: Comparison of 904 nm laser and 850 nm light-emitting diode (LED). J. Photochem. Photobiol. B Biol. 2018, 187, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Sacco, A.M.; Belviso, I.; Romano, V.; Carfora, A.; Schonauer, F.; Nurzynska, D.; Montagnani, S.; Di Meglio, F.; Castaldo, C. Diversity of dermal fibroblasts as major determinant of variability in cell reprogramming. J. Cell Mol. Med. 2019, 23, 4256–4268. [Google Scholar] [CrossRef]
- Kirk, T.; Ahmed, A.; Rognoni, E. Fibroblast Memory in Development, Homeostasis and Disease. Cells 2021, 10, 2840. [Google Scholar] [CrossRef]
- Li, B.; Bian, X.; Hu, W.; Wang, X.; Li, Q.; Wang, F.; Sun, M.; Ma, K.; Zhang, C.; Fu, X.; et al. Regenerative and protective effects of calcium silicate on senescent fibroblasts induced by high glucose. Wound Repair Regen. 2020, 28, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Phang, S.J.; Arumugam, B.; Kuppusamy, U.R.; Fauzi, M.B.; Looi, M.L. A review of diabetic wound models-Novel insights into diabetic foot ulcer. J. Tissue Eng. Regen. Med. 2021, 15, 1051–1068. [Google Scholar] [CrossRef] [PubMed]
- Maitra, S.; Dutta, D. Downregulation of Hexose Sugar Metabolism in Diabetes Decreases the Rate of Wound Healing; Academic Press: Cambridge, MA, USA, 2020; pp. 259–270. [Google Scholar]
- Soydas, T.; Sayitoglu, M.; Sarac, E.Y.; Cınar, S.; Solakoglu, S.; Tiryaki, T.; Sultuybek, G.K. Metformin reverses the effects of high glucose on human dermal fibroblasts of aged skin via downregulating RELA/p65 expression. J. Physiol. Biochem. 2021, 77, 443–450. [Google Scholar] [CrossRef]
- Vujčić, S.; Kotur-Stevuljević, J.; Vekić, J.; Perović-Blagojević, I.; Stefanović, T.; Ilić-Mijailović, S.; Uzelac, B.K.; Bosic, S.; Antonic, T.; Guzonjic, A.; et al. Oxidative Stress and Inflammatory Biomarkers in Patients with Diabetic Foot. Medicina 2022, 58, 1866. [Google Scholar] [CrossRef] [PubMed]
- Theocharidis, G.; Baltzis, D.; Roustit, M.; Tellechea, A.; Dangwal, S.; Khetani, R.S.; Shu, B.; Zhao, W.; Fu, J.; Bhasin, S.; et al. Integrated Skin Transcriptomics and Serum Multiplex Assays Reveal Novel Mechanisms of Wound Healing in Diabetic Foot Ulcers. Diabetes 2020, 69, 2157–2169. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.-C.; Lan, C.-C.E. The Epidermal Keratinocyte as a Therapeutic Target for Management of Diabetic Wounds. Int. J. Mol. Sci. 2023, 24, 4290. [Google Scholar] [CrossRef]
- Dhariwala, M.O.; Scharschmidt, T.C. Baby’s skin bacteria: First impressions are long-lasting. Trends Immunol. 2021, 42, 1088–1099. [Google Scholar] [CrossRef]
- Haftek, M. ‘Memory’ of the stratum corneum: Exploration of the epidermis’ past. Br. J. Dermatol. 2014, 171 (Suppl. S3), 6–9. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Huang, S.-M.; Cheng, Y.-W.; Yen, M.-C.; Hsu, Y.-L.; Lan, C.-C.E. Investigation of the keratinocyte transcriptome altered in high-glucose environment: An in-vitro model system for precision medicine. J. Dermatol. Sci. 2023, 109, 37–46. [Google Scholar] [CrossRef]
- Cibrian, D.; de la Fuente, H.; Sánchez-Madrid, F. Metabolic Pathways That Control Skin Homeostasis and Inflammation. Trends Mol. Med. 2020, 26, 975–986. [Google Scholar] [CrossRef]
- Mansoub, N.H. The role of keratinocyte function on the defected diabetic wound healing. Int. J. Burns Trauma. 2021, 11, 430–441. [Google Scholar]
