

The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment


Abstract
:1. Introduction
2. Protective Mechanisms of EGCG in Breast Cancer
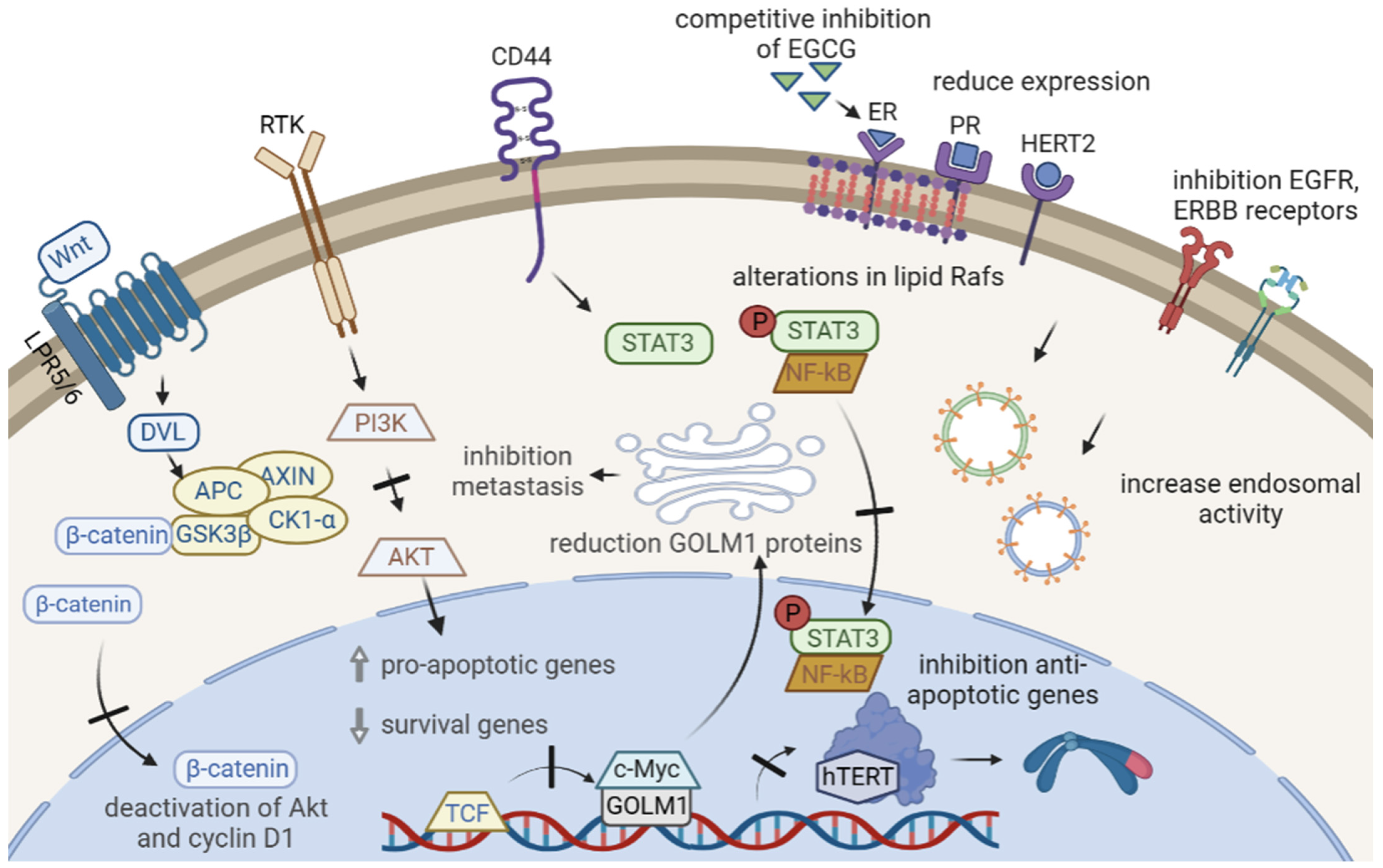
2.1. Suppressive Effects of EGCG on Proliferative Signals
2.2. EGCG Inhibits Evasion of Apoptosis in Cancer
2.3. EGCG Controls the Replicative Potential of Cancer
2.4. EGCG Inhibits Tissue Invasion and Metastasis
2.5. Effects of EGCG on the Immune System
2.6. Epigenetic Regulation of Cancer
2.7. EGCG on Cancer Metabolism
3. Synergistic Effect of EGCG in the Treatment of Breast Cancer
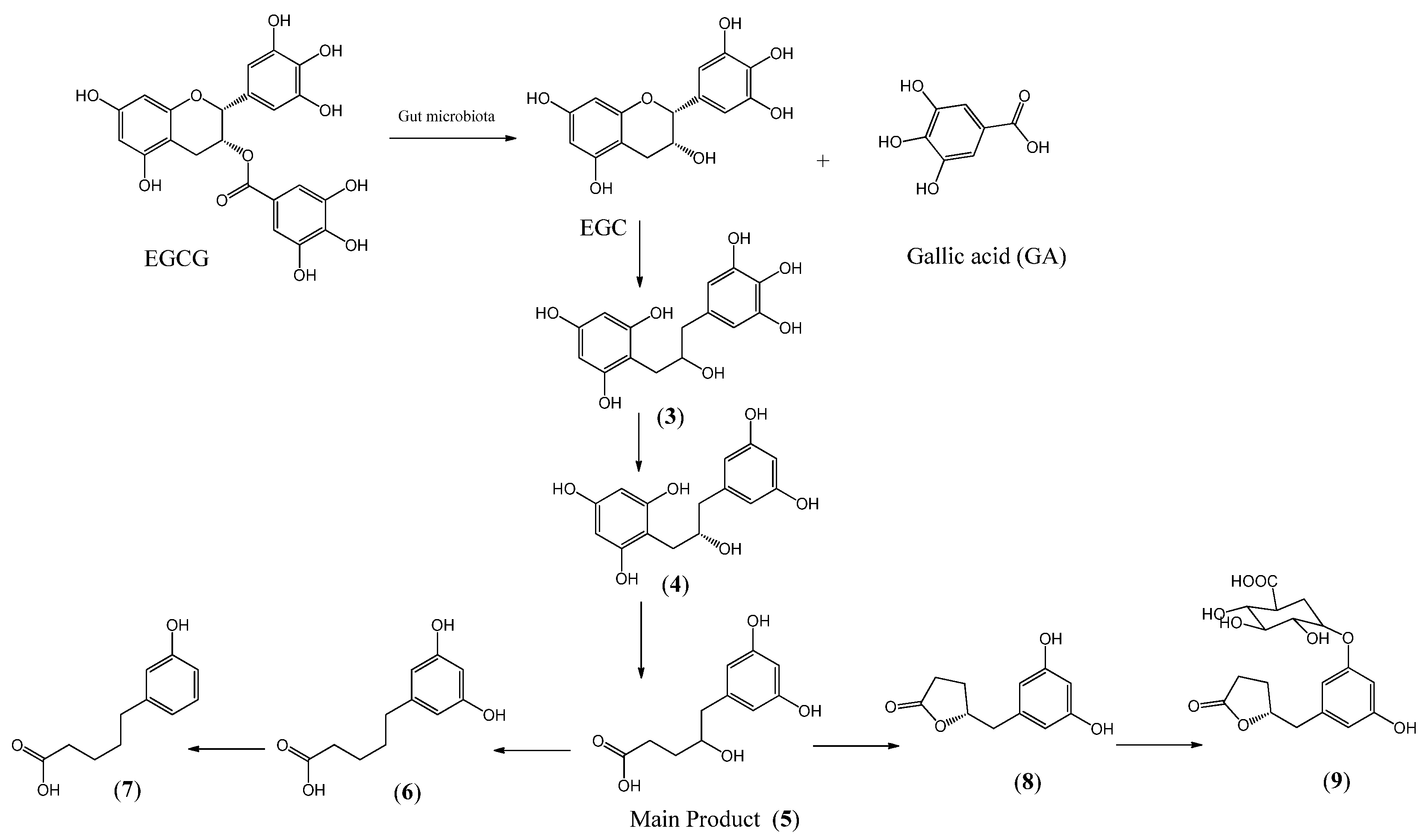
4. Absorption and Metabolism of EGCG
5. Strategies for EGCG Delivery
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- DeSantis, C.E.; Lin, C.C.; Mariotto, A.B.; Siegel, R.L.; Stein, K.D.; Kramer, J.L. Cancer treatment and survivorship statistics, 2014. CA Cancer J. Clin. 2014, 64, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; Van De Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Botstein, D. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Xue, X.; Hu, C.; Xu, H.; Kou, D.; Li, R.; Li, M. Comparison of clinicopathological features and prognosis in triple-negative and non-triple negative breast cancer. J. Cancer 2016, 7, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-negative breast cancer: Challenges and opportunities of a heterogeneous disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Mukai, H.; Uemura, Y.; Akabane, H.; Watanabe, T.; Park, Y.; Takahashi, M.; Sagara, Y.; Nishimura, R.; Takashima, T.; Fujisawa, T.; et al. Anthracycline-containing regimens or taxane versus S-1 as first-line chemotherapy for metastatic breast cancer. Br. J. Cancer 2021, 125, 1217–1225. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. NEJM 2010, 363, 1938–1948. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chan, T.H.; Dou, Q.P. EGCG, green tea polyphenols and their synthetic analogs and prodrugs for human cancer prevention and treatment. Adv. Clin. Chem. 2011, 53, 155–177. [Google Scholar]
- Nakachi, K.; Matsuyama, S.; Miyake, S.; Suganuma, M.; Imai, K. Preventive effects of drinking green tea on cancer and cardiovascular disease: Epidemiological evidence for multiple targeting prevention. Biofactors 2000, 13, 49–54. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2000, 21, 1744. [Google Scholar] [CrossRef] [Green Version]
- Avramovich-Tirosh, Y.; Reznichenko, L.; Amit, T.; Zheng, H.; Fridkin, M.; Weinreb, O.; Youdim, M.B. Neurorescue activity, APP regulation and amyloid-β peptide reduction by novel multi-functional brain permeable iron-chelating-antioxidants, M-30 and green tea polyphenol, EGCG. Curr. Alzheimer Res. 2007, 4, 403–411. [Google Scholar] [CrossRef]
- Kaya, Z.; Yayla, M.; Cinar, I.; Atila, N.E.; Ozmen, S.; Bayraktutan, Z.; Bilici, D. Epigallocatechin-3-gallate (EGCG) exert therapeutic effect on acute inflammatory otitis media in rats. Int. J. Pediatr. Otorhinolaryngol. 2019, 124, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Holman, C.D.A.J.; Huang, J.P.; Xie, X. Green tea and the prevention of breast cancer: A case–control study in Southeast China. Carcinogenesis 2007, 28, 1074–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R.; Thabrew, I.; De Silva, E.D. Protective effects of six selected dietary compounds against leptin-induced proliferation of oestrogen receptor positive (MCF-7) breast cancer cells. Medicines 2017, 4, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodin, M.G.; Fertuck, K.C.; Zacharewski, T.R.; Rosengren, R.J. Estrogen receptor-mediated actions of polyphenolic catechins in vivo and in vitro. Toxicol. Sci. 2002, 69, 354–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Afaq, F.; Saleem, M.; Ahmad, N.; Mukhtar, H. Targeting multiple signaling pathways by green tea polyphenol (−)-epigallocatechin-3-gallate. Cancer Res. 2006, 66, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.C.; Liou, Y.M. The anti-cancer effects of (−)-Epigalocathine-3-gallate on the signaling pathways associated with membrane receptors in MCF-7 cells. J. Cell. Physiol. 2011, 226, 2721–2730. [Google Scholar] [CrossRef]
