
A Mannose Receptor from Litopenaeus vannamei Involved in Innate Immunity by Pathogen Recognition and Inflammation Regulation


Abstract
:1. Introduction
2. Results
2.1. cDNA Cloning and Sequence Analysis of LvMR
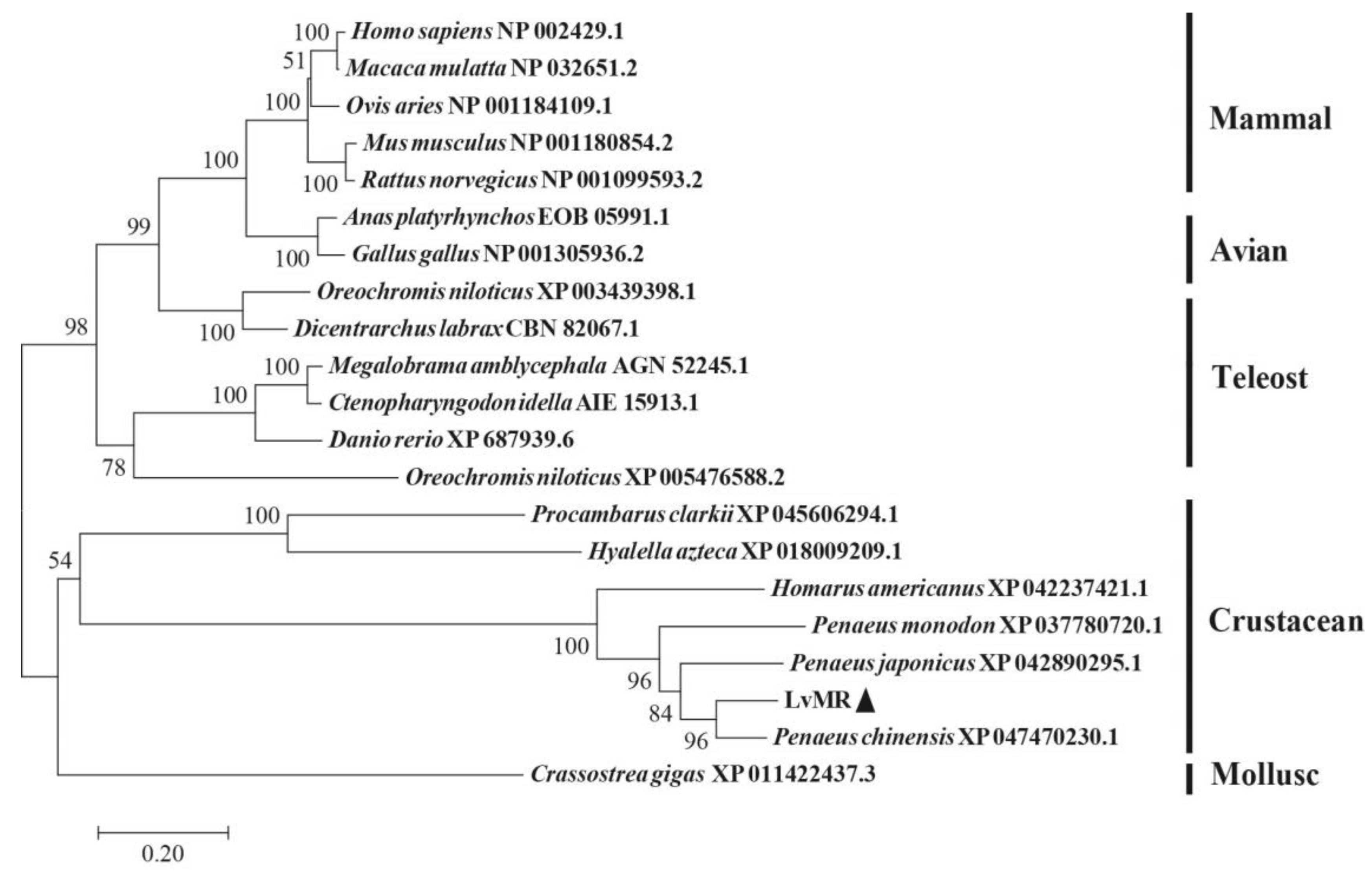
2.2. Homology and Phylogenetic Analysis of LvMR
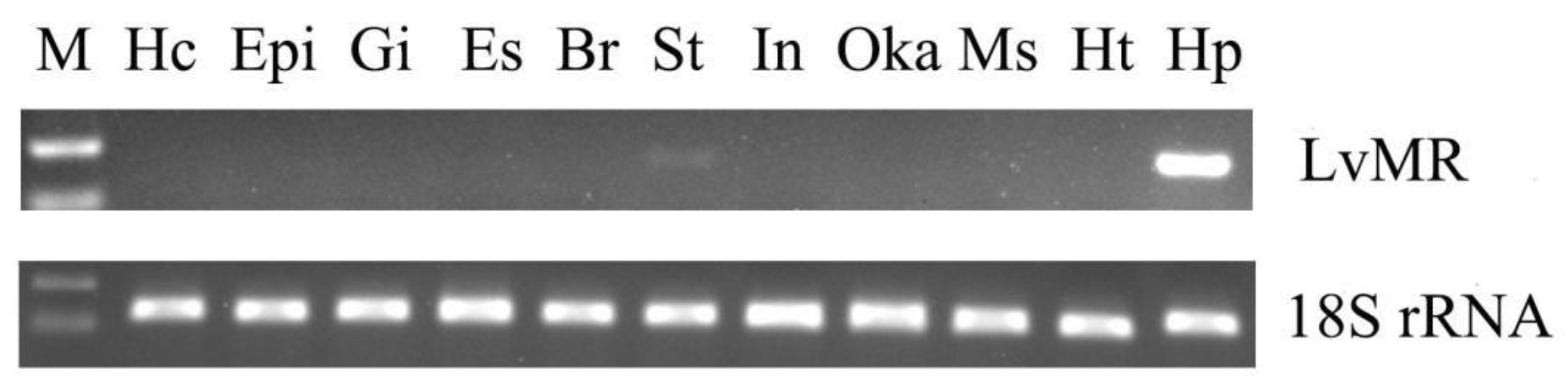
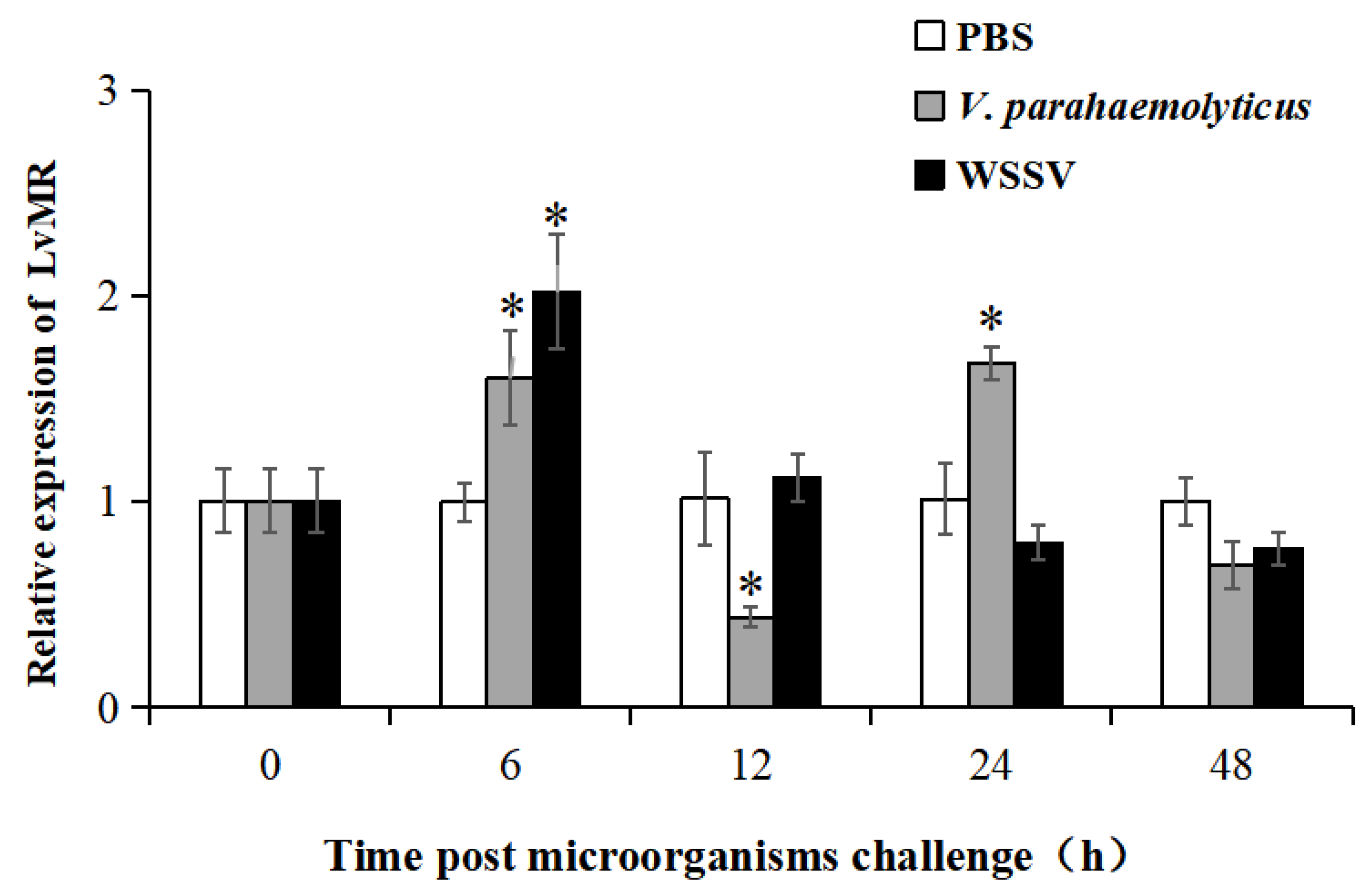
2.3. Tissue Expression and Immune Responses of LvMR
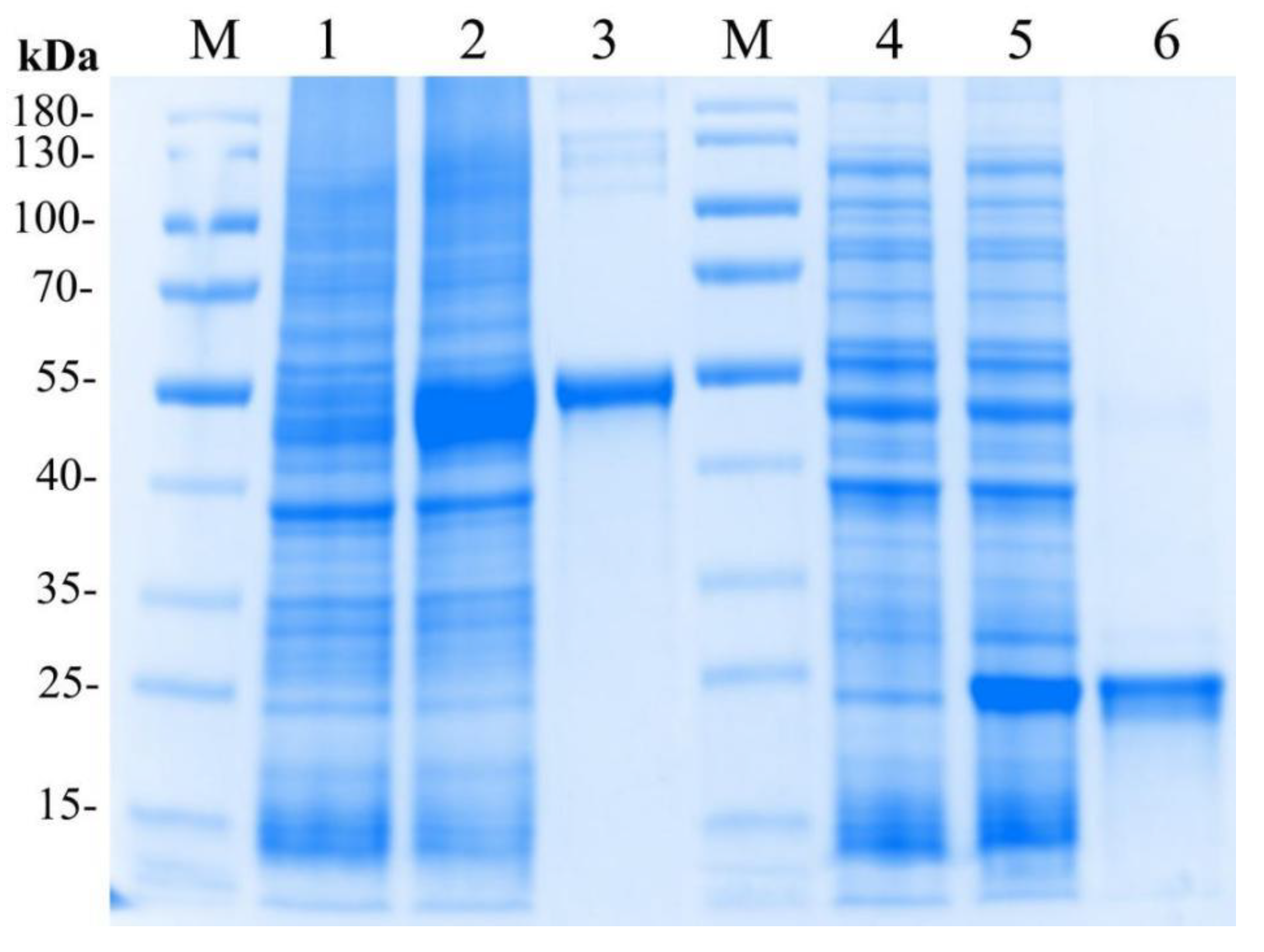
2.4. Expression and Purification of the Recombinant LvMR
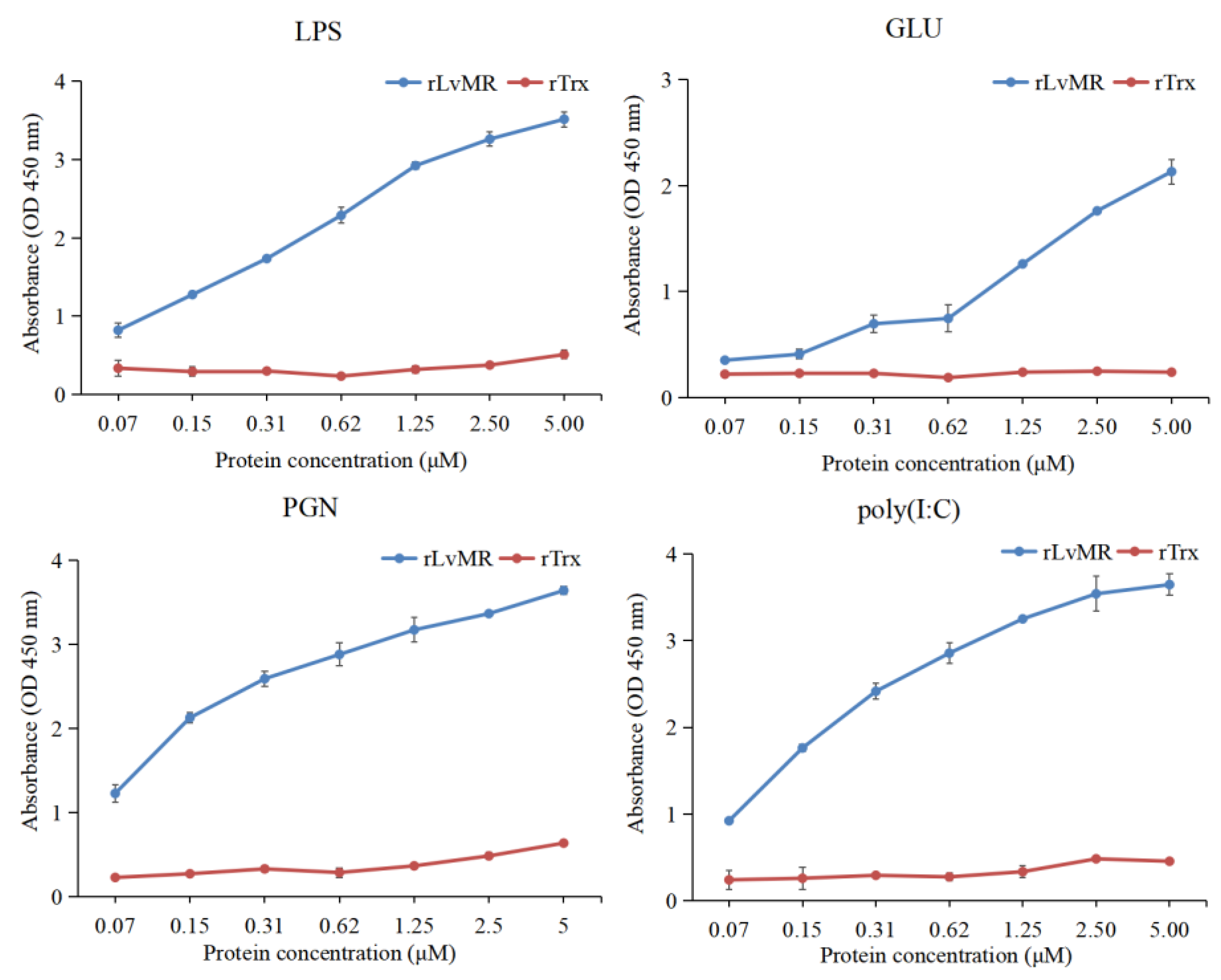
2.5. Binding Activity of rLvMR to PAMPs
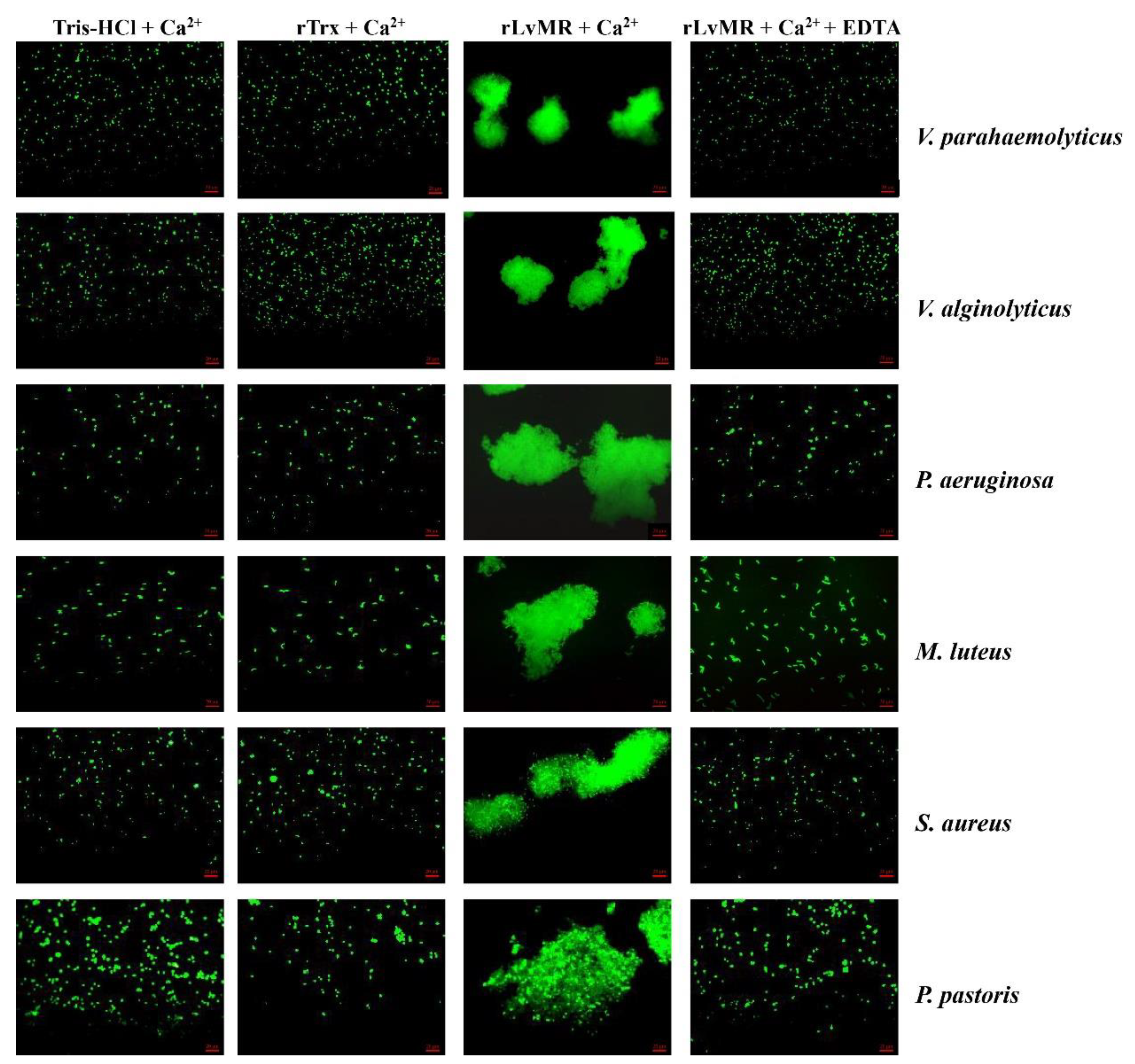
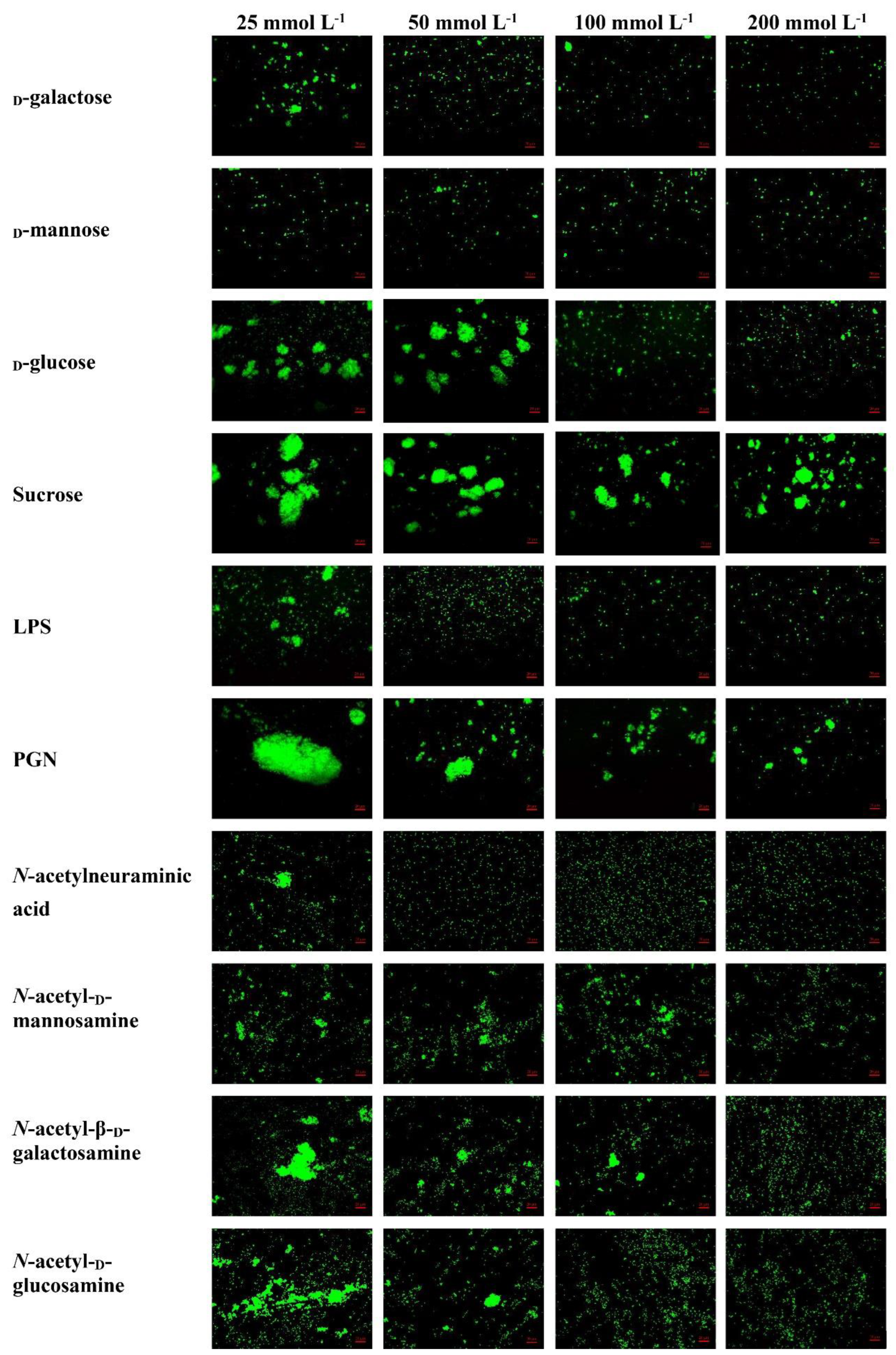
2.6. Agglutinating Activity of rLvMR to Microorganisms
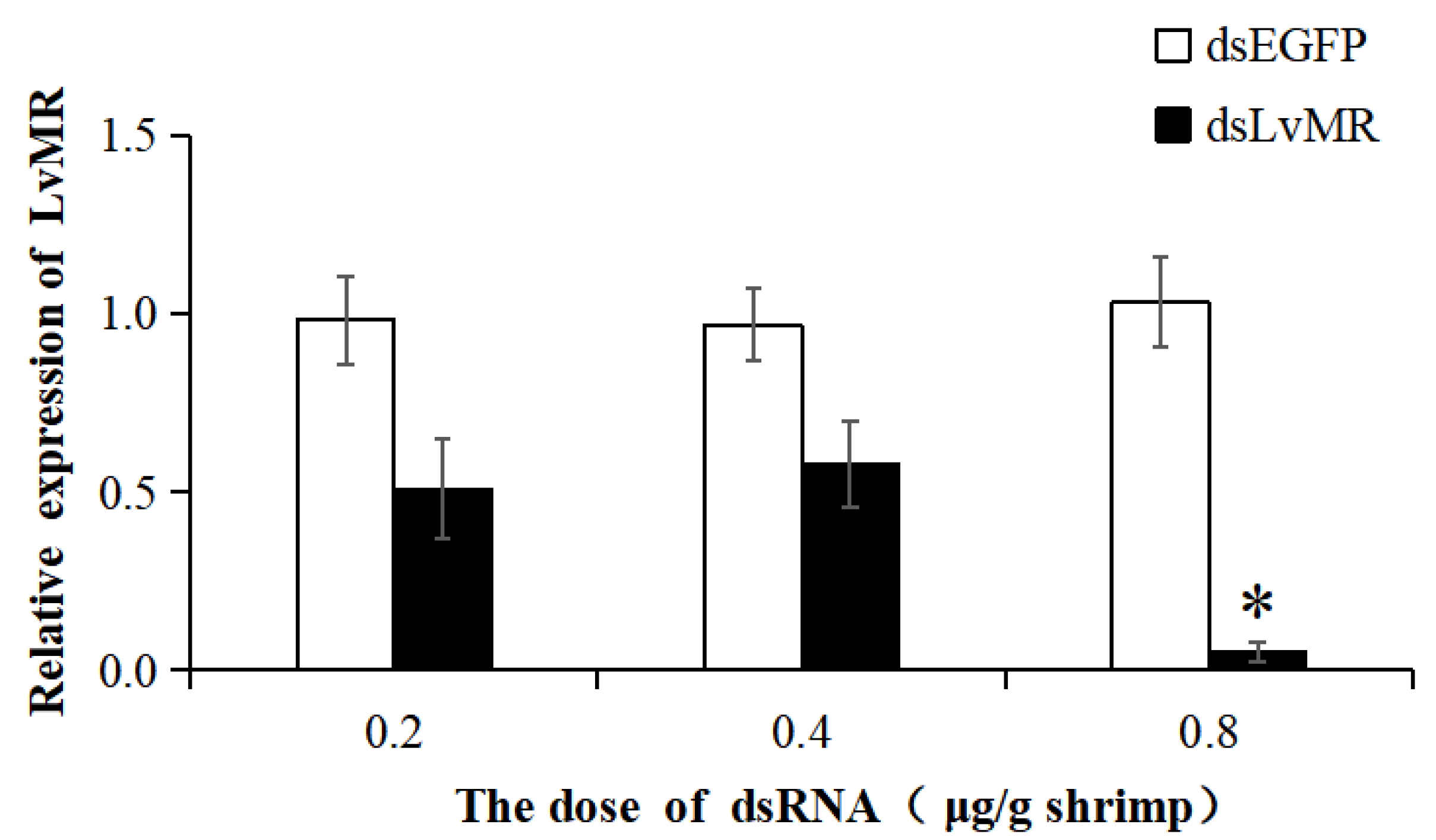
2.7. Gene Knockdown of LvMR
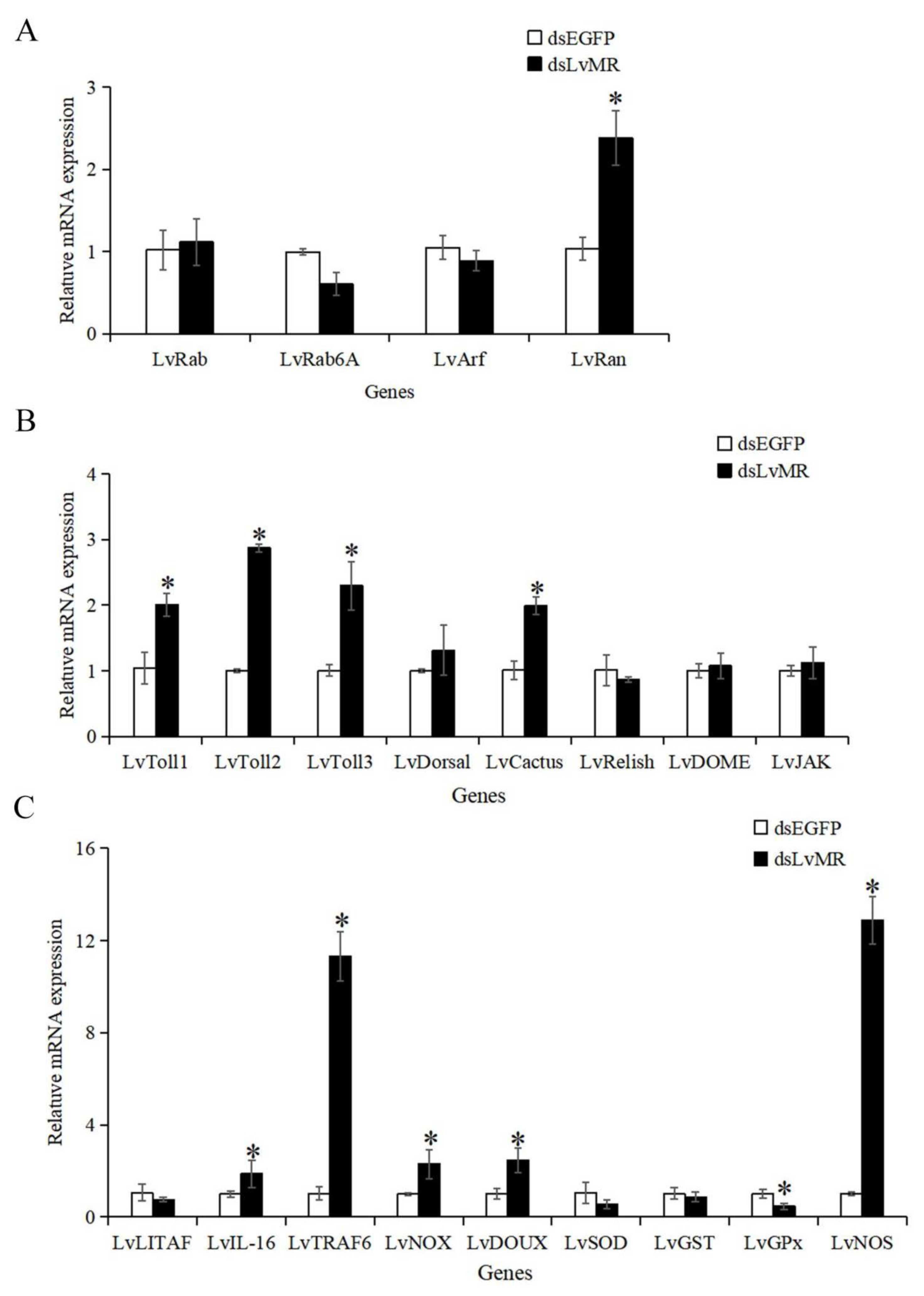
2.8. Effects of LvMR Interference on the Expression of Immune Genes
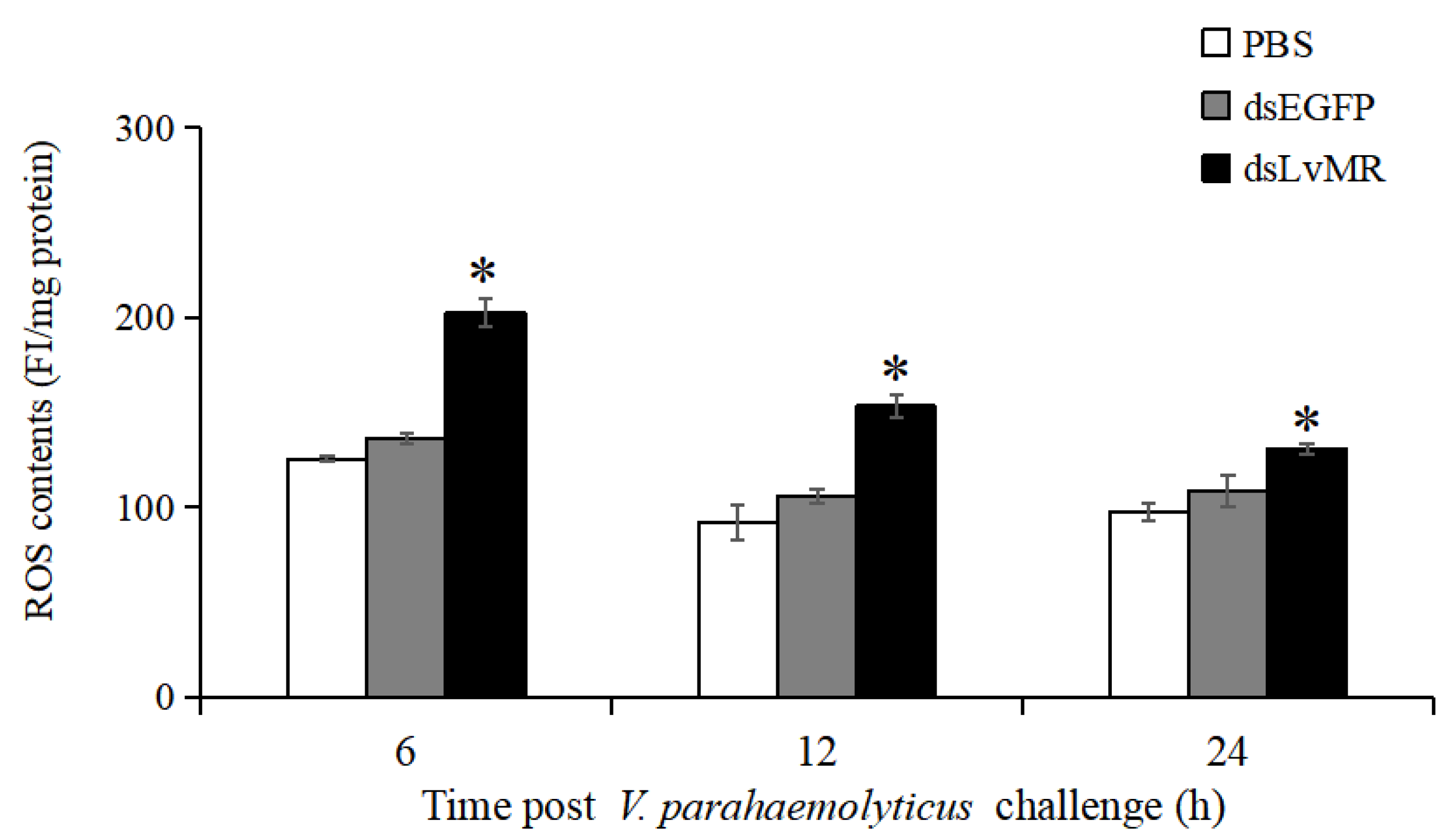
2.9. Effects of LvMR Interference on ROS Production
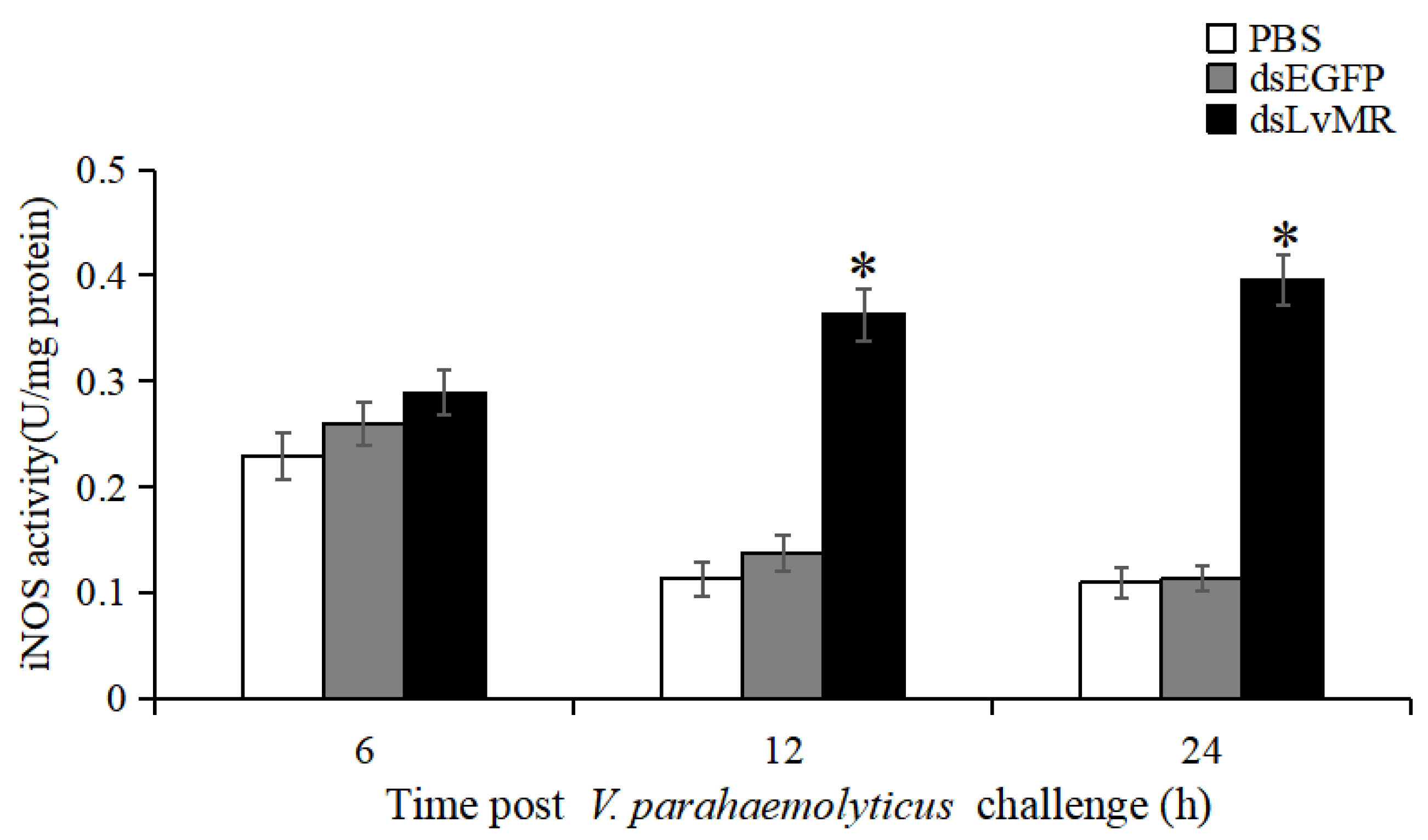
2.10. Effects of LvMR Interference on iNOS Activity
3. Discussion
4. Materials and Methods
4.1. Shrimp, Immune Stimulation and Sample Collection
4.2. RNA Extraction, cDNA Synthesis and Gene Cloning
4.3. Bioinformatics Analysis
4.4. Semi-Quantitative RT-PCR
4.5. Real-Time Quantitative PCR Analysis (qRT-PCR)
4.6. Recombinant Expression and Protein Purification
4.7. PAMP Binding Assay
4.8. Microorganism Agglutination and Agglutination Inhibition Assays
