
Impact of Exposomes on Ocular Surface Diseases



Abstract
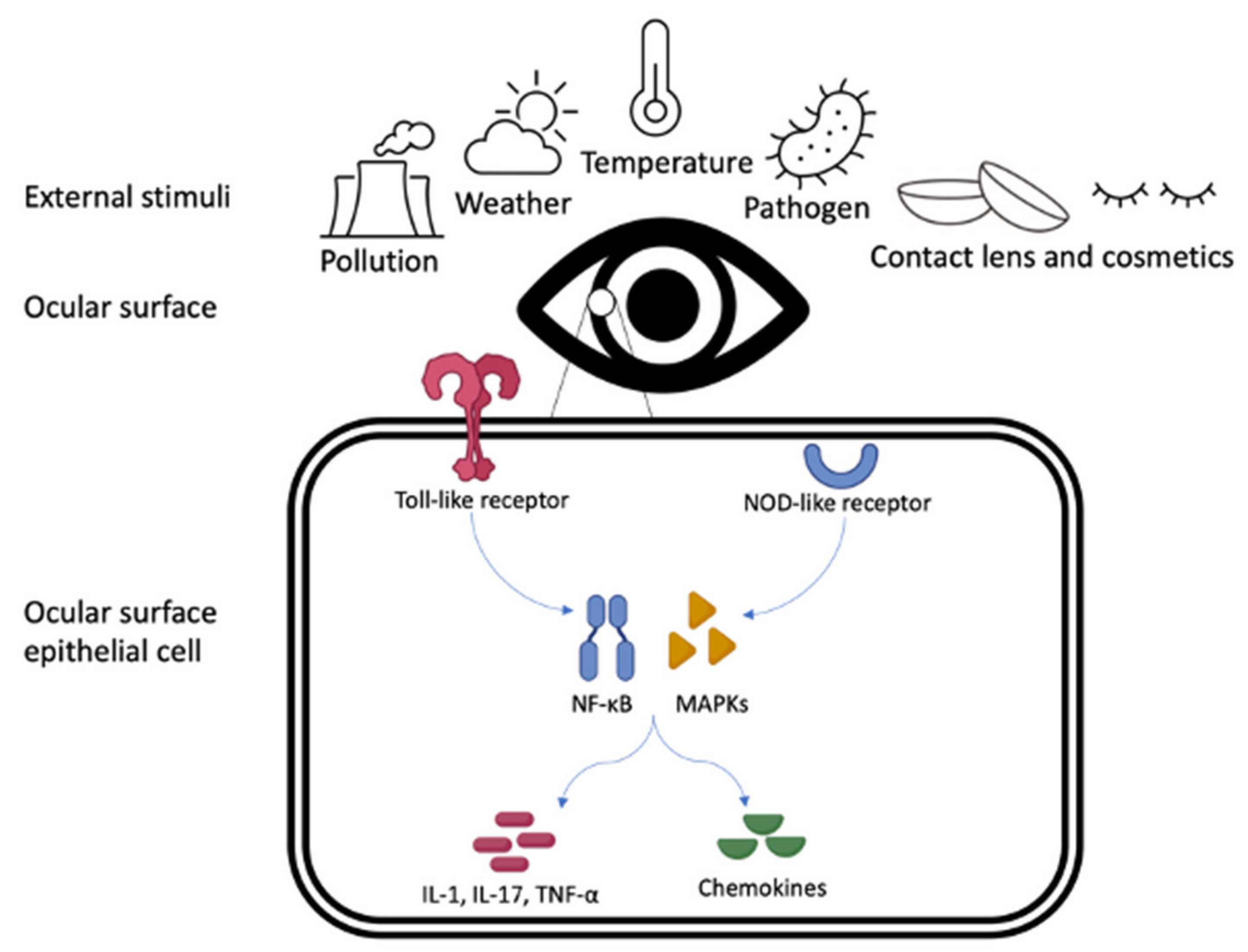
:1. Introduction
2. External Exposomes and the Ocular Surface
3. Internal Exposomes and the Ocular Surface
3.1. The Ocular Surface Microbiome in Health
3.2. Alterations in Ocular Surface Microbiome in Disease
4. Dry Eye Disease and Blepharitis/Meibomian Gland Dysfunction
4.1. Impact of Exposomes on Dry Eye Disease
4.2. External Exposome on Dry Eye Disease and Meibomian Gland Dysfunction
4.2.1. Environmental Pollution
4.2.2. Humidity, Temperature, Cleanliness and Screen Time
4.2.3. Contact Lens Wear
4.3. Internal Exposomes on Dry Eye Disease and Meibomian Gland Dysfunction
4.3.1. Ocular Surface Microbiome
4.3.2. Gut Microflora, Laryngopharyngeal Reflux (LPR) Disease
4.3.3. Impact of Exposome and the Drive towards Chronic Inflammation in Dry Eye Disease
5. Allergic Eye Disease
5.1. External Exposomes and Allergic Eye Disease
5.1.1. Contact Lens Wear, Pollution, Lack of Exposure to Microbes
5.1.2. Diet
5.2. Internal Exposomes and Allergic Eye Disease
Ocular Surface Microbiome
6. Cicatrising Conjnctivitis
6.1. External Exposomes on Cicatrising Conjunctivitis
Viral Infections and Drugs
6.2. Internal Exposomes on Cicatrising Conjunctivitis
6.2.1. Gut Microbiome
6.2.2. Ocular Surface Microbiome
7. Cosmetics: Common but Often Overlooked External Exposome with Impact on the Ocular Surface
8. Contact Lens Wear: An Example of External Exposome Impacting upon Internal Exposome of the Ocular Surface
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gipson, I.K. The ocular surface: The challenge to enable and protect vision: The Friedenwald lecture. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4390, 4391–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolaños-Jiménez, R.; Navas, A.; López-Lizárraga, E.P.; De Ribot, F.M.; Peña, A.; O Graue-Hernández, E.; Garfias, Y. Ocular Surface as Barrier of Innate Immunity. Open Ophthalmol. J. 2015, 9, 49–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, H.S.; Dart, J.K. Managing ocular surface disease: A common-sense approach. Community Eye Health 2016, 29, 44–46. [Google Scholar] [PubMed]
- Alves, M.; Asbell, P.; Dogru, M.; Giannaccare, G.; Grau, A.; Gregory, D.; Kim, D.H.; Marini, M.C.; Ngo, W.; Nowinska, A.; et al. TFOS Lifestyle Report: Impact of environmental conditions on the ocular surface. Ocul. Surf. 2023, 29, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Queiruga, J.; Pena-Verdeal, H.; Sabucedo-Villamarin, B.; Giraldez, M.J.; Garcia-Resua, C.; Yebra-Pimentel, E. A cross-sectional study of non-modifiable and modifiable risk factors of dry eye disease states. Cont. Lens. Anterior Eye 2023, 46, 101800. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Diebold, Y.; Sahu, S.K.; Leonardi, A. Epithelial barrier dysfunction in ocular allergy. Allergy 2022, 77, 1360–1372. [Google Scholar] [CrossRef]
