
Comparison of Four Methods of RNA Extraction and cDNA Synthesis from The Venom of Peruvian Snakes of the Genus Bothrops of Clinical Importance

 , , , , and
, , , , and
Abstract
:1. Introduction
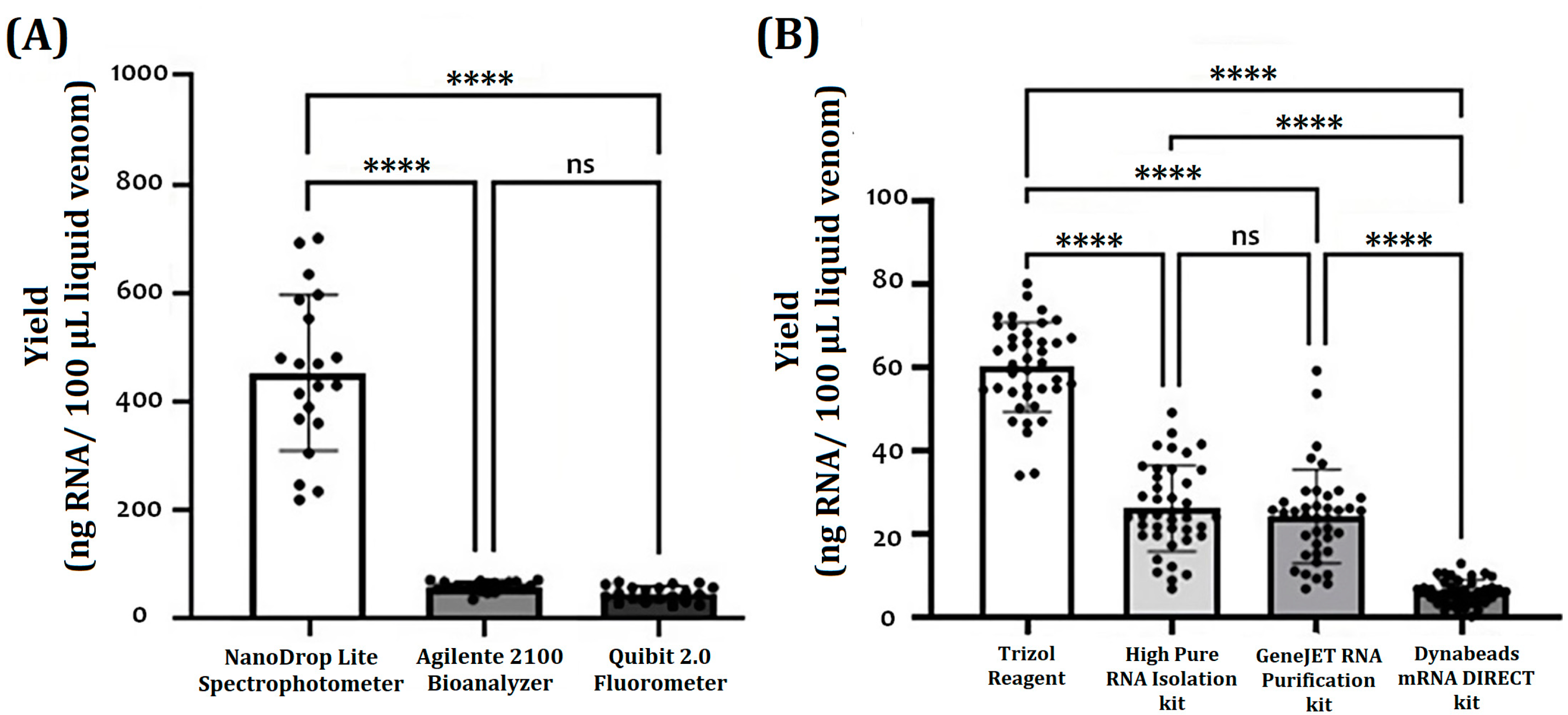
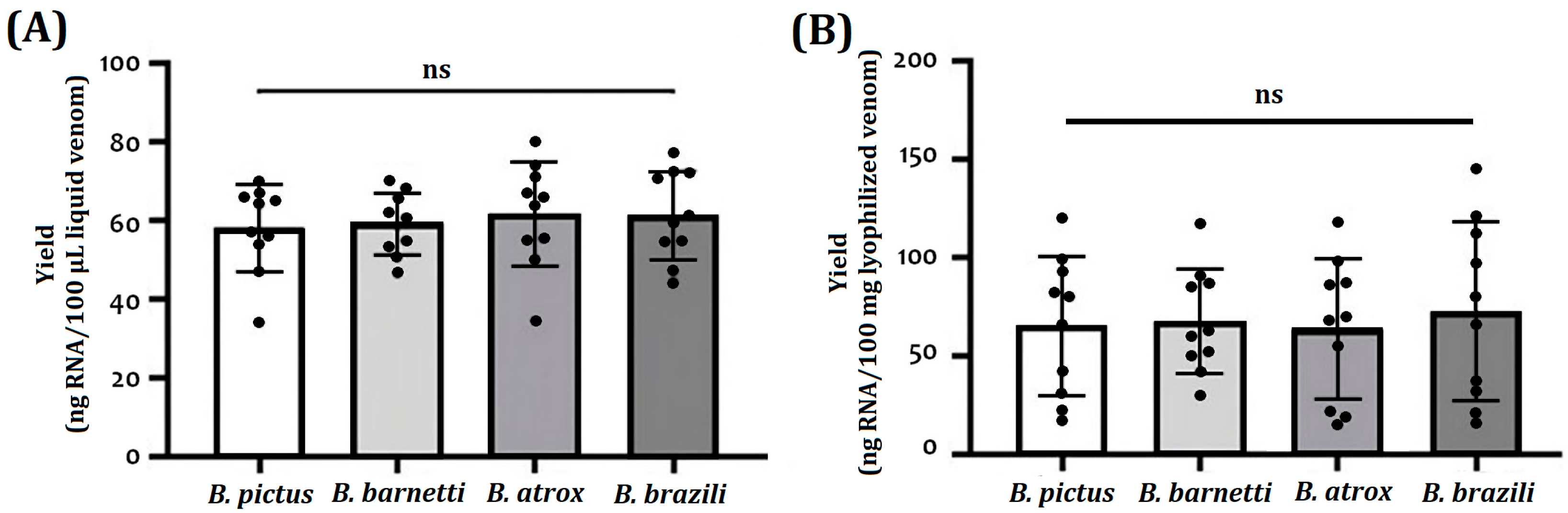
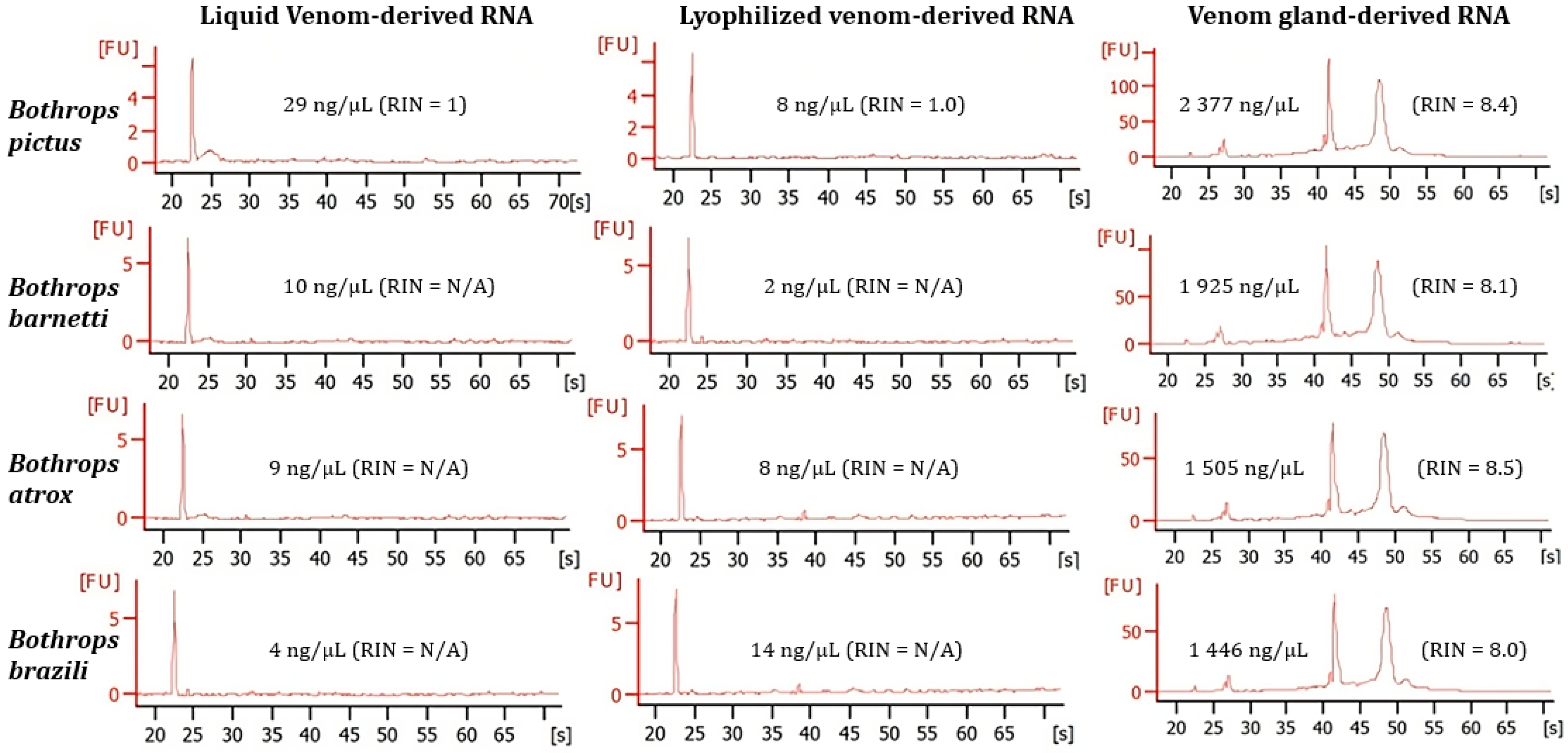
2. Results
2.1. RNA Purification
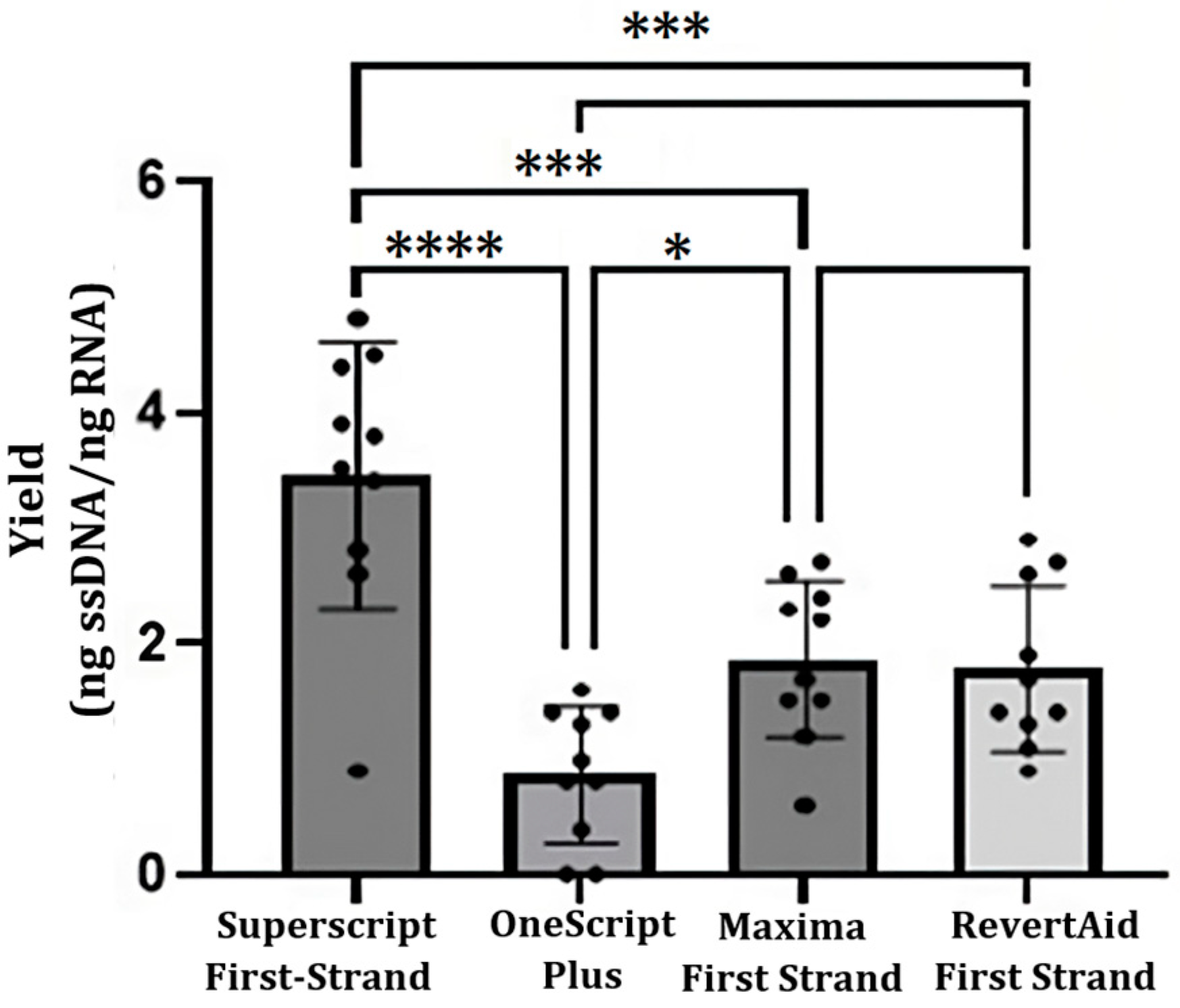
2.2. cDNA Synthesis
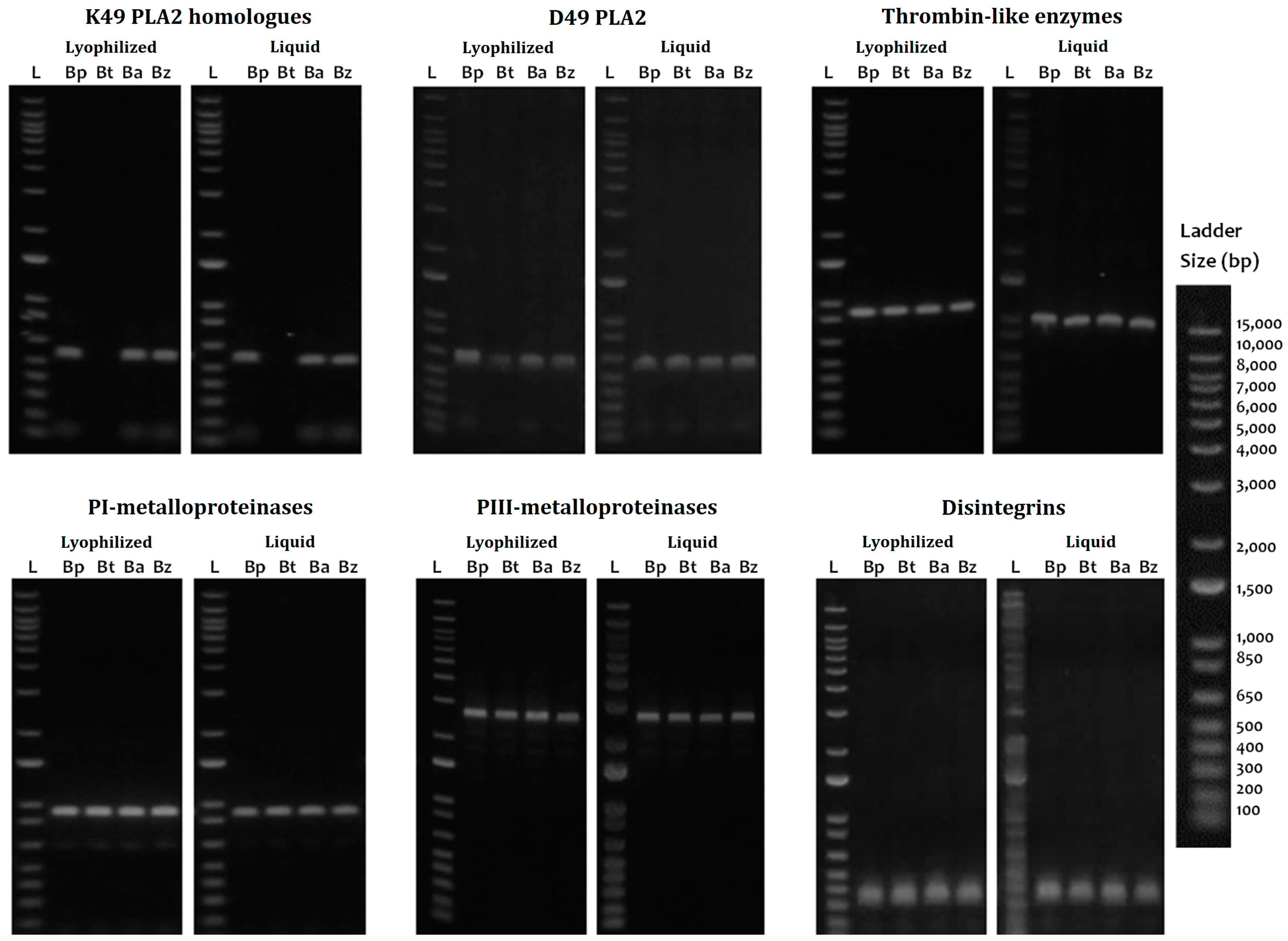
2.3. PCR Amplification
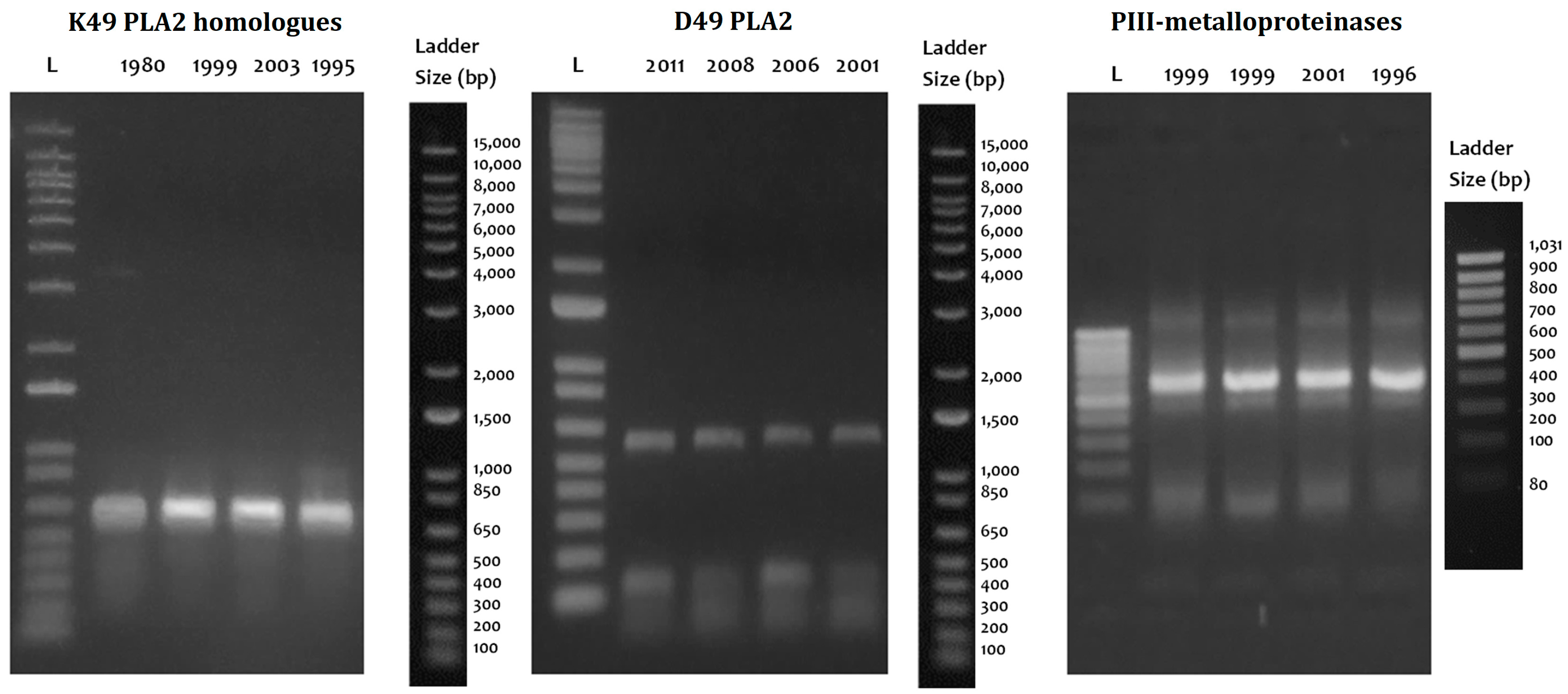
2.4. Cloning and Sequences Identification
3. Discussion
4. Materials and Methods
4.1. Venoms and Venom Gland Collection
4.2. RNA Isolation
4.3. cDNA Synthesis
4.4. Nucleic Acids Quality Control
4.5. Endpoint PCR Amplification
4.6. Cloning of Amplified Products and Sequencing
4.7. Sequence Analysis
4.8. Statistical Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Resiere, D.; Monteiro, W.; Houcke, S.; Pujo, J.M.; Mathien, C.; Mayence, C.; Neviere, R.; Hommel, D.; de Almeida Gonçalves Sachett, J.; Mehdaoui, H.; et al. Bothrops Snakebite Envenomings in the Amazon Region. Curr. Trop. Med. Rep. 2020, 7, 48–60. [Google Scholar] [CrossRef]
- Chippaux, J.P. Snakebite Envenomation Turns Again into a Neglected Tropical Disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef]
- Luna, K.P.O.; da Silva, M.B.; Pereira, V.R.A. Clinical and Immunological Aspects of Envenomations by Bothrops Snakes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Larréché, S.; Chippaux, J.-P.; Chevillard, L.; Mathé, S.; Résière, D.; Siguret, V.; Mégarbane, B. Bleeding and Thrombosis: Insights into Pathophysiology of Bothrops Venom-Related Hemostasis Disorders. Int. J. Mol. Sci. 2021, 22, 9643. [Google Scholar] [CrossRef] [PubMed]