- Lan, C.-C.E.; Huang, S.-M.; Wu, C.-S.; Wu, C.-H.; Chen, G.-S. High-glucose environment increased thrombospondin-1 expression in keratinocytes via DNA hypomethylation. Transl. Res. 2016, 169, 91–101.e3. [Google Scholar] [CrossRef]
- Li, D.; Kular, L.; Vij, M.; Herter, E.K.; Li, X.; Wang, A.; Chu, T.; Toma, M.-A.; Zhang, L.; Liapi, E.; et al. Human skin long noncoding RNA WAKMAR1 regulates wound healing by enhancing keratinocyte migration. Proc. Natl. Acad. Sci. USA 2019, 116, 9443–9452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Chen, C.; Wu, D.; Chen, G.; Tan, R.; Ran, J. AGEs-induced MMP-9 activation mediated by Notch1 signaling is involved in impaired wound healing in diabetic rats. Diabetes Res. Clin. Pr. 2022, 186, 109831. [Google Scholar] [CrossRef]
- Tan, Q.; Wang, W.; Yang, C.; Zhang, J.; Sun, K.; Luo, H.C.; Mai, L.F.; Lao, Y.; Yan, L.; Ren, M. alpha-ketoglutarate is associated with delayed wound healing in diabetes. Clin. Endocrinol. 2016, 85, 54–61. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, P.; Liu, D.; Gao, M.; Wang, J.; Wang, X.; Liu, Y.; Zhang, X. c-Myc Upregulated by High Glucose Inhibits HaCaT Differentiation by S100A6 Transcriptional Activation. Front. Endocrinol. 2021, 12, 676403. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Brem, H.; Vouthounis, C.; Lee, B.; Fallon, J.; Stallcup, M.; Merchant, A.; Galiano, R.D.; Tomic-Canic, M. Molecular pathogenesis of chronic wounds: The role of beta-catenin and c-myc in the inhibition of epithelialization and wound healing. Am. J. Pathol. 2005, 167, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Graves, D.T. Keratinocyte Function in Normal and Diabetic Wounds and Modulation by FOXO1. J. Diabetes Res. 2020, 2020, 1–9. [Google Scholar]
- Fu, X.; Ding, H.; Miao, W.; Mao, C.; Zhan, M.; Chen, H. Global recurrence rates in diabetic foot ulcers: A systematic review and meta-analysis. Diabetes Metab. Res. Rev. 2019, 35, e3160. [Google Scholar] [CrossRef]
- Bus, S.A.; van Netten, J.J.; Lavery, L.A.; Monteiro-Soares, M.; Rasmussen, A.; Jubiz, Y.; Price, P.E.; International Working Group on the Diabetic Foot. IWGDF guidance on the prevention of foot ulcers in at-risk patients with diabetes. Diabetes Metab. Res. Rev. 2016, 32 (Suppl. S1), 16–24. [Google Scholar] [CrossRef] [Green Version]
- Mahdipour, E.; Sahebkar, A. The Role of Recombinant Proteins and Growth Factors in the Management of Diabetic Foot Ulcers: A Systematic Review of Randomized Controlled Trials. J. Diabetes Res. 2020, 2020, 6320514. [Google Scholar] [CrossRef] [PubMed]
- Berlanga-Acosta, J.; Fernandez, M.J.; Santana, H.; Pérez, C.; Savigne, W.; Mendoza-Mari, Y.; Garcia-Ojalvo, A.; de Leon Perez, L.P.; Pena, Y.O.; Acosta, O.R.; et al. A narrative review on Epidermal Growth Factor (EGF) intralesional infiltrations for diabetic complex wounds: The rational of an innovative delivery route. Vasc. Dis. Ther. 2022, 7, 1–9. [Google Scholar] [CrossRef]
- Ferrer-Tasies, L.; Santana, H.; Cabrera-Puig, I.; González-Mira, E.; Ballell-Hosa, L.; Castellar-Álvarez, C.; Cordoba, A.; Merlo-Mas, J.; Geronimo, H.; Chinea, G.; et al. Recombinant Human Epidermal Growth Factor/Quatsome Nanoconjugates: A Robust Topical Delivery System for Complex Wound Healing. Adv. Ther. 2021, 4, 2000260. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.; Camacho-Rodriguez, H.; Mendoza-Mari, Y.; Falcon-Cama, V.; Garcia-Ojalvo, A.; Herrera-Martinez, L.; Guillen-Nieto, G. Epidermal Growth Factor in Healing Diabetic Foot Ulcers: From Gene Expression to Tissue Healing and Systemic Biomarker Circulation. MEDICC Rev. 2020, 22, 24–31. [Google Scholar]
