
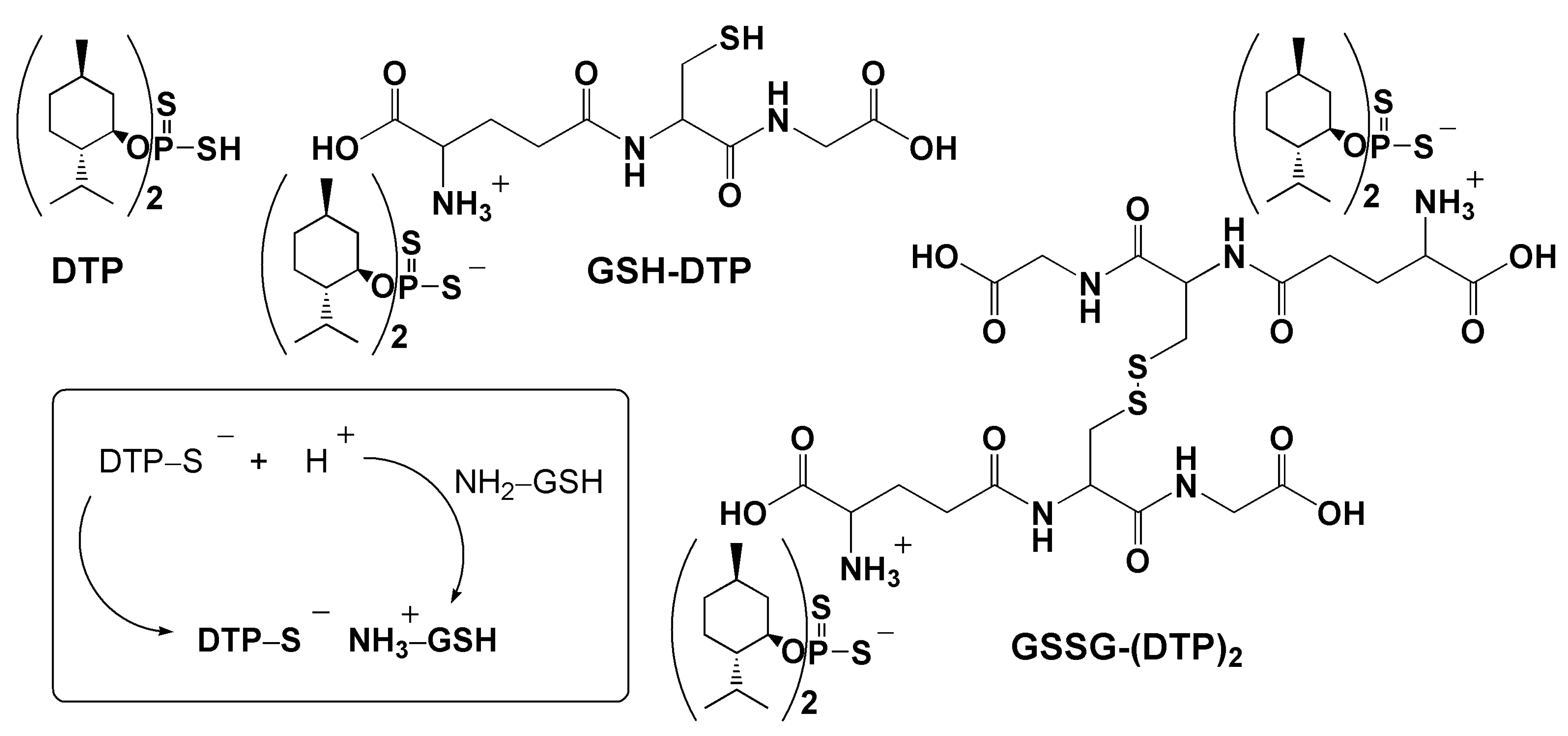
Characterization of Glutathione Dithiophosphates as Long-Acting H2S Donors

 ,
,  and
and 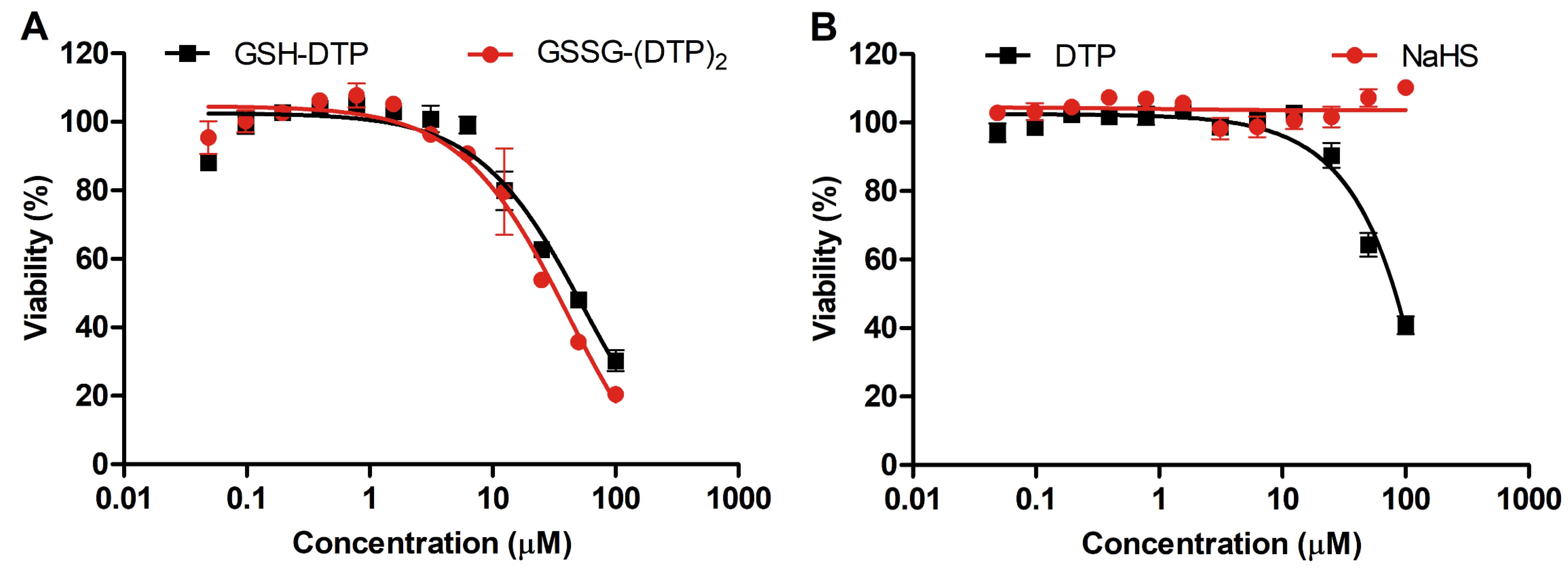{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cytotoxicity of Glutathione Dithiophosphates
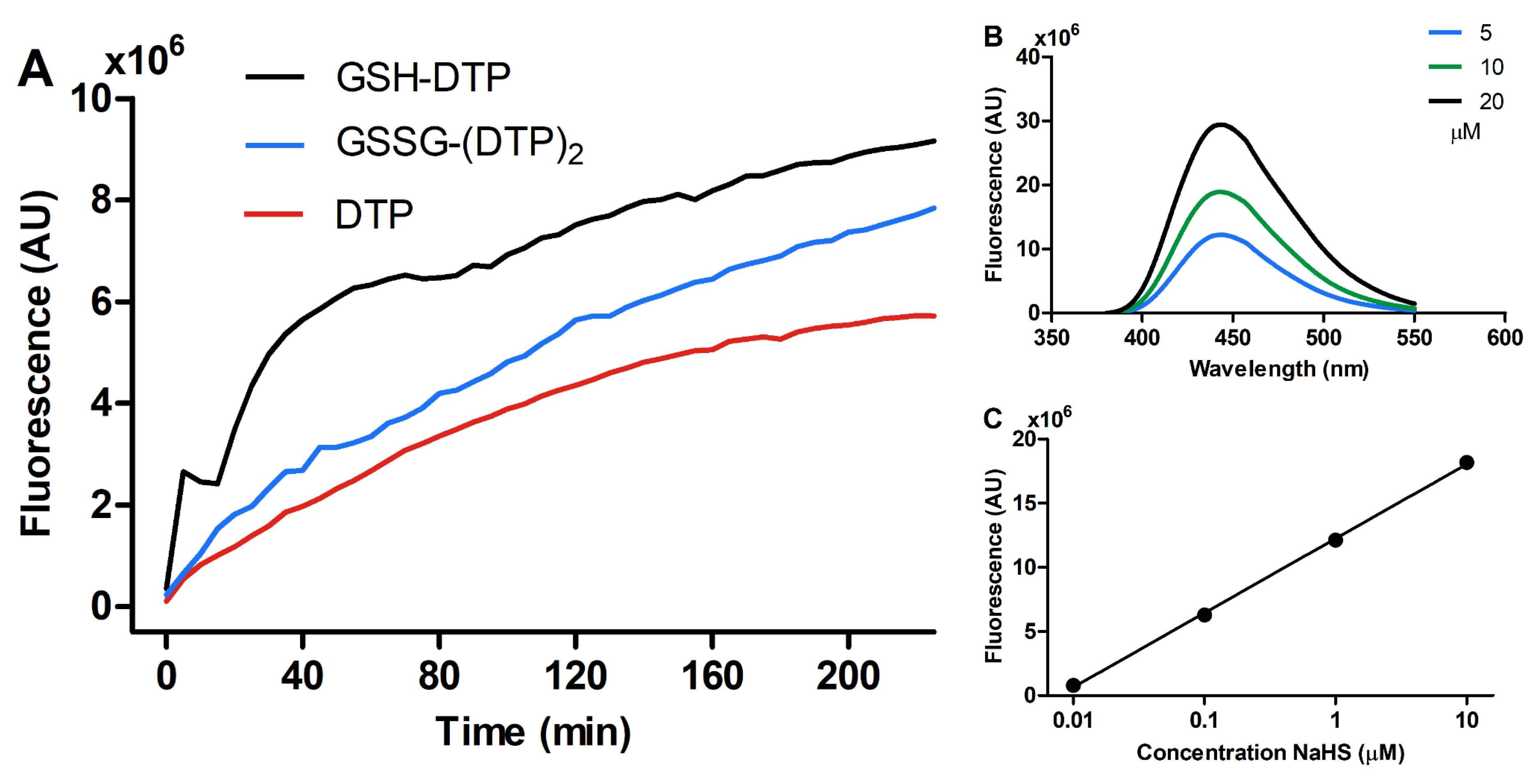
2.2. H2S-Releasing Profile
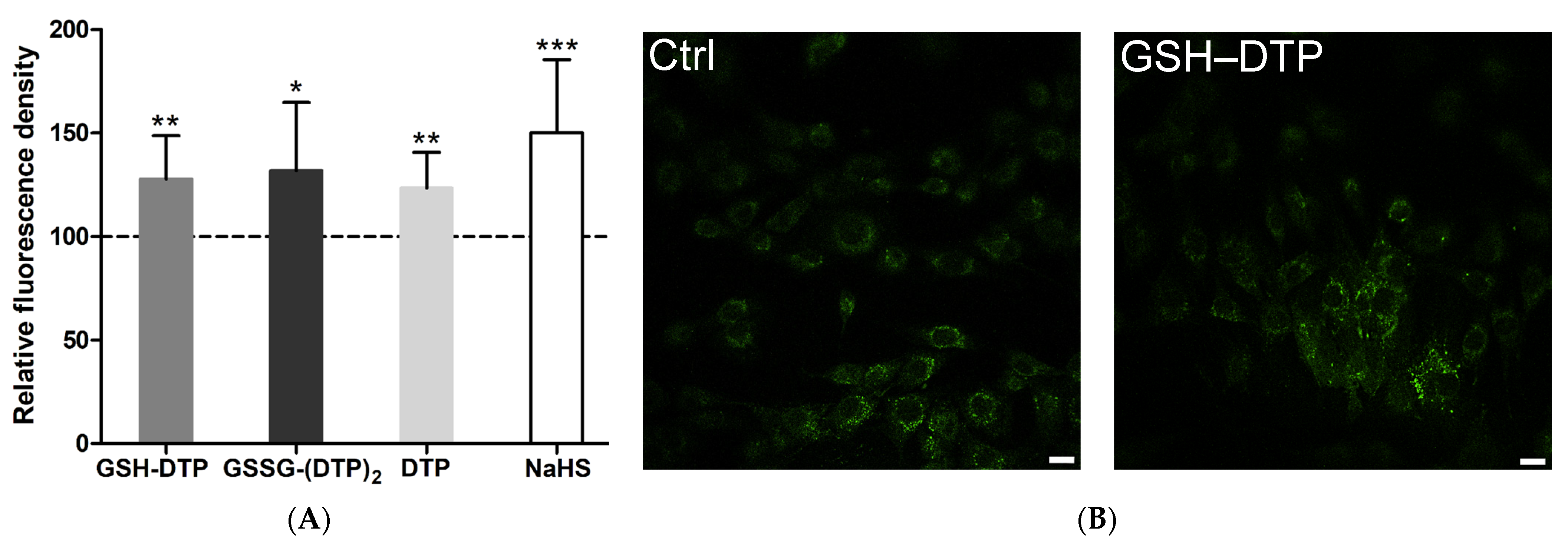
2.3. Effect on Cellular H2S
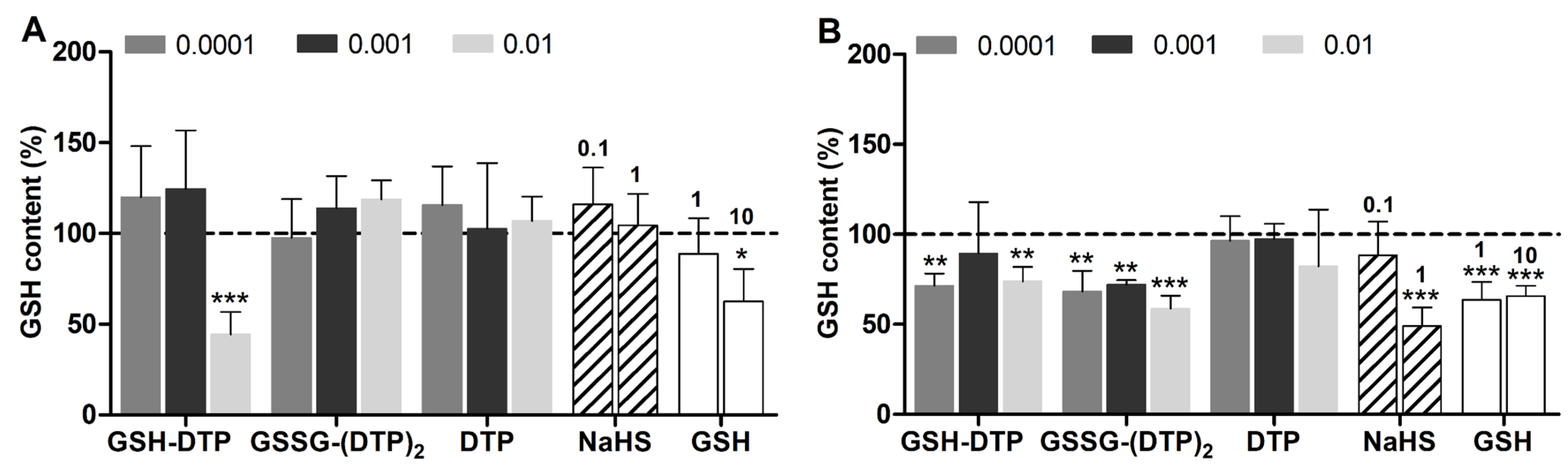
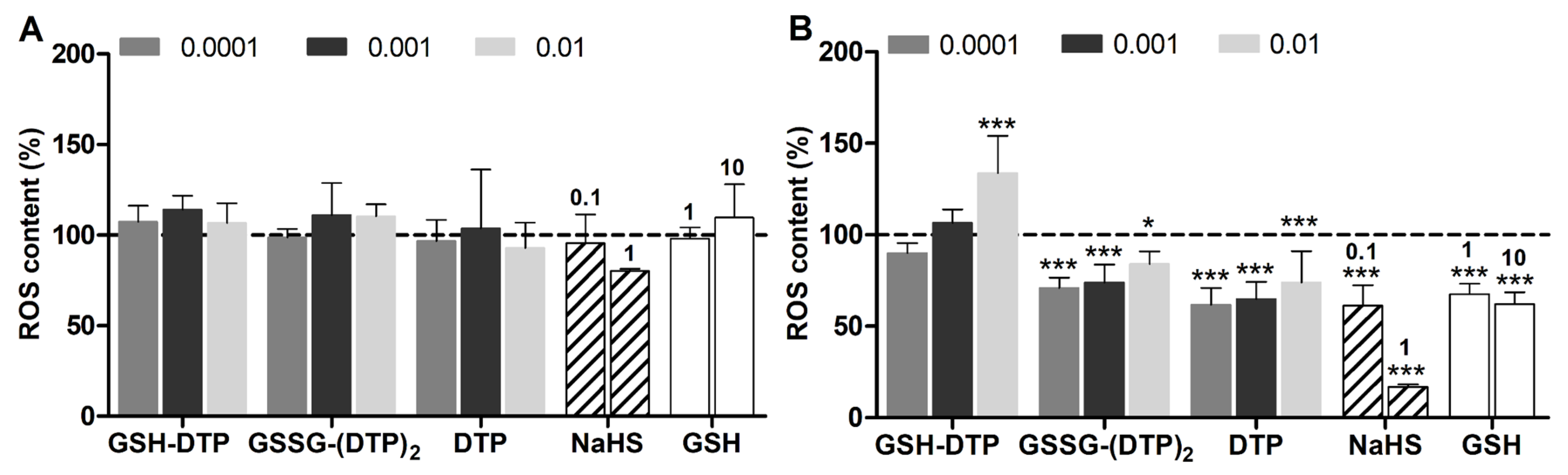
2.4. Effect on Cellular GSH and ROS
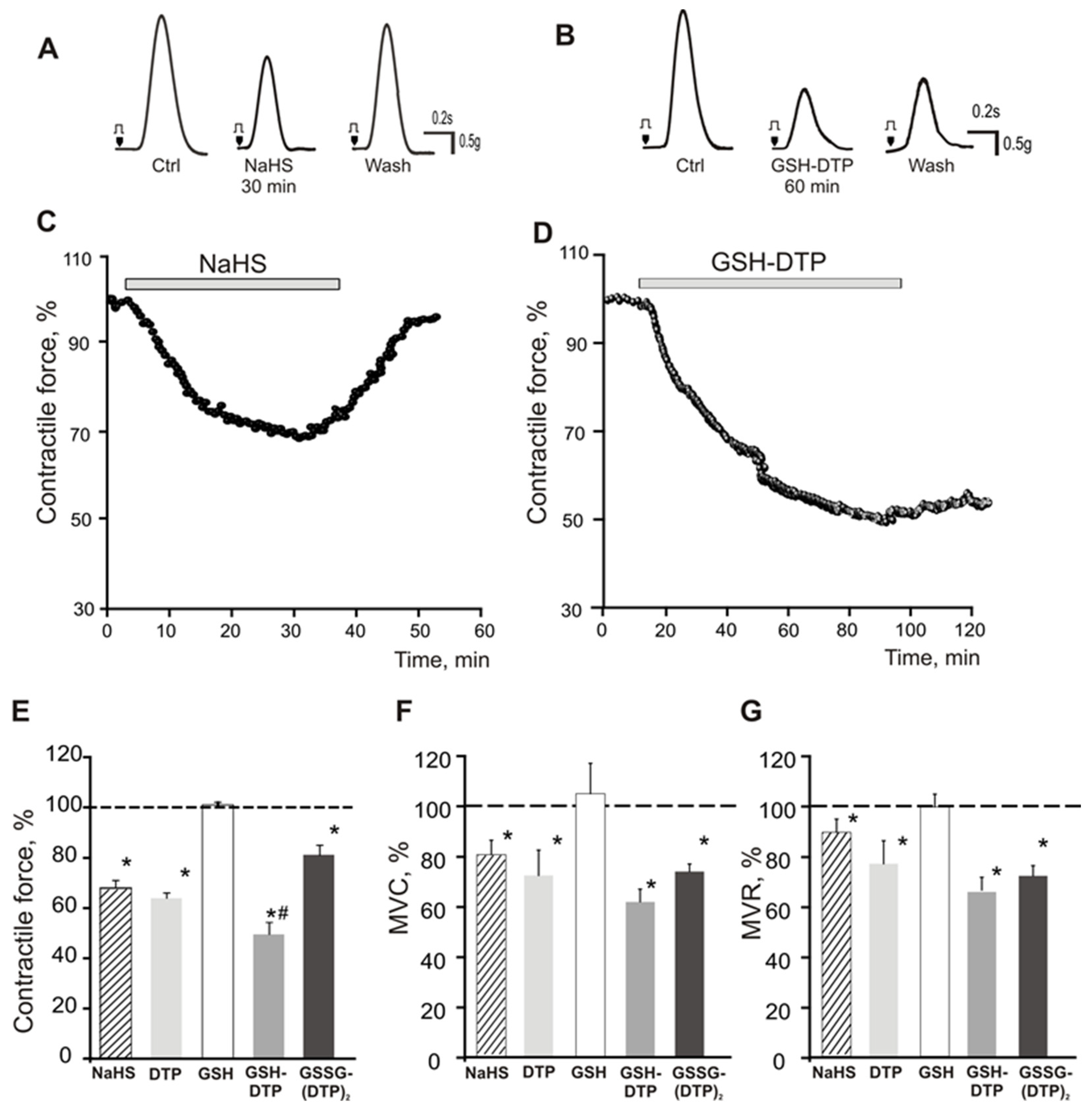
2.5. Effect on Contractile Activity of Myocardium Stripes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Viability Assessment
4.3. Detection of H2S Release
4.4. Laser Scanning Confocal Microscopy (LSCM)
4.5. GSH and ROS Detection
4.6. Study of Isolated Rat Right Atrial Contraction
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hermann, A.; Sitdikova, G.F.; Weiger, T.M. Gasotransmitters: Physiology and Pathophysiology; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Cirino, G.; Szabo, C.; Papapetropoulos, A. Physiological roles of hydrogen sulfide in mammalian cells, tissues, and organs. Physiol. Rev. 2023, 103, 31–276. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Mesfin, F.M.; Hunter, C.E.; Olson, K.R.; Shelley, W.C.; Brokaw, J.P.; Manohar, K.; Markel, T.A. Recent Development of the Molecular and Cellular Mechanisms of Hydrogen Sulfide Gasotransmitter. Antioxidants 2022, 11, 1788. [Google Scholar] [CrossRef] [PubMed]
- Sitdikova, G.F.; Fuchs, R.; Kainz, V.; Weiger, T.M.; Hermann, A. Phosphorylation of BK channels modulates the sensitivity to hydrogen sulfide (H2S). Front. Physiol. 2014, 5, 431. [Google Scholar] [CrossRef] [Green Version]
- Kabil, O.; Motl, N.; Banerjee, R. H2S and its role in redox signaling. Biochim. Biophys. Acta BBA Proteins Proteom. 2014, 1844, 1355–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuevasanta, E.; Benchoam, D.; Semelak, J.A.; Möller, M.N.; Zeida, A.; Trujillo, M.; Alvarez, B.; Estrin, D.A. Possible molecular basis of the biochemical effects of cysteine-derived persulfides. Front. Mol. Biosci. 2022, 9, 975988. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen Sulfide (H2S) and Polysulfide (H2Sn) Signaling: The First 25 Years. Biomolecules 2021, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kowalczyk-Pachel, D.; Bilska-Wilkosz, A.; Kwiecień, I.; Górny, M.; Włodek, L. S-sulfhydration as a cellular redox regulation. Biosci. Rep. 2016, 36, e00304. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a Physiologic Vasorelaxant: Hypertension in Mice with Deletion of Cystathionine γ-Lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Alsaeedi, A.; Welham, S.; Rose, P.; Zhu, Y.-Z. The Impact of Drugs on Hydrogen Sulfide Homeostasis in Mammals. Antioxidants 2023, 12, 908. [Google Scholar] [CrossRef]
