
Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern


Abstract
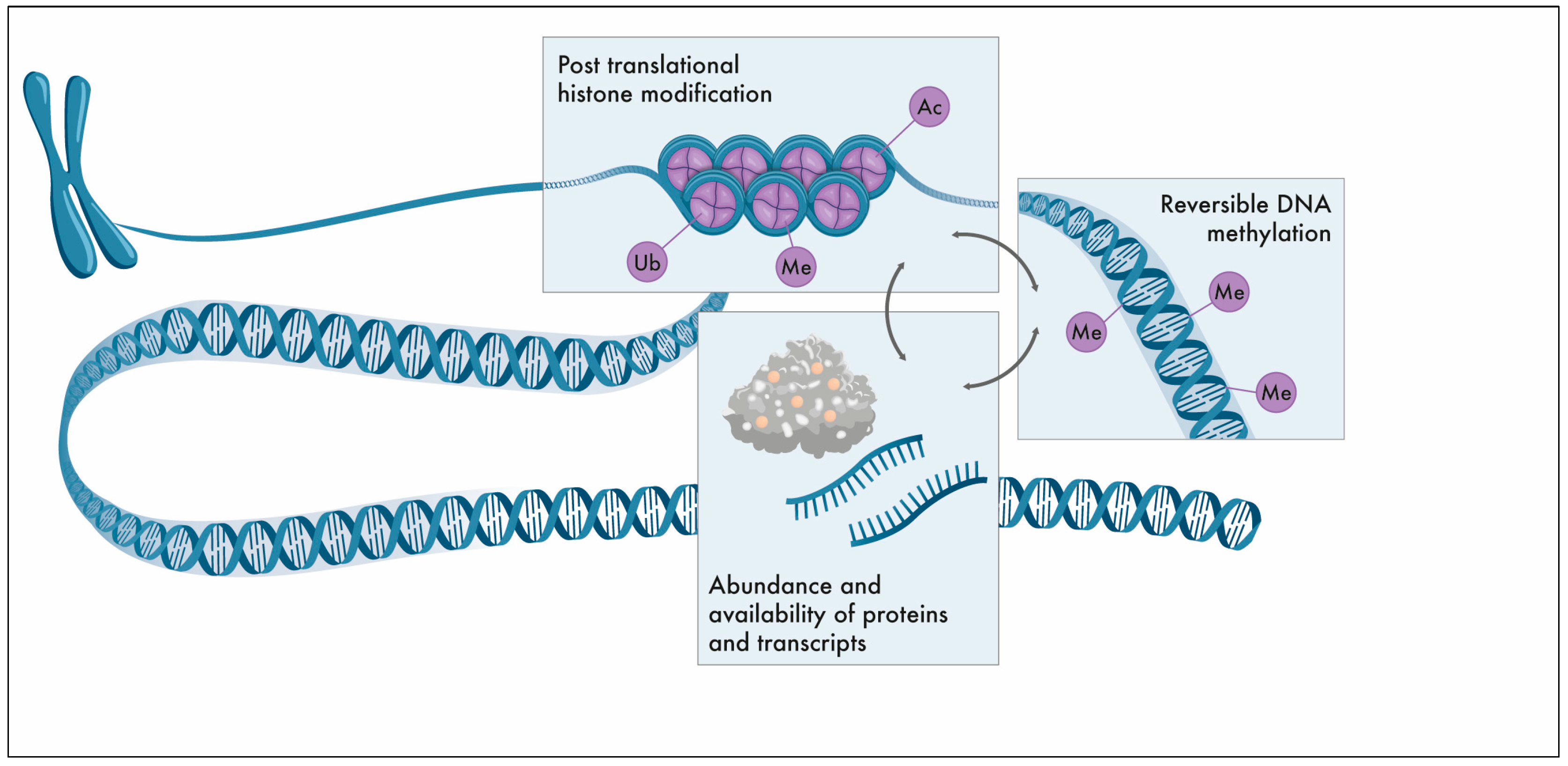
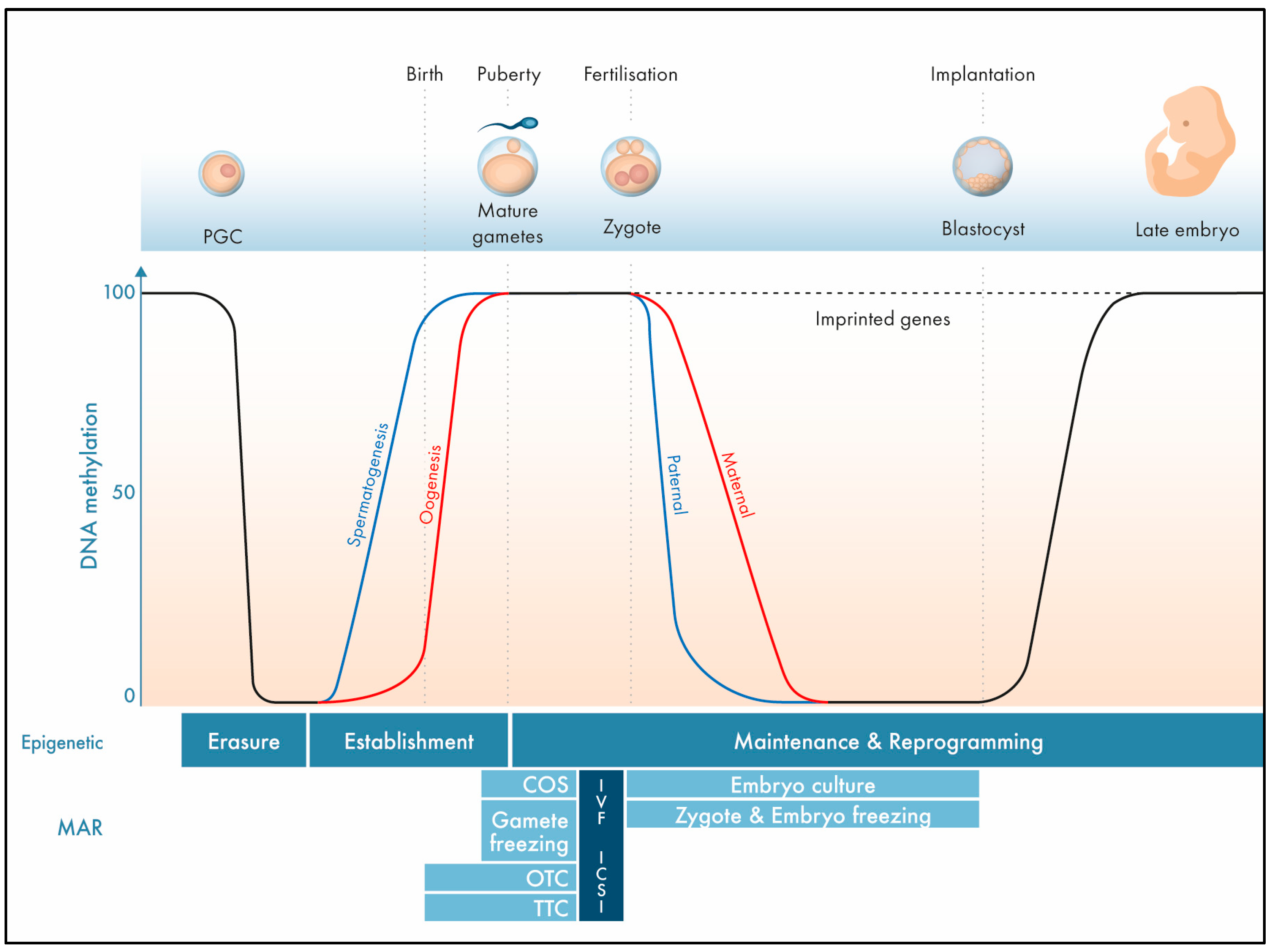
:1. The Epigenome
2. Imprinting Disorders
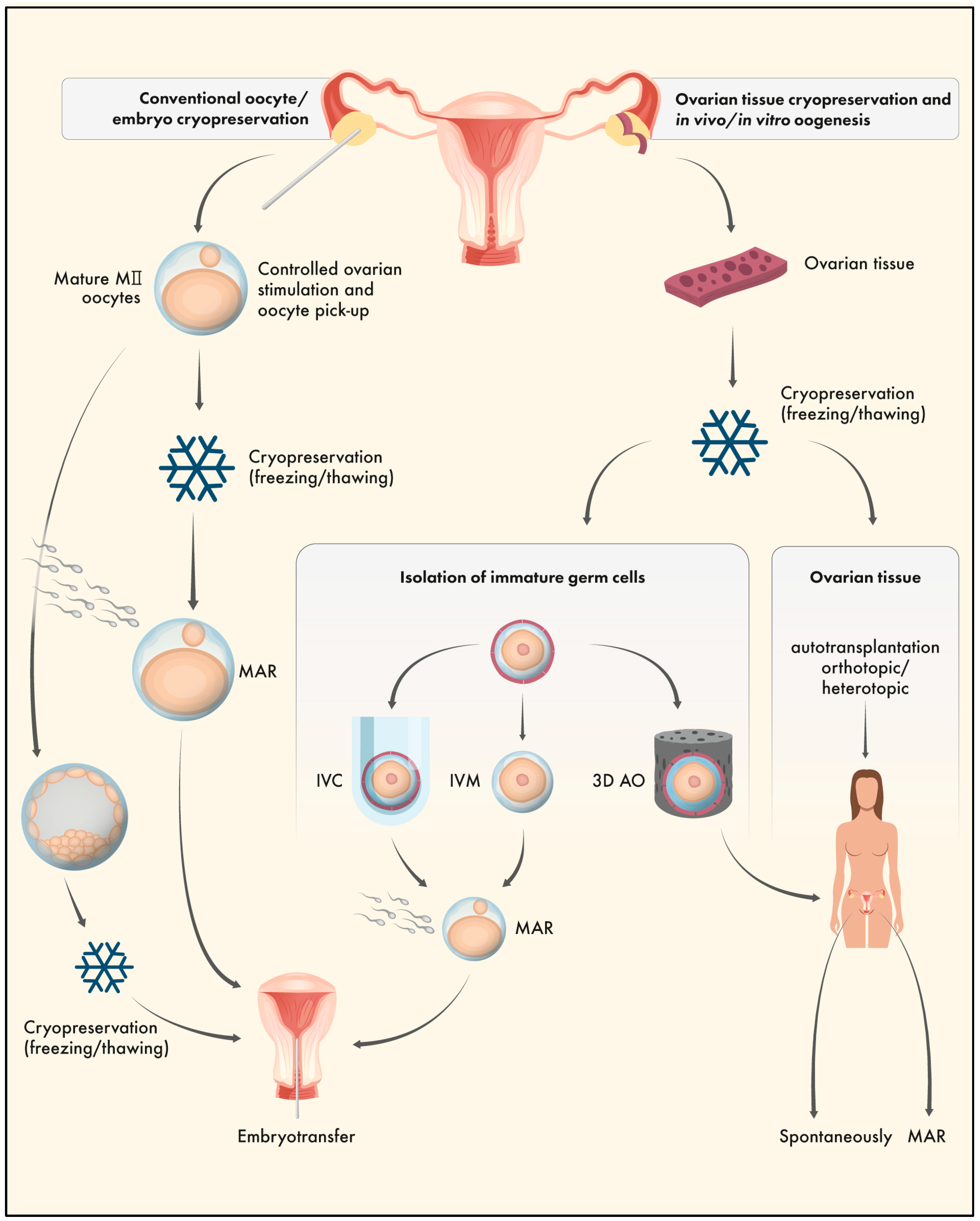
3. Ovarian Tissue Cryopreservation (OTC)
4. OTC and the Epigenome
4.1. Gene Expression and Protein Abundance
4.2. DNA Methylation Patterns
4.3. Post-Translational Histone Modifications

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Type/Species (Cryopreservation Technique) | Analysis (Technique) | Main Outcome |
|---|---|---|---|
| Shirazi et al. (2016) [51] | Ovine GV oocytes, IVM (vitrification including 20% DMSO and 20% EG) | Epigenetically-relevant mRNA abundance in GV/embryos (real-time qPCR) | Alteration of DNMT3B and HDAC1 |
| Demant et al. (2012) [53] | Murine pre-antral follicles, IVC (vitrification including 15% DMSO and 15% EG) | Proteome analysis in GV/MII (LC-MS/MS and 2D DIGE) | No differences between vitrified and non-vitrified GV/MII |
| He et al. (2018) [50] | Murine OTC (vitrification including 20% DMSO and 20% EG) | mRNA expression and protein abundance (Western blotting and real-time qPCR) | Decreased mRNA/protein levels for Dnmt1 |
| Yan et al. (2020) [52] | Murine OTC and orthotopic transplantation (slow freezing including 1.5M DMSO; vitrification including 15% DMSO and 15% EG) | Epigenetically relevant mRNA abundance in offspring (real-time qPCR) | mRNA differences in H19, Igf2r and PLAGL1 but normal Snrpn expression |
| Yodrug et al. (2021) [58] | Bovine GV oocytes, IVM (vitrification including 15% DMSO and 15% EG) | Global DNA methylation in MII and embryos (immunofluorescent assay) | Normal gDNA pattern in MII but altered in blastocysts |
| Al-Khtib et al. (2011) [57] | Human GV oocytes, IVM (vitrification including PrOH and EG; concentrations n/a; DMSO free) | Imprinted genes in MII (bisulphite mutagenesis and sequencing) | Normal pattern for H19 and KCNQ1OT1 |
| Yan et al. (2014) [56] | Murine GV oocytes, IVM (vitrification including 15% PrOH and 15% EG; DMSO free) | Global DNA methylation in MII (immunofluorescent assay) | Normal gDNA pattern |
| Trapphoff et al. (2010) [59] | Murine pre-antral follicles, IVC (vitrification including 15% DMSO and 15% EG) | Imprinted genes in GV (bisulphite pyrosequencing) | Normal establishment of H19 and Igf2r imprinting but some single CpG errors in Snrpn |
| Yan et al. (2020) [52] | Murine OTC and orthotopic transplantation (slow freezing including 1.5M DMSO; Vitrification including 15% DMSO and 15% EG) | Imprinted genes in offspring (MethylDetector PCR after bisulphite treatment) | Significant variations in H19, Igf2r, and PLAGL1 but normal Snrpn methylation |
| He et al. (2018) [50] | Murine OTC (vitrification including 20% DMSO and 20% EG) | Methylation pattern (Western blotting and real-time qPCR; indirect) | Hypermethylation of the Grb10 promoter region |
| Wang et al. (2013) [55] | Murine OTC (vitrification including 20% DMSO and 20% EG or 5.5M EG) | Methylation pattern in GV after vitrification/warming (bisulphite sequencing PCR) | Normal Snrpn methylation |
| Sauvat et al. (2008) [61] | Murine OTC and grafting (slow freezing including 1.5M DMSO) | Imprinted genes in offspring (Southern blotting) | Normal H19 and Lit1 methylation |
| Sauvat et al. (2013) [62] | OTC and grafting in ewes (slow freezing including 1.5M DMSO) | Imprinted gene in offspring (bisulphite mutagenesis and sequencing) | Normal Igf2r methylation |