- Mamede, C.C.N.; de Sousa Simamoto, B.B.; da Cunha Pereira, D.F.; de Oliveira Costa, J.; Ribeiro, M.S.M.; de Oliveira, F. Edema, Hyperalgesia and Myonecrosis Induced by Brazilian Bothropic Venoms: Overview of the Last Decade. Toxicon 2020, 187, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.L.M.M.; da Silva Junior, G.B.; Meneses, G.C.; Martins, A.M.C.; Lima, D.B.; Raubenheimer, J.; Fathima, S.; Buckley, N.; Daher, E.D.F. Acute Kidney Injury Induced by Bothrops Venom: Insights into the Pathogenic Mechanisms. Toxins 2019, 11, 148. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Aziz, T.M.; Soares, A.G.; Stockand, J.D. Advances in Venomics: Modern Separation Techniques and Mass Spectrometry. J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 2020, 1160, 122352. [Google Scholar] [CrossRef]
- Brahma, R.K.; McCleary, R.J.R.; Kini, R.M.; Doley, R. Venom Gland Transcriptomics for Identifying, Cataloging, and Characterizing Venom Proteins in Snakes. Toxicon 2015, 93, 1–10. [Google Scholar] [CrossRef]
- Tan, C.H. Snake Venomics: Fundamentals, Recent Updates, and a Look to the Next Decade. Toxins 2022, 14, 247. [Google Scholar] [CrossRef]
- Whiteley, G.; Logan, R.A.E.; Leung, K.Y.D.; Newberry, F.J.; Rowley, P.D.; Dunbar, J.P.; Wagstaff, S.C.; Casewell, N.R.; Harrison, R.A. Stabilising the Integrity of Snake Venom MRNA Stored under Tropical Field Conditions Expands Research Horizons. PLoS Negl. Trop. Dis. 2016, 10, e0004615. [Google Scholar] [CrossRef] [Green Version]
- Sells, P.G.; Hommel, M.; Theakston, R.D.G. Venom Production in Snake Venom Gland Cells Cultured In Vitro. Toxicon 1989, 27, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Post, Y.; Puschhof, J.; Beumer, J.; Kerkkamp, H.M.; de Bakker, M.A.G.; Slagboom, J.; de Barbanson, B.; Wevers, N.R.; Spijkers, X.M.; Olivier, T.; et al. Snake Venom Gland Organoids. Cell 2020, 180, 233–247.e21. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Mackessy, S.P. Full-Length Venom Protein CDNA Sequences from Venom-Derived MRNA: Exploring Compositional Variation and Adaptive Multigene Evolution. PLoS Negl. Trop. Dis. 2016, 10, e0004587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proleón, A.; Torrejón, D.; Urra, F.A.; Lazo, F.; López-Torres, C.; Fuentes-Retamal, S.; Quispe, E.; Bautista, L.; Agurto, A.; Gavilan, R.G.; et al. Functional, Immunological Characterization, and Anticancer Activity of BaMtx: A New Lys49- PLA2 Homologue Isolated from the Venom of Peruvian Bothrops atrox Snake (Serpentes: Viperidae). Int. J. Biol. Macromol. 2022, 206, 990–1002. [Google Scholar] [CrossRef] [PubMed]