4.9. Double Strand RNA (dsRNA) Synthesis and RNA Interference Assay
4.10. Expression Changes of Immune Genes in the LvMR-Knockdown Shrimp
4.11. Assay of Reactive Oxygen Species (ROS)
4.12. Assay of Inducible NOS (iNOS) Activity
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, P.R.; Gordon, S.; Martinez-Pomares, L. The mannose receptor: Linking homeostasis and immunity through sugar recognition. Trends Immunol. 2005, 26, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Gazi, U.; Martinez-Pomares, L. Influence of the mannose receptor in host immune responses. Immunobiology 2009, 214, 554–561. [Google Scholar] [CrossRef] [PubMed]
- East, L.; Isacke, C.M. The mannose receptor family. Biochim. Biophys. Acta 2002, 1572, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Navarrete, A.M.; Bayry, J.; Delignat, S.; Wootla, B.; Andre, S.; Christophe, O.; Nascimbeni, M.; Jacquemin, M.; Martinez-Pomares, L. A role for exposed mannosylations in presentation of human therapeutic self-proteins to CD4+ T lymphocytes. Proc. Natl. Acad. Sci. USA 2007, 104, 8965–8970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wileman, T.E.; Lennartz, M.R.; Stahl, P.D. Identification of the macrophage mannose receptor as a 175-kDa membrane protein. Proc. Natl. Acad. Sci. USA 1986, 83, 2501–2505. [Google Scholar] [CrossRef] [Green Version]