| Damavandi et al. (2021) [60] | Murine pre-antral follicles, IVC (vitrification including 15% DMSO and 15% EG) | CpG methylation in granulosa cells (direct PCR sequencing after bisulphite treatment) | Altered Inhba/Inhbb methylation |
| Yodrug et al. (2021) [58] | Bovine GV oocytes, IVM (vitrification including 15% DMSO and 15% EG) | Histone modifications (immunofluorescent assay) | Normal H3K9me pattern in MII/blastocysts |
| Tian et al. (2022) [63] | Murine pre-antral follicles, IVC (vitrification including 0.75M EG and 0.75M PrOH) | Histone modifications in embryos (immunofluorescent assay) | Normal histone pattern (H3K9me3, H3K4me3, H3K27ac) |
| Lee and Comizzoli (2019) [64] | Domestic cat GV (vitrification including 20% DMSO and 20% EG) | Histone modifications after vitrification (immunofluorescent assay) | Normal H3K9me3 but altered H3K4me3 |
5. Testicular Tissue Cryopreservation (TTC)
6. TTC and the Epigenome
7. Conclusions—OTC and TTC and the Epigenome
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tucci, V.; Isles, A.R.; Kelsey, G.; Ferguson-Smith, A.C. Genomic Imprinting and Physiological Processes in Mammals. Cell 2019, 176, 952–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Gao, Z.; Qin, D.; Li, L. A Maternal Functional Module in the Mammalian Oocyte-To-Embryo Transition. Trends Mol. Med. 2017, 23, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Swales, A.K.E.; Spears, N. Genomic Imprinting and Reproduction. Reproduction 2005, 130, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anvar, Z.; Chakchouk, I.; Demond, H.; Sharif, M.; Kelsey, G.; Van den Veyver, I.B. DNA Methylation Dynamics in the Female Germline and Maternal-Effect Mutations That Disrupt Genomic Imprinting. Genes 2021, 12, 1214. [Google Scholar] [CrossRef]
- Demond, H.; Kelsey, G. The Enigma of DNA Methylation in the Mammalian Oocyte. F1000Research 2020, 9, F1000 Faculty Rev-146. [Google Scholar] [CrossRef]
- Weaver, J.R.; Bartolomei, M.S. Chromatin Regulators of Genomic Imprinting. Biochim. Biophys. Acta 2014, 1839, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Bonnet-Garnier, A.; Feuerstein, P.; Chebrout, M.; Fleurot, R.; Jan, H.-U.; Debey, P.; Beaujean, N. Genome Organization and Epigenetic Marks in Mouse Germinal Vesicle Oocytes. Int. J. Dev. Biol. 2012, 56, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, S.; Liu, H.; Kaneko, N.; Ooga, M.; Nagata, M.; Aoki, F. Alterations in Epigenetic Modifications during Oocyte Growth in Mice. Reproduction 2007, 133, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Stewart, K.R.; Veselovska, L.; Kim, J.; Huang, J.; Saadeh, H.; Tomizawa, S.; Smallwood, S.A.; Chen, T.; Kelsey, G. Dynamic Changes in Histone Modifications Precede de Novo DNA Methylation in Oocytes. Genes Dev. 2015, 29, 2449–2462. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, R.D.; Saitoh, S.; Horsthemke, B. Imprinting in Prader-Willi and Angelman Syndromes. Trends Genet. 1998, 14, 194–200. [Google Scholar] [CrossRef]
- Poole, R.L.; Leith, D.J.; Docherty, L.E.; Shmela, M.E.; Gicquel, C.; Splitt, M.; Temple, I.K.; Mackay, D.J.G. Beckwith-Wiedemann Syndrome Caused by Maternally Inherited Mutation of an OCT-Binding Motif in the IGF2/H19-Imprinting Control Region, ICR1. Eur. J. Hum. Genet. 2012, 20, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Ishida, M. New Developments in Silver-Russell Syndrome and Implications for Clinical Practice. Epigenomics 2016, 8, 563–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begemann, M.; Rezwan, F.I.; Beygo, J.; Docherty, L.E.; Kolarova, J.; Schroeder, C.; Buiting, K.; Chokkalingam, K.; Degenhardt, F.; Wakeling, E.L.; et al. Maternal Variants in NLRP and Other Maternal Effect Proteins Are Associated with Multilocus Imprinting Disturbance in Offspring. J. Med. Genet. 