- Sobrinho, J.C.; Kayano, A.M.; Simões-Silva, R.; Alfonso, J.J.; Gomez, A.F.; Gomez, M.C.V.; Zanchi, F.B.; Moura, L.A.; Souza, V.R.; Fuly, A.L.; et al. Anti-Platelet Aggregation Activity of Two Novel Acidic Asp49-Phospholipases A2 from Bothrops brazili Snake Venom. Int. J. Biol. Macromol. 2018, 107, 1014–1022. [Google Scholar] [CrossRef]
- Silva, M.A.; Lopes, D.S.; Teixeira, S.C.; Gimenes, S.N.C.; Azevedo, F.V.P.V.; Polloni, L.; Borges, B.C.; da Silva, M.S.; Barbosa, M.J.; de Oliveira Júnior, R.J.; et al. Genotoxic Effects of BnSP-6, a Lys-49 Phospholipase A2 (PLA2) Homologue from Bothrops pauloensis Snake Venom, on MDA-MB-231 Breast Cancer Cells. Int. J. Biol. Macromol. 2018, 118, 311–319. [Google Scholar] [CrossRef]
- Vivas-Ruiz, D.E.; Rosas, P.; Proleón, A.; Torrejón, D.; Lazo, F.; Tenorio-Ricca, A.B.; Guajardo, F.; Almarza, C.; Andrades, V.; Astorga, J.; et al. Pictolysin-III, a Hemorrhagic Type-III Metalloproteinase Isolated from Bothrops pictus (Serpentes: Viperidae) Venom, Reduces Mitochondrial Respiration and Induces Cytokine Secretion in Epithelial and Stromal Cell Lines. Pharmaceutics 2023, 15, 1533. [Google Scholar] [CrossRef]
- Cerda, P.A.; Crowe-Riddell, J.M.; Gonçalves, D.J.P.; Larson, D.A.; Duda, T.F.; Davis Rabosky, A.R. Divergent Specialization of Simple Venom Gene Profiles among Rear-Fanged Snake Genera (Helicops and Leptodeira, Dipsadinae, Colubridae). Toxins 2022, 14, 489. [Google Scholar] [CrossRef]
- Li, M.; Fry, B.G.; Kini, R.M. Putting the Brakes on Snake Venom Evolution: The Unique Molecular Evolutionary Patterns of Aipysurus eydouxii (Marbled Sea Snake) Phospholipase A2 Toxins. Mol. Biol. Evol. 2005, 22, 934–941. [Google Scholar] [CrossRef] [Green Version]
