
Low-Field Magnetic Stimulation Alleviates MPTP-Induced Alterations in Motor Function and Dopaminergic Neurons in Male Mice


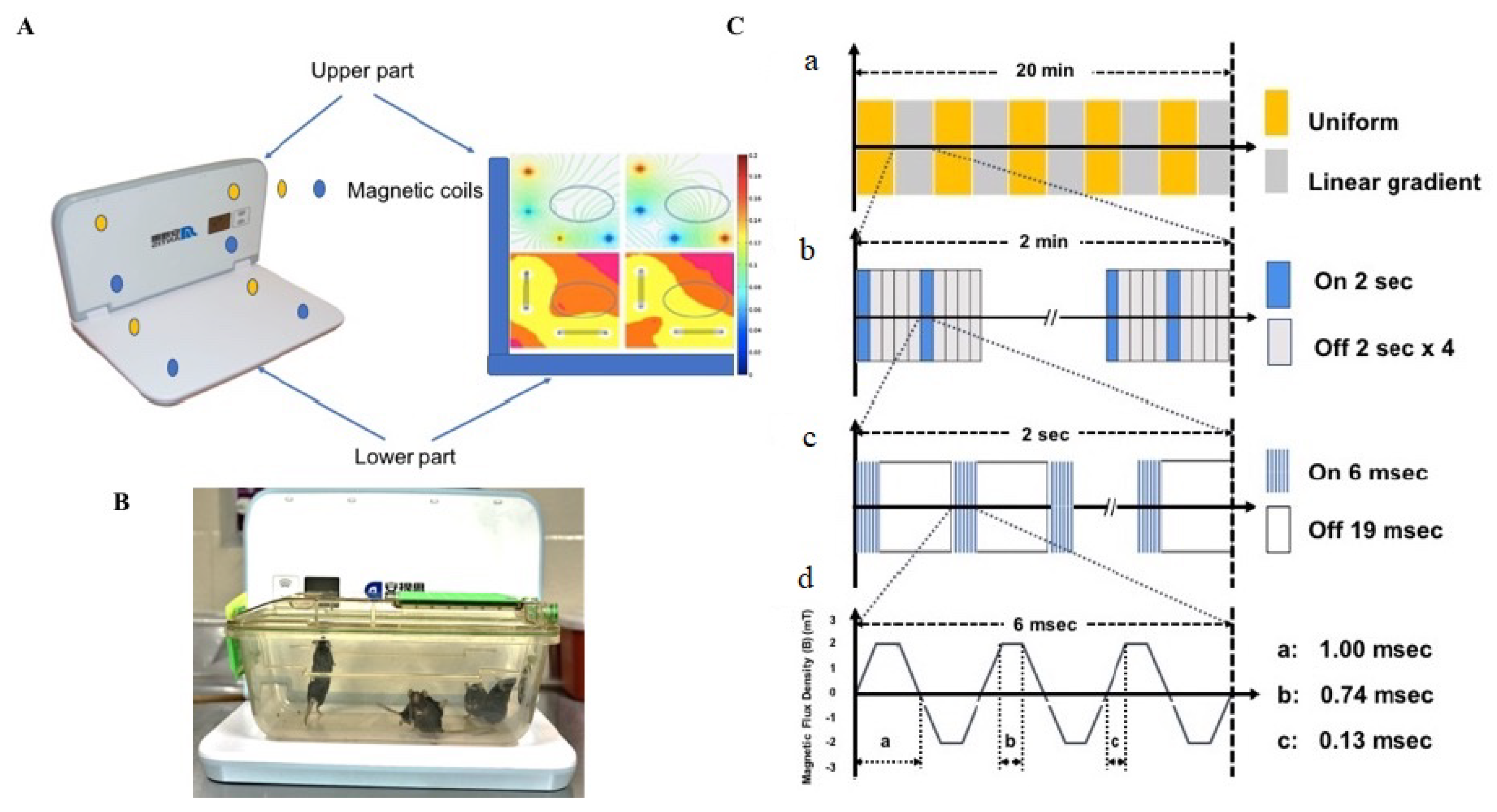{kind=link}
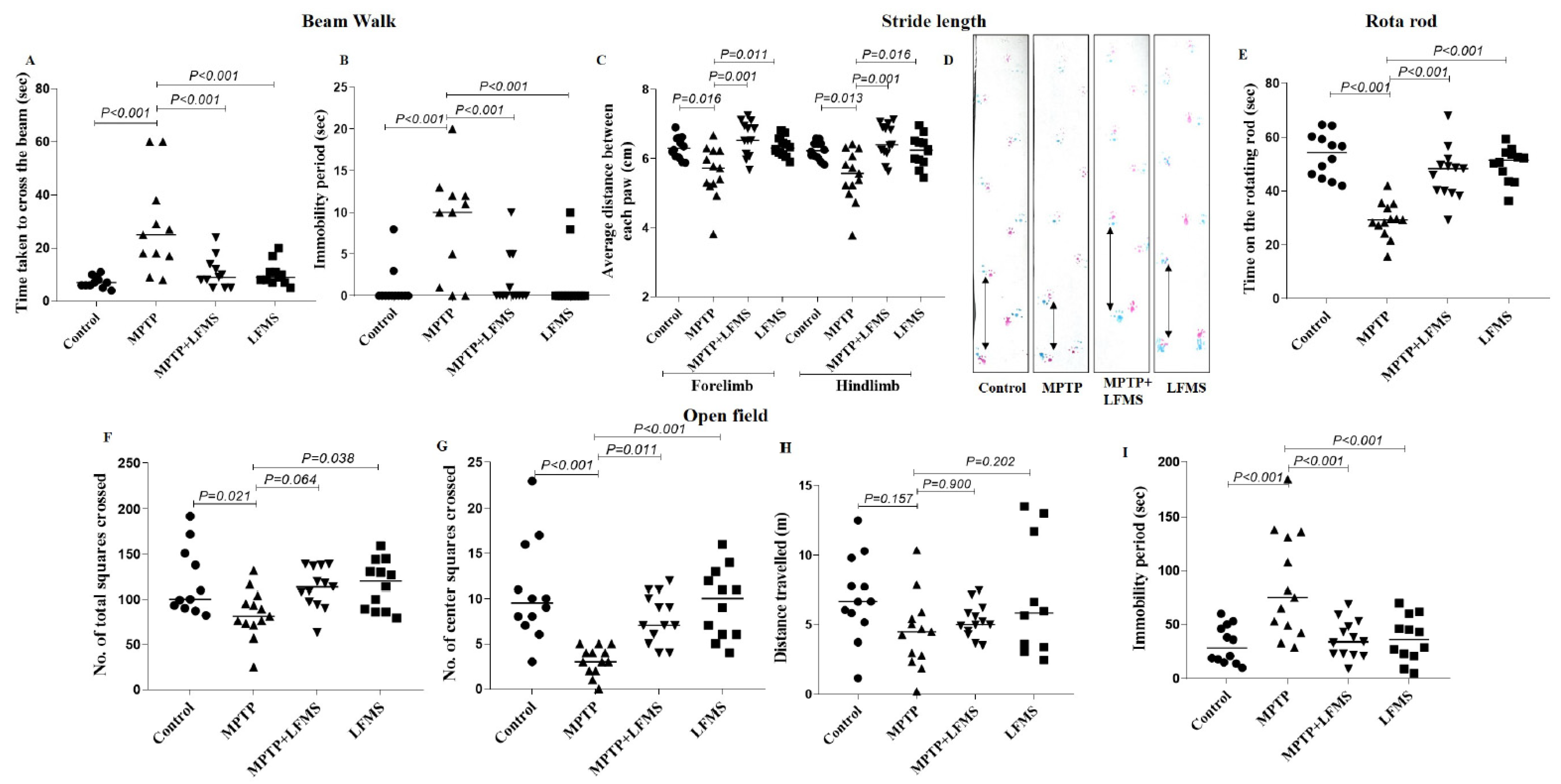{kind=link}
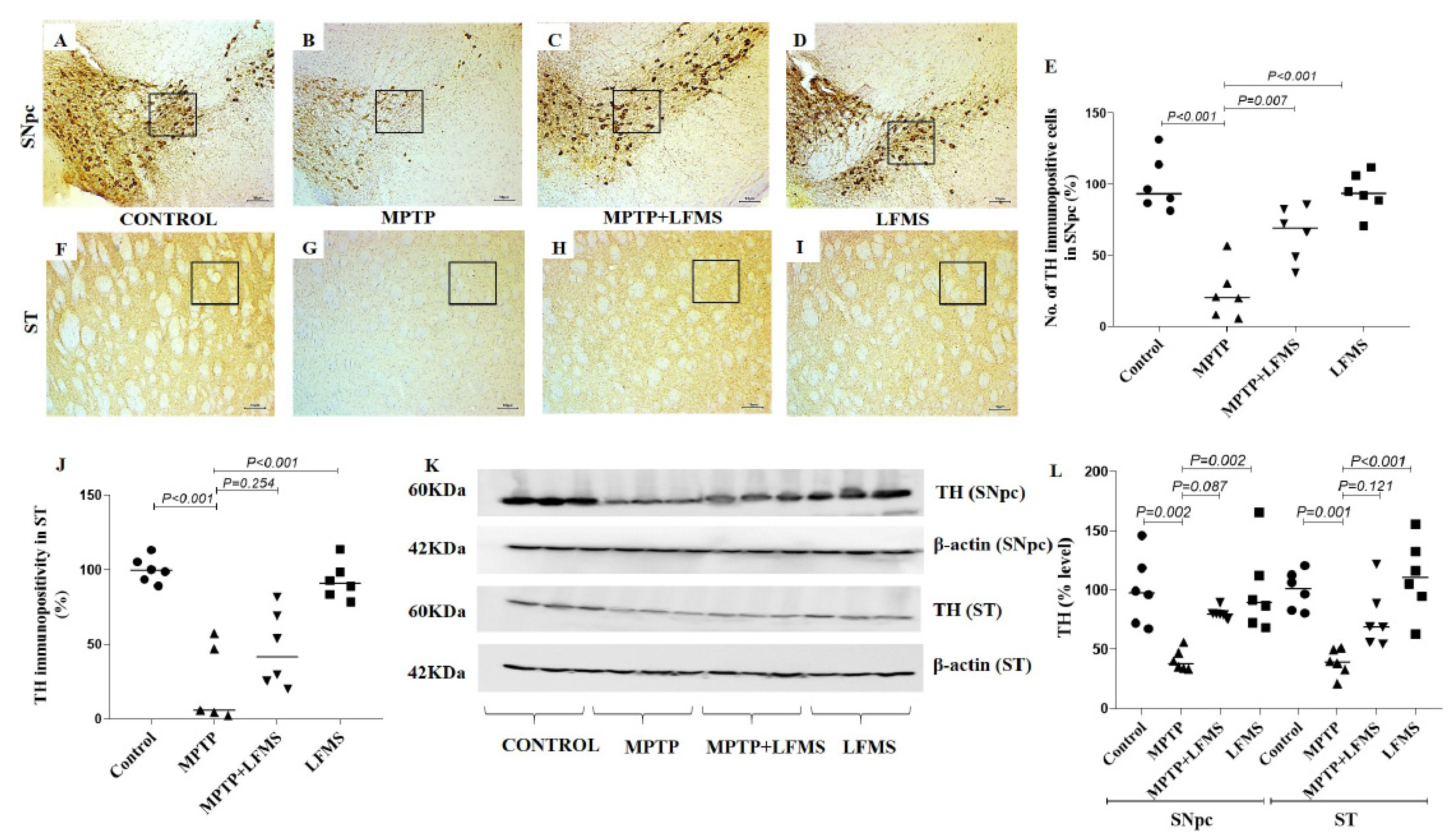{kind=link}
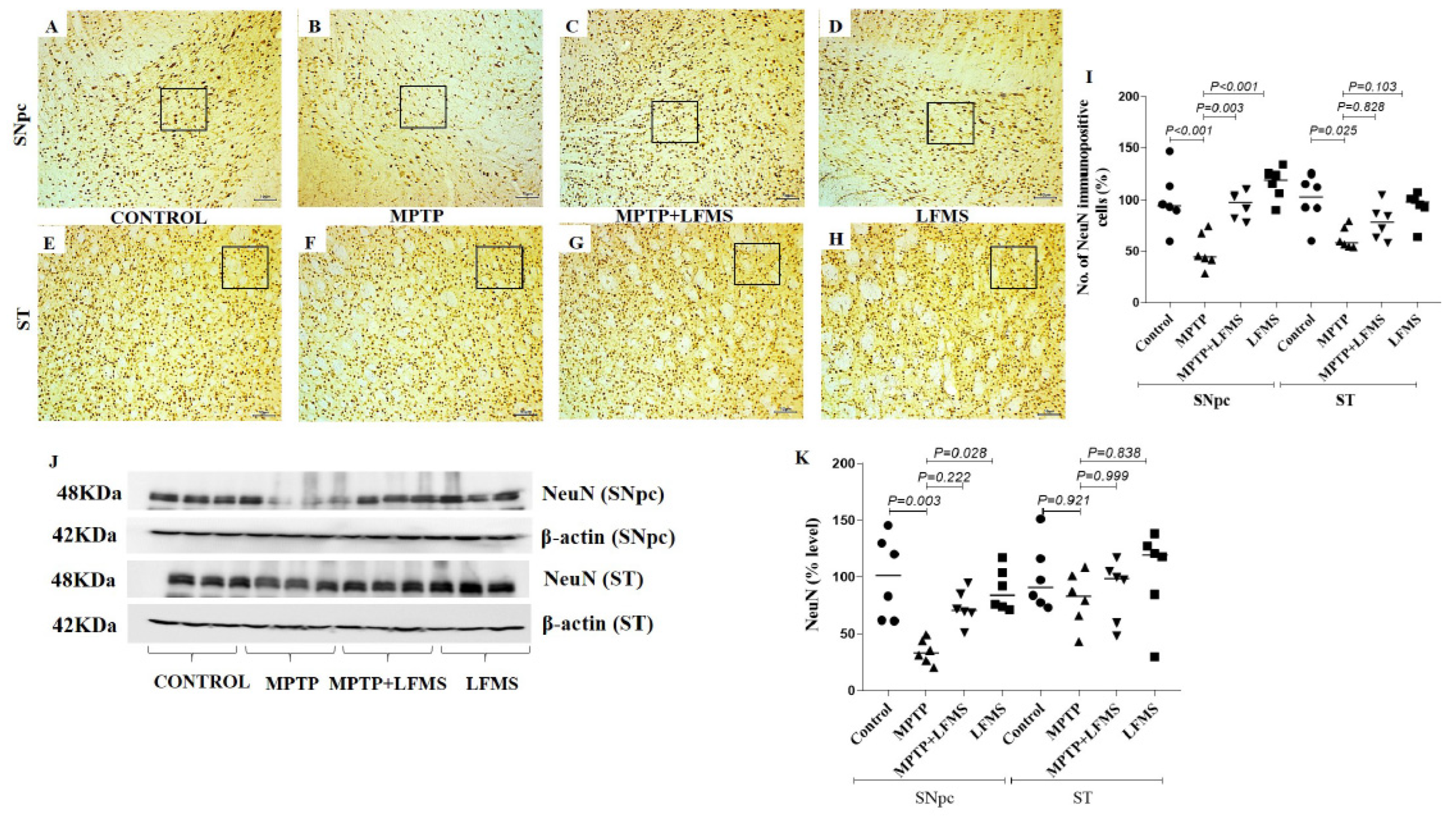{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. LFMS Treatment Improved Motor Function in MPTP-Treated Mice
2.1.1. Beam Walk Test
2.1.2. Stride Length
2.1.3. Rotarod Test
2.1.4. Open Field Locomotor Activity
2.2. LFMS Treatment Significantly Improved TH and NeuN Levels in the SNpc and ST Regions of MPTP-Treated Mouse Brain
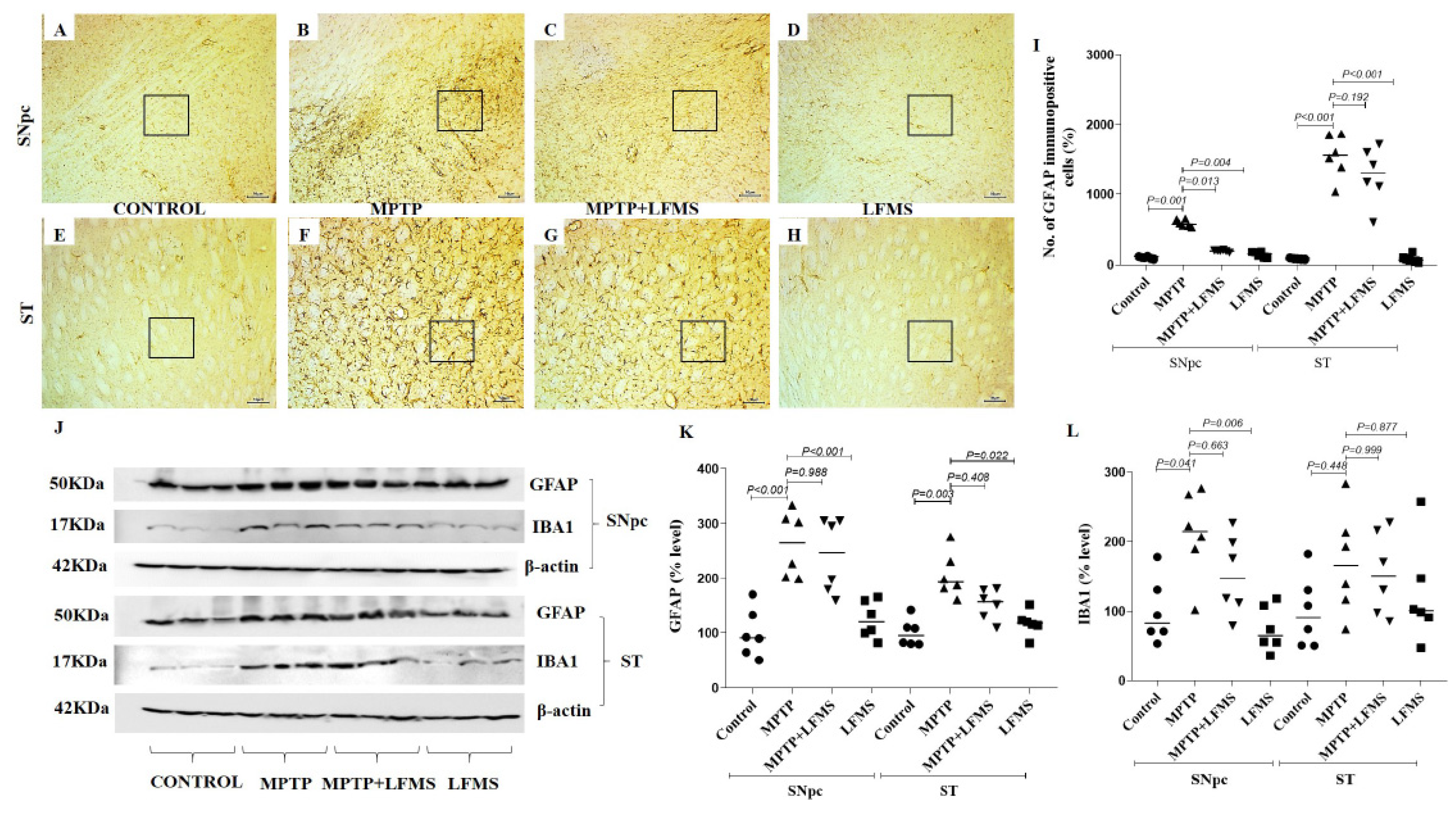
2.3. LFMS Treatment Reduced GFAP Level, Thereby Suppressing Gliosis in MPTP-Treated Mouse Brain
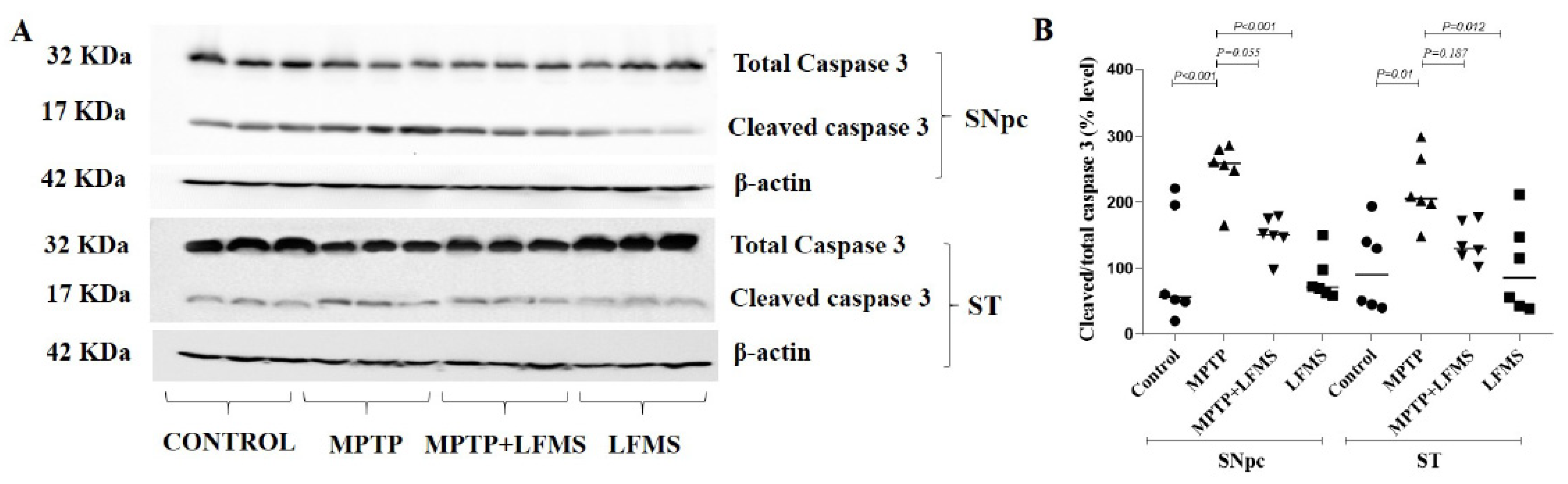
2.4. Effect of LFMS Treatment on Caspase-3 Activation in SNpc and ST Regions of MPTP-Treated Mouse Brains
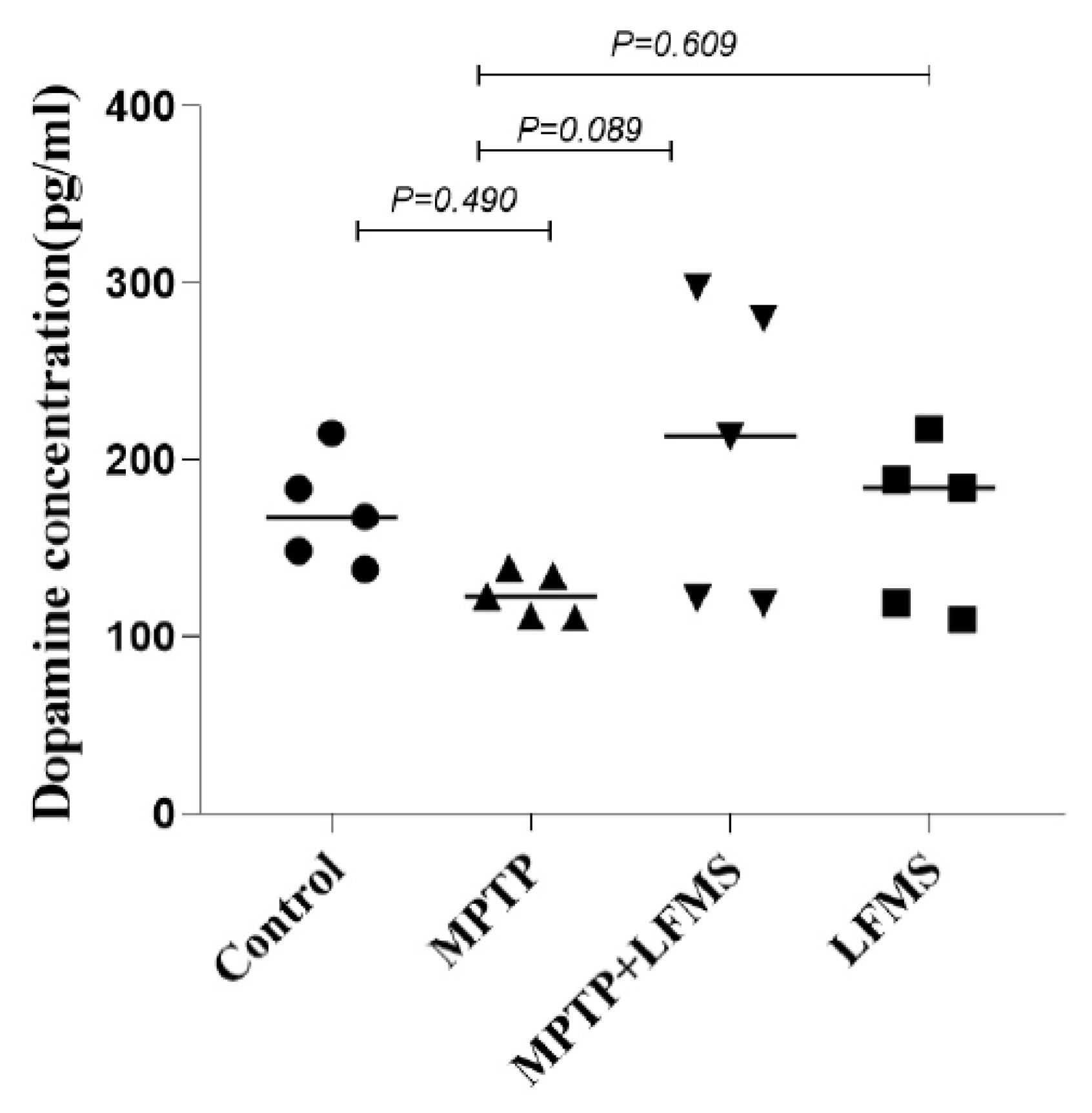
2.5. Effect of LFMS Treatment on Dopamine Level in the ST Region of MPTP Mouse Brain
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Experiment Design and Treatment
4.4. Motor Function Analysis
4.4.1. Beam Walk Test
4.4.2. Stride Length
4.4.3. Rotarod
4.4.4. Open Field Test
4.5. Western Blotting
4.6. Immunohistochemistry
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Lau, L.M.; Breteler, M.M. Epidemiology of Parkinson’s disease. Lancet Neurol. 2006, 5, 525–535. [Google Scholar] [CrossRef]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Kalia, L.V.; Lang, A.E. Parkinson disease in 2015: Evolving basic, pathological and clinical concepts in PD. Nat. Rev. Neurol. 2016, 12, 65–66. [Google Scholar] [CrossRef]
- Marino, B.L.B.; de Souza, L.R.; Sousa, K.P.A.; Ferreira, J.V.; Padilha, E.C.; da Silva, C.H.T.P.; Taft, C.A.; Hage-Melim, L.I.S. Parkinson’s disease: A Review from the Pathophysiology to Diagnosis, New Perspectives for Pharmacological Treatment. Mini-Rev. Med. Chem. 2019, 20, 754–767. [Google Scholar] [CrossRef]