- Gao, Y.; Yao, X.; Zhang, Y.; Li, W.; Kang, K.; Sun, L.; Sun, X. The protective role of hydrogen sulfide in myocardial ischemia–reperfusion-induced injury in diabetic rats. Int. J. Cardiol. 2011, 152, 177–183. [Google Scholar] [CrossRef]
- Kolluru, G.K.; Shackelford, R.E.; Shen, X.; Dominic, P.; Kevil, C.G. Sulfide regulation of cardiovascular function in health and disease. Nat. Rev. Cardiol. 2023, 20, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Bull, R.; Rains, J.L.; Bass, P.F.; Levine, S.N.; Reddy, S.; McVie, R.; Bocchini, J.A. Low Levels of Hydrogen Sulfide in the Blood of Diabetes Patients and Streptozotocin-Treated Rats Causes Vascular Inflammation? Antioxid. Redox Signal. 2010, 12, 1333–1337. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Two’s company, three’s a crowd: Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, J.L.; Vaughan, D.; Dicay, M.; MacNaughton, W.K.; de Nucci, G. Hydrogen Sulfide-Releasing Therapeutics: Translation to the Clinic. Antioxid. Redox Signal. 2017, 28, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.W.; Deng, N.H.; Tian, K.J.; Liu, L.S.; Wang, Z.; Wei, D.H.; Liu, H.T.; Jiang, Z.S. Development of hydrogen sulfide donors for anti-atherosclerosis therapeutics research: Challenges and future priorities. Front. Cardiovasc. Med. 2022, 9, 909178. [Google Scholar] [CrossRef]
- Sun, Y.-G.; Cao, Y.-X.; Wang, W.-W.; Ma, S.-F.; Yao, T.; Zhu, Y.-C. Hydrogen sulphide is an inhibitor of L-type calcium channels and mechanical contraction in rat cardiomyocytes. Cardiovasc. Res. 2008, 79, 632–641. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Lukesh, J.C. H2S Donors with Cytoprotective Effects in Models of MI/R Injury and Chemotherapy-Induced Cardiotoxicity. Antioxidants 2023, 12, 650. [Google Scholar] [CrossRef]
- Mazza, R.; Pasqua, T.; Cerra, M.C.; Angelone, T.; Gattuso, A. Akt/eNOS signaling and PLN S-sulfhydration are involved in H2S-dependent cardiac effects in frog and rat. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2013, 305, R443–R451. [Google Scholar] [CrossRef] [Green Version]
- Sitdikova, G.F.; Khaertdinov, N.N.; Zefirov, A.L. Role of Calcium and Potassium Channels in Effects of Hydrogen Sulfide on Frog Myocardial Contractility. Bull. Exp. Biol. Med. 2011, 151, 163–166. [Google Scholar] [CrossRef]