- Matoori, S.; Veves, A.; Mooney, D.J. Advanced bandages for diabetic wound healing. Sci. Transl. Med. 2021, 13. [Google Scholar] [CrossRef]
- Zidarič, T.; Kleinschek, K.S.; Maver, U.; Maver, T. (Eds.) Commercial Skin Equivalents. In Function-Oriented Bioengineered Skin Equivalents: Continuous Development Towards Complete Skin Replication; Springer International Publishing: Berlin/Heidelberg, Germany, 2023; pp. 103–122. [Google Scholar]
- Sarkar, N.; Bhumiratana, S.; Geris, L.; Papantoniou, I.; Grayson, W.L. Bioreactors for engineering patient-specific tissue grafts. Nat. Rev. Bioeng. 2023, 1, 361–377. [Google Scholar] [CrossRef]
- Heras, K.L.; Igartua, M.; Santos-Vizcaino, E.; Hernandez, R.M. Cell-based dressings: A journey through chronic wound management. Biomater. Adv. 2022, 135, 212738. [Google Scholar] [CrossRef]
- Nilforoushzadeh, M.A.; Amirkhani, M.A.; Khodaverdi, E.; Razzaghi, Z.; Afzali, H.; Izadpanah, S.; Zare, S. Tissue engineering in dermatology—From lab to market. Tissue Cell. 2022, 74, 101717. [Google Scholar] [CrossRef]
- Wynn, M.; Freeman, S. The efficacy of negative pressure wound therapy for diabetic foot ulcers: A systematised review. J. Tissue Viability 2019, 28, 152–160. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.; Hou, L.; Yan, C. Vacuum-assisted closure versus conventional dressing in necrotizing fasciitis: A systematic review and meta-analysis. J. Orthop. Surg. Res. 2023, 18, 1–9. [Google Scholar] [CrossRef]
- Wieman, T.J.; Smiell, J.M.; Su, Y. Efficacy and safety of a topical gel formulation of recombinant human platelet-derived growth factor-BB (becaplermin) in patients with chronic neuropathic diabetic ulcers. A phase III randomized placebo-controlled double-blind study. Diabetes Care 1998, 21, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Bui, T.Q.; Bui, Q.V.P.; Németh, D.; Hegyi, P.; Szakács, Z.; Rumbus, Z.; Toth, B.; Emri, G.; Parniczky, A.; Sarlos, P.; et al. Epidermal Growth Factor is Effective in the Treatment of Diabetic Foot Ulcers: Meta-Analysis and Systematic Review. Int. J. Environ. Res. Public Health 2019, 16, 2584. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, V.; Pendsey, S.; Sekar, N.; Murthy, G.S.R. A phase III study to evaluate the safety and efficacy of recombinant human epidermal growth factor (REGEN-D™ 150) in healing diabetic foot ulcers. Wounds 2006, 18, 186–196. [Google Scholar]
- Al-Jalodi, O.; Kupcella, M.; Breisinger, K.; Serena, T.E. A multicenter clinical trial evaluating the durability of diabetic foot ulcer healing in ulcers treated with topical oxygen and standard of care versus standard of care alone 1 year post healing. Int. Wound J. 2022, 19, 1838–1842. [Google Scholar] [CrossRef] [PubMed]
- Tsang, M.W.; Wong, W.K.R.; Hung, C.S.; Lai, K.-M.; Tang, W.; Cheung, E.Y.N.; Kam, G.; Leung, L.; Chan, C.W.; Chu, C.M.; et al. Human epidermal growth factor enhances healing of diabetic foot ulcers. Diabetes Care 2003, 26, 1856–1861. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.P.; Jung, H.D.; Kim, Y.W. Recombinant human epidermal growth factor (EGF) to enhance healing for diabetic foot ulcers. Ann. Plast. Surg. 2006, 56, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Han, S.H.; Hong, J.P.; Lee, D.-H.; Kim, B.S.; Ahn, J.H.; Lee, J.W. Topical epidermal growth factor spray for the treatment of chronic diabetic foot ulcers: A phase III multicenter, double-blind, randomized, placebo-controlled trial. Diabetes Res. Clin. Pr. 2018, 142, 335–344. [Google Scholar] [CrossRef]
