
Interplay between the RNA Chaperone Hfq, Small RNAs and Transcriptional Regulator OmpR Modulates Iron Homeostasis in the Enteropathogen Yersinia enterocolitica

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
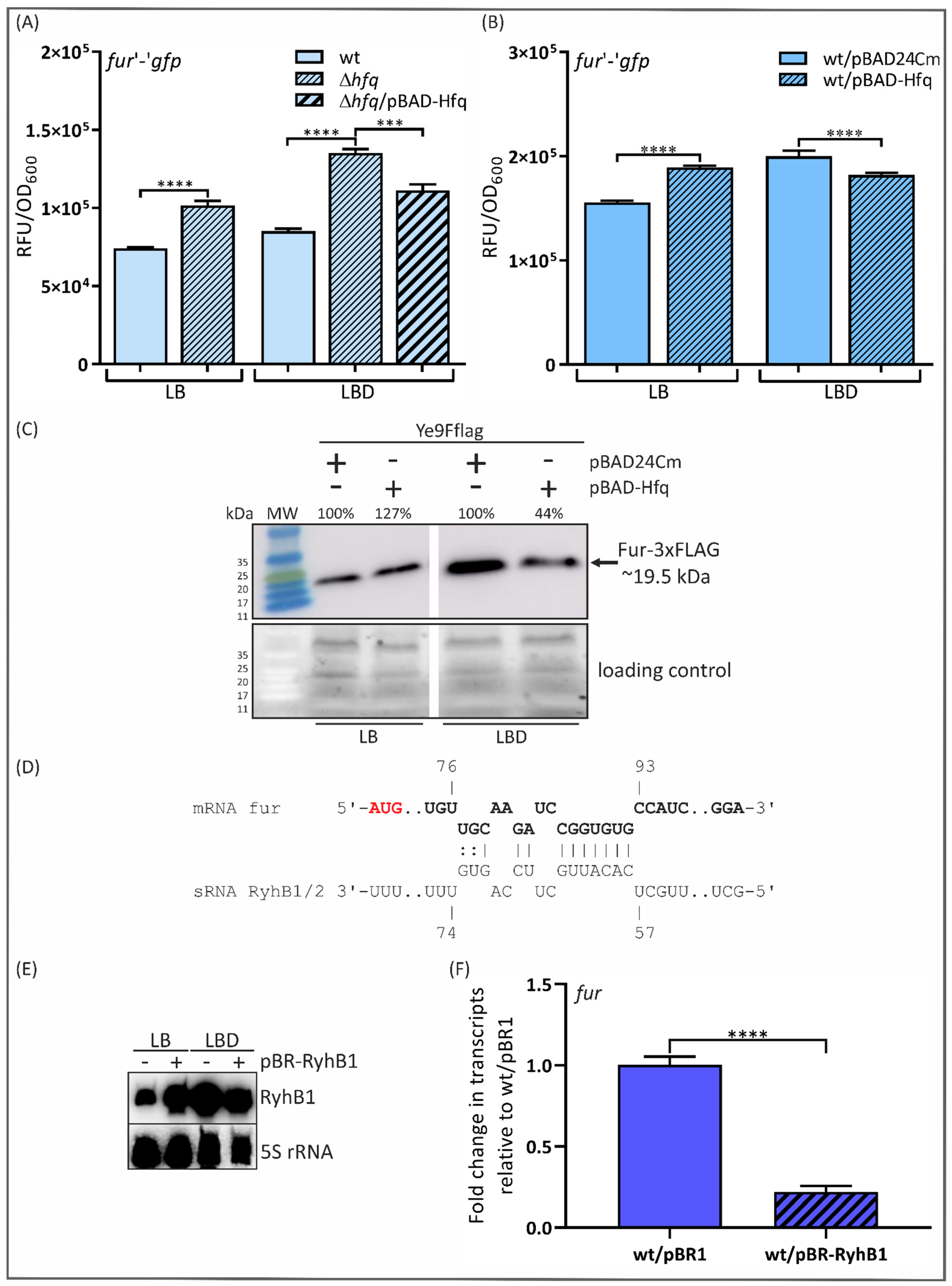
2.1. The RNA Chaperone Hfq and sRNA RyhB1 Impact Fur Expression in Y. enterocolitica 2/O:9, Strain Ye9N
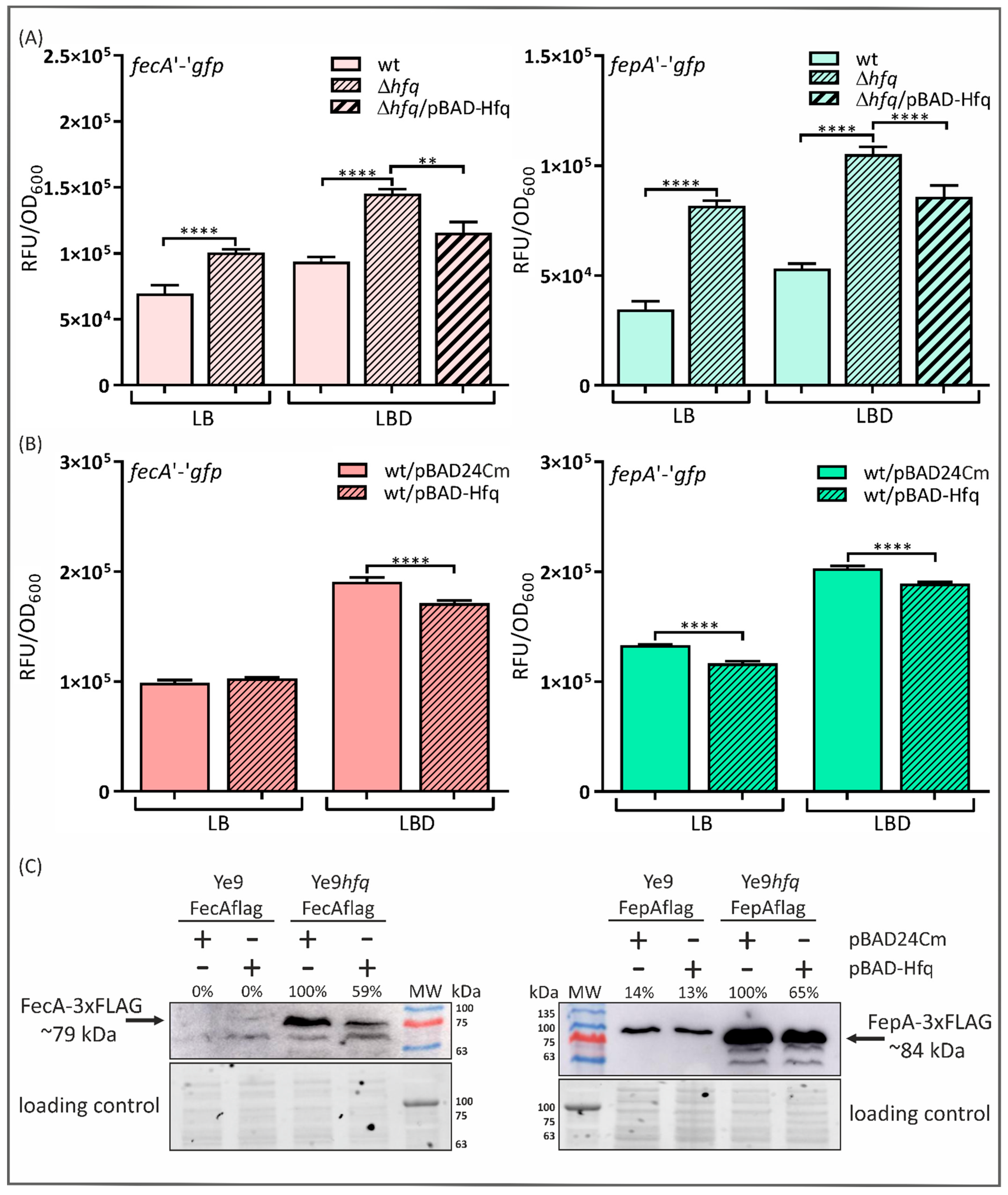
2.2. The Expression of fecA and fepA Is Silenced by Hfq
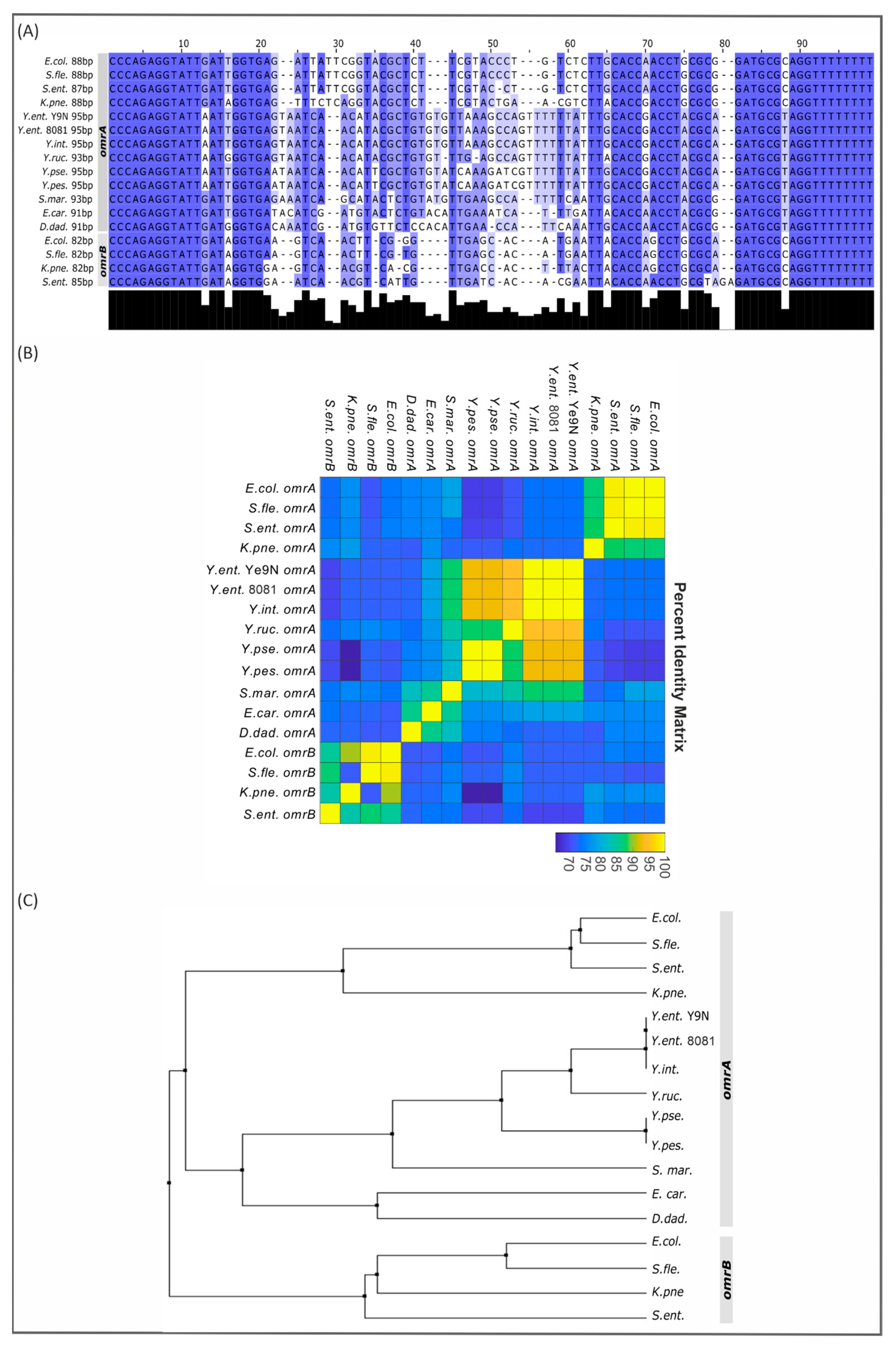
2.3. Partial Conservation of the omrA/B Locus in Gammaproteobacteria
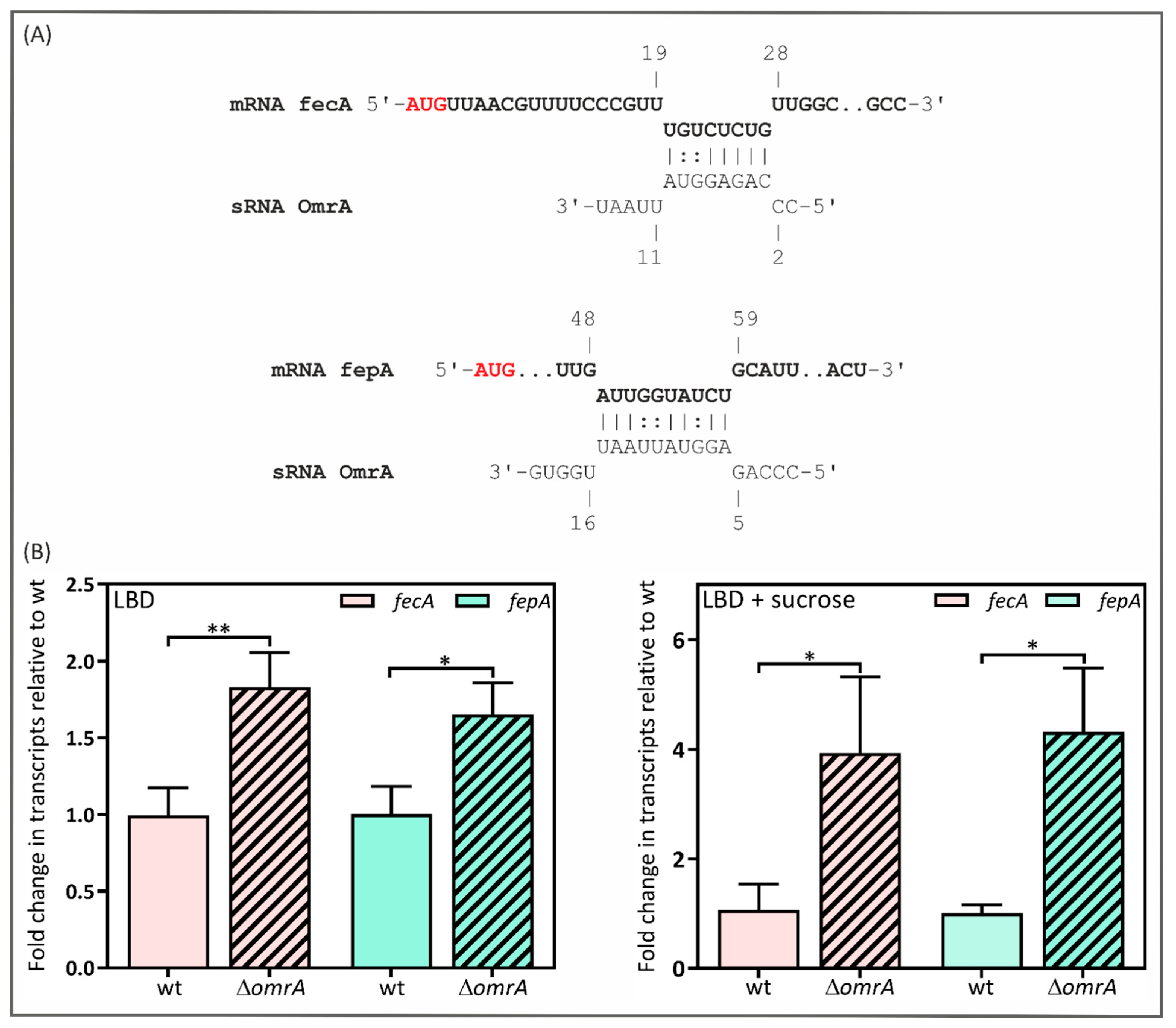
2.4. Y. enterocolitica’s OmrA sRNA Decreases fecA and fepA Levels
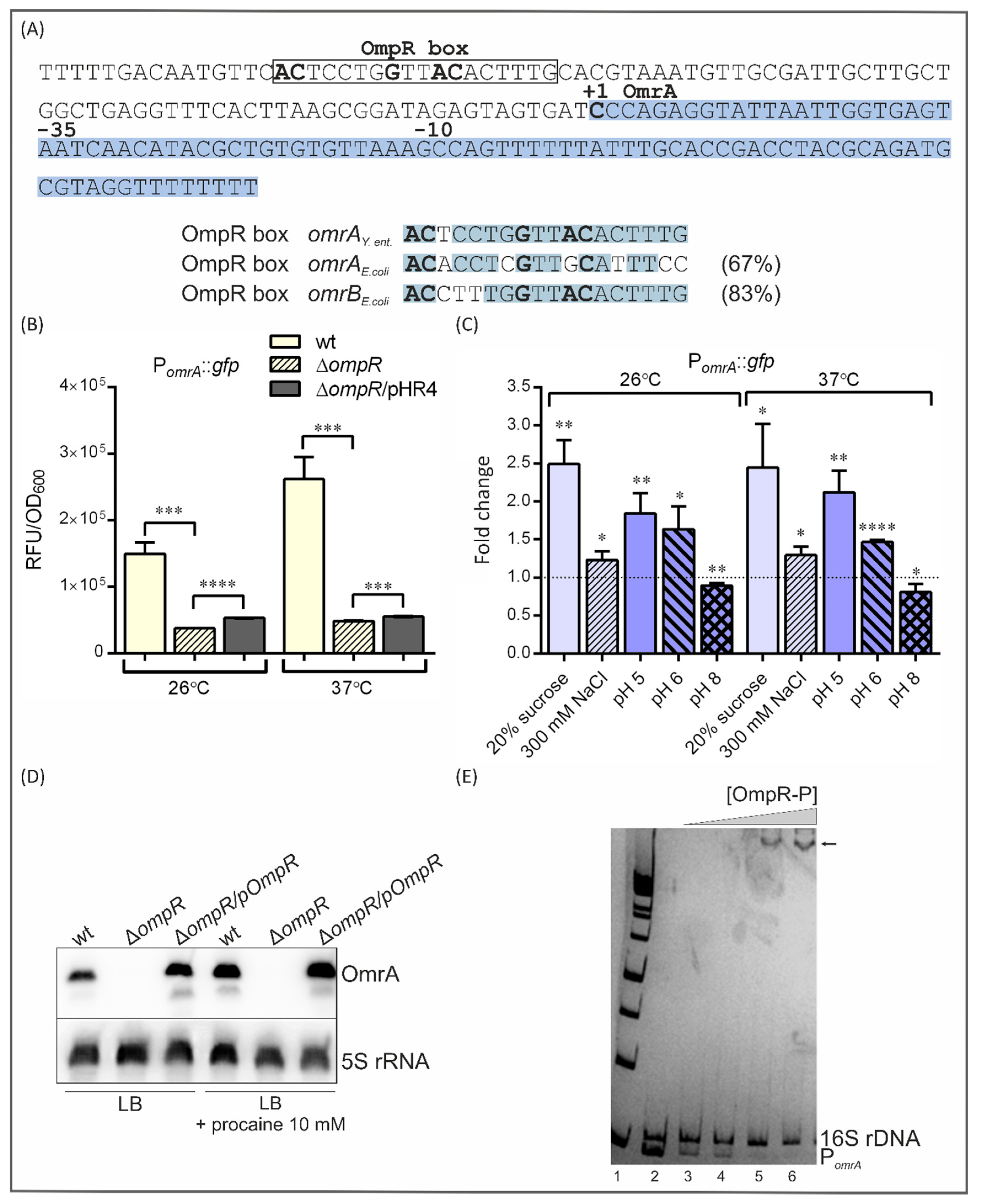
2.5. Influence of OmpR and Environmental Conditions on omrA Promoter Function
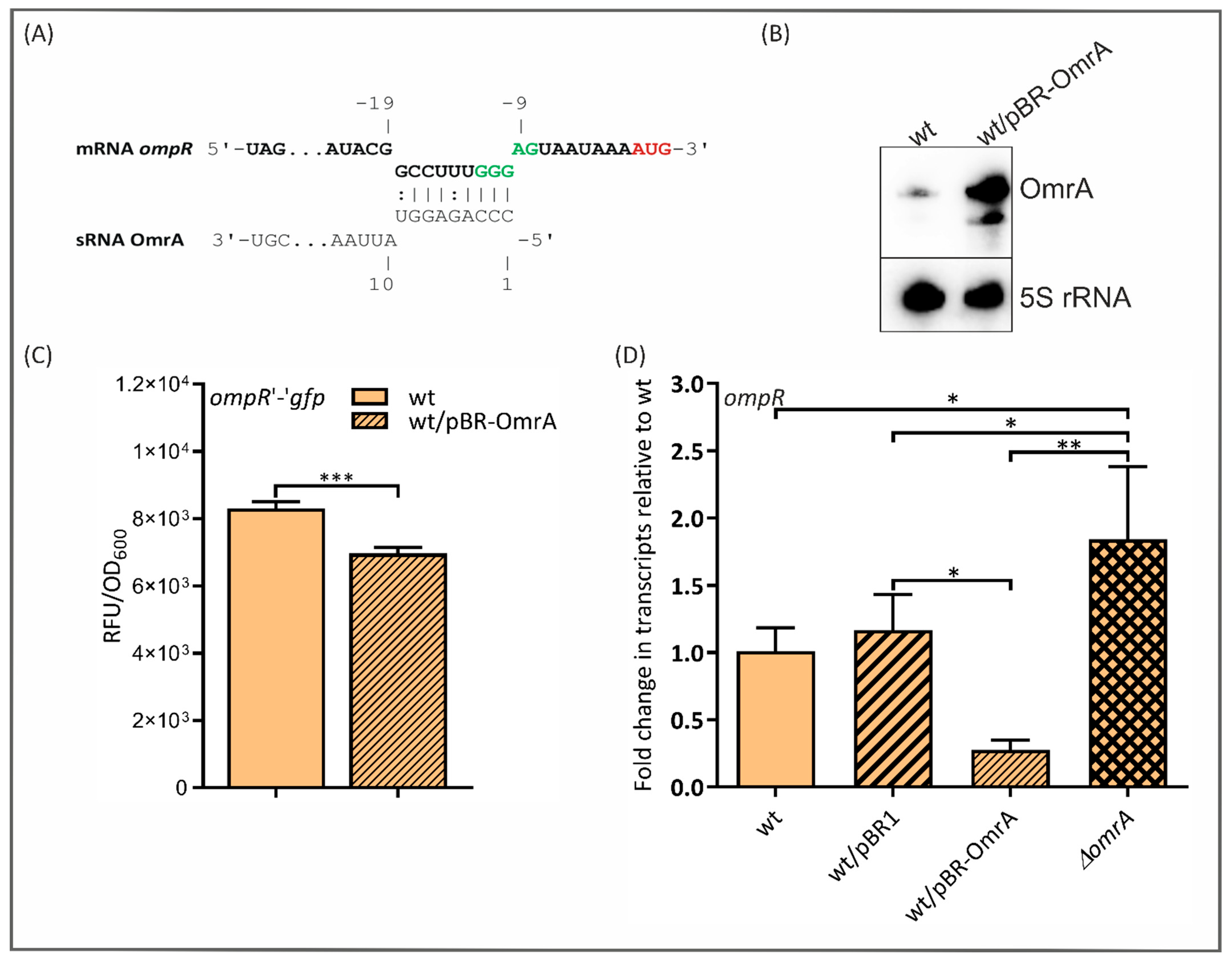
2.6. OmrA Regulates ompR Expression Post-Transcriptionally
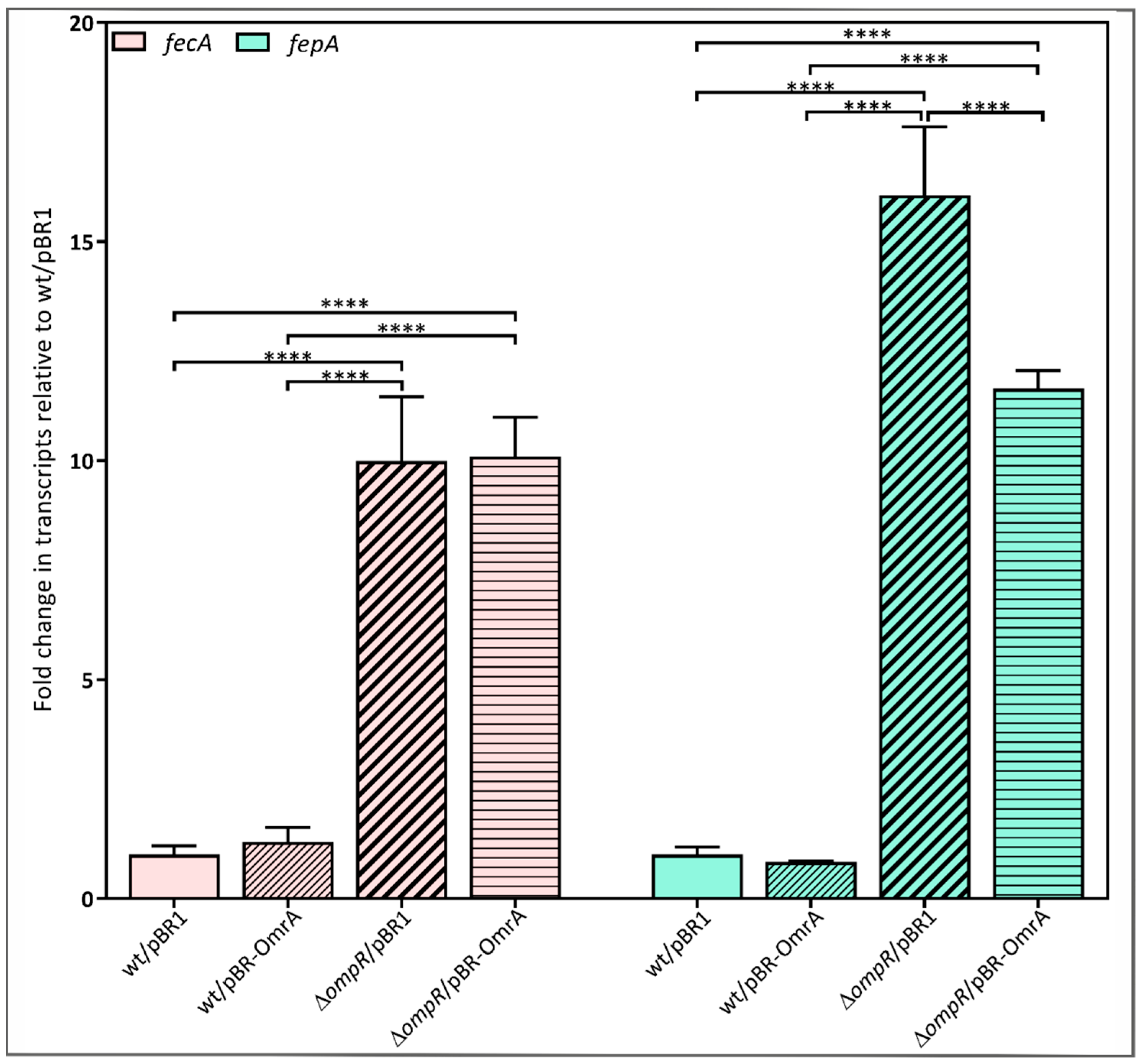
2.7. Requirement for OmpR in the OmrA-Dependent Regulation of fecA and fepA Expression
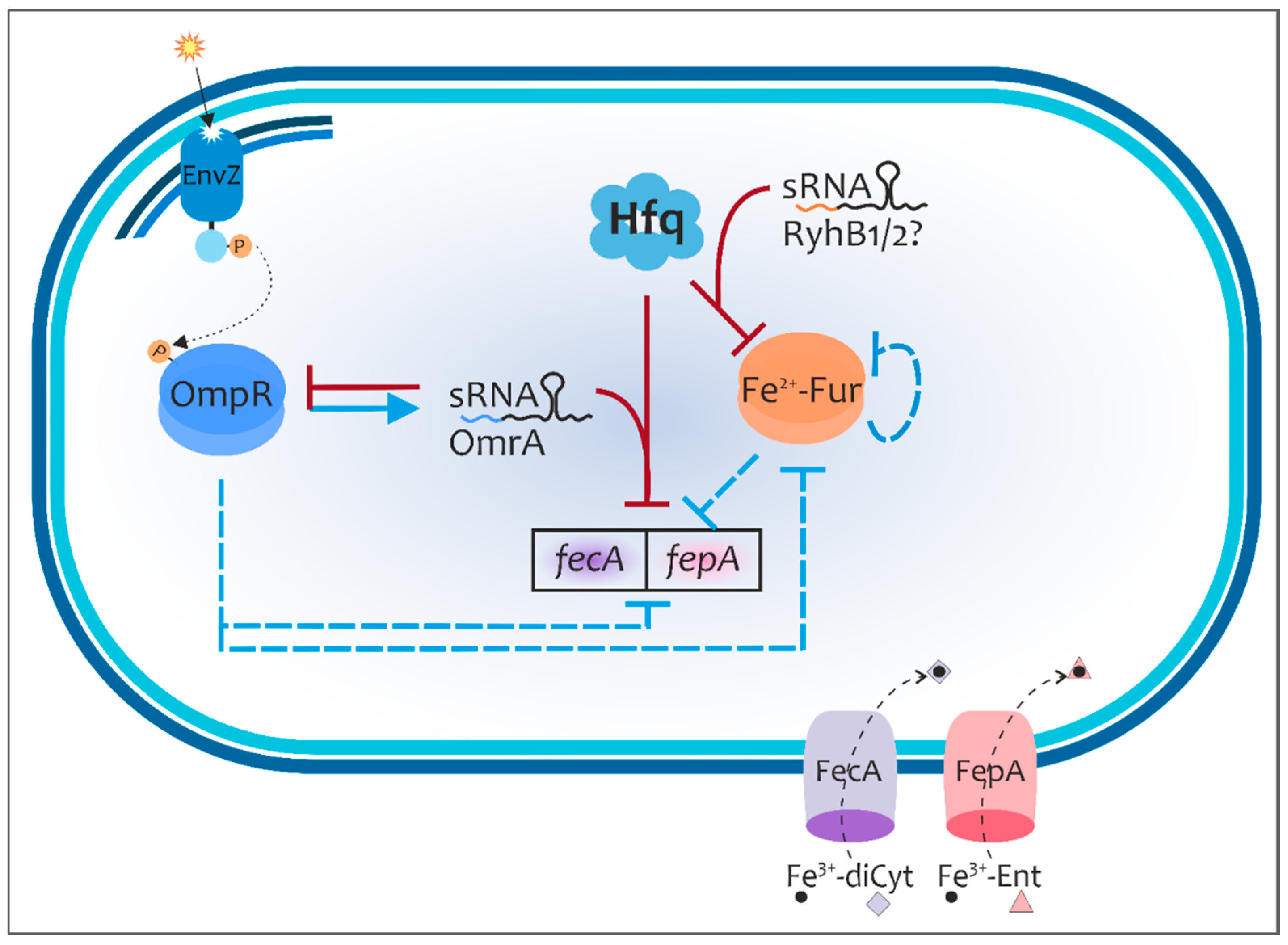
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, Media and Growth Conditions
4.2. Molecular Biology Techniques
4.3. Generation of Null hfq and omrA Mutants in Y. enterocolitica Strains
4.4. Construction of Plasmid-Borne gfp Translational Fusions with fepA and fecA
4.5. Construction of Strains Expressing FecA and FepA Carrying a 3×FLAG Epitope
4.6. Western Blotting
4.7. Construction of a PomrA::gfp Transcriptional Fusion in Plasmid pPROBE-TT’
4.8. Measurement of GFP Fluorescence
4.9. Generation of a Plasmid Overexpressing ryhB1 and omrA from a Foreign Promoter
4.10. RNA Isolation and Northern Blot Analysis
4.11. RT-qPCR
4.12. Electrophoretic Mobility Shift Assay (EMSA)
4.13. Bioinformatic and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrews, S.; Norton, I.; Salunkhe, A.S.; Goodluck, H.; Aly, W.S.; Mourad-Agha, H.; Cornelis, P. Control of iron metabolism in bacteria. Metal Ions Life Sci. 2013, 12, 203–239. [Google Scholar] [CrossRef]
- Frawley, E.R.; Fang, F.C. The ins and outs of bacterial iron metabolism. Mol. Microbiol. 2014, 93, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef]
- Hantke, K. Iron and metal regulation in bacteria. Curr. Opin. Microbiol. 2001, 4, 172–177. [Google Scholar] [CrossRef]
- Andrews, S.C.; Robinson, A.K.; Rodriguez-Quinones, F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003, 27, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Braun, V.; Hantke, K. Recent insights into iron import by bacteria. Curr. Opin. Chem. Biol. 2011, 15, 328–334. [Google Scholar] [CrossRef]
- Finkelstein, R.A.; Sciortino, C.V.; McIntosh, M.A. Role of iron in microbe-host interactions. Rev. Infect. Dis. 1983, 5 (Suppl. S4), S759–S777. [Google Scholar] [CrossRef]
- Sheldon, J.R.; Laakso, H.A.; Heinrichs, D.E. Iron Acquisition Strategies of Bacterial Pathogens. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Noinaj, N.; Guillier, M.; Barnard, T.J.; Buchanan, S.K. TonB-dependent transporters: Regulation, structure, and function. Annu. Rev. Microbiol. 2010, 64, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Doan, B.; Schneider, T.D.; Storz, G. OxyR and SoxRS regulation of fur. J. Bacteriol. 1999, 181, 4639–4643. [Google Scholar] [CrossRef] [Green Version]