- Zheng, F.; Asim, M.; Lan, J.; Zhao, L.; Wei, S. Molecular cloning and functional characterization of mannose receptor in zebra fish (Danio rerio) during infection with Aeromonas sobria. Int. J. Mol. Sci. 2015, 16, 10997–11012. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Li, J.; He, J.; Liu, W.; Jiang, L.; Ye, Y.; Wu, C. Anti-infective mannose receptor immune mechanism in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2016, 54, 257–265. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, L.; Hegazy, A.M.; Wang, H.; Li, J.; Zheng, F.; Zhou, Y.; Wang, W.; Li, J.; Liu, X. Mannose receptor mediated phagocytosis of bacteria in macrophages of blunt snout bream (Megalobrama amblycephala) in a Ca2+-dependent manner. Fish Shellfish Immunol. 2015, 43, 357–363. [Google Scholar] [CrossRef]
- Yin, X.; Bai, H.; Mu, L.; Chen, N.; Qi, W.; Huang, Y.; Xu, H.; Jian, J.; Wang, A.; Ye, J. Expression and functional characterization of the mannose receptor (MR) from Nile tilapia (Oreochromis niloticus) in response to bacterial infection. Dev. Comp. Immunol. 2022, 126, 104257. [Google Scholar] [CrossRef]
- Liu, R.; Qi, Y.; Zhai, Y.; Li, H.; An, L.; Yang, G.; Shan, S. Identification and functional analysis of mannose receptor in Asian swamp eel (Monopterus albus) in response to bacterial infection. Fish Shellfish Immunol. 2022, 127, 463–473. [Google Scholar] [CrossRef]
- Man, X.; Pan, X.T.; Zhang, H.W.; Wang, Y.; Li, X.C.; Zhang, X.W. A mannose receptor is involved in the anti-Vibrio defense of red swamp crayfish. Fish Shellfish Immunol. 2018, 82, 258–266. [Google Scholar] [CrossRef]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 2016, 452, 69–87. [Google Scholar] [CrossRef]