- Aragona, P.; Baudouin, C.; del Castillo, J.M.B.; Messmer, E.; Barabino, S.; Merayo-Lloves, J.; Brignole-Baudouin, F.; Inferrera, L.; Rolando, M.; Mencucci, R.; et al. The ocular microbiome and microbiota and their effects on ocular surface pathophysiology and disorders. Surv. Ophthalmol. 2021, 66, 907–925. [Google Scholar] [CrossRef]
- Hou, A.; Tin, M.Q.; Tong, L. Toll-like receptor 2-mediated NF-kappa B pathway activation in ocular surface epithelial cells. Eye Vis. 2017, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Busanello, A.; Santucci, D.; Bonini, S.; Micera, A. Review: Environmental impact on ocular surface disorders: Possible epigenetic mechanism modulation and potential biomarkers. Ocul. Surf. 2017, 15, 680–687. [Google Scholar] [CrossRef]
- Ueta, M.; Kinoshita, S. Innate immunity of the ocular surface. Brain Res. Bull. 2010, 81, 219–228. [Google Scholar] [CrossRef]
- Kugadas, A.; Christiansen, S.H.; Sankaranarayanan, S.; Surana, N.K.; Gauguet, S.; Kunz, R.; Fichorova, R.; Vorup-Jensen, T.; Gadjeva, M. Impact of Microbiota on Resistance to Ocular Pseudomonas aeruginosa-Induced Keratitis. PLOS Pathog. 2016, 12, e1005855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kugadas, A.; Wright, Q.; Geddes-McAlister, J.; Gadjeva, M. Role of Microbiota in Strengthening Ocular Mucosal Barrier Function Through Secretory IgA. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4593–4600. [Google Scholar] [CrossRef]
- Leger, A.J.S.; Desai, J.V.; Drummond, R.A.; Kugadas, A.; Almaghrabi, F.; Silver, P.; Raychaudhuri, K.; Gadjeva, M.; Iwakura, Y.; Lionakis, M.S.; et al. An Ocular Commensal Protects against Corneal Infection by Driving an Interleukin-17 Response from Mucosal γδ T Cells. Immunity 2017, 47, 148–158.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, L.; Constancias, F.; Hou, A.; Chua, S.L.; Drautz-Moses, D.I.; Schuster, S.C.; Yang, L.; Williams, R.B.H.; Kjelleberg, S. Shotgun metagenomic sequencing analysis of ocular surface microbiome in Singapore residents with mild dry eye. Front. Med. 2022, 9, 1034131. [Google Scholar] [CrossRef] [PubMed]
- Cavuoto, K.M.; Mendez, R.; Miller, D.; Galor, A.; Banerjee, S. Effect of clinical parameters on the ocular surface microbiome in children and adults. Clin. Ophthalmol. 2018, 12, 1189–1197. [Google Scholar] [CrossRef] [Green Version]
- Cavuoto, K.M.; Banerjee, S.; Miller, D.; Galor, A. Composition and Comparison of the Ocular Surface Microbiome in Infants and Older Children. Transl. Vis. Sci. Technol. 2018, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Holland, M.J.; Makalo, P.; Joof, H.; Roberts, C.H.; Mabey, D.C.; Bailey, R.L.; Burton, M.J.; Weinstock, G.M.; E Burr, S. The conjunctival microbiome in health and trachomatous disease: A case control study. Genome Med. 2014, 6, 99. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Wen, X.; Hu, X.; Zou, Y.; Zhao, C.; Chen, X.; Miao, L.; Li, X.; Deng, X.; Bible, P.W.; et al. Geographic Difference Shaped Human Ocular Surface Metagenome of Young Han Chinese From Beijing, Wenzhou, and Guangzhou Cities. Investig. Opthalmol. Vis. Sci. 2020, 61, 47. [Google Scholar] [CrossRef] [Green Version]