- Vecerek, B.; Moll, I.; Blasi, U. Control of Fur synthesis by the non-coding RNA RyhB and iron-responsive decoding. EMBO J. 2007, 26, 965–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottone, E.J. Yersinia enterocolitica: A panoramic view of a charismatic microorganism. CRC Crit. Rev. Microbiol. 1977, 5, 211–241. [Google Scholar] [CrossRef] [PubMed]
- Bottone, E.J. Yersinia enterocolitica: Revisitation of an Enduring Human Pathogen. Clin. Microbiol. Newsl. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- ECDC. Annual Epidemiological Report on Communicable Diseases, European Centre for Disease Prevention and Control. 2020. Available online: http://www.ecdc.europa.eu (accessed on 9 September 2022).
- Pelludat, C.; Rakin, A.; Jacobi, C.A.; Schubert, S.; Heesemann, J. The yersiniabactin biosynthetic gene cluster of Yersinia enterocolitica: Organization and siderophore-dependent regulation. J. Bacteriol. 1998, 180, 538–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caza, M.; Kronstad, J.W. Shared and distinct mechanisms of iron acquisition by bacterial and fungal pathogens of humans. Front. Cell. Infect. Microbiol. 2013, 19, 80. [Google Scholar] [CrossRef]
- Jaworska, K.; Nieckarz, M.; Ludwiczak, M.; Raczkowska, A.; Brzostek, K. OmpR-Mediated Transcriptional Regulation and Function of Two Heme Receptor Proteins of Yersinia enterocolitica Bio-Serotype 2/O:9. Front. Cell. Infect. Microbiol. 2018, 8, 333. [Google Scholar] [CrossRef]
- Carniel, E.; Guilvout, I.; Prentice, M. Characterization of a large chromosomal “high-pathogenicity island” in biotype 1B Yersinia enterocolitica. J. Bacteriol. 1996, 178, 6743–6751. [Google Scholar] [CrossRef] [Green Version]
- Bäumler, A.; Koebnik, R.; Stojiljkovic, I.; Heesemann, J.; Bottone, V.; Hantke, K. Survey on Newly Characterized Iron Uptake Systems of Yersinia enterocolitica. Zent. Bakteriol. 1993, 278, 416–424. [Google Scholar] [CrossRef]
- Straley, S.C.; Perry, R.D. Environmental modulation of gene expression and pathogenesis in Yersinia. Trends Microbiol. 1995, 3, 310–317. [Google Scholar] [CrossRef]
- Staggs, T.M.; Perry, R.D. Fur regulation in Yersinia species. Mol. Microbiol. 1992, 6, 2507–2516. [Google Scholar] [CrossRef]
- Jaworska, K.; Ludwiczak, M.; Murawska, E.; Raczkowska, A.; Brzostek, K. The regulator OmpR in Yersinia enterocolitica participates in iron homeostasis by modulating fur level and affecting the expression of genes involved in iron uptake. Int. J. Mol. Sci. 2021, 22, 1475. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.S.; Storz, G. Regulatory RNAs in bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guillier, M.; Gottesman, S. Remodelling of the Escherichia coli outer membrane by two small regulatory RNAs. Mol. Microbiol. 2006, 59, 231–247. [Google Scholar] [CrossRef]
- Guillier, M.; Gottesman, S.; Storz, G. Modulating the outer membrane with small RNAs. Genes Dev. 2006, 20, 2338–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görke, B.; Vogel, J. Noncoding RNA control of the making and breaking of sugars. Genes Dev. 2008, 22, 2914–2925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romby, P.; Vandenesch, F.; Wagner, E.G. The role of RNAs in the regulation of virulence-gene expression. Curr. Opin. Microbiol. 2006, 9, 229–236. [Google Scholar] [CrossRef]