2018, 55, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Boonen, S.E.; Mackay, D.J.G.; Hahnemann, J.M.D.; Docherty, L.; Grønskov, K.; Lehmann, A.; Larsen, L.G.; Haemers, A.P.; Kockaerts, Y.; Dooms, L.; et al. Transient Neonatal Diabetes, ZFP57, and Hypomethylation of Multiple Imprinted Loci: A Detailed Follow-Up. Diabetes Care 2013, 36, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Kou, Y.C.; Shao, L.; Peng, H.H.; Rosetta, R.; del Gaudio, D.; Wagner, A.F.; Al-Hussaini, T.K.; Van den Veyver, I.B. A Recurrent Intragenic Genomic Duplication, Other Novel Mutations in NLRP7 and Imprinting Defects in Recurrent Biparental Hydatidiform Moles. Mol. Hum. Reprod. 2008, 14, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, D.J.G.; Callaway, J.L.A.; Marks, S.M.; White, H.E.; Acerini, C.L.; Boonen, S.E.; Dayanikli, P.; Firth, H.V.; Goodship, J.A.; Haemers, A.P.; et al. Hypomethylation of Multiple Imprinted Loci in Individuals with Transient Neonatal Diabetes Is Associated with Mutations in ZFP57. Nat. Genet. 2008, 40, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Barberet, J.; Binquet, C.; Guilleman, M.; Doukani, A.; Choux, C.; Bruno, C.; Bourredjem, A.; Chapusot, C.; Bourc’his, D.; Duffourd, Y.; et al. Do Assisted Reproductive Technologies and in Vitro Embryo Culture Influence the Epigenetic Control of Imprinted Genes and Transposable Elements in Children? Hum. Reprod. 2021, 36, 479–492. [Google Scholar] [CrossRef]
- Barberet, J.; Romain, G.; Binquet, C.; Guilleman, M.; Bruno, C.; Ginod, P.; Chapusot, C.; Choux, C.; Fauque, P. Do Frozen Embryo Transfers Modify the Epigenetic Control of Imprinted Genes and Transposable Elements in Newborns Compared with Fresh Embryo Transfers and Natural Conceptions? Fertil. Steril. 2021, 116, 1468–1480. [Google Scholar] [CrossRef]
- Hattori, H.; Hiura, H.; Kitamura, A.; Miyauchi, N.; Kobayashi, N.; Takahashi, S.; Okae, H.; Kyono, K.; Kagami, M.; Ogata, T.; et al. Association of Four Imprinting Disorders and ART. Clin. Epigenetics 2019, 11, 21. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.-F.; Huang, Y.-F.; Huang, R.-F.; Lin, S.-X.; Guo, C.-Q.; Hua, C.-Z.; Wu, P.-Y.; Hu, J.-F.; Li, Y.-Z. Effects of Vitrification on the Imprinted Gene Snrpn in Neonatal Placental Tissue. Reprod. Dev. Med. 2020, 4, 25–31. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, Y.; Wen, L.; Lei, H.; Chen, S.; Wang, X. Changes in DNA Methylation and Imprinting Disorders in E9.5 Mouse Fetuses and Placentas Derived from Vitrified Eight-Cell Embryos. Mol. Reprod. Dev. 2019, 86, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xu, L.; He, F. Embryo Vitrification Affects the Methylation of the H19/Igf2 Differentially Methylated Domain and the Expression of H19 and Igf2. Fertil. Steril. 2010, 93, 2729–2733. [Google Scholar] [CrossRef] [PubMed]
- Henningsen, A.A.; Gissler, M.; Rasmussen, S.; Opdahl, S.; Wennerholm, U.B.; Spangsmose, A.L.; Tiitinen, A.; Bergh, C.; Romundstad, L.B.; Laivuori, H.; et al. Imprinting Disorders in Children Born after ART: A Nordic Study from the CoNARTaS Group. Hum. Reprod. 2020, 35, 1178–1184. [Google Scholar] [CrossRef]