- Melander, M.; Jürgensen, H.J.; Madsen, D.; Engelholm, L.; Behrendt, N. The collagen receptor uPARAP/Endo180 in tissue degradation and cancer (Review). Int. J. Oncol. 2015, 47, 1177–1188. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Pomares, L.; Wienke, D.; Stillion, R.; McKenzie, E.; Arnold, J.; Harris, J.; McGreal, E.; Sim, R.; Isacke, C.; Gordon, S. Carbohydrate-independent recognition of collagens by the macrophage mannose receptor. Eur. J. Immunol. 2006, 36, 1074–1082. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Kong, P.; Wang, L.; Zhang, H.; Song, X.; Zhou, Z.; Yang, J.; Qiu, L.; Wang, L.; Song, L. A novel C-type lectin from bay scallop Argopecten irradians (AiCTL-7) agglutinating fungi with mannose specificity. Fish Shellfish Immunol. 2011, 30, 836–844. [Google Scholar] [CrossRef]
- Song, K.K.; Li, D.F.; Zhang, M.C.; Yang, H.J.; Xu, X. Cloning and characterization of three novel WSSV recognizing lectins from shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2010, 28, 596–603. [Google Scholar] [CrossRef]
- Rattanaporn, O.; Utarabhand, P. Molecular cloning of a C-type lectin with two CRD domains from the banana shrimp Fenneropenaeus merguiensis: Early gene up-regulation after Vibrio harveyi infection. J. Invertebr. Pathol. 2011, 106, 196–204. [Google Scholar] [CrossRef]