- Abramochkin, D.V.; Moiseenko, L.S.; Kuzmin, V.S. The Effect of Hydrogen Sulfide on Electrical Activity of Rat Atrial Myocardium. Bull. Exp. Biol. Med. 2009, 147, 683–686. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, R. H2S-induced vasorelaxation and underlying cellular and molecular mechanisms. Am. J. Physiol.-Heart Circ. Physiol. 2002, 283, H474–H480. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; Maulik, S.K. Effect of garlic on cardiovascular disorders: A review. Nutr. J. 2002, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rossoni, G.; Sparatore, A.; Lee, L.C.; Del Soldato, P.; Moore, P.K. Anti-inflammatory and gastrointestinal effects of a novel diclofenac derivative. Free. Radic. Biol. Med. 2007, 42, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef]
- Zhao, Y.; Bhushan, S.; Yang, C.; Otsuka, H.; Stein, J.D.; Pacheco, A.; Peng, B.; Devarie-Baez, N.O.; Aguilar, H.C.; Lefer, D.J.; et al. Controllable Hydrogen Sulfide Donors and Their Activity against Myocardial Ischemia-Reperfusion Injury. ACS Chem. Biol. 2013, 8, 1283–1290. [Google Scholar] [CrossRef] [Green Version]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Pugliesi, I.; Barresi, E.; Nesi, G.; Rapposelli, S.; Taliani, S.; Da Settimo, F.; et al. Arylthioamides as H2S Donors: L-Cysteine-Activated Releasing Properties and Vascular Effects in Vitro and in Vivo. ACS Med. Chem. Lett. 2013, 4, 904–908. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, H.; Xian, M. Cysteine-Activated Hydrogen Sulfide (H2S) Donors. J. Am. Chem. Soc. 2011, 133, 15–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chun, O.K.; Song, W.O. Plasma and Dietary Antioxidant Status as Cardiovascular Disease Risk Factors: A Review of Human Studies. Nutrients 2013, 5, 2969–3004. [Google Scholar] [CrossRef] [Green Version]
- Levinn, C.M.; Cerda, M.M.; Pluth, M.D. Activatable Small-Molecule Hydrogen Sulfide Donors. Antioxid. Redox Signal. 2019, 32, 96–109. [Google Scholar] [CrossRef]
- Kang, J.; Li, Z.; Organ, C.L.; Park, C.-M.; Yang, C.-t.; Pacheco, A.; Wang, D.; Lefer, D.J.; Xian, M. pH-Controlled Hydrogen Sulfide Release for Myocardial Ischemia-Reperfusion Injury. J. Am. Chem. Soc. 2016, 138, 6336–6339. [Google Scholar] [CrossRef]
- Lane, T.; Fontana, M.; Martinez-Naharro, A.; Quarta, C.C.; Whelan, C.J.; Petrie, A.; Rowczenio, D.M.; Gilbertson, J.A.; Hutt, D.F.; Rezk, T.; et al. Natural History, Quality of Life, and Outcome in Cardiac Transthyretin Amyloidosis. Circulation 2019, 140, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; von Wantoch Rekowski, M.; Coletta, C.; Szabo, C.; Bucci, M.; Cirino, G.; Topouzis, S.; Papapetropoulos, A.; Giannis, A. Thioglycine and l-thiovaline: Biologically active H2S-donors. Bioorgan. Med. Chem. 2012, 20, 2675–2678. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.-H.; Moore, P.K. Characterization of a Novel, Water-Soluble Hydrogen Sulfide–Releasing Molecule (GYY4137). Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [Green Version]