- Schröder, L.; Marahrens, P.; Koch, J.G.; Heidegger, H.; Vilsmeier, T.; Phan-Brehm, T.; Richter, D.U. Effects of green tea, matcha tea and their components epigallocatechin gallate and quercetin on MCF-7 and MDA-MB-231 breast carcinoma cells. Oncol. Rep. 2019, 41, 387–396. [Google Scholar]
- Pan, X.; Zhao, B.; Song, Z.; Han, S.; Wang, M. Estrogen receptor-α36 is involved in epigallocatechin-3-gallate induced growth inhibition of ER-negative breast cancer stem/progenitor cells. J. Pharmacol. Sci. 2016, 130, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Filippi, A.; Picot, T.; Aanei, C.M.; Nagy, P.; Szöllősi, J.; Campos, L.; Mocanu, M.M. Epigallocatechin-3-O-gallate alleviates the malignant phenotype in A-431 epidermoid and SK-BR-3 breast cancer cell lines. Int. J. Food Sci. Nutr. 2018, 69, 584–597. [Google Scholar] [CrossRef]
- Roskoski, R. The ErbB/HER family of protein-tyrosine kinases and cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef]
- Wang, Z. ErbB Receptors and Cancer. Methods Mol. Biol. 2017, 1652, 3–35. [Google Scholar] [PubMed]
- Yang, C.S.; Wang, H. Mechanistic issues concerning cancer prevention by tea catechins. Mol. Nutr. Food Res. 2011, 55, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Mocanu, M.M.; Ganea, C.; Georgescu, L.; Varadi, T.; Shrestha, D.; Baran, I.; Szöllősi, J. Epigallocatechin 3-O-gallate induces 67 kDa laminin receptor-mediated cell death accompanied by downregulation of ErbB proteins and altered lipid raft clustering in mammary and epidermoid carcinoma cells. J. Nat. Prod. 2014, 77, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Haque, I.; Ray, P.; Ghosh, A.; Dutta, D.; Quadir, M.; Banerjee, S.K. CCN5 activation by free or encapsulated EGCG is required to render triple-negative breast cancer cell viability and tumor progression. Pharmacol. Res. Perspect. 2021, 9, e00753. [Google Scholar] [CrossRef] [PubMed]
- Hallman, K.; Aleck, K.; Quigley, M.; Dwyer, B.; Lloyd, V.; Szmyd, M.; Dinda, S. The regulation of steroid receptors by epigallocatechin-3-gallate in breast cancer cells. Breast Cancer Targets Ther. 2017, 9, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Moradzadeh, M.; Hosseini, A.; Erfanian, S.; Rezaei, H. Epigallocatechin-3-gallate promotes apoptosis in human breast cancer T47D cells through down-regulation of PI3K/AKT and Telomerase. Pharmacol. Rep. 2017, 69, 924–928. [Google Scholar] [CrossRef]
- Wang, K.; Wang, R.L.; Liu, J.J.; Zhou, J.; Li, X.; Hu, W.W.; Jiang, W.J.; Hao, N.B. The prognostic significance of hTERT overexpression in cancers: A systematic review and meta-analysis. Medicine 2018, 97, e11794. [Google Scholar] [CrossRef]
- Chung, S.S.; Vadgama, J.V. Curcumin and epigallocatechin gallate inhibit the cancer stem cell phenotype via down-regulation of STAT3–NFκB signaling. Anticancer Res. 2015, 35, 39–46. [Google Scholar] [PubMed]
- Fukada, T.; Ohtani, T.; Yoshida, Y.; Shirogane, T.; Nishida, K.; Nakajima, K.; Hirano, T. STAT3 orchestrates contradictory signals in cytokine-induced G1 to S cell-cycle transition. EMBO J. 1998, 17, 6670–6677. [Google Scholar] [CrossRef] [Green Version]