- Su, Y.; Liu, Y.; Gao, F.; Cui, Z. A novel C-type lectin with a YPD motif from Portunus trituberculatus (PtCLec1) mediating pathogen recognition and opsonization. Dev. Comp. Immunol. 2020, 106, 103609. [Google Scholar] [CrossRef]
- Huang, Y.; Huang, X.; Wang, Z.; Tan, J.M.; Hui, K.M.; Wang, W.; Ren, Q. Function of two novel single-CRD containing C-type lectins in innate immunity from Eriocheir sinensis. Fish Shellfish Immunol. 2014, 37, 313–321. [Google Scholar] [CrossRef]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Stavenhagen, K.; Mehta, A.Y.; Laan, L.; Gao, C.; Heimburg-Molinaro, J.; van Die, I.; Cummings, R.D. N-glycosylation of mannose receptor (CD206) regulates glycan binding by C-type lectin domains. J. Biol. Chem. 2022, 298, 102591. [Google Scholar] [CrossRef] [PubMed]
- Paveley, R.A.; Aynsley, S.A.; Turner, J.D.; Bourke, C.D.; Jenkins, S.J.; Cook, P.C.; Martinez-Pomares, L.; Mountford, A.P. The Mannose Receptor (CD206) is an important pattern recognition receptor (PRR) in the detection of the infective stage of the helminth Schistosoma mansoni and modulates IFNγ production. Int. J. Parasit. 2011, 41, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Zelensky, A.N.; Gready, J.E. Comparative analysis of structural properties of the C-type-lectin-like domain (CTLD). Proteins 2003, 52, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Stahl, P.D.; Ezekowitz, R.A. The mannose receptor is a pattern recognition receptor involved in host defense. Curr. Opin. Immunol. 1998, 10, 50. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, Z.; Tang, M.; Pan, G.; Zhao, L.; Qin, Z.; Lin, L. Mannose receptor of Epinephelus coioides exerts antiviral activity against red-spotted grouper nervous necrosis virus and regulates apoptosis and inflammation. Aquaculture 2022, 546, 737264. [Google Scholar] [CrossRef]
- Zhang, J.; Tachado, S.D.; Patel, N.; Zhu, J.; Koziel, H. Negative regulatory role of mannose receptors on human alveolar macrophage proinflammatory cytokine release in vitro. J. Leukoc. Biol. 2005, 78, 665–674. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, H.; Song, L.S.; Li, C.H.; Zhao, J.M.; Wang, H.; Qiu, L.M.; Ni, D.J.; Zhang, Y. A novel C1q-domain-containing protein from Zhikong scallop Chlamys farreri with lipopolysaccharide binding activity. Fish Shellfish Immunol. 2008, 25, 281–289. [Google Scholar] [CrossRef]
- Wang, M.; Wang, L.; Huang, M.M.; Song, L.S. A galectin from Eriocheir sinensis functions as pattern recognition receptor enhancing microbe agglutination and haemocytes encapsulation. Fish Shellfish Immunol. 2016, 55, 10–20. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Sequence (5′–3′) |
|---|---|
| LvMR_F | CGGAACTACATATAAATAGGCGTCG |
| LvMR_R | ATTCCCCTTTCTATTCAAGCACG |
| LvMR_ReF | GCTGATATCGGATCCGAATTCTGCAACCCGCCGTTCACCATG |
| LvMR_ReR | CTCGAGTGCGGCCGCAAGCTTGAGGAACCGCAGCTTGCAGAT |
| LvMR_qF | CTGCGTCAGCATCTCTTTG |
| LvMR_qR | GGTTTCCAGTCGGTCATTG |
| LvMR_dsF1 | TAATACGACTCACTATAGGGTACCTTGGATTCAACGCTCC |
| LvMR_dsR1 | TAATACGACTCACTATAGGGCACCCAGTACGAGTTCGCTT |