- Mika, F.; Hengge, R. Small regulatory RNAs in the control of motility and biofilm formation in E. coli and Salmonella. Int. J. Mol. Sci. 2013, 26, 4560–4579. [Google Scholar] [CrossRef] [Green Version]
- Masse, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef]
- Koo, J.T.; Alleyne, T.M.; Schiano, C.A.; Jafari, N.; Lathem, W.W. Global discovery of small RNAs in Yersinia pseudotuberculosis identifies Yersinia-specific small, noncoding RNAs required for virulence. Proc. Natl. Acad. Sci. USA 2011, 108, E709–E717. [Google Scholar] [CrossRef]
- Schiano, C.A.; Lathem, W.W. Post-transcriptional regulation of gene expression in Yersinia species. Front. Cell. Infect. Microbiol. 2012, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Chavarría, L.C.; Vadyvaloo, V. Yersinia pestis and Yersinia pseudotuberculosis infection: A regulatory RNA perspective. Front. Microbiol. 2015, 6, 956. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Su, S.; Meng, X.; Ji, X.; Qu, Y.; Liu, Z.; Wang, X.; Cui, Y.; Deng, Z.; Zhou, D.; et al. Determination of sRNA expressions by RNA-seq in Yersinia pestis grown in vitro and during infection. PLoS ONE 2013, 8, e74495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmühl, C.; Beckstette, M.; Heroven, A.K.; Bunk, B.; Spröer, C.; McNally, A.; Overmann, J.; Dersch, P. Comparative transcriptomic profiling of Yersinia enterocolitica O:3 and O:8 reveals major expression differences of fitness- and virulence-relevant genes indicating ecological separation. Msystems 2019, 23, e00239-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massé, E.; Escorcia, F.E.; Gottesman, S. Coupled degradation of a small regulatory RNA and its mRNA targets in Escherichia coli. Genes Dev. 2003, 17, 2374–2383. [Google Scholar] [CrossRef] [Green Version]
- Moll, I.; Leitsch, D.; Steinhauser, T.; Bläsi, U. RNA chaperone activity of the Sm-like Hfq protein. EMBO Rep. 2003, 4, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Aiba, H. Mechanism of RNA silencing by Hfq-binding small RNAs. Curr. Opin. Microbiol. 2007, 10, 13413–13419. [Google Scholar] [CrossRef]
- Franze de Fernandez, M.T.; Eoyang, L.; August, J.T. Factor fraction required for the synthesis of bacteriophage Qbeta-RNA. Nature 1968, 219, 588–590. [Google Scholar] [CrossRef]
- Vogel, J.; Luisi, B.F. Hfq and its constellation of RNA. Nat. Rev. Microbiol. 2011, 9, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhulin, I.; Wartell, R.M. Predicted structure and phyletic distribution of the RNA-binding protein Hfq. Nucleic Acids Res. 2002, 30, 3662–3671. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.; Vogel, J. The role of Hfq in bacterial pathogens. Curr. Opin. Microbiol. 2010, 13, 24–33. [Google Scholar] [CrossRef]
- Geng, J.; Song, Y.; Yang, L.; Feng, Y.; Qiu, Y.; Li, G.; Guo, J.; Bi, Y.; Qu, Y.; Wang, W.; et al. Involvement of the post-transcriptional regulator Hfq in Yersinia pestis virulence. PLoS ONE 2009, 4, e6213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiano, C.A.; Bellows, L.E.; Lathem, W.W. The small RNA chaperone Hfq is required for the virulence of Yersinia pseudotuberculosis. Infect. Immun. 2010, 78, 2034–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakoschke, T.; Kakoschke, S.; Magistro, G.; Schubert, S.; Borath, M.; Heesemann, J.; Rossier, O. The RNA chaperone Hfq impacts growth, metabolism and production of virulence factors in Yersinia enterocolitica. PLoS ONE 2014, 9, e86113. [Google Scholar] [CrossRef] [Green Version]