- Boras-Granic, K.; Wysolmerski, J.J. Wnt signaling in breast organogenesis. Organogenesis 2008, 4, 116–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amado, N.G.; Fonseca, B.F.; Cerqueira, D.M.; Neto, V.M.; Abreu, J.G. Flavonoids: Potential Wnt/beta-catenin signaling modulators in cancer. Life Sci. 2011, 89, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, O.Y.; Noh, E.M.; Jang, H.Y.; Lee, Y.R.; Lee, B.K.; Jung, S.H.; Youn, H.J. Epigallocatechin gallate inhibits the growth of MDA-MB-231 breast cancer cells via inactivation of the β-catenin signaling pathway. Oncol. Lett. 2017, 14, 441–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Guo, L.; Zhang, L.; Hu, Y.; Shang, D.; Ji, D. Bioinformatics analysis of microarray profiling identifies the mechanism of focal adhesion kinase signalling pathway in proliferation and apoptosis of breast cancer cells modulated by green tea polyphenol epigallocatechin 3-gallate. J. Pharm. Pharmacol. 2018, 70, 1606–1618. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yi, J.; Song, Y.; Zhao, M.; Fan, L.; Zhao, L. Suppression of GOLM1 by EGCG through HGF/HGFR/AKT/GSK-3β/β-catenin/c-Myc signaling pathway inhibits cell migration of MDA-MB-231. FCT 2021, 157, 112574. [Google Scholar] [CrossRef] [PubMed]
- Nowakowska, A.; Tarasiuk, J. Comparative effects of selected plant polyphenols, gallic acid and epigallocatechin gallate, on matrix metalloproteinases activity in multidrug resistant MCF7/DOX breast cancer cells. Acta Biochim. Pol. 2016, 63, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Roomi, M.W.; Monterrey, J.C.; Kalinovsky, T.; Rath, M.; Niedzwiecki, A. Patterns of MMP-2 and MMP-9 expression in human cancer cell lines. Oncol. Rep. 2009, 21, 1323–1333. [Google Scholar]
- Xu, P.; Yan, F.; Zhao, Y.; Chen, X.; Sun, S.; Wang, Y.; Ying, L. Green Tea Polyphenol EGCG Attenuates MDSCs-mediated Immunosuppression through Canonical and Non-Canonical Pathways in a 4T1 Murine Breast Cancer Model. Nutrients 2020, 12, 1042. [Google Scholar] [CrossRef] [Green Version]
- Parker, K.H.; Beury, D.W.; Ostrand-Rosenberg, S. Myeloid-derived suppressor cells: Critical cells driving immune suppression in the tumor microenvironment. Adv. Cancer Res. 2015, 128, 95–139. [Google Scholar]
- Chen, J.; Ye, Y.; Liu, P.; Yu, W.; Wei, F.; Li, H.; Yu, J. Suppression of T cells by myeloid-derived suppressor cells in cancer. Hum. Immunol. 2017, 78, 113–119. [Google Scholar] [CrossRef]
- Solito, S.; Falisi, E.; Diaz-Montero, C.M.; Doni, A.; Pinton, L.; Rosato, A.; Francescato, S.; Basso, G.; Zanovello, P.; Onicescu, G. A human promyelocytic-like population is responsible for the immune suppression mediated by myeloid-derived suppressor cells. Blood 2011, 118, 2254–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.; Shi, W.; Guo, H.; Long, W.; Wang, Y.; Qi, J.; Xu, Y. The inhibitory effect of (−)-epigallocatechin-3-gallate on breast cancer progression via reducing SCUBE2 methylation and DNMT activity. Molecules 2019, 24, 2899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldívar-González, F.I.; Gómez-García, A.; Chavez-Ponce de Leon, D.E.; Sánchez-Cruz, N.; Ruiz-Rios, J.; Pilón-Jiménez, B.A.; Medina-Franco, J.L. Inhibitors of DNA methyltransferases from natural sources: A computational perspective. Front. Pharmacol. 2018, 9, 1144. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.; Nur, S.M.; Abdulaal, W.H. A study on DNA methylation modifying natural compounds identified EGCG for induction of IFI16 gene expression related to the innate immune response in cancer cells. Oncol. Lett. 2022, 24, 1–10. [Google Scholar] [CrossRef]
- Assumpção, J.H.M.; Takeda, A.A.S.; Sforcin, J.M.; Rainho, C.A. Effects of propolis and phenolic acids on triple-negative breast cancer cell lines: Potential involvement of epigenetic mechanisms. Molecules 2022, 25, 1289. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Meeran, S.M.; Tollefsbol, T.O. Combinatorial bioactive botanicals re-sensitize tamoxifen treatment in ER-negative breast cancer via epigenetic reactivation of ERα expression. Sci. Rep. 2017, 7, 9345. [Google Scholar] [CrossRef] [Green Version]