- Le Tuyet, H.; Quynh, T.T.N.; Minh, H.V.H.; Bich, D.N.T.; Dinh, T.D.; Le Tan, D.; Van, H.L.; Le Huy, T.; Huu, H.D.; Trong, T.N.T. The efficacy and safety of epidermal growth factor in treatment of diabetic foot ulcers: The preliminary results. Int. Wound J. 2009, 6, 159–166. [Google Scholar] [CrossRef]
- Fernandez-Montequin, J.I.; Valenzuela-Silva, C.M.; Diaz, O.G.; Savigne, W.; Sancho-Soutelo, N.; Rivero-Fernandez, F.; Sanchez-Penton, P.; Morejon-Vega, L.; Artaza-Sanz, H.; Garcia-Herrera, A.; et al. Intra-lesional injections of recombinant human epidermal growth factor promote granulation and healing in advanced diabetic foot ulcers: Multicenter, randomised, placebo-controlled, double-blind study. Int. Wound J. 2009, 6, 432–443. [Google Scholar] [CrossRef]
- Gomez-Villa, R.; Aguilar-Rebolledo, F.; Lozano-Platonoff, A.; Teran-Soto, J.M.; Fabian-Victoriano, M.R.; Kresch-Tronik, N.S.; Garrido-Espindola, X.; Garcia-Solis, A.; Bondani-Guasti, A.; Bierzwinsky-Sneider, G.; et al. Response to Uzun G. Wound Repair Regen. 2014, 22, 767. [Google Scholar] [CrossRef]
- Bartın, M.K.; Okut, G. The effect of human recombinant epidermal growth factor in the treatment of diabetic foot ulcers. J. Clin. Investig. Surg. 2022, 7, 38–42. [Google Scholar] [CrossRef]
- Yera-Alos, I.B.; Alonso-Carbonell, L.; Valenzuela-Silva, C.M.; Tuero-Iglesias, A.D.; Moreira-Martinez, M.; Marrero-Rodriguez, I.; Lopez-Mola, E.; Lopez-Saura, P.A. Active post-marketing surveillance of the intralesional administration of human recombinant epidermal growth factor in diabetic foot ulcers. BMC Pharmacol. Toxicol. 2013, 14, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yera-Alos, I.B.; A López-Saura, P.; Valenzuela-Silva, C.; González-Díaz, O.; Del Río-Martín, A.; Berlanga-Acosta, J.; Fernandez-Montequin, J.I.; Acevedo-Castro, B.; Lopez-Mola, E.; Herrera-martinez, L. Medical Practice Confirms Clinical Trial Results of the Use of Intralesional Human Recombinant Epidermal Growth Factor in Advanced Diabetic Foot Ulcers. Adv. Pharmacoepidemiol. Drug Saf. 2013, 2. [Google Scholar]
- Kahraman, M.; Misir, A.; Kizkapan, T.B.; Ozcamdalli, M.; Uzun, E.; Mutlu, M. The Long-Term Outcomes Following the Application of Intralesional Epidermal Growth Factor in Patients with Diabetic Foot Ulcers. J. Foot Ankle Surg. 2019, 58, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Embil, J.M.; Papp, K.; Sibbald, G.; Tousignant, J.; Smiell, J.M.; Wong, B.; Lau, C.Y. Recombinant human platelet-derived growth factor-BB (becaplermin) for healing chronic lower extremity diabetic ulcers: An open-label clinical evaluation of efficacy. Wound Repair Regen. 2000, 8, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Smiell, J.M.; Wieman, T.J.; Steed, D.L.; Perry, B.H.; Sampson, A.R.; Schwab, B.H. Sampson AR, Schwab BH. Efficacy and safety of becaplermin (recombinant human platelet-derived growth factor-BB) in patients with nonhealing, lower extremity diabetic ulcers: A combined analysis of four randomized studies. Wound Repair Regen. 1999, 7, 335–346. [Google Scholar] [CrossRef]