- Kakoschke, T.K.; Kakoschke, S.C.; Zeuzem, C.; Bouabe, H.; Adler, K.; Heesemann, J.; Rossier, O. The RNA chaperone Hfq is essential for virulence and modulates the expression of four adhesins in Yersinia enterocolitica. Sci. Rep. 2016, 6, 29275. [Google Scholar] [CrossRef] [Green Version]
- Brantl, S. Bacterial chromosome-encoded small regulatory RNAs. Future Microbiol. 2009, 4, 85–103. [Google Scholar] [CrossRef]
- Wilderman, P.J.; Sowa, N.A.; FitzGerald, D.J.; FitzGerald, P.C.; Gottesman, S.; Ochsner, U.A.; Vasil, M.L. Identification of tandem duplicate regulatory small RNAs in Pseudomonas aeruginosa involved in iron homeostasis. Proc. Natl. Acad. Sci. USA 2004, 101, 9792–9797. [Google Scholar] [CrossRef] [PubMed]
- Mellin, J.R.; Goswami, S.; Grogan, S.; Tjaden, B.; Genco, C.A. A novel Fur- and iron-regulated small RNA, NrrF, is required for indirect Fur-mediated regulation of the sdhA and sdhC genes in Neisseria meningitides. J. Bacteriol. 2007, 189, 3686–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaballa, A.; Antelmann, H.; Aguilar, C.; Khakh, S.K.; Song, K.B.; Smaldone, G.T.; Helmann, J.D. The Bacillus subtilis iron-sparing response is mediated by a Fur-regulated small RNA and three small, basic proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 11927–11932. [Google Scholar] [CrossRef]
- Mey, A.R.; Craig, S.A.; Payne, S.M. Characterization of Vibrio cholerae RyhB: The RyhB regulon and role of RyhB in biofilm formation. Infect. Immun. 2005, 73, 5706–5719. [Google Scholar] [CrossRef] [Green Version]
- Davis, B.M.; Quinones, M.; Pratt, J.; Ding, Y.; Waldor, M.K. Characterization of the small untranslated RNA RyhB and its regulon in Vibrio cholera. J. Bacteriol. 2005, 187, 4005–4014. [Google Scholar] [CrossRef] [Green Version]
- Oglesby, A.G.; Murphy, E.R.; Iyer, V.R.; Payne, S.M. Fur regulates acid resistance in Shigella flexneri via RyhB and ydeP. Mol. Microbiol. 2005, 58, 1354–1367. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.R.; Payne, S.M. RyhB, an iron-responsive small RNA molecule, regulates Shigella dysenteriae virulence. Infect. Immun. 2007, 75, 470–3477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argaman, L.; Hershberg, R.; Vogel, J.; Bejerano, G.; Wagner, E.G.; Margalit, H.; Altuvia, S. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 2001, 12, 941–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassarman, K.M.; Repoila, F.; Rosenow, C.; Storz, G.; Gottesman, S. Identification of novel small RNAs using comparative genomics and microarrays. Genes Dev. 2001, 15, 1637–1651. [Google Scholar] [CrossRef] [Green Version]
- Guillier, M.; Gottesman, S. The 5′ end of two redundant sRNAs is involved in the regulation of multiple targets, including their own regulator. Nucleic Acids Res. 2008, 36, 6781–6794. [Google Scholar] [CrossRef] [Green Version]
- Holmqvist, E.; Reimegård, J.; Sterk, M.; Grantcharova, N.; Römling, U.; Wagner, E.G. Two antisense RNAs target the transcriptional regulator CsgD to inhibit curli synthesis. EMBO J. 2010, 29, 1840–1850. [Google Scholar] [CrossRef] [Green Version]