- Cavuoto, K.M.; Galor, A.; Banerjee, S. Anatomic Characterization of the Ocular Surface Microbiome in Children. Microorganisms 2019, 7, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Yang, B.; Li, W. Defining the normal core microbiome of conjunctival microbial communities. Clin. Microbiol. Infect. 2016, 22, 643.e7–643.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.; Brulc, J.M.; Iovieno, A.; Bates, B.; Garoutte, A.; Miller, D.; Revanna, K.V.; Gao, X.; Antonopoulos, D.A.; Slepak, V.Z.; et al. Diversity of Bacteria at Healthy Human Conjunctiva. Investig. Opthalmology Vis. Sci. 2011, 52, 5408–5413. [Google Scholar] [CrossRef] [Green Version]
- Ozkan, J.; Nielsen, S.; Diez-Vives, C.; Coroneo, M.; Thomas, T.; Willcox, M. Temporal Stability and Composition of the Ocular Surface Microbiome. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Miao, L.; Deng, Y.; Bible, P.W.; Hu, X.; Zou, Y.; Liu, Y.; Guo, S.; Liang, J.; Chen, T.; et al. The Influence of Age and Sex on Ocular Surface Microbiota in Healthy Adults. Investig. Opthalmology Vis. Sci. 2017, 58, 6030–6037. [Google Scholar] [CrossRef]
- Doan, T.; Hinterwirth, A.; Worden, L.; Arzika, A.M.; Maliki, R.; Chen, C.; Zhong, L.; Zhou, Z.; Acharya, N.R.; Porco, T.C.; et al. Post-antibiotic Ocular Surface Microbiome in Children: A Cluster-Randomized Trial. Ophthalmology 2020, 127, 1127–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Price, K.; Albert, L.; Dodick, J.; Park, L.; Dominguez-Bello, M.G. Changes in the Eye Microbiota Associated with Contact Lens Wearing. mBio 2016, 7, e00198-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhao, F.; Hutchinson, D.S.; Sun, W.; Ajami, N.J.; Lai, S.; Wong, M.C.; Petrosino, J.F.; Fang, J.; Jiang, J.; et al. Conjunctival Microbiome Changes Associated With Soft Contact Lens and Orthokeratology Lens Wearing. Investig. Opthalmol. Vis. Sci. 2017, 58, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Graham, J.E.; Moore, J.E.; Jiru, X.; Moore, J.E.; Goodall, E.A.; Dooley, J.S.G.; Hayes, V.E.A.; Dartt, D.A.; Downes, C.S.; Moore, T.C.B. Ocular Pathogen or Commensal: A PCR-Based Study of Surface Bacterial Flora in Normal and Dry Eyes. Investig. Opthalmol. Vis. Sci. 2007, 48, 5616–5623. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, D.H.; Jung, J.Y.; Kim, J.C.; Jeon, C.O. Comparative ocular microbial communities in humans with and without blepharitis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5585–5593. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gong, Y.; Chen, S.; Li, S.; Zhang, Y.; Zhong, H.; Wang, Z.; Chen, Y.; Deng, Q.; Jiang, Y.; et al. Comparative portrayal of ocular surface microbe with and without dry eye. J. Microbiol. 