- Zan, L.; Chen, Q.; Zhang, L.; Li, X. Epigallocatechin gallate (EGCG) suppresses growth and tumorigenicity in breast cancer cells by downregulation of miR-25. Bioengineered 2019, 10, 374–382. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Tsuneoka, M. Gallic acid derivatives propyl gallate and epigallocatechin gallate reduce rRNA transcription via induction of KDM2A activation. Biomolecules 2022, 12, 30. [Google Scholar] [CrossRef]
- Wei, R.; Mao, L.; Xu, P.; Zheng, X.; Hackman, R.M.; Mackenzie, G.G.; Wang, Y. Suppressing glucose metabolism with epigallocatechin-3-gallate (EGCG) reduces breast cancer cell growth in preclinical models. Food Funct. 2018, 9, 5682–5696. [Google Scholar] [CrossRef] [PubMed]
- Pouysségur, J.; Dayan, F.; Mazure, N.M. Hypoxia signalling in cancer and approaches to enforce tumour regression. Nature 2006, 441, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Cheng, T.C.; Yen, Y.; Fang, C.L.; Liao, Y.C.; Kuo, C.C.; Ho, Y.S. Tea polyphenol epigallocatechin-3-gallate inhibits cell proliferation in a patient-derived triple-negative breast cancer xenograft mouse model via inhibition of proline-dehydrogenase-induced effects. JFDA 2021, 29, 113. [Google Scholar] [CrossRef] [PubMed]
- Kuban-Jankowska, A.; Kostrzewa, T.; Musial, C.; Barone, G.; Lo-Bosco, G.; Lo-Celso, F.; Gorska-Ponikowska, M. Green tea catechins induce inhibition of PTP1B phosphatase in breast cancer cells with potent anti-cancer properties: In Vitro assay, molecular docking, and dynamics studies. Antioxidants 2020, 9, 1208. [Google Scholar] [CrossRef] [PubMed]
- Lessard, L.; Stuible, M.; Tremblay, M.L. The two faces of PTP1B in cancer. Biochim. Biophys. Acta BBA-Proteins Proteom. 2010, 1804, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, A.; Sánchez-Jiménez, F.; Maymó, J.; Dueñas, J.L.; Varone, C.; Sánchez-Margalet, V. Role of leptin in female reproduction. Clin. Chem. Lab. Med. 2015, 53, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Lorincz, A.M.; Sukumar, S. Molecular links between obesity and breast cancer. Endocr. Relat. Cancer 2006, 13, 279–292. [Google Scholar] [CrossRef]
- Gonzalez Suarez, N.; Rodriguez Torres, S.; Ouanouki, A.; El Cheikh-Hussein, L.; Annabi, B. EGCG inhibits adipose-derived mesenchymal stem cells differentiation into adipocytes and prevents a STAT3-mediated paracrine oncogenic control of triple-negative breast cancer cell invasive phenotype. Molecules 2021, 26, 1506. [Google Scholar] [CrossRef]
- Gonzalez Suarez, N.; Fernandez-Marrero, Y.; Torabidastgerdooei, S.; Annabi, B. EGCG prevents the onset of an inflammatory and cancer-associated adipocyte-like phenotype in adipose-derived mesenchymal stem/stromal cells in response to the triple-negative breast cancer secretome. Nutrients 2022, 14, 1099. [Google Scholar] [CrossRef]
- Chisholm, K.; Bray, B.; Rosengren, R. Tamoxifen and epigallocatechin gallate are synergistically cytotoxic to MDA-MB-231 human breast cancer cells. Anti-Cancer Drugs 2004, 15, 889–897. [Google Scholar] [CrossRef]
- Lubecka, K.; Kaufman-Szymczyk, A.; Cebula-Obrzut, B.; Smolewski, P.; Szemraj, J.; Fabianowska-Majewska, K. Novel clofarabine-based combinations with polyphenols epigenetically reactivate retinoic acid receptor beta, inhibit cell growth, and induce apoptosis of breast cancer cells. Int. J. Mol. Sci. 2018, 19, 3970. [Google Scholar] [CrossRef] [Green Version]
- Bimonte, S.; Cascella, M.; Barbieri, A.; Arra, C.; Cuomo, A. Shining a light on the effects of the combination of (–)-epigallocatechin-3-gallate and tapentadol on the growth of human triple-negative breast cancer cells. In Vivo 2019, 33, 1463–1468. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Tang, J.; Du, Y.; Ding, J.; Liu, J.Y. The green tea polyphenol EGCG potentiates the antiproliferative activity of sunitinib in human cancer cells. Tumor Biol. 2017, 37, 8555–8566. [Google Scholar] [CrossRef]
- Li, Y.; Yuan, Y.Y.; Meeran, S.M.; Tollefsbol, T.O. Synergistic epigenetic reactivation of estrogen receptor-α (ERα) by combined green tea polyphenol and histone deacetylase inhibitor in ERα-negative breast cancer cells. Mol. Cancer 2010, 9, 274. [Google Scholar] [CrossRef] [Green Version]