- De Lay, N.; Gottesman, S. A complex network of small non-coding RNAs regulate motility in Escherichia coli. Mol. Microbiol. 2012, 86, 524–538. [Google Scholar] [CrossRef] [Green Version]
- Romilly, C.; Hoekzema, M.; Holmqvist, E.; Wagner, E.G.H. Small RNAs OmrA and OmrB promote class III flagellar gene expression by inhibiting the synthesis of anti-Sigma factor FlgM. RNA Biol. 2020, 17, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Brosse, A.; Korobeinikova, A.; Gottesman, S.; Guillier, M. Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system. Nucleic Acids Res. 2016, 44, 9650–9666. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.J.; Inouye, M. EnvZ-OmpR interaction and osmoregulation in Escherichia coli. J. Biol. Chem. 2002, 277, 24155–24161. [Google Scholar] [CrossRef] [Green Version]
- Stincone, A.; Daudi, N.; Rahman, A.S.; Antczak, P.; Henderson, I.; Cole, J.; Johnson, M.D.; Lund, P.; Falciani, F. A systems biology approach sheds new light on Escherichia coli acid resistance. Nucleic Acids Res. 2011, 39, 7512–7528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, S.; Kenney, L.J. A New Role of OmpR in Acid and Osmotic Stress in Salmonella and E. coli. Front. Microbiol. 2018, 9, 2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slauch, J.M.; Silhavy, T.J. Genetic analysis of the switch that controls porin gene expression in Escherichia coli K-12. J. Mol. Biol. 1989, 210, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Park, C. Modulation of flagellar expression in Escherichia coli by acetyl phosphate and the osmoregulator OmpR. J. Bacteriol. 1995, 177, 4696–4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Nagura, R.; Tanabe, H.; Fujita, N.; Ishihama, A.; Utsumi, R. Negative regulation of the bolA1p of Escherichia coli K-12 by the transcription factor OmpR for osmolarity response genes. FEMS Microbiol. Lett. 2000, 186, 257–262. [Google Scholar] [CrossRef]
- Bernardini, M.L.; Fontaine, A.; Sansonetti, P.J. The two-component regulatory system ompR-envZ controls the virulence of Shigella flexneri. J. Bacteriol. 1990, 172, 6274–6281. [Google Scholar] [CrossRef] [Green Version]
- Perkins, T.T.; Davies, M.R.; Klemm, E.J.; Rowley, G.; Wileman, T.; James, K.; Keane, T.; Maskell, D.; Hinton, J.C.D.; Dougan, G.; et al. ChIP-seq and transcriptome analysis of the OmpR regulon of Salmonella enterica serovars Typhi and Typhimurium reveals accessory genes implicated in host colonization. Mol. Microbiol. 2013, 87, 526–538. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Lu, P.; Wang, Y.; Ding, L.; Atkinson, S.; Chen, S. OmpR positively regulates urease expression to enhance acid survival of Yersinia pseudotuberculosis. Microbiology 2009, 155, 2522–2531. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Zhang, Y.; Han, Y.; Yang, L.; Liu, X.; Guo, Z.; Tan, Y.; Huang, X.; Zhou, D.; Yang, R. Phenotypic and transcriptional analysis of the osmotic regulator OmpR in Yersinia pestis. BMC Microbiol. 2011, 11, 39. [Google Scholar] [CrossRef] [Green Version]
- Brzostek, K.; Skorek, K.; Raczkowska, A. OmpR, a central integrator of several cellular responses in Yersinia enterocolitica. Adv. Exp. Med. Biol. 2012, 954, 325–334. [Google Scholar] [CrossRef]