| LvMR_dsF2 | GGGTCCAAATGCCTCTAC |
| LvMR_dsR2 | ATCGCAGGAAGGGAAGGA |
| M13_47 | CGCCAGGGTTTTCCCAGTCACGAC |
| RV_M | GAGCGGATAACAATTTCACACAGG |
| T7 promoter | TAATACGACTCACTATAGGG |
| T7 terminator | GCTAGTTATTGCTCAGCGGT |
| 18S_F | TATACGCTAGTGGAGCTGGAA |
| 18S_R | GGGGAGGTAGTGACGAAAAAT |
| LvRab6A_qF | CGCAGCCTTATTCCCTCATAT |
| LvRab6A_qR | CGCACATCATCTATCCATT |
| LvRab_qF | GACAGTGGTGTTGGAAAG |
| LvRab_qR | GCCGTCTAGTTCAATTGTTCG |
| LvRan_qF | CCATACAAATAGAGGACCCAT |
| LvRan_qR | CATTCTTGTACGTGACTCTAG |
| LvArf_qF | GTCTTGATGCTGCTGGTAA |
| LvArf_qR | CAAAGATGAGACCCTGAGTA |
| LvLITAF_qF | GCAGTCAACGCACATGATCT |
| LvITAF_qR | TTGTATTTGCCCAGGAAAGC |
| LvIL-16_qF | AGCAAGAGCCTCGTGTCAGAC |
| LvIL-16_qR | CCTCCAGAGAAAAGCCCAGT |
| LvTRAF6-qF | ACATCACCAATCCCAGAG |
| LvTRAF6-qR | GTCAGCACCGCCTTTATC |
| LvSOD_qF | ATTGCCGCTACGAAGAAG |
| LvSOD_qR | AGATGGTGTGGTTCAAGTG |
| LvGPx_qF | GCACCAGGAGAACACTAC |
| LvGPx_qR | TTCCAGGCAATGTCAGAG |
| LvGST_qF | AGAAAAACTACCCTGTCGG |
| LvGST_qR | CCTTGCTCTGCGTTATCTT |
| LvDUOX_qF | GACTTGGCAGCAAACCTA |
| LvDUOX_qR | TGCGGGAAAGGTCGTAGAT |
| LvNOX_qF | CCAACGATGTGCCTGATAGTG |
| LvNOX_qR | ATGTCGGTCTTCTGAAGGGCT |
| LvNOS_qF | GAGCAAGTTATTCGGCAAGGC |
| LvNOS_qR | TCTCTCCCAGTTTCTTGGCGT |
| LvToll1_qF | CTATTGTGGTGCTTTCGT |
| LvToll1_qR | TGGAGATGTACAGTCGTAAC |
| LvToll2_qF | CATGCCTGCAGGACTGTTTA |
| LvToll2_qR | GGCCTGAGGGTAAGGTCTTC |
| LvToll3_qF | GTGAATCTGACCCGAGTTGA |
| LvToll3_qR | TGCTGCCTTCGGTGTTCTA |
| LvCactus_qF | GCCTGTCTTACGCCCCT |