- Lazaraviciute, G.; Kauser, M.; Bhattacharya, S.; Haggarty, P.; Bhattacharya, S. A Systematic Review and Meta-Analysis of DNA Methylation Levels and Imprinting Disorders in Children Conceived by IVF/ICSI Compared with Children Conceived Spontaneously. Hum. Reprod. Update 2014, 20, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Steptoe, P.C.; Edwards, R.G. Birth after the Reimplantation of a Human Embryo. Lancet 1978, 2, 366. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Rosenwaks, Z. Assisted Reproductive Techniques. Methods Mol. Biol. 2014, 1154, 171–231. [Google Scholar]
- El Tokhy, O.; Salman, M.; El-Toukhy, T. Preimplantation Genetic Diagnosis. Obstet. Gynaecol. Reprod. Med. 2021, 31, 157–161. [Google Scholar] [CrossRef]
- Rodriguez-Wallberg, K.A.; Waterstone, M.; Anastácio, A. Ice Age: Cryopreservation in Assisted Reproduction—An Update. Reprod. Biol. 2019, 19, 119–126. [Google Scholar] [CrossRef]
- Yding Andersen, C.; Mamsen, L.S.; Kristensen, S.G. Fertility preservation: Freezing of Ovarian Tissue and Clinical Opportunities. Reproduction 2019, 158, 27–34. [Google Scholar] [CrossRef]
- Dolmans, M.-M.; Donnez, J.; Cacciottola, L. Fertility Preservation: The Challenge of Freezing and Transplanting Ovarian Tissue. Trends Mol. Med. 2021, 27, 777–791. [Google Scholar] [CrossRef]
- Deanesly, R. Immature Rat Ovaries Grafted after Freezing and Thawing. J. Endocrinol. 1954, 11, 197–200. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.M.; Demylle, D.; Jadoul, P.; Pirard, C.; Squifflet, J.; Martinez-Madrid, B.; van Langendonckt, A. Livebirth after Orthotopic Transplantation of Cryopreserved Ovarian Tissue. Lancet 2004, 364, 1405–1410. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo Signaling Disruption and Akt Stimulation of Ovarian Follicles for Infertility Treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful Fertility Preservation Following Ovarian Tissue Vitrification in Patients with Primary Ovarian Insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas Leonel, E.C.; Lucci, C.M.; Amorim, C.A. Cryopreservation of Human Ovarian Tissue: A Review. Transfus. Med. Hemotherapy 2019, 46, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Han, Y.; Zhang, J.; Zhang, S.; Zhai, Y.; An, X.; Li, Q.; Duan, J.; Zhang, X.; Li, Z.; et al. Effects of Dimethyl Sulfoxide (DMSO) on DNA Methylation and Histone Modification in Parthenogenetically Activated Porcine Embryos. Reprod. Fertil. Dev. 2022, 34, 598–607. [Google Scholar] [CrossRef]
- Iwatani, M.; Ikegami, K.; Kremenska, Y.; Hattori, N.; Tanaka, S.; Yagi, S.; Shiota, K. Dimethyl Sulfoxide Has an Impact on Epigenetic Profile in Mouse Embryoid Body. Stem Cells 2006, 24, 2549–2556. [Google Scholar] [CrossRef] [PubMed]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO Induces Drastic Changes in Human Cellular Processes and Epigenetic Landscape in Vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Obata, R.; Okuyama, N.; Aono, N.; Hashimoto, T.; Kyono, K. Residual Ethylene Glycol and Dimethyl Sulphoxide Concentration in Human Ovarian Tissue during Warming/Thawing Steps Following Cryopreservation. Reprod. Biomed. Online 2017, 35, 311–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, M.J.; Pendola, J.K.; Eppig, J.J. A Revised Protocol for in Vitro Development of Mouse Oocytes from Primordial Follicles Dramatically Improves Their Developmental Competence. Biol. Reprod. 2003, 68, 1682–1686. [Google Scholar] [CrossRef] [Green Version]