- Braicu, C.; Pileczki, V.; Pop, L.; Petric, R.C.; Chira, S.; Pointiere, E.; Berindan-Neagoe, I. Dual targeted therapy with p53 siRNA and Epigallocatechingallate in a triple negative breast cancer cell model. PLoS ONE 2015, 10, e0120936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farabegoli, F.; Govoni, M.; Spisni, E.; Papi, A. EGFR inhibition by (−)-epigallocatechin-3-gallate and IIF treatments reduces breast cancer cell invasion. Biosci. Rep. 2017, 37, BSR20170168. [Google Scholar] [CrossRef] [Green Version]
- Steed, K.L.; Jordan, H.R.; Tollefsbol, T.O. SAHA and EGCG promote apoptosis in triple-negative breast cancer cells, possibly through the modulation of cIAP2. Anticancer Res. 2020, 40, 9–26. [Google Scholar] [CrossRef]
- Tyagi, T.; Treas, J.N.; Mahalingaiah, P.K.S.; Singh, K.P. Potentiation of growth inhibition and epigenetic modulation by combination of green tea polyphenol and 5-aza-2′-deoxycytidine in human breast cancer cells. Breast Cancer Res. Treat. 2015, 149, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Deb, G.; Thakur, V.S.; Limaye, A.M.; Gupta, S. Epigenetic induction of tissue inhibitor of matrix metalloproteinase-3 by green tea polyphenols in breast cancer cells. Mol. Carcinog. 2015, 54, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Foygel, K.; Sekar, T.V.; Paulmurugan, R. Monitoring the antioxidant mediated chemosensitization and ARE-signaling in triple negative breast cancer therapy. PLoS ONE 2015, 10, e0141913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.C.; Choi, J.S. Effects of epigallocatechin gallate on the oral bioavailability and pharmacokinetics of tamoxifen and its main metabolite, 4-hydroxytamoxifen, in rats. Anti-Cancer Drugs 2009, 20, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Nakayama, K.; Nakamura, M.; Sookwong, P.; Tsuduki, T.; Niino, H.; Miyazawa, T. Effects of co-administration of tea epigallocatechin-3-gallate (EGCG) and caffeine on absorption and metabolism of EGCG in humans. Biosci. Biotechnol. Biochem. 2009, 73, 2014–2017. [Google Scholar] [CrossRef] [Green Version]
- Kohri, T.; Matsumoto, N.; Yamakawa, M.; Suzuki, M.; Nanjo, F.; Hara, Y.; Oku, N. Metabolic fate of (−)-[4-3H] epigallocatechin gallate in rats after oral administration. J. Agric. Food Chem. 2001, 49, 4102–4112. [Google Scholar] [CrossRef]
- Takagaki, A.; Nanjo, F. Metabolism of (−)-epigallocatechin gallate by rat intestinal flora. J. Agric. Food Chem. 2010, 58, 1313–1321. [Google Scholar] [CrossRef]
- Sang, S.; Lee, M.J.; Hou, Z.; Ho, C.T.; Yang, C.S. Stability of tea polyphenol (−)-epigallocatechin-3-gallate and formation of dimers and epimers under common experimental conditions. J. Agric. Food Chem. 2005, 53, 9478–9484. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, X.; Pang, Q.; Zhu, Q.; Liang, Q.; Zi, C.; Sheng, J. Proliferation of MDA-MB-231 can be suppressed by dimeric-epigallocatechin gallate through competitive inhibition of amphiregulin-epidermal growth factor receptor signaling. Anticancer Drugs 2021, 32, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Landis-Piwowar, K.R.; Huo, C.; Chen, D.I.; Milacic, V.; Shi, G.; Chan, T.H.; Dou, Q.P. A novel prodrug of the green tea polyphenol (−)-epigallocatechin-3-gallate as a potential anticancer agent. Cancer Res. 2007, 67, 4303–4310. [Google Scholar] [CrossRef] [Green Version]
- Gan, N.; Wakayama, C.; Inubushi, S.; Kunihisa, T.; Mizumoto, S.; Baba, M.; Ooya, T. Size dependency of selective cellular uptake of epigallocatechin gallate-modified gold nanoparticles for effective radiosensitization. ACS Appl. Bio Mater. 2021, 5, 355–365. [Google Scholar] [CrossRef]