- Simon, D.K.; Tanner, C.M.; Brundin, P. Parkinson Disease Epidemiology, Pathology, Genetics, and Pathophysiology. Clin. Geriatr. Med. 2020, 36, 1–12. [Google Scholar] [CrossRef]
- Rascol, O.; Payoux, P.; Ory, F.; Ferreira, J.J.; Brefel-Courbon, C.; Montastruc, J. Limitations of current Parkinson’s disease therapy. Ann. Neurol. 2003, 53, S3–S12. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.L.; Mak, M.K. Effect of Repetitive Transcranial Magnetic Stimulation on Physical Function and Motor Signs in Parkinson’s Disease: A Systematic Review and Meta-Analysis. Brain Stimul. 2016, 9, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Hai-Jiao, W.; Ge, T.; Li-Na, Z.; Deng, C.; Da, X.; Shan-Shan, C.; Liu, L. The efficacy of repetitive transcranial magnetic stimulation for Parkinson disease patients with depression. Int. J. Neurosci. 2020, 130, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, J.P.; Aleman, A.; Baeken, C.; Benninger, D.H.; Brunelin, J.; Di Lazzaro, V.; Filipović, S.R.; Grefkes, C.; Hasan, A.; Hummel, F.C.; et al. Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS): An update (2014–2018). Clin. Neurophysiol. 2020, 131, 474–528. [Google Scholar] [CrossRef] [PubMed]
- Dinkelbach, L.; Brambilla, M.; Manenti, R.; Brem, A.K. Non-invasive brain stimulation in Parkinson’s disease: Exploiting crossroads of cognition and mood. Neurosci. Biobehav. Rev. 2017, 75, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Goodwill, A.M.; Lum, J.A.G.; Hendy, A.M.; Muthalib, M.; Johnson, L.; Albein-Urios, N.; Teo, W. Using non-invasive transcranial stimulation to improve motor and cognitive function in Parkinson’s disease: A systematic review and meta-analysis. Sci. Rep. 2017, 7, 14840. [Google Scholar] [CrossRef] [Green Version]
- Rektorova, I.; Anderkova, L. Noninvasive Brain Stimulation and Implications for Nonmotor Symptoms in Parkinson’s Disease. Int. Rev. Neurobiol. 2017, 134, 1091–1110. [Google Scholar] [PubMed]