- Lazo, F.; Vivas-Ruiz, D.E.; Sandoval, G.A.; Rodríguez, E.F.; Kozlova, E.E.G.; Costal-Oliveira, F.; Chávez-Olórtegui, C.; Severino, R.; Yarlequé, A.; Sanchez, E.F. Biochemical, Biological and Molecular Characterization of an L-Amino Acid Oxidase (LAAO) Purified from Bothrops pictus Peruvian Snake Venom. Toxicon 2017, 139, 74–86. [Google Scholar] [CrossRef]
- Fonseca, A.; Renjifo-Ibáñez, C.; Renjifo, J.M.; Cabrera, R. Protocol to Obtain Targeted Transcript Sequence Data from Snake Venom Samples Collected in the Colombian Field. Toxicon 2018, 148, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Currier, R.; Calvete, J.; Harrison, R.; Rowley, P. Unusual Stability of Messenger RNA in Snake Venom Reveals Gene Expression Dynamics of Venom Replenishment. PLoS ONE 2012, 7, e41888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amazonas, D.R.; Portes-Junior, J.A.; Nishiyama, M.Y., Jr.; Nicolau, C.A.; Chalkidis, H.M.; Mourão, R.H.V.; Grazziotin, F.G.; Rokyta, D.R.; Gibbs, H.L.; Valente, R.H.; et al. Molecular Mechanisms Underlying Intraspecific Variation in Snake Venom. J. Proteom. 2018, 181, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Freitas-De-Sousa, L.A.; Amazonas, D.R.; Sousa, L.F.; Sant’Anna, S.S.; Nishiyama, M.Y.; Serrano, S.M.T.; Junqueira-De-Azevedo, I.L.M.; Chalkidis, H.M.; Moura-Da-Silva, A.M.; Mourão, R.H.V. Comparison of Venoms from Wild and Long-Term Captive Bothrops atrox Snakes and Characterization of Batroxrhagin, the Predominant Class PIII Metalloproteinase from the Venom of This Species. Biochimie 2015, 118, 60–70. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.S.; In, M.; Alvarenga, G.; Eble, J.A.; Sanchez, E.F. Atroxlysin-III, A Metalloproteinase from the Venom of the Peruvian Pit Viper Snake Bothrops atrox (Jergón) Induces Glycoprotein VI Shedding and Impairs Platelet Function. Molecules 2019, 24, 3489. [Google Scholar] [CrossRef] [Green Version]