- Ma, C.; A Hernandez, M.; E Kirkpatrick, V.; Liang, L.-J.; Nouvong, A.L.; Gordon, I. Topical platelet-derived growth factor vs placebo therapy of diabetic foot ulcers offloaded with windowed casts: A randomized, controlled trial. Wounds 2015, 27, 83–91. [Google Scholar]
- Veves, A.; Falanga, V.; Armstrong, D.G.; Sabolinski, M.L.; Apligraf Diabetic Foot Ulcer Study. Graftskin, a human skin equivalent, is effective in the management of noninfected neuropathic diabetic foot ulcers: A prospective randomized multicenter clinical trial. Diabetes Care 2001, 24, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Marston, W.A.; Hanft, J.; Norwood, P.; Pollak, R. Dermagraft Diabetic Foot Ulcer Study G. The efficacy and safety of Dermagraft in improving the healing of chronic diabetic foot ulcers: Results of a prospective randomized trial. Diabetes Care 2003, 26, 1701–1705. [Google Scholar] [CrossRef] [Green Version]
- Zelen, C.M.; Serena, T.E.; Gould, L.; Le, L.; Carter, M.J.; Keller, J.; Li, W.W. Treatment of chronic diabetic lower extremity ulcers with advanced therapies: A prospective, randomised, controlled, multi-centre comparative study examining clinical efficacy and cost. Int. Wound J. 2016, 13, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Zelen, C.M.; Serena, T.E.; Fetterolf, D.E. Dehydrated human amnion/chorion membrane allografts in patients with chronic diabetic foot ulcers: A long-term follow-up study. Wound Med. 2014, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Blume, P.A.; Walters, J.; Payne, W.; Ayala, J.; Lantis, J. Comparison of negative pressure wound therapy using vacuum-assisted closure with advanced moist wound therapy in the treatment of diabetic foot ulcers: A multicenter randomized controlled trial. Diabetes Care 2008, 31, 631–636. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.G.; A Lavery, L. Diabetic Foot Study C. Negative pressure wound therapy after partial diabetic foot amputation: A multicentre, randomised controlled trial. Lancet 2005, 366, 1704–1710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Z.C.; Chen, D.; Guo, D.; Zhu, J.Y.; Tang, B. Effectiveness and safety of negative-pressure wound therapy for diabetic foot ulcers: A meta-analysis. Plast. Reconstr. Surg. 2014, 134, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; He, C.-Z.; Cai, Y.-T.; Xing, Q.-P.; Guo, Y.-Z.; Chen, Z.-L.; Su, J.-L.; Yang, L.-P. Evaluation of negative-pressure wound therapy for patients with diabetic foot ulcers: Systematic review and meta-analysis. Ther. Clin. Risk Manag. 2017, 13, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Meloni, M.; Izzo, V.; Vainieri, E.; Giurato, L.; Ruotolo, V.; Uccioli, L. Management of negative pressure wound therapy in the treatment of diabetic foot ulcers. World J. Orthop. 2015, 6, 387–393. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Recombinant Epidermal Growth Factor (EGF) | ||||
|---|---|---|---|---|
| Reference | Major Outcome | Administration Route | Follow Up Period | Recurrences |
| Tsang, M.W. et al., 2003 [117] | 20 of 21 diabetic foot ulcers healed with daily application of 0.04% (wt/wt) hEGF for 12 weeks | Topical | 6 months | ND |
| Hong, J.P. et al., 2006 [118] | Topical treatment with EGF combined with advanced dressing may have positive effects in promoting healing | Topical | 6 months | No recurrences were observed in EGF group |