- Malling, A.S.B.; Morberg, B.M.; Wermuth, L.; Gredal, O.; Bech, P.; Jensen, B.R. Effect of transcranial pulsed electromagnetic fields (T-PEMF) on functional rate of force development and movement speed in persons with Parkinson’s disease: A randomized clinical trial. PLoS ONE 2018, 13, e0204478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manenti, R.; Cotelli, M.S.; Cobelli, C.; Gobbi, E.; Brambilla, M.; Rusich, D.; Alberici, A.; Padovani, A.; Borroni, B.; Cotelli, M. Transcranial direct current stimulation combined with cognitive training for the treatment of Parkinson Disease: A randomized, placebo-controlled study. Brain Stimul. 2018, 11, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.J.; Antal, A.; Bikson, M.; Boggio, P.S.; Brunoni, A.R.; Celnik, P.; Cohen, L.G.; Fregni, F.; Herrmann, C.S.; Kappenman, E.S.; et al. A technical guide to tDCS, and related non-invasive brain stimulation tools. Clin. Neurophysiol. 2016, 127, 1031–1048. [Google Scholar] [CrossRef] [Green Version]
- Martiny, K.; Lunde, M.; Bech, P. Transcranial low voltage pulsed electromagnetic fields in patients with treatment-resistant depression. Biol. Psychiatry 2010, 68, 163–169. [Google Scholar] [CrossRef]
- Rohan, M.; Parow, A.; Stoll, A.L.; Demopulos, C.; Friedman, S.; Dager, S.; Hennen, J.; Cohen, B.M.; Renshaw, P.F. Low-field magnetic stimulation in bipolar depression using an MRI-based stimulator. Am. J. Psychiatry 2004, 161, 93–98. [Google Scholar] [CrossRef]
- Shafi, M.; Stern, A.P.; Pascual-Leone, A. Adding low-field magnetic stimulation to noninvasive electromagnetic neuromodulatory therapies. Biol. Psychiatry 2014, 76, 170–171. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Phillips, B. A pilot study of the use of EEG-based synchronized Transcranial Magnetic Stimulation (sTMS) for treatment of Major Depression. BMC Psychiatry 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Leuchter, A.F.; Cook, I.A.; Feifel, D.; Goethe, J.W.; Husain, M.; Carpenter, L.L.; Thase, M.E.; Krystal, A.D.; Philip, N.S.; Bhati, M.T.; et al. Efficacy and Safety of Low-field Synchronized Transcranial Magnetic Stimulation (sTMS) for Treatment of Major Depression. Brain Stimul. 2015, 8, 787–794. [Google Scholar] [CrossRef]