- Quispe, E.; Torrejón, D.; Bautista, L.; Sandoval, G.; Rodríguez, E.; Lazo, F.; vivas-Ruiz, D.; Yarlequé, A.; Quispe, E.; Torrejón, D.; et al. Purificación y Algunas Propiedades Bioquímicas y Moleculares de una Nueva Fosfolipasa A2 No Miotóxica del Veneno de la Serpiente Bothrops atrox. Rev. Soc. Química Perú 2019, 85, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Vivas-Ruiz, D.E.; Sandoval, G.A.; Mendoza, J.; Inga, R.R.; Gontijo, S.; Richardson, M.; Eble, J.A.; Yarleque, A.; Sanchez, E.F. Coagulant Thrombin-like Enzyme (Barnettobin) from Bothrops barnetti Venom: Molecular Sequence Analysis of Its CDNA and Biochemical Properties. Biochimie 2013, 95, 1476–1486. [Google Scholar] [CrossRef] [Green Version]
- Vandenplas, M.L.; Vandenplas, S.; Brebner, K.; Bester, A.J.; Boyd, C.D. Characterization of the Messenger RNA Population Coding for Components of Viperid Snake Venom. Toxicon 1985, 23, 289–305. [Google Scholar] [CrossRef]
- Vivas-Ruiz, D.; Sandoval, G.A.; Gonzalez-Kozlova, E.; Zarria-Romero, J.; Lazo, F.; Rodríguez, E.; Magalhães, H.P.B.; Chávez-Olortegui, C.; Oliveira, L.S.; Alvarenga, V.G.; et al. Fibrinogen-Clotting Enzyme, Pictobin, from Bothrops pictus Snake Venom. Structural and Functional Characterization. Int. J. Biol. Macromol. 2020, 153, 779–795. [Google Scholar] [CrossRef]
- Vivas-Ruiz, D.; Gonzalez-Kozlova, E.E.; Delgadillo, J.; Palermo, P.M.; Sandoval, G.A.; Lazo, F.; Rodríguez, E.; Chávez-Olórtegui, C.; Yarlequé, A.; Sanchez, E.F. Biochemical and Molecular Characterization of the Hyaluronidase from Bothrops atrox Peruvian Snake Venom. Biochimie 2019, 162, 33–45. [Google Scholar] [CrossRef]
- Kałużna, M.; Kuras, A.; Mikiciński, A.; Puławska, J. Evaluation of Different RNA Extraction Methods for High-Quality Total RNA and MRNA from Erwinia Amylovora in Planta. Eur. J. Plant Pathol. 2016, 146, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Scholes, A.N.; Lewis, J.A. Comparison of RNA Isolation Methods on RNA-Seq: Implications for Differential Expression and Meta-Analyses. BMC Genom. 