| Park, K.H. et al., 2018 [119] | The phase III study supports the efficacy and safety of spray-applied EGF treatment for DFUs | Topical | ND | ND |
| Viswanathan, V. et al., 2006 [115] | In EGF group, 90% of ulcers healed in 15 weeks compared with 22 weeks in control group | Topical | ND | ND |
| Tuyet HL et al. 2009 [120] | Topical EGF spray has positive effects on healing of moderate-to-severe foot ulcers | Topical | ND | ND |
| Fernández-Montequin, J. et al., 2009 [121] | Locally infiltrated EGF at 75 µg enhanced granulation tissue growth and wound closure | Intralesional injection | 12 months | No recurrences reported for EGF groups |
| Gomez-Villa, R. et al., 2014 [122] | Patients with DFU who received intralesional rhEGF application resulted in complete healing | Intralesional injection | ND | ND |
| Bartın, M. & Okut, G. 2022 [123] | Intralesional administration of EGF in T2DM can prevent amputations in DFU | Intralesional injection | 6 months | Two cases in the group receiving EGF |
| Yera-Alos, I.B. et al., 2013 [124] | Post-marketing study including 1788 patients treated with intranuclear-injected EGF. Re-epithelization was documented in 61% of the 1659 followed-up cases | Intralesional injection | 14 months | 5%/year |
| López-Saura, P.A. et al., 2013 [125] | Intralesional use of EGF for high-grade DFU in more than 2000 subjects. It confirms the 75% probability of complete granulation response, 61% healing, and a 16% absolute, and 71% relative reduction in amputation risk | Intralesional injection | 12 months | The frequency of relapseswas significantly lower (p < 0.001) in patients that received rhEGF: |
| Kahraman, M. et al., 2019 [126] | Study aimed to investigate the long-term outcomes after intralesional epidermal growth factor injections in the treatment of 34 diabetic patients with foot ulcers. | Intralesional injection | 60 months | Of 29 patients involved in the 5-year follow-up, 27 were ulcer free |
| Regranex or Becaplermin (rh-PDGF-BB) | ||||
| Reference | Major Outcome | Administration Route | Follow Up Period | Recurrences |
| Embil, J.M. et al., 2000 [127] | Confirms the efficacy and safety of becaplermin gel for the treatment of lower extremity diabetic ulcers | Topical | 6 months | 21% of recurrence in Becaplermin-treated patients |
| Smiell, J.M. et al., 1999 [128] | Becaplermin gel at a dose of 100 μg/g once daily is effective in patients with lower extremity diabetic ulcers | Topical | 3 months | ND |
| Wieman, T.J. et al., 1998 [113] | Becaplermin gel 100 μg/g significantly increased the incidence of complete wound closure | Topical | 3 months | Ulcer recurrence incidence was ≈30% in all the groups |
| Ma, C. et al., 2015 [129] | Topical platelet-derived growth factor does not appear to significantly improve healing of Wagner grade I diabetic foot ulcers | Topical | 6 months | No difference was observed between groups in recurrence |
| Cellular and Tissue-Based Products | ||||
| Reference | Major Outcome | Administration Route | Follow Up Period | Recurrences |
| Veves, A. et al., 2001 [130] | At the 12-week follow-up visit, 63 (56%) Graftskin-treated patients achieved complete wound healing compared with 36 (38%) in the control group (p = 0.0042). | Topical–bioengineered skin substitutes | 6 months | The incidence of ulcer recurrence was similar for Graftskin and control groups |