- Gaucher, C.; Boudier, A.; Bonetti, J.; Clarot, I.; Leroy, P.; Parent, M. Glutathione: Antioxidant Properties Dedicated to Nanotechnologies. Antioxidants 2018, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Dronamraju, V.; Xie, W.; More, S.S. Sulfur-containing therapeutics in the treatment of Alzheimer’s disease. Med. Chem. Res. 2021, 30, 305–352. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Du, Y.-s.; Xu, W.-s.; Li, C.-j.; Sun, H.; Hu, K.-r.; Hu, Y.-z.; Yu, T.-j.; Guo, H.-m.; Xie, L.; et al. Exogenous glutathione exerts a therapeutic effect in ischemic stroke rats by interacting with intrastriatal dopamine. Acta Pharmacol. Sin. 2022, 43, 541–551. [Google Scholar] [CrossRef]
- Cacciatore, I.; Cornacchia, C.; Pinnen, F.; Mollica, A.; Di Stefano, A. Prodrug Approach for Increasing Cellular Glutathione Levels. Molecules 2010, 15, 1242–1264. [Google Scholar] [CrossRef] [Green Version]
- Ishkaeva, R.A.; Nizamov, I.S.; Blokhin, D.S.; Urakova, E.A.; Klochkov, V.V.; Nizamov, I.D.; Gareev, B.I.; Salakhieva, D.V.; Abdullin, T.I. Dithiophosphate-Induced Redox Conversions of Reduced and Oxidized Glutathione. Molecules 2021, 26, 2973. [Google Scholar] [CrossRef]
- Ishkaeva, R.A.; Zoughaib, M.; Laikov, A.V.; Angelova, P.R.; Abdullin, T.I. Probing Cell Redox State and Glutathione-Modulating Factors Using a Monochlorobimane-Based Microplate Assay. Antioxidants 2022, 11, 391. [Google Scholar] [CrossRef]
- Lifanova, A.; Khaertdinov, N.; Sitdikova, G. Interplay Between Hydrogen Sulfide and Adrenergic and Muscarinic Receptors in the Mouse Atrium. BioNanoScience 2017, 7, 306–308. [Google Scholar] [CrossRef]
- Lifanova, A.; Khaertdinov, N.; Zakharov, A.; Gizzatullin, A.; Sitdikova, G. Role of potassium channels in the negative inotropic effect of hydrogen sulfide in mouse atrium. Genes Cells 2014, 9, 94–98. [Google Scholar]
- DeLeon, E.R.; Stoy, G.F.; Olson, K.R. Passive loss of hydrogen sulfide in biological experiments. Anal. Biochem. 2012, 421, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Rose, P.; Tan, C.-H.; Parkinson, D.B.; Moore, P.K. The Effect of Hydrogen Sulfide Donors on Lipopolysaccharide-Induced Formation of Inflammatory Mediators in Macrophages. Antioxid. Redox Signal. 2010, 12, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Hydrogen Sulfide and Persulfides Oxidation by Biologically Relevant Oxidizing Species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [Green Version]
- Cuevasanta, E.; Möller, M.N.; Alvarez, B. Biological chemistry of hydrogen sulfide and persulfides. Arch. Biochem. Biophys. 2017, 617, 9–25. [Google Scholar] [CrossRef]
- Mishanina, T.V.; Libiad, M.; Banerjee, R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 2015, 11, 457–464. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Shen, Z.; Luo, S.; Guo, W.; Zhu, Y.Z. The Cardioprotective Effects of Hydrogen Sulfide in Heart Diseases: From Molecular Mechanisms to Therapeutic Potential. Oxid. Med. Cell. Longev. 2015, 2015, 925167. [Google Scholar] [CrossRef] [Green Version]