2020, 21, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, X.; Qiu, D.; Hegele, R.D.; Tan, W.C. Comparison of Different Methods of Total RNA Extraction for Viral Detection in Sputum. J. Virol. Methods 2001, 94, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.; Sun, S.; Chen, J.; Chen, J.; Zhou, E. Comparison of Different Methods for Total RNA Extraction from Sclerotia of Rhizoctonia solani. Electron. J. Biotechnol. 2014, 17, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Toni, L.S.; Garcia, A.M.; Jeffrey, D.A.; Jiang, X.; Stauffer, B.L.; Miyamoto, S.D.; Sucharov, C.C. Optimization of Phenol-Chloroform RNA Extraction. MethodsX 2018, 5, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Tomo, S.; Modi, A.; Purohit, P.; Sharma, P. Optimising Total RNA Quality and Quantity by Phenol-Chloroform Extraction Method from Human Visceral Adipose Tissue: A Standardisation Study. MethodsX 2020, 7, 101113. [Google Scholar] [CrossRef]
- Macedo, N.; Ferreira, T. Maximizing Total RNA Yield from TRIzol® Reagent Protocol: A Feasibility Study. In Proceedings of the ASEE Zone I Conference, Bridgpeort, CT, USA, 3–5 April 2014. [Google Scholar]
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef]
- Perry, B.W.; Schield, D.R.; Westfall, A.K.; Mackessy, S.P.; Castoe, T.A. Physiological Demands and Signaling Associated with Snake Venom Production and Storage Illustrated by Transcriptional Analyses of Venom Glands. Sci. Rep. 2020, 10, 18083. [Google Scholar] [CrossRef]
- Aranda IV, R.; Dineen, S.M.; Craig, R.L.; Guerrieri, R.A.; Robertson, J.M. Comparison and Evaluation of RNA Quantification Methods Using Viral, Prokaryotic, and Eukaryotic RNA over a 10(4) Concentration Range. Anal. Biochem. 2009, 387, 122–127. [Google Scholar] [CrossRef]
- Hasegawa, T.; Hapsari, D.; Iwahashi, H. Challenges for Accurate Quantification of RNA. Rev. Agric. Sci. 2021, 9, 1–19. [Google Scholar] [CrossRef]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA Delivery by Extracellular Vesicles in Mammalian Cells and Its Applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef] [PubMed]