| Marston, W.A. et al., 2003 [131] | Patients experienced a significant clinical benefit when treated with Dermagraft versus patients treated with conventional therapy alone. | Topical–bioengineered skin substitutes | ND | ND |
| Zelen, C.M. et al., 2016 [132] | EpiFix® (dehydrated human amnion/chorion membrane) is superior to standard wound care SWC and Apligraf®, in achieving complete wound closure within 4–6 weeks. | Topical–bioengineered skin substitutes | ND | ND |
| Zelen, C.M. et al., 2014 [133] | DFU healed with use of dehydrated human amnion/chorion membrane (EpiFix) in 18 available subjects with healed DFU. Wound median size of 1.7 cm2. | Topical–bioengineered skin substitutes | 9–12 months | 17 wounds remained healed |
| Vacuum Assisted Closure (VAC)/Negative Pressure Wound Therapy (NPWT) | ||||
| Reference | Major Outcome | Administration Route | Follow Up Period | Recurrences |
| Blume, P. et al., 2008 [134] | A greater proportion of foot ulcers achieved complete ulcer closure with NPWT (73 of 169, 43.2%) than with advanced moist therapy within the 112-day active treatment | Topical–sub-atmospheric pressure over the wound area | ND | ND |
| Armstrong, D.G. et al., 2005 [135] | More patients healed in the NPWT group than in the control group (43 [56%] vs. 33 [39%], p = 0·040) | Topical–sub-atmospheric pressure over the wound area | ND | ND |
| Zhang, J. et al., 2014 [136] | Meta-analysis concludes that NPWT appears to be more effective for diabetic foot ulcers compared with non–negative-pressure wound therapy and has a similar safety profile. | Topical–sub-atmospheric pressure over the wound area | ND | ND |
| Liu, S. et al., 2017 [137] | Meta-analysis concludes that NPWT is efficacious, safe, and cost-effective in treating DFUs. | Topical–sub-atmospheric pressure over the wound area | ND | ND |
| Meloni, M. et al., 2015 [138] | A comprehensive review. DFUs managed with NPWT benefit from a significant reduction in the ulcer size and often a shorter treatment in comparison with ulcers treated with traditional gauze dressing | Topical–sub-atmospheric pressure over the wound area | ND | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berlanga-Acosta, J.; Garcia-Ojalvo, A.; Guillen-Nieto, G.; Ayala-Avila, M. Endogenous Biological Drivers in Diabetic Lower Limb Wounds Recurrence: Hypothetical Reflections. Int. J. Mol. Sci. 2023, 24, 10170. https://doi.org/10.3390/ijms241210170
Berlanga-Acosta J, Garcia-Ojalvo A, Guillen-Nieto G, Ayala-Avila M. Endogenous Biological Drivers in Diabetic Lower Limb Wounds Recurrence: Hypothetical Reflections. International Journal of Molecular Sciences. 2023; 24(12):10170. https://doi.org/10.3390/ijms241210170
Chicago/Turabian StyleBerlanga-Acosta, Jorge, Ariana Garcia-Ojalvo, Gerardo Guillen-Nieto, and Marta Ayala-Avila. 2023. "Endogenous Biological Drivers in Diabetic Lower Limb Wounds Recurrence: Hypothetical Reflections" International Journal of Molecular Sciences 24, no. 12: 10170. https://doi.org/10.3390/ijms241210170
APA StyleBerlanga-Acosta, J., Garcia-Ojalvo, A., Guillen-Nieto, G., & Ayala-Avila, M. (2023). Endogenous Biological Drivers in Diabetic Lower Limb Wounds Recurrence: Hypothetical Reflections. International Journal of Molecular Sciences, 24(12), 10170. https://doi.org/10.3390/ijms241210170