- Kashfi, K.; Olson, K.R. Biology and therapeutic potential of hydrogen sulfide and hydrogen sulfide-releasing chimeras. Biochem. Pharmacol. 2013, 85, 689–703. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Yuan, Z.; Wang, B. Persulfide Prodrugs. In Hydrogen Sulfide; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2022; pp. 293–319. [Google Scholar] [CrossRef]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H.; Kashfi, K. Effects of hydrogen sulfide on mitochondrial function and cellular bioenergetics. Redox Biol. 2021, 38, 101772. [Google Scholar] [CrossRef]
- Olas, B. Chapter Six—Hydrogen Sulfide as a “Double-Faced” Compound: One with Pro- and Antioxidant Effect. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 78, pp. 187–196. [Google Scholar] [CrossRef]
- Wusheng Xiao, J.L. Metabolic Responses to Reductive Stress. Antioxid. Redox Signal. 2020, 32, 1330–1347. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Ye, T.; Li, Y.; Lin, Y.; Liu, D.; Zhou, H.; Wang, J.; Li, L. A New Ratiometric Fluorescent Probe Based on BODIPY for Highly Selective Detection of Hydrogen Sulfide. Molecules 2022, 27, 7499. [Google Scholar] [CrossRef] [PubMed]
- Lin, V.S.; Lippert, A.R.; Chang, C.J. Cell-trappable fluorescent probes for endogenous hydrogen sulfide signaling and imaging H2O2-dependent H2S production. Proc. Natl. Acad. Sci. USA 2013, 110, 7131–7135. [Google Scholar] [CrossRef] [PubMed]
- Trummer, M.; Galardon, E.; Fischer, A.; Toegel, S.; Mayer, B.; Steiner, G.; Kloesch, B. Characterization of the Inducible and Slow-Releasing Hydrogen Sulfide and Persulfide Donor P*: Insights into Hydrogen Sulfide Signaling. Antioxidants 2021, 10, 1049. [Google Scholar] [CrossRef] [PubMed]
- Star, B.S.; van der Slikke, E.C.; Ransy, C.; Schmitt, A.; Henning, R.H.; Bouillaud, F.; Bouma, H.R. GYY4137-Derived Hydrogen Sulfide Donates Electrons to the Mitochondrial Electron Transport Chain via Sulfide: Quinone Oxidoreductase in Endothelial Cells. Antioxidants 2023, 12, 587. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, L.; Zhou, J.; Xu, Y.; Zhu, W.; Qian, X. A highly selective fluorescent probe for fast detection of hydrogen sulfide in aqueous solution and living cells. Chem. Commun. 2012, 48, 10871–10873. [Google Scholar] [CrossRef]
- Zhao, Y.; Steiger, A.K.; Pluth, M.D. Colorimetric Carbonyl Sulfide (COS)/Hydrogen Sulfide (H2S) Donation from γ-Ketothiocarbamate Donor Motifs. Angew. Chem. Int. Ed. 2018, 57, 13101–13105. [Google Scholar] [CrossRef]