- Telfer, E.E.; Andersen, C.Y. In Vitro Growth and Maturation of Primordial Follicles and Immature Oocytes. Fertil. Steril. 2021, 115, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.; Albertini, D.F.; Wallace, W.H.B.; Anderson, R.A.; Telfer, E.E. Metaphase II Oocytes from Human Unilaminar Follicles Grown in a Multi-Step Culture System. Mol. Hum. Reprod. 2018, 24, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhang, L.; Wang, Z.; Chang, H.; Xie, X.; Fu, L.; Zhang, Y.; Quan, F. Resveratrol Improved the Developmental Potential of Oocytes after Vitrification by Modifying the Epigenetics. Mol. Reprod. Dev. 2019, 86, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.-R.; Fu, X.-W.; Zhang, R.-N.; Jia, G.-X.; Hou, Y.-P.; Zhu, S.-E. Effect of Oocyte Vitrification on Deoxyribonucleic Acid Methylation of H19, Peg3, and Snrpn Differentially Methylated Regions in Mouse Blastocysts. Fertil. Steril. 2014, 102, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Long, C.; Liu, G.; Bai, H.; Ma, L.; Bai, T.; Zuo, Y.; Li, S. WGBS Combined with RNA-Seq Analysis Revealed That Dnmt1 Affects the Methylation Modification and Gene Expression Changes during Mouse Oocyte Vitrification. Theriogenology 2022, 177, 11–21. [Google Scholar] [CrossRef]
- Bebbere, D.; Masala, L.; Albertini, D.F.; Ledda, S. The Subcortical Maternal Complex: Multiple Functions for One Biological Structure? J. Assist. Reprod. Genet. 2016, 33, 1431–1438. [Google Scholar] [CrossRef] [Green Version]
- Schultz, R.M.; Stein, P.; Svoboda, P. The Oocyte-to-Embryo Transition in Mouse: Past, Present, and Future. Biol. Reprod. 2018, 99, 160–174. [Google Scholar] [CrossRef]
- Gu, R.; Ge, N.; Huang, B.; Fu, J.; Zhang, Y.; Wang, N.; Xu, Y.; Li, L.; Peng, X.; Zou, Y.; et al. Impacts of Vitrification on the Transcriptome of Human Ovarian Tissue in Patients with Gynecological Cancer. Front. Genet. 2023, 14, 1114650. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, W.; Todorov, P.; Pei, C.; Isachenko, E.; Rahimi, G.; Mallmann, P.; Nawroth, F.; Isachenko, V. RNA Transcripts in Human Ovarian Cells: Two-Time Cryopreservation Does Not Affect Developmental Potential. Int. J. Mol. Sci. 2023, 24, 6880. [Google Scholar] [CrossRef]
- He, Z.-Y.; Wang, H.-Y.; Zhou, X.; Liang, X.-Y.; Yan, B.; Wang, R.; Ma, L.-H.; Wang, Y.-L. Evaluation of Vitrification Protocol of Mouse Ovarian Tissue by Effect of DNA Methyltransferase-1 and Paternal Imprinted Growth Factor Receptor-Binding Protein 10 on Signaling Pathways. Cryobiology 2018, 80, 89–95. [Google Scholar] [CrossRef]
- Shirazi, A.; Naderi, M.M.; Hassanpour, H.; Heidari, M.; Borjian, S.; Sarvari, A.; Akhondi, M.M. The Effect of Ovine Oocyte Vitrification on Expression of Subset of Genes Involved in Epigenetic Modifications during Oocyte Maturation and Early Embryo Development. Theriogenology 2016, 86, 2136–2146. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Li, Q.; Zhang, L.; Kang, B.; Fan, W.; Deng, T.; Zhu, J.; Wang, Y. The Growth and Development Conditions in Mouse Offspring Derived from Ovarian Tissue Cryopreservation and Orthotopic Transplantation. J. Assist. Reprod. Genet. 2020, 37, 923–932. [Google Scholar] [CrossRef]
- Demant, M.; Trapphoff, T.; Fröhlich, T.; Arnold, G.J.; Eichenlaub-Ritter, U. Vitrification at the Pre-Antral Stage Transiently Alters Inner Mitochondrial Membrane Potential but Proteome of in Vitro Grown and Matured Mouse Oocytes Appears Unaffected. Hum. Reprod. 2012, 27, 1096–1111. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, M.J.; Siatkowski, M.; Paudel, Y.; Balbach, S.T.; Baeumer, N.; Crosetto, N.; Drexler, H.C.A.; Fuellen, G.; Boiani, M. Proteomic Analysis of Mouse Oocytes Reveals 28 Candidate Factors of the “Reprogrammome”. J. Proteome Res. 2011, 10, 2140–2153. [Google Scholar] [CrossRef]