- Rohan, M.L.; Yamamoto, R.T.; Ravichandran, C.T.; Cayetano, K.R.; Morales, O.G.; Olson, D.P.; Vitaliano, G.; Paul, S.M.; Cohen, B.M. Rapid mood-elevating effects of low field magnetic stimulation in depression. Biol. Psychiatry 2014, 76, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Straaso, B.; Lauritzen, L.; Lunde, M.; Vinberg, M.; Lindberg, L.; Larsen, E.R.; Dissing, S.; Bech, P. Dose-remission of pulsating electromagnetic fields as augmentation in therapy-resistant depression: A randomized, double-blind controlled study. Acta Neuropsychiatr. 2014, 26, 272–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.; Wang, Y.; Gu, P.; Shao, R.; Zhao, L.; Liu, X.; Wang, Z.; Wang, M. The Neuroprotective Mechanism of Low-Frequency rTMS on Nigral Dopaminergic Neurons of Parkinson’s Disease Model Mice. Park. Dis. 2015, 2015, 564095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.C.; Luan, F.; Xie, C.; Geng, D.; Wang, Y.; Ma, J. Low-frequency transcranial magnetic stimulation is beneficial for enhancing synaptic plasticity in the aging brain. Neural Regen. Res. 2015, 10, 916–924. [Google Scholar] [PubMed]
- Sekar, S.; Zhang, Y.; Miranzadeh Mahabadi, H.; Parvizi, A.; Taghibiglou, C. Low-Field Magnetic Stimulation Restores Cognitive and Motor Functions in the Mouse Model of Repeated Traumatic Brain Injury: Role of Cellular Prion Protein. J. Neurotrauma 2019, 36, 3103–3114. [Google Scholar] [CrossRef] [PubMed]
- Mooshekhian, A.; Sandhini, T.; Wei, Z.; Van Bruggen, R.; Li, H.; Li, X.; Zhang, Y. Low-field magnetic stimulation improved cuprizone-induced depression-like symptoms and demylination in female mice. Exp. Ther. Med. 2022, 23, 210. [Google Scholar] [CrossRef]
- Wang, Z.; Baharani, A.; Truong, D.; Bi, X.; Wang, F.; Li, X.; Verge, V.M.K.; Zhang, Y. Low field magnetic stimulation promotes myelin repair and cognitive recovery in chronic cuprizone mouse model. Clin. Exp. Pharmacol. Physiol. 2021, 48, 1090–1102. [Google Scholar] [CrossRef]