- Skorek, K.; Raczkowska, A.; Dudek, B.; Miętka, K.; Guz-Regner, K.; Pawlak, A.; Klausa, E.; Bugla-Płoskońska, G.; Brzostek, K. Regulatory protein OmpR influences the serum resistance of Yersinia enterocolitica O:9 by modifying the structure of the outer membrane. PLoS ONE 2013, 8, e79525. [Google Scholar] [CrossRef]
- Raczkowska, A.; Trzos, J.; Lewandowska, O.; Nieckarz, M.; Brzostek, K. Expression of the AcrAB components of the AcrAB-TolC multidrug efflux pump of Yersinia enterocolitica is subject to dual regulation by OmpR. PLoS ONE 2015, 10, e0124248. [Google Scholar] [CrossRef] [PubMed]
- Nieckarz, M.; Raczkowska, A.; Debski, J.; Kistowski, M.; Dadlez, M.; Heesemann, J.; Rossier, O.; Brzostek, K. Impact of OmpR on the membrane proteome of Yersinia enterocolitica in different environments: Repression of major adhesin YadA and heme receptor HemR. Environ. Microbiol. 2016, 18, 997–1021. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, V.; Herrero, M.; Giovannini, F.; Neilands, J.B. Fur (ferric uptake regulation) protein and CAP (catabolite-activator protein) modulate transcription of fur gene in Escherichia coli. Eur. J. Biochem. 1988, 173, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Meng, X.; Su, S.; Liu, Z.; Ji, X.; Zhang, Y.; Zhao, X.; Wang, X.; Yang, R.; Han, Y. Two sRNA RyhB homologs from Yersinia pestis biovar microtus expressed in vivo have differential Hfq-dependent stability. Res. Microbiol. 2012, 163, 413–418. [Google Scholar] [CrossRef]
- Acuña, L.G.; Barros, M.J.; Montt, F.; Peñaloza, D.; Núñez, P.; Valdés, I.; Gil, F.; Fuentes, J.A.; Calderón, I.L. Participation of two sRNA RyhB homologs from the fish pathogen Yersinia ruckeri in bacterial physiology. Microbiol. Res. 2021, 242, 126629. [Google Scholar] [CrossRef] [PubMed]
- Arraiano, C.M.; Andrade, J.M.; Domingues, S.; Guinote, I.B.; Malecki, M.; Matos, R.G.; Moreira, R.N.; Pobre, V.; Reis, F.P.; Saramago, M.; et al. The critical role of RNA processing and degradation in the control of gene expression. FEMS Microbiol. Rev. 2010, 34, 883–923. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview version 2: A multiple sequence alignment and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Harlocker, S.L.; Bergstrom, L.; Inouye, M. Tandem binding of six OmpR proteins to the ompF upstream regulatory sequence of Escherichia coli. J. Biol. Chem. 1995, 270, 26849–26856. [Google Scholar] [CrossRef] [Green Version]
- Pratt, L.A.; Silhavy, T.J. Identification of base pairs important for OmpR-DNA interaction. Mol. Microbiol. 1995, 17, 565–573. [Google Scholar] [CrossRef]
- Kenney, L.J. Structure/function relationships in OmpR and other winged-helix transcription factors. Curr. Opin. Microbiol. 2002, 5, 135–141. [Google Scholar] [CrossRef]
- Brzostek, K.; Raczkowska, A.; Zasada, A. The osmotic regulator OmpR is involved in the response of Yersinia enterocolitica O:9 to environmental stresses and survival within macrophages. FEMS Microbiol. Lett. 2003, 228, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Taylor, R.K.; Hall, M.N.; Silhavy, T.J. Isolation and characterization of mutations altering expression of the major outer membrane porin proteins using the local anaesthetic procaine. J. Mol. Biol. 1983, 166, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Rampersaud, A.; Inouye, M. Procaine, a local anesthetic, signals through the EnvZ receptor to change the DNA binding affinity of the transcriptional activator protein OmpR. J. Bacteriol. 