- Wang, H.-Y.; Li, Y.-H.; Sun, L.; Gao, X.; You, L.; Wang, Y.; Ma, J.-L.; Chen, Z.-J. Allotransplantation of Cryopreserved Prepubertal Mouse Ovaries Restored Puberty and Fertility without Affecting Methylation Profile of Snrpn-DMR. Fertil. Steril. 2013, 99, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, L.; Wang, T.; Li, R.; Liu, P.; Yan, L.; Qiao, J. Effect of Vitrification at the Germinal Vesicle Stage on the Global Methylation Status in Mouse Oocytes Subsequently Matured in Vitro. Chin. Med. J. 2014, 127, 4019–4024. [Google Scholar] [PubMed]
- Al-Khtib, M.; Perret, A.; Khoueiry, R.; Ibala-Romdhane, S.; Blachère, T.; Greze, C.; Lornage, J.; Lefèvre, A. Vitrification at the Germinal Vesicle Stage Does Not Affect the Methylation Profile of H19 and KCNQ1OT1 Imprinting Centers in Human Oocytes Subsequently Matured in Vitro. Fertil. Steril. 2011, 95, 1955–1960. [Google Scholar] [CrossRef]
- Yodrug, T.; Parnpai, R.; Hirao, Y.; Somfai, T. Effect of Vitrification at Different Meiotic Stages on Epigenetic Characteristics of Bovine Oocytes and Subsequently Developing Embryos. Anim. Sci. J. 2021, 92, e13596. [Google Scholar] [CrossRef]
- Trapphoff, T.; El Hajj, N.; Zechner, U.; Haaf, T.; Eichenlaub-Ritter, U. DNA Integrity, Growth Pattern, Spindle Formation, Chromosomal Constitution and Imprinting Patterns of Mouse Oocytes from Vitrified Pre-Antral Follicles. Hum. Reprod. 2010, 25, 3025–3042. [Google Scholar] [CrossRef] [Green Version]
- Damavandi, M.; Farrokh, P.; Zavareh, S. Effect of Mouse Ovarian Vitrification on Promoter Methylation of Inhba and Inhbb in Granulosa Cells of Follicles. Cryo Lett. 2021, 42, 67–72. [Google Scholar]
- Sauvat, F.; Capito, C.; Sarnacki, S.; Poirot, C.; Bachelot, A.; Meduri, G.; Dandolo, L.; Binart, N. Immature Cryopreserved Ovary Restores Puberty and Fertility in Mice without Alteration of Epigenetic Marks. PLoS ONE 2008, 3, e1972. [Google Scholar] [CrossRef] [PubMed]
- Sauvat, F.; Bouilly, J.; Capito, C.; Lefèvre, A.; Blachère, T.; Borenstein, N.; Sarnacki, S.; Dandolo, L.; Binart, N. Ovarian Function Is Restored after Grafting of Cryopreserved Immature Ovary in Ewes. FASEB J. 2013, 27, 1511–1518. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Shen, L.; Gong, C.; Cao, Y.; Shi, Q.; Zhao, G. Microencapsulation and Nanowarming Enables Vitrification Cryopreservation of Mouse Preantral Follicles. Nat. Commun. 2022, 13, 7515. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-C.; Comizzoli, P. Desiccation and Supra-Zero Temperature Storage of Cat Germinal Vesicles Lead to Less Structural Damage and Similar Epigenetic Alterations Compared to Cryopreservation. Mol. Reprod. Dev. 2019, 86, 1822–1831. [Google Scholar] [CrossRef] [PubMed]
- Tamburrino, L.; Traini, G.; Marcellini, A.; Vignozzi, L.; Baldi, E.; Marchiani, S. Cryopreservation of Human Spermatozoa: Functional, Molecular and Clinical Aspects. Int. J. Mol. Sci. 2023, 24, 4656. [Google Scholar] [CrossRef]
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European Perspective on Testicular Tissue Cryopreservation for Fertility Preservation in Prepubertal and Adolescent Boys. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef] [Green Version]
- Delgouffe, E.; Braye, A.; Goossens, E. Testicular Tissue Banking for Fertility Preservation in Young Boys: Which Patients Should Be Included? Front. Endocrinol. 2022, 13, 854186. [Google Scholar] [CrossRef]