- Jia, X.; Kohn, A. Gamma rhythms in the brain. PLoS Biol. 2011, 9, e1001045. [Google Scholar] [CrossRef]
- Uhlhaas, P.J.; Singer, W. Neural synchrony in brain disorders: Relevance for cognitive dysfunctions and pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Nowak, M.; Zich, C.; Stagg, C.J. Motor Cortical Gamma Oscillations: What Have We Learnt and Where Are We Headed? Curr. Behav. Neurosci. Rep. 2018, 5, 136–142. [Google Scholar] [CrossRef]
- Chartove, J.A.K.; McCarthy, M.M.; Pittman-Polletta, B.R.; Kopell, N.J. A biophysical model of striatal microcircuits suggests gamma and beta oscillations interleaved at delta/theta frequencies mediate periodicity in motor control. PLoS Comput. Biol. 2020, 16, e1007300. [Google Scholar] [CrossRef] [PubMed]
- Sulzer, D. Multiple hit hypotheses for dopamine neuron loss in Parkinson’s disease. Trends Neurosci. 2007, 30, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.D., Jr.; Chang, M.L.; Klaidman, L. Parkinson’s disease—Redox mechanisms. Curr. Med. Chem. 2001, 8, 809–814. [Google Scholar] [CrossRef]
- Lofredi, R.; Neumann, W.; Bock, A.; Horn, A.; Huebl, J.; Siegert, S.; Schneider, G.; Krauss, J.K.; Kühn, A.A. Dopamine-dependent scaling of subthalamic gamma bursts with movement velocity in patients with Parkinson’s disease. Elife 2018, 7, e31895. [Google Scholar] [CrossRef]
- Tabrez, S.; Jabir, N.R.; Shakil, S.; Greig, N.H.; Alam, Q.; Abuzenadah, A.M.; Damanhouri, G.A.; Kamal, M.A. A synopsis on the role of tyrosine hydroxylase in Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2012, 11, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Gebreyesus, H.H.; Gebremichael, T.G. The Potential Role of Astrocytes in Parkinson’s Disease (PD). Med. Sci. 2020, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udovin, L.; Quarracino, C.; Herrera, M.I.; Capani, F.; Otero-Losada, M.; Perez-Lloret, S. Role of Astrocytic Dysfunction in the Pathogenesis of Parkinson’s Disease Animal Models from a Molecular Signaling Perspective. Neural Plast. 2020, 2020, 1859431. [Google Scholar] [CrossRef] [Green Version]