- Ding, J.; Yao, J.; Xue, J.; Li, R.; Bao, B.; Jiang, L.; He, Z. Tumor-homing cell-penetrating peptide linked to colloidal mesoporous silica encapsulated (-)-epigallocatechin-3-gallate as drug delivery system for breast cancer therapy in vivo. ACS Appl. Mater. Interfaces 2015, 7, 18145–18155. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Yao, Z.; Ding, J.; Min, Q.; Jiang, L.; Zhu, J.J. Cascaded aptamers-governed multistage drug-delivery system based on biodegradable envelope-type nanovehicle for targeted therapy of HER2-overexpressing breast cancer. ACS Appl. Mater. Interfaces 2018, 10, 34050–34059. [Google Scholar] [CrossRef]
- Ding, J.; Liang, T.; Min, Q.; Jiang, L.; Zhu, J.J. “Stealth and fully-laden” drug carriers: Self-assembled nanogels encapsulated with epigallocatechin gallate and siRNA for drug-resistant breast cancer therapy. ACS Appl. Mater. Interfaces 2018, 10, 9938–9948. [Google Scholar] [CrossRef] [PubMed]
- Hajipour, H.; Hamishehkar, H.; Nazari Soltan Ahmad, S.; Barghi, S.; Maroufi, N.F.; Taheri, R.A. Improved anticancer effects of epigallocatechin gallate using RGD-containing nanostructured lipid carriers. Artif. Cells Nanomed. Biotechnol. 2018, 46, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, B.P.; Puvvada, N.; Rajput, S.; Sarkar, S.; Mahto, M.K.; Yallapu, M.M.; Mandal, M. Targeting of EGFR, VEGFR2, and Akt by engineered dual drug encapsulated mesoporous silica–gold nanoclusters sensitizes tamoxifen-resistant breast cancer. Mol. Pharm. 2018, 15, 2698–2713. [Google Scholar] [CrossRef]
- Zeng, L.; Yan, J.; Luo, L.; Ma, M.; Zhu, H. Preparation and characterization of (−)-Epigallocatechin-3-gallate (EGCG)-loaded nanoparticles and their inhibitory effects on Human breast cancer MCF-7 cells. Sci. Rep. 2017, 7, 45521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, B.; Ying, H.; Yu, C.; Fan, Z.; Zhang, W.; Shi, J.; Du, Q. (−)-Epigallocatechin gallate (EGCG)-nanoethosomes as a transdermal delivery system for docetaxel to treat implanted human melanoma cell tumors in mice. Int. J. Pharm. 2016, 512, 22–31. [Google Scholar] [CrossRef]
- Chen, Q.; Li, Q.; Liang, Y.; Zu, M.; Chen, N.; Canup, B.S.; Xiao, B. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer via ROS generation and microbiota modulation. Acta Pharm. Sin. B 2022, 12, 907–923. [Google Scholar] [CrossRef]
- El-Kayal, M.; Nasr, M.; Elkheshen, S.; Mortada, N. Colloidal (−)-epigallocatechin-3-gallate vesicular systems for prevention and treatment of skin cancer: A comprehensive experimental study with preclinical investigation. Eur. J. Pharm. Sci. 2019, 137, 104972. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; He, J.; Jiang, S.; Gao, Y.; Zhang, L.K.; Yin, L.; Guan, Y.Q. Multi-ligand modified PC@ DOX-PA/EGCG micelles effectively inhibit the growth of ER+, PR+ or HER 2+ breast cancer. J. Mater. Chem. 2022, 10, 418–429. [Google Scholar]
- Kazi, J.; Sen, R.; Ganguly, S.; Jha, T.; Ganguly, S.; Debnath, M.C. Folate decorated epigallocatechin-3-gallate (EGCG) loaded PLGA nanoparticles; in-vitro and in-vivo targeting efficacy against MDA-MB-231 tumor xenograft. Int. J. Pharm. 2020, 585, 119449. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Pooja, D.; Kulhari, H.; Gudem, S.; Ravuri, H.G.; Bhargava, S.; Ramakrishna, S. Bombesin conjugated solid lipid nanoparticles for improved delivery of epigallocatechin gallate for breast cancer treatment. Chem. Phys. Lipids 2019, 224, 104770. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Study | Treatment | Models | Outcomes | References |
|---|---|---|---|---|
| in vitro studies | ||||
| Cell viability by cell proliferation assay | 4-hydroxytamoxifen (1 μM) + 5–25 μM EGCG. 7 days | MCF-7, 47D, MDA-MB-231 and HS578T | Decrease cell viability. Synergistic activity of EGCG in MDA-MB-231 cells at 25 μM. | [58] |
| Cell growth, apoptosis, and epigenetic regulation of transcriptional activity of DNA | Clofarabine/ EGCG 10 μM. 4 days | MCF7 and MDA-MB-231 | Decrease cell growth with low toxicity. Increase in apoptosis and reactivation of DNA methylation-silenced tumor suppressor genes such as RARB. | [59] |