- Carregari, V.C.; Rosa-Fernandes, L.; Baldasso, P.; Bydlowski, S.P.; Marangoni, S.; Larsen, M.R.; Palmisano, G. Snake Venom Extracellular Vesicles (SVEVs) Reveal Wide Molecular and Functional Proteome Diversity. Sci. Rep. 2018, 8, 12067. [Google Scholar] [CrossRef] [Green Version]
- Kohlhoff, M.; Borges, M.H.; Yarleque, A.; Cabezas, C.; Richardson, M.; Sanchez, E.F. Exploring the Proteomes of the Venoms of the Peruvian Pit Vipers Bothrops atrox, B. barnetti and B. pictus. J. Proteom. 2012, 75, 2181–2195. [Google Scholar] [CrossRef] [PubMed]
- Núñez, V.; Cid, P.; Sanz, L.; De La Torre, P.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J. Snake Venomics and Antivenomics of Bothrops atrox Venoms from Colombia and the Amazon Regions of Brazil, Perú and Ecuador Suggest the Occurrence of Geographic Variation of Venom Phenotype by a Trend towards Paedomorphism. J. Proteom. 2009, 73, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.F.; Portes-Junior, J.A.; Nicolau, C.A.; Bernardoni, J.L.; Nishiyama-Jr, M.Y.; Amazonas, D.R.; Freitas-de-Sousa, L.A.; Mourão, R.H.; Chalkidis, H.M.; Valente, R.H.; et al. Functional Proteomic Analyses of Bothrops atrox Venom Reveals Phenotypes Associated with Habitat Variation in the Amazon. J. Proteom. 2017, 159, 32–46. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Chippaux, J.P. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins. Biol. Aujourdhui. 2010, 204, 87–91. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA Isolation Protocols | RNA Yield * per Species | |||
|---|---|---|---|---|
| B. pictus | B. barnetti | B. atrox | B. brazili | |
| Freshly collected venoms (ng RNA/100 µL venom) | 58 ± 11 | 59 ± 8 | 62 ± 13 | 61 ± 11 |
| Lyophilized venoms RNA Yield (ng RNA/10 mg venom) | 65 ± 35 | 68 ± 27 | 83 ± 21 | 73 ± 46 |
| Snake venom glands (ng RNA/mg tissue) | 120 ± 25 | 143 ± 26 | 156 ± 31 | 136 ± 28 |
| cDNA Yield (ng cDNA 1/ng RNA 1) | ||||
|---|---|---|---|---|
| Isolation Method | TRIzol Reagent Protocol | High Pure RNA Isolation Kit | GeneJET RNA Purification Kit | Dynabeads mRNA DIRECT Kit |