- Haque, M.E.; Akther, M.; Jakaria, M.; Kim, I.; Azam, S.; Choi, D. Targeting the microglial NLRP3 inflammasome and its role in Parkinson’s disease. Mov. Disord. 2020, 35, 20–33. [Google Scholar] [CrossRef]
- Lazdon, E.; Stolero, N.; Frenkel, D. Microglia and Parkinson’s disease: Footprints to pathology. J. Neural Transm. 2020, 127, 149–158. [Google Scholar] [CrossRef]
- Bachiller, S.; Jiménez-Ferrer, I.; Paulus, A.; Yang, Y.; Swanberg, M.; Deierborg, T.; Boza-Serrano, A. Microglia in Neurological Diseases: A Road Map to Brain-Disease Dependent-Inflammatory Response. Front. Cell. Neurosci. 2018, 12, 488. [Google Scholar] [CrossRef] [Green Version]
- Sofroniew, M.V. Astrogliosis. Cold Spring Harb. Perspect. Biol. 2014, 7, a020420. [Google Scholar] [CrossRef] [Green Version]
- Cheyne, D.; Ferrari, P. MEG studies of motor cortex gamma oscillations: Evidence for a gamma “fingerprint” in the brain? Front. Hum. Neurosci. 2013, 7, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, C.; Duesmann, C.; Woost, T.B.; Hofen-Hohloch, J.V.; Rumpf, J.; Weise, D.; Classen, J. Dual-Site Transcranial Magnetic Stimulation for the Treatment of Parkinson’s Disease. Front. Neurol. 2019, 10, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Guo, Z.; Peng, H.; Xing, G.; Chen, H.; McClure, M.A.; He, B.; He, L.; Du, F.; Xiong, L.; et al. Repetitive transcranial magnetic stimulation therapy for motor recovery in Parkinson’s disease: A Meta-analysis. Brain Behav. 2018, 8, e01132. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Ugawa, Y. Repetitive Transcranial Magnetic Stimulation for Parkinson’s Disease: A Review. Brain Nerve 2017, 69, 219–225. [Google Scholar] [PubMed]
- Hanajima, R.; Terao, Y.; Shirota, Y.; Ohminami, S.; Tsutsumi, R.; Shimizu, T.; Tanaka, N.; Okabe, S.; Tsuji, S.; Ugawa, Y. Triad-conditioning transcranial magnetic stimulation in Parkinson’s disease. Brain Stimul. 2014, 7, 74–79. [Google Scholar] [CrossRef]