1991, 173, 6882–6888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, B.; Wu, H.S.; Xin, Y.Q.; Zhang, Q.W.; Zhang, Y.Q. Reciprocal regulation between Fur and two RyhB homologs in Yersinia pestis, and roles of RyhBs in biofilm formation. Biomed. Environ. Sci. 2021, 34, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Hussein, R.; Lim, H.N. Disruption of small RNA signaling caused by competition for Hfq. Proc. Natl. Acad. Sci. USA 2011, 108, 1110–1115. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Gottesman, S. Competition among Hfq-binding small RNAs in Escherichia coli. Mol. Microbiol. 2011, 82, 1545–1562. [Google Scholar] [CrossRef] [PubMed]
- Carniel, E. The Yersinia high-pathogenicity island: An iron-uptake island. Microbes Infect. 2001, 3, 561–569. [Google Scholar] [CrossRef]
- Vecerek, B.; Moll, I.; Afonyushkin, T.; Kaberdin, V.; Bläsi, U. Interaction of the RNA chaperone Hfq with mRNAs: Direct and indirect roles of Hfq in iron metabolism of Escherichia coli. Mol. Microbiol. 2003, 50, 897–909. [Google Scholar] [CrossRef]
- Mellin, J.R.; McClure, R.; Lopez, D.; Green, O.; Reinhard, B.; Genco, C. Role of Hfq in iron-dependent and -independent gene regulation in Neisseria meningitidis. Microbiology 2010, 156, 2316–2326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oglesby-Sherrouse, A.G.; Murphy, E.R. Iron-responsive bacterial small RNAs: Variations on a theme. Metallomics 2013, 4, 276–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachterle, J.K.; Zeng, Q.; Sundin, G.W. Three Hfq-dependent small RNAs regulate flagellar motility in the fire blight pathogen Erwinia amylovora. Mol. Microbiol. 2019, 111, 1476–1492. [Google Scholar] [CrossRef]
- Jagodnik, J.; Chiaruttini, C.; Guillier, M. Stem-loop structures within mRNA coding sequences activate translation initiation and mediate control by small regulatory RNAs. Mol. Cell. 2017, 5, 158–170.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottesman, S.; Storz, G. Bacterial small RNA regulators: Versatile roles and rapidly evolving variations. Cold Spring Harb. Perspect. Biol. 2011, 3, a003798. [Google Scholar] [CrossRef] [Green Version]
- Russo, F.D.; Silhavy, T.J. EnvZ controls the concentration of phosphorylated OmpR to mediate osmoregulation of the porin genes. J. Mol. Biol. 1991, 222, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, L.; Lin, L.; Tang, H.; Osei-Adjei, G.; Xu, S.; Zhang, Y.; Huang, X. Reciprocal regulation of OmpR and Hfq and their regulatory actions on the Vi polysaccharide capsular antigen in Salmonella enterica serovar Typhi. Curr. Microbiol. 2018, 6, 773–778. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in Gram-negative bacteria. Bio/Technology 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Herrero, M.; de Lorenzo, V.; Timmis, K.N. Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in Gram-negative bacteria. J. Bacteriol. 1990, 172, 6557–6567. [Google Scholar] [CrossRef] [Green Version]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russel, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Philippe, N.; Alcaraz, J.-P.; Coursange, E.; Geiselmann, J.; Schneider, D. Improvement of pCVD442, a suicide plasmid for gene allele exchange in bacteria. Plasmid 2004, 51, 246–255. [Google Scholar] [CrossRef]