- Lacerda, L.; Smith, R.M.; Opie, L.; Lecour, S. TNFα-induced cytoprotection requires the production of free radicals within mitochondria in C2C12 myotubes. Life Sci. 2006, 79, 2194–2201. [Google Scholar] [CrossRef]
- Zebedin, E.; Mille, M.; Speiser, M.; Zarrabi, T.; Sandtner, W.; Latzenhofer, B.; Todt, H.; Hilber, K. C2C12 skeletal muscle cells adopt cardiac-like sodium current properties in a cardiac cell environment. Am. J. Physiol.-Heart Circ. Physiol. 2007, 292, H439–H450. [Google Scholar] [CrossRef] [Green Version]
- Koh, G.Y.; Klug, M.G.; Soonpaa, M.H.; Field, L.J. Differentiation and long-term survival of C2C12 myoblast grafts in heart. J. Clin. Investig. 1993, 92, 1548–1554. [Google Scholar] [CrossRef] [Green Version]
- Reinecke, H.; Minami, E.; Poppa, V.; Murry, C.E. Evidence for Fusion Between Cardiac and Skeletal Muscle Cells. Circ. Res. 2004, 94, e56–e60. [Google Scholar] [CrossRef] [PubMed]
- Menasché, P. Skeletal muscle satellite cell transplantation. Cardiovasc. Res. 2003, 58, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.-Y.; Chun, Y.-S.; Kim, J.-K.; Lee, J.-O.; Lee, Y.-J.; Ku, S.-K.; Shim, S.-M. Curcumin Ameliorated Oxidative Stress and Inflammation-Related Muscle Disorders in C2C12 Myoblast Cells. Antioxidants 2021, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- Mizugaki, A.; Kato, H.; Takeda, T.; Inoue, Y.; Hasumura, M.; Hasegawa, T.; Murakami, H. Cystine reduces mitochondrial dysfunction in C2C12 myotubes under moderate oxidative stress induced by H2O2. Amino Acids 2022, 54, 1203–1213. [Google Scholar] [CrossRef]
- Majumder, A.; Singh, M.; Behera, J.; Theilen, N.T.; George, A.K.; Tyagi, N.; Metreveli, N.; Tyagi, S.C. Hydrogen sulfide alleviates hyperhomocysteinemia-mediated skeletal muscle atrophy via mitigation of oxidative and endoplasmic reticulum stress injury. Am. J. Physiol.-Cell Physiol. 2018, 315, C609–C622. [Google Scholar] [CrossRef]
- Parsanathan, R.; Jain, S.K. Hydrogen sulfide increases glutathione biosynthesis, and glucose uptake and utilisation in C2C12 mouse myotubes. Free Radic. Res. 2018, 52, 288–303. [Google Scholar] [CrossRef]
- Ardite, E.; Barbera, J.A.; Roca, J.; Fernández-Checa, J.C. Glutathione Depletion Impairs Myogenic Differentiation of Murine Skeletal Muscle C2C12 Cells through Sustained NF-κB Activation. Am. J. Pathol. 2004, 165, 719–728. [Google Scholar] [CrossRef]
- Li, X.; Zhang, S.; Zhang, Y.; Liu, P.; Li, M.; Lu, Y.; Han, J. Myoblast differentiation of C2C12 cell may related with oxidative stress. Intractable Rare Dis. Res. 2021, 10, 173–178. [Google Scholar] [CrossRef]
- Pérez-Torres, I.; Guarner-Lans, V.; Rubio-Ruiz, M.E. Reductive Stress in Inflammation-Associated Diseases and the Pro-Oxidant Effect of Antioxidant Agents. Int. J. Mol. Sci. 2017, 18, 2098. [Google Scholar] [CrossRef] [Green Version]