| Effects of Tapentadol on viability and migration | Tapentadol 1–80 μg/mL + EGCG 1–160 μM | MDA-MB-231 | Reduction of proliferation by impairing cell-cycle progression. Increase in apoptosis. | [60] |
| Inhibition insulin receptor substrate (IRS)/MAPK | 24 h of sunitinib and then 12 h with pulsed EGCG 0–50 μM | MCF-7, H460, and H1975, with PIK3CA mutations | Downregulation of insulin receptor substrate (IRS), suppressed mitogenic effects, and inhibition of IRS/MAPK/p-S6K1 signaling. | [61] |
| Reactivation of ERa by EGCG and histone deacetylase inhibitor | Trichostatin A (TSA) (100 ng/mL for 12 h) + 10 μM EGCG | MDA-MB-231 | Sensibilization of ERα-negative breast cancer cells to the activator 17β-estradiol (E2) and antagonist tamoxifen. | [62] |
| Effects of p53 gene silencing in conjunction with EGCG. | p53 siRNA 40 nmol + EGCG 24 h | Hs578T | Activation of pro-apoptotic and inhibition of anti-apoptotic genes, autophagy, and cell network formation. | [63] |
| Effects of EGCG and IIF in the EGFR inhibition | IIF 15 or 30 μM + 25 μg/mL EGCG by 24 h | MCF-7 and MDA-MB-231 | Inhibition of EGFR phosphorylation, invasion, and metastasis. | [64] |
| Synergism of SAHA and EGCG in TNBC cells | Suberoylanilide hydroxamic acid (SAHA) 25 mM + EGCG 100 mM, every 24 h/3 days | MDA-MB-157, MDA-MB-231, and HCC1806 | Increase apoptosis by decreasing cIAP2 and increasing pro-apoptotic caspase 7. Inhibition of cell migration. | [65] |
| Synergism of 5-aza 2’dC with EGCG | 5-aza 2′dC 5 µM+ EGCG 50 µM. 7 days | MCF-7, MDA-MB 231 and MCF-10A | Synergic effects in cell growth inhibition by epigenetic mechanisms. | [66] |
| Evaluation of epigenetic induction of matrix metalloproteinase-3 | green tea polyphenols 10 µg/mL + EGCG 20 µM 24 h | MCF-7 and MDA-MB-231 | Activation of TIMP-3 and reduction of zeste homolog 2 (EZH2) and class I histone deacetylases (HDACs). | [67] |
| in vivo studies | ||||
| FLuc2 fusion with N-terminal 100-aa of Nrf2 and activation of Nrf2-ARE signaling | Oral cisplatin 5 mg/kg + EGCG 100 mg/kg. 11 days | MDA-MB231 tumor xenografts | Synergistic activity in vivo by Tumor size reduction in TNBC tumor xenografts. | [68] |
| Effects of EGCG on oral bioavailability of Tamoxifen | Tamoxifen (IV, 2 mg/kg and PO, 10 mg/kg), followed by EGCG (0.5, 3 and 10 mg/kg). | Male Sprague Dawley rats | Increase of bioavailability 1.48–1.77-fold of Tamoxifen in the presence of EGCG. | [69] |
| Evaluate angiogenesis and VEGF levels | EGCG single doses 4 h after sunitinib treatment | MCF-7 and H460 xenograft tumors | Downregulation of IRS-1 levels and suppressed mitogenic effects. Marked suppression of the IRS/MAPK/p-S6K1 signaling cascade. | [61] |
| Formulation | Study | Results | References |
|---|---|---|---|
| In vitro studies | |||
| Dimeric-EGCG oxidized and polymerized. | Competitive inhibition of Amphiregulin (AREG) in MDA-MB-231 cells. | Proliferation and migration were significantly inhibited by dimeric-EGCG at 10 μM. | [74] |
| Peracetate-protected (−)-EGCG (Pro-EGCG). | Anticancerogenic effects in MDA-MB-231 tumors. | Enhanced tumor and proteasome inhibition, apoptosis induction, and accumulation. | [75] |
| Gold nanoparticles (AuNPs) with ratios EGCG/gold 1:2 to 10:1. | Study in MDA-MB-231 cells. | Particles of 39 nm in diameter enhanced irradiation-induced cell death. | [76] |
| Colloidal mesoporous silica (CMS) and breast tumor-homing cell-penetrating peptide (PEGA-pVEC peptide). | Comparison of anticancerogenic properties of EGCG into CMS and CMS@peptide. | CMS@peptide enhanced the efficacy of EGCG on breast tumors by targeted accumulation and release. | [77] |
| Specific aptamers to HER2 and ATP organized in a hierarchical manner loaded with EGCG and protamine sulfate. | SK-BR-3; MDA-MB-231. | Improved inhibitory tumor growth and minimum side effects to normal organs and tissues. | [78] |