- Sanou, I.; van Maaren, J.; Eliveld, J.; Lei, Q.; Meißner, A.; de Melker, A.A.; Hamer, G.; van Pelt, A.M.M.; Mulder, C.L. Spermatogonial Stem Cell-Based Therapies: Taking Preclinical Research to the Next Level. Front. Endocrinol. 2022, 13, 850219. [Google Scholar] [CrossRef]
- Wyns, C.; Kanbar, M.; Giudice, M.G.; Poels, J. Fertility Preservation for Prepubertal Boys: Lessons Learned from the Past and Update on Remaining Challenges towards Clinical Translation. Hum. Reprod. Update 2021, 27, 433–459. [Google Scholar] [CrossRef]
- Onofre, J.; Baert, Y.; Faes, K.; Goossens, E. Cryopreservation of Testicular Tissue or Testicular Cell Suspensions: A Pivotal Step in Fertility Preservation. Hum. Reprod. Update 2016, 22, 744–761. [Google Scholar] [CrossRef] [Green Version]
- Onofre, J.; Kadam, P.; Baert, Y.; Goossens, E. Testicular Tissue Cryopreservation Is the Preferred Method to Preserve Spermatogonial Stem Cells Prior to Transplantation. Reprod. Biomed. Online 2020, 40, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayomi, A.P.; Peters, K.; Sukhwani, M.; Valli-Pulaski, H.; Shetty, G.; Meistrich, M.L.; Houser, L.; Robertson, N.; Roberts, V.; Ramsey, C.; et al. Autologous Grafting of Cryopreserved Prepubertal Rhesus Testis Produces Sperm and Offspring. Science 2019, 363, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Goossens, E.; De Rycke, M.; Haentjens, P.; Tournaye, H. DNA Methylation Patterns of Spermatozoa and Two Generations of Offspring Obtained after Murine Spermatogonial Stem Cell Transplantation. Hum. Reprod. 2009, 24, 2255–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, J.B.; van Eekelen, R.; de Winter-Korver, C.M.; van Daalen, S.K.M.; Tabeling, N.C.; Catsburg, L.A.E.; Gijbels, M.J.J.; Mulder, C.L.; van Pelt, A.M.M. Impact of Restoring Male Fertility with Transplantation of in Vitro Propagated Spermatogonial Stem Cells on the Health of Their Offspring throughout Life. Clin. Transl. Med. 2021, 11, e531. [Google Scholar] [CrossRef]
- Nickkholgh, B.; Mizrak, S.C.; van Daalen, S.K.M.; Korver, C.M.; Sadri-Ardekani, H.; Repping, S.; van Pelt, A.M.M. Genetic and Epigenetic Stability of Human Spermatogonial Stem Cells during Long-Term Culture. Fertil. Steril. 2014, 102, 1700–1707. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Ogonuki, N.; Iwano, T.; Lee, J.; Kazuki, Y.; Inoue, K.; Miki, H.; Takehashi, M.; Toyokuni, S.; Shinkai, Y.; et al. Genetic and Epigenetic Properties of Mouse Male Germline Stem Cells during Long-Term Culture. Development 2005, 132, 4155–4163. [Google Scholar] [CrossRef] [Green Version]
- Langenstroth-Röwer, D.; Gromoll, J.; Wistuba, J.; Tröndle, I.; Laurentino, S.; Schlatt, S.; Neuhaus, N. De Novo Methylation in Male Germ Cells of the Common Marmoset Monkey Occurs during Postnatal Development and Is Maintained in Vitro. Epigenetics 2017, 12, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In Vitro Production of Functional Sperm in Cultured Neonatal Mouse Testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef]
- Yokonishi, T.; Sato, T.; Komeya, M.; Katagiri, K.; Kubota, Y.; Nakabayashi, K.; Hata, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; et al. Offspring Production with Sperm Grown in Vitro from Cryopreserved Testis Tissues. Nat. Commun. 2014, 5, 4320. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Goodyear, S.M.; Abramowitz, L.K.; Bartolomei, M.S.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Fertile Offspring Derived from Mouse Spermatogonial Stem Cells Cryopreserved for More than 14 Years. Hum. Reprod. 2012, 27, 1249–1259. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.B.; Tabeling, N.C.; de Winter-Korver, C.M.; van Daalen, S.K.M.; van Pelt, A.M.M.; Mulder, C.L. Sperm DNA Methylation Is Predominantly Stable in Mice Offspring Born after Transplantation of Long-Term Cultured Spermatogonial Stem Cells. Clin. Epigenetics 2023, 15, 58. [Google Scholar] [CrossRef]