| SuperScript First-Strand Synthesis System | 3.2 ± 1.2 | 2.1 ± 1.5 | 2.7 ± 1.1 | 4.8 ± 2.0 |
| OneScript Plus cDNA Synthesis Kit | 0.8 ± 0.7 | 0.6 ± 0.4 | 0.9 ± 0.3 | 2.0 ± 1.0 |
| Maxima First Strand cDNA Synthesis Kit | 1.9 ± 0.9 | 2.1 ± 0.6 | n.d. | n.d. |
| RevertAid First Strand cDNA Synthesis Kit | 1.6 ± 0.4 | 2.4 ± 0.2 | n.d. | n.d. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torrejón, D.; Cárdenas, J.; Juárez, D.; Espinoza, J.; Proleón, A.; Agurto-Arteaga, A.; Lazo, F.; Leguía, M.; Urra, F.A.; Sánchez, E.F.; et al. Comparison of Four Methods of RNA Extraction and cDNA Synthesis from The Venom of Peruvian Snakes of the Genus Bothrops of Clinical Importance. Int. J. Mol. Sci. 2023, 24, 11161. https://doi.org/10.3390/ijms241311161
Torrejón D, Cárdenas J, Juárez D, Espinoza J, Proleón A, Agurto-Arteaga A, Lazo F, Leguía M, Urra FA, Sánchez EF, et al. Comparison of Four Methods of RNA Extraction and cDNA Synthesis from The Venom of Peruvian Snakes of the Genus Bothrops of Clinical Importance. International Journal of Molecular Sciences. 2023; 24(13):11161. https://doi.org/10.3390/ijms241311161
Chicago/Turabian StyleTorrejón, Daniel, Javier Cárdenas, Diana Juárez, Jordano Espinoza, Alex Proleón, Andrés Agurto-Arteaga, Fanny Lazo, Mariana Leguía, Félix A. Urra, Eladio F. Sánchez, and et al. 2023. "Comparison of Four Methods of RNA Extraction and cDNA Synthesis from The Venom of Peruvian Snakes of the Genus Bothrops of Clinical Importance" International Journal of Molecular Sciences 24, no. 13: 11161. https://doi.org/10.3390/ijms241311161
APA StyleTorrejón, D., Cárdenas, J., Juárez, D., Espinoza, J., Proleón, A., Agurto-Arteaga, A., Lazo, F., Leguía, M., Urra, F. A., Sánchez, E. F., Chávez-Olortegui, C., Vivas-Ruiz, D. E., & Yarlequé, A. (2023). Comparison of Four Methods of RNA Extraction and cDNA Synthesis from The Venom of Peruvian Snakes of the Genus Bothrops of Clinical Importance. International Journal of Molecular Sciences, 24(13), 11161. https://doi.org/10.3390/ijms241311161