- Chervyakov, A.V.; Chernyavsky, A.Y.; Sinitsyn, D.O.; Piradov, M.A. Possible Mechanisms Underlying the Therapeutic Effects of Transcranial Magnetic Stimulation. Front. Hum. Neurosci. 2015, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- Morberg, B.M.; Malling, A.S.; Jensen, B.R.; Gredal, O.; Bech, P.; Wermuth, L. Parkinson’s disease and transcranial pulsed electromagnetic fields: A randomized clinical trial. Mov. Disord. 2017, 32, 625–626. [Google Scholar] [CrossRef]
- Huang, D.; Xu, J.; Wang, J.; Tong, J.; Bai, X.; Li, H.; Wang, Z.; Huang, Y.; Wu, Y.; Yu, M.; et al. Dynamic Changes in the Nigrostriatal Pathway in the MPTP Mouse Model of Parkinson’s Disease. Park. Dis. 2017, 2017, 9349487. [Google Scholar] [CrossRef] [Green Version]
- Sathiya, S.; Ranju, V.; Kalaivani, P.; Priya, R.J.; Sumathy, H.; Sunil, A.G.; Babu, C.S. Telmisartan attenuates MPTP induced dopaminergic degeneration and motor dysfunction through regulation of alpha-synuclein and neurotrophic factors (BDNF and GDNF) expression in C57BL/6J mice. Neuropharmacology 2013, 73, 98–110. [Google Scholar] [CrossRef]
- Zhen, J.; Qian, Y.; Weng, X.; Su, W.; Zhang, J.; Cai, L.; Dong, L.; An, H.; Su, R.; Wang, J.; et al. Gamma rhythm low field magnetic stimulation alleviates neuropathologic changes and rescues memory and cognitive impairments in a mouse model of Alzheimer’s disease. Alzheimer’s Dement. 2017, 3, 487–497. [Google Scholar] [CrossRef]
- Zhang, Y.; Schlussman, S.D.; Rabkin, J.; Butelman, E.R.; Ho, A.; Kreek, M.J. Chronic escalating cocaine exposure, abstinence/withdrawal, and chronic re-exposure: Effects on striatal dopamine and opioid systems in C57BL/6J mice. Neuropharmacology 2013, 67, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Fernagut, P.O.; Diguet, E.; Labattu, B.; Tison, F. A simple method to measure stride length as an index of nigrostriatal dysfunction in mice. J. Neurosci. Methods 2002, 113, 123–130. [Google Scholar] [CrossRef]
- Deacon, R.M. Measuring motor coordination in mice. J. Vis. Exp. 2013, 75, e2609. [Google Scholar]