- Bellezza, I.; Riuzzi, F.; Chiappalupi, S.; Arcuri, C.; Giambanco, I.; Sorci, G.; Donato, R. Reductive stress in striated muscle cells. Cell. Mol. Life Sci. 2020, 77, 3547–3565. [Google Scholar] [CrossRef]
- Ma, W.-X.; Li, C.-Y.; Tao, R.; Wang, X.-P.; Yan, L.-J. Reductive Stress-Induced Mitochondrial Dysfunction and Cardiomyopathy. Oxidative Med. Cell. Longev. 2020, 2020, 5136957. [Google Scholar] [CrossRef]
- Yakovleva, O.V.; Ziganshina, A.R.; Dmitrieva, S.A.; Arslanova, A.N.; Yakovlev, A.V.; Minibayeva, F.V.; Khaertdinov, N.N.; Ziyatdinova, G.K.; Giniatullin, R.A.; Sitdikova, G.F. Hydrogen Sulfide Ameliorates Developmental Impairments of Rat Offspring with Prenatal Hyperhomocysteinemia. Oxidative Med. Cell. Longev. 2018, 2018, 2746873. [Google Scholar] [CrossRef] [Green Version]
- Yakovleva, O.; Bogatova, K.; Mukhtarova, R.; Yakovlev, A.; Shakhmatova, V.; Gerasimova, E.; Ziyatdinova, G.; Hermann, A.; Sitdikova, G. Hydrogen Sulfide Alleviates Anxiety, Motor, and Cognitive Dysfunctions in Rats with Maternal Hyperhomocysteinemia via Mitigation of Oxidative Stress. Biomolecules 2020, 10, 995. [Google Scholar] [CrossRef]
- Yakovleva, O.V.; Bogatova, K.S.; Skripnikova, V.V.; Sitdikova, G.F. Effects of Moderate Chronic Stressing of Female Rats Before and During Pregnancy of Sensorimotor Development, Anxiety Levels, and Cognitive Functions in Their Offspring. Neurosci. Behav. Physiol. 2022, 52, 251–261. [Google Scholar] [CrossRef]
- Geng, B.; Chang, L.; Pan, C.; Qi, Y.; Zhao, J.; Pang, Y.; Du, J.; Tang, C. Endogenous hydrogen sulfide regulation of myocardial injury induced by isoproterenol. Biochem. Biophys. Res. Commun. 2004, 318, 756–763. [Google Scholar] [CrossRef] [PubMed]
- Elsey, D.J.; Fowkes, R.C.; Baxter, G.F. Regulation of cardiovascular cell function by hydrogen sulfide (H2S). Cell Biochem. Funct. 2010, 28, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Marino, A.; Piano, I.; Brancaleone, V.; Tomita, K.; Di Cesare Mannelli, L.; Martelli, A.; Citi, V.; Breschi, M.C.; Levi, R.; et al. The novel H2S-donor 4-carboxyphenyl isothiocyanate promotes cardioprotective effects against ischemia/reperfusion injury through activation of mitoKATP channels and reduction of oxidative stress. Pharmacol. Res. 2016, 113, 290–299. [Google Scholar] [CrossRef]
- Geng, B.; Yang, J.; Qi, Y.; Zhao, J.; Pang, Y.; Du, J.; Tang, C. H2S generated by heart in rat and its effects on cardiac function. Biochem. Biophys. Res. Commun. 2004, 313, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Cerra, M.C.; Imbrogno, S. Phospholamban and cardiac function: A comparative perspective in vertebrates. Acta Physiol. 2012, 205, 9–25. [Google Scholar] [CrossRef]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen Sulfide and Cell Signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef] [Green Version]