- Oblette, A.; Rives-Feraille, A.; Dumont, L.; Delessard, M.; Saulnier, J.; Rives, N.; Rondanino, C. Dynamics of Epigenetic Modifications in ICSI Embryos from in Vitro-Produced Spermatozoa. Andrology 2021, 9, 640–656. [Google Scholar] [CrossRef]
- Oblette, A.; Rondeaux, J.; Dumont, L.; Delessard, M.; Saulnier, J.; Rives, A.; Rives, N.; Rondanino, C. DNA Methylation and Histone Post-Translational Modifications in the Mouse Germline Following in-Vitro Maturation of Fresh or Cryopreserved Prepubertal Testicular Tissue. Reprod. Biomed. Online 2019, 39, 383–401. [Google Scholar] [CrossRef]
- Kawai, K.; Li, Y.-S.; Song, M.-F.; Kasai, H. DNA Methylation by Dimethyl Sulfoxide and Methionine Sulfoxide Triggered by Hydroxyl Radical and Implications for Epigenetic Modifications. Bioorg. Med. Chem. Lett. 2010, 20, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A Preliminary Study on Epigenetic Changes during Boar Spermatozoa Cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Khosravizadeh, Z.; Hassanzadeh, G.; Tavakkoly Bazzaz, J.; Alizadeh, F.; Totonchi, M.; Salehi, E.; Khodamoradi, K.; Khanehzad, M.; Hosseini, S.R.; Abolhassani, F. The Effect of Cryopreservation on DNA Methylation Patterns of the Chromosome 15q11-Q13 Region in Human Spermatozoa. Cell Tissue Bank. 2020, 21, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Kläver, R.; Bleiziffer, A.; Redmann, K.; Mallidis, C.; Kliesch, S.; Gromoll, J. Routine Cryopreservation of Spermatozoa Is Safe-Evidence from the DNA Methylation Pattern of Nine Spermatozoa Genes. J. Assist. Reprod. Genet. 2012, 29, 943–950. [Google Scholar] [CrossRef] [Green Version]




| Reference | Type/Species (Cryopreservation Technique) | Analysis (Technique) | Main Outcome |
|---|---|---|---|
| Oblette et al. (2019) [83] | Murine in vitro culture of TT (solid surface vitrification including 2.1 mol/L DMSO and 2.7 mol/L EG or slow freezing including 1.5 mol/L DMSO) | Testicular tissue after IVC (qPCR and immunofluorescent assay) | Normal expression of epigenetic modification enzymes and gDNA methylation, but differences in histone modification |
| Oblette et al. (2021) [82] | Murine in vitro culture of TT (slow freezing including 1.5M DMSO) | Pre-implantation embryo (immunofluorescent assay) | Normal post-translational histone modifications and altered gDNA methylation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trapphoff, T.; Dieterle, S. Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern. Int. J. Mol. Sci. 2023, 24, 11061. https://doi.org/10.3390/ijms241311061
Trapphoff T, Dieterle S. Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern. International Journal of Molecular Sciences. 2023; 24(13):11061. https://doi.org/10.3390/ijms241311061
Chicago/Turabian StyleTrapphoff, Tom, and Stefan Dieterle. 2023. "Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern" International Journal of Molecular Sciences 24, no. 13: 11061. https://doi.org/10.3390/ijms241311061
APA StyleTrapphoff, T., & Dieterle, S. (2023). Cryopreservation of Ovarian and Testicular Tissue and the Influence on Epigenetic Pattern. International Journal of Molecular Sciences, 24(13), 11061. https://doi.org/10.3390/ijms241311061