- Salas, R.; Pieri, F.; Fung, B.; Dani, J.A.; De Biasi, M. Altered anxiety-related responses in mutant mice lacking the beta4 subunit of the nicotinic receptor. J. Neurosci. 2003, 23, 6255–6263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Franklin, K. The Mouse Brain in Stereotaxic Coordinates, 2nd ed.; Academic Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Bachman, J. Immunohistochemistry on freely floating fixed tissue sections. Methods Enzymol. 2013, 533, 207–215. [Google Scholar] [PubMed]
- Nichkova, M.; Wynveen, P.M.; Marc, D.T.; Huisman, H.; Kellermann, G.H. Validation of an ELISA for urinary dopamine: Applications in monitoring treatment of dopamine-related disorders. J. Neurochem. 2013, 125, 724–735. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekar, S.; Zhang, Y.; Miranzadeh Mahabadi, H.; Buettner, B.; Taghibiglou, C. Low-Field Magnetic Stimulation Alleviates MPTP-Induced Alterations in Motor Function and Dopaminergic Neurons in Male Mice. Int. J. Mol. Sci. 2023, 24, 10328. https://doi.org/10.3390/ijms241210328
Sekar S, Zhang Y, Miranzadeh Mahabadi H, Buettner B, Taghibiglou C. Low-Field Magnetic Stimulation Alleviates MPTP-Induced Alterations in Motor Function and Dopaminergic Neurons in Male Mice. International Journal of Molecular Sciences. 2023; 24(12):10328. https://doi.org/10.3390/ijms241210328
Chicago/Turabian StyleSekar, Sathiya, Yanbo Zhang, Hajar Miranzadeh Mahabadi, Benson Buettner, and Changiz Taghibiglou. 2023. "Low-Field Magnetic Stimulation Alleviates MPTP-Induced Alterations in Motor Function and Dopaminergic Neurons in Male Mice" International Journal of Molecular Sciences 24, no. 12: 10328. https://doi.org/10.3390/ijms241210328
APA StyleSekar, S., Zhang, Y., Miranzadeh Mahabadi, H., Buettner, B., & Taghibiglou, C. (2023). Low-Field Magnetic Stimulation Alleviates MPTP-Induced Alterations in Motor Function and Dopaminergic Neurons in Male Mice. International Journal of Molecular Sciences, 24(12), 10328. https://doi.org/10.3390/ijms241210328