- Furne, J.; Saeed, A.; Levitt, M.D. Whole tissue hydrogen sulfide concentrations are orders of magnitude lower than presently accepted values. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2008, 295, R1479–R1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhmadishina, R.A.; Kuznetsova, E.V.; Sadrieva, G.R.; Sabirzyanova, L.R.; Nizamov, I.S.; Akhmedova, G.R.; Nizamov, I.D.; Abdullin, T.I. Glutathione salts of O,O-diorganyl dithiophosphoric acids: Synthesis and study as redox modulating and antiproliferative compounds. Peptides 2018, 99, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Witt, R.; Weigand, A.; Boos, A.M.; Cai, A.; Dippold, D.; Boccaccini, A.R.; Schubert, D.W.; Hardt, M.; Lange, C.; Arkudas, A.; et al. Mesenchymal stem cells and myoblast differentiation under HGF and IGF-1 stimulation for 3D skeletal muscle tissue engineering. BMC Cell Biol. 2017, 18, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharov, A.V. Elph: An open-source program for acquisition control and analysis of electrophysiological signals. Uchenye Zap. Kazan. Univ. Seriya Estestv. Nauk. 2019, 161, 245–254. [Google Scholar] [CrossRef] [Green Version]
- Abramochkin, D.V.; Haertdinov, N.N.; Porokhnya, M.V.; Zefirov, A.L.; Sitdikova, G.F. Carbon monoxide affects electrical and contractile activity of rat myocardium. J. Biomed. Sci. 2011, 18, 40. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishkaeva, R.A.; Khaertdinov, N.N.; Yakovlev, A.V.; Esmeteva, M.V.; Salakhieva, D.V.; Nizamov, I.S.; Sitdikova, G.F.; Abdullin, T.I. Characterization of Glutathione Dithiophosphates as Long-Acting H2S Donors. Int. J. Mol. Sci. 2023, 24, 11063. https://doi.org/10.3390/ijms241311063
Ishkaeva RA, Khaertdinov NN, Yakovlev AV, Esmeteva MV, Salakhieva DV, Nizamov IS, Sitdikova GF, Abdullin TI. Characterization of Glutathione Dithiophosphates as Long-Acting H2S Donors. International Journal of Molecular Sciences. 2023; 24(13):11063. https://doi.org/10.3390/ijms241311063
Chicago/Turabian StyleIshkaeva, Rezeda A., Nail N. Khaertdinov, Aleksey V. Yakovlev, Marina V. Esmeteva, Diana V. Salakhieva, Ilyas S. Nizamov, Guzel F. Sitdikova, and Timur I. Abdullin. 2023. "Characterization of Glutathione Dithiophosphates as Long-Acting H2S Donors" International Journal of Molecular Sciences 24, no. 13: 11063. https://doi.org/10.3390/ijms241311063
APA StyleIshkaeva, R. A., Khaertdinov, N. N., Yakovlev, A. V., Esmeteva, M. V., Salakhieva, D. V., Nizamov, I. S., Sitdikova, G. F., & Abdullin, T. I. (2023). Characterization of Glutathione Dithiophosphates as Long-Acting H2S Donors. International Journal of Molecular Sciences, 24(13), 11063. https://doi.org/10.3390/ijms241311063