| Biodegradable gel: EGCG + siRNA + protamine. | MDA-MB-231 and xenograft MDA-MB-231 tumor-bearing mice. | The formulation enhanced cytotoxicity to cancer cells 15-fold, with little toxicity to normal tissues. | [79] |
| Nanostructured lipid carriers Arginyl-glycyl-aspartic acid + EGCG; EGCG-loaded NLC-RGD. | Cytotoxic and apoptotic effects and uptake into MDA-MB-231 cells were evaluated. | Nanoparticles with a size of 85 nm enhanced the apoptotic activity of EGCG with higher accumulation in tumors. | [80] |
| Mesoporous silica gold cluster nanodrug loaded with dual drugs, ZD6474 and EGCG. | Adjuvant treatment to Tamoxifen in MCF-7 and T-47D cells. | The nanoformulation enhanced the toxicity of drugs against chemoresistant cancers. | [81] |
| 2 EGCG nanoparticles FA-NPS-PEG and FA-PEG-NPS. | Modulation of PI3K-Akt pathway and regulatory proteins in MCF-7 cells. | EGCG-FA-NPS-PEG, with a size of 185.0 nm and an encapsulation efficiency of 90.36%, enhanced the cytotoxic activity with IC50 of 65.9 μg/mL. | [82] |
| FA-NPS-PEG and FA-PEG-NPS nanoparticles. | CNN5 gene activation in MCF-7 (ER-α positive) and MDA-MB-231 (TNBC). | EGCG makes TNBC cells sensitive to estrogen via activating ER-α, reducing the viability and enhancing tumor formation. | [25] |
| In vivo studies | |||
| EGCG-nanoethosomes, loaded with docetaxel (DT). | Transdermal delivery using mouse skin and treatment of skin cancer growth. | Mice treated with DT-EGCG-nanoethosomes exhibited a significant tumor size reduction by 31.5% after 14 d. | [83] |
| Natural nanovehicles (exosome-like) from tea flowers (TFENs), particle sizes 131 nm. | Evaluation of tumor growth and metastasis. | Inhibition of growth and tumor metastasis. | [84] |
| Encapsulation of EGCG in ultradeformable colloidal vesicular systems or penetration enhancer-containing vesicles (PEVs). | Study of photodegradation, stability, and anticancer properties. | EGCG-loaded PEVs increase the cytotoxic activity of epidermoid carcinoma cells (A431) and reduce tumor sizes. | [85] |
| PC@DOX-PA/EGCG nanoparticles: Phosphatidylcholine, doxorubicin, and procyanidin with HER2, ER, and PR ligands on the surface. | Antitumor evaluation activity in BT-474, MCF-7, EMT-6, and MDA-MB-231. | Nanoparticles can target breast cancer cells and inhibit tumoral growth. | [86] |
| Folate peptide nanoparticles loaded with EGCG (FP-EGCG-NPs). | Antitumor activity in MDA-MB-231 and MCF-7 cells. | FP-EGCG-NPs enhanced the antitumor activity. | [87] |
| EGCG in solid lipid nanoparticles conjugated to gastrin-releasing peptide receptors (GRPR). | Tumoral studies on C57/BL6 mice. | Enhanced cytotoxicity to cancer cells, reduction in tumor volume, and greater animal survivability. | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marín, V.; Burgos, V.; Pérez, R.; Maria, D.A.; Pardi, P.; Paz, C. The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment. Int. J. Mol. Sci. 2023, 24, 10737. https://doi.org/10.3390/ijms241310737
Marín V, Burgos V, Pérez R, Maria DA, Pardi P, Paz C. The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment. International Journal of Molecular Sciences. 2023; 24(13):10737. https://doi.org/10.3390/ijms241310737
Chicago/Turabian StyleMarín, Víctor, Viviana Burgos, Rebeca Pérez, Durvanei Augusto Maria, Paulo Pardi, and Cristian Paz. 2023. "The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment" International Journal of Molecular Sciences 24, no. 13: 10737. https://doi.org/10.3390/ijms241310737
APA StyleMarín, V., Burgos, V., Pérez, R., Maria, D. A., Pardi, P., & Paz, C. (2023). The Potential Role of Epigallocatechin-3-Gallate (EGCG) in Breast Cancer Treatment. International Journal of Molecular Sciences, 24(13), 10737. https://doi.org/10.3390/ijms241310737