| LvCactus_qR | CCGTCCGACCACTCTTG |
| LvRelish_qF | CATGCAAGACTTCGCAA |
| LvRelish_qR | CTGGTAATGTAACAGGACG |
| LvDorsal_qF | TGGGGAAGGAAGGATGC |
| LvDorsal_qR | CGTAACTTGAGGGCATCTTC |
| LvJAK_qF | CCTTAATTCGAGCGCAATGGG |
| LvJAK_qR | CTAGCGACAGAGGGTTTAGCG |
| LvDOME_qF | CTCAGGCTATGTTTCTCAGGATTCA |
| LvDOME_qR | CACGGCAGTTCCTTTATGGTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, N.; Liu, Y.; Hao, Q.; Sun, M.; Li, F. A Mannose Receptor from Litopenaeus vannamei Involved in Innate Immunity by Pathogen Recognition and Inflammation Regulation. Int. J. Mol. Sci. 2023, 24, 10665. https://doi.org/10.3390/ijms241310665
Guo N, Liu Y, Hao Q, Sun M, Li F. A Mannose Receptor from Litopenaeus vannamei Involved in Innate Immunity by Pathogen Recognition and Inflammation Regulation. International Journal of Molecular Sciences. 2023; 24(13):10665. https://doi.org/10.3390/ijms241310665
Chicago/Turabian StyleGuo, Na, Yuan Liu, Qiang Hao, Mingzhe Sun, and Fuhua Li. 2023. "A Mannose Receptor from Litopenaeus vannamei Involved in Innate Immunity by Pathogen Recognition and Inflammation Regulation" International Journal of Molecular Sciences 24, no. 13: 10665. https://doi.org/10.3390/ijms241310665
APA StyleGuo, N., Liu, Y., Hao, Q., Sun, M., & Li, F. (2023). A Mannose Receptor from Litopenaeus vannamei Involved in Innate Immunity by Pathogen Recognition and Inflammation Regulation. International Journal of Molecular Sciences, 24(13), 10665. https://doi.org/10.3390/ijms241310665