- Engler, C.; Kandzia, R.; Marillonnet, S. A one pot, one step, precision cloning method with high throughput capability. PLoS ONE 2008, 3, e3647. [Google Scholar] [CrossRef] [Green Version]
- Schmidtke, C.; Abendroth, U.; Brock, J.; Serrania, J.; Becker, A.; Bonas, U. Small RNA sX13: A multifaceted regulator of virulence in the plant pathogen Xanthomonas. PLoS Pathog. 2013, 9, e1003626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figurski, D.H.; Helinski, D.R. Replication of an origin-containing derivative of plasmid RK2 dependent on a plasmid function provided in trans. Proc. Natl. Acad. Sci. USA 1979, 76, 1648–1652. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.G.; Leveau, J.H.; Lindow, S.E. Improved gfp and inaZ broad-host-range promoter-probe vectors. Mol. Plant-Microbe Interact. 2000, 11, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, R.; Bujard, H. Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. Nucleic Acids Res. 1997, 25, 1203–1210. [Google Scholar] [CrossRef]
- Chomczynski, P. A reagent for the single-step simultaneous isolation of RNA, DNA and proteins from cell and tissue samples. Biotechniques 1993, 15, 532–537. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Brzostek, K.; Brzostkowska, M.; Bukowska, I.; Karwicka, E.; Raczkowska, A. OmpR negatively regulates expression of invasin in Yersinia enterocolitica. Microbiology 2007, 153, 2416–2425. [Google Scholar] [CrossRef] [Green Version]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaworska, K.; Konarska, J.; Gomza, P.; Rożen, P.; Nieckarz, M.; Krawczyk-Balska, A.; Brzostek, K.; Raczkowska, A. Interplay between the RNA Chaperone Hfq, Small RNAs and Transcriptional Regulator OmpR Modulates Iron Homeostasis in the Enteropathogen Yersinia enterocolitica. Int. J. Mol. Sci. 2023, 24, 11157. https://doi.org/10.3390/ijms241311157
Jaworska K, Konarska J, Gomza P, Rożen P, Nieckarz M, Krawczyk-Balska A, Brzostek K, Raczkowska A. Interplay between the RNA Chaperone Hfq, Small RNAs and Transcriptional Regulator OmpR Modulates Iron Homeostasis in the Enteropathogen Yersinia enterocolitica. International Journal of Molecular Sciences. 2023; 24(13):11157. https://doi.org/10.3390/ijms241311157
Chicago/Turabian StyleJaworska, Karolina, Julia Konarska, Patrycja Gomza, Paula Rożen, Marta Nieckarz, Agata Krawczyk-Balska, Katarzyna Brzostek, and Adrianna Raczkowska. 2023. "Interplay between the RNA Chaperone Hfq, Small RNAs and Transcriptional Regulator OmpR Modulates Iron Homeostasis in the Enteropathogen Yersinia enterocolitica" International Journal of Molecular Sciences 24, no. 13: 11157. https://doi.org/10.3390/ijms241311157
APA StyleJaworska, K., Konarska, J., Gomza, P., Rożen, P., Nieckarz, M., Krawczyk-Balska, A., Brzostek, K., & Raczkowska, A. (2023). Interplay between the RNA Chaperone Hfq, Small RNAs and Transcriptional Regulator OmpR Modulates Iron Homeostasis in the Enteropathogen Yersinia enterocolitica. International Journal of Molecular Sciences, 24(13), 11157. https://doi.org/10.3390/ijms241311157