2019, 57, 1025–1032. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Y.; Wang, W.; Lin, P.; Huang, Y. Composition and Diversity of Bacterial Community on the Ocular Surface of Patients with Meibomian Gland Dysfunction. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4774–4783. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zhang, D.; Ge, C.; Zhang, L.; Reinach, P.S.; Tian, X.; Tao, C.; Zhao, Z.; Zhao, C.; Fu, W.; et al. Metagenomic Profiling of Ocular Surface Microbiome Changes in Meibomian Gland Dysfunction. Investig. Opthalmol. Vis. Sci. 2020, 61, 22. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Li, J.; Zhang, S.; Liao, Y.; Guo, S.; Liang, J.; Deng, X.; Liu, Y.; Zou, B.; Wen, X.; et al. Characterization of conjunctival microbiome dysbiosis associated with allergic conjunctivitis. Allergy 2020, 76, 596–600. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, A.; Modugno, R.L.; Cavarzeran, F.; Rosani, U. Metagenomic analysis of the conjunctival bacterial and fungal microbiome in vernal keratoconjunctivitis. Allergy 2021, 76, 3215–3217. [Google Scholar] [CrossRef]
- Zilliox, M.J.; Gange, W.; Kuffel, G.; Mores, C.R.; Joyce, C.; de Bustros, P.; Bouchard, C.S. Assessing the ocular surface microbiome in severe ocular surface diseases. Ocul. Surf. 2020, 18, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Kittipibul, T.; Puangsricharern, V.; Chatsuwan, T. Comparison of the ocular microbiome between chronic Stevens-Johnson syndrome patients and healthy subjects. Sci. Rep. 2020, 10, 4353. [Google Scholar] [CrossRef] [Green Version]
- Ueta, M.; Hosomi, K.; Park, J.; Mizuguchi, K.; Sotozono, C.; Kinoshita, S.; Kunisawa, J. Categorization of the Ocular Microbiome in Japanese Stevens–Johnson Syndrome Patients With Severe Ocular Complications. Front. Cell. Infect. Microbiol. 2021, 11, 741654. [Google Scholar] [CrossRef]
- Berry, M.; Harris, A.; Lumb, R.; Powell, K. Commensal ocular bacteria degrade mucins. Br. J. Ophthalmol. 2002, 86, 1412–1416. [Google Scholar] [CrossRef]
- Aristoteli, L.P.; Bojarski, B.; Willcox, M.D. Isolation of conjunctival mucin and differential interaction with Pseudomonas aeruginosa strains of varied pathogenic potential. Exp. Eye Res. 2003, 77, 699–710. [Google Scholar] [CrossRef]
- Vidal-Rohr, M.; Craig, J.P.; Davies, L.N.; Wolffsohn, J.S. The epidemiology of dry eye disease in the UK: The Aston dry eye study. Cont. Lens. Anterior Eye 2023, 46, 101837. [Google Scholar] [CrossRef]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.-K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Berg, E.J.; Ying, G.-S.; Maguire, M.G.; Sheffield, P.E.; Szczotka-Flynn, L.B.; Asbell, P.A.; Shen, J.F.; the DREAM Study Research Group. Climatic and Environmental Correlates of Dry Eye Disease Severity: A Report from the Dry Eye Assessment and Management (DREAM) Study. Transl. Vis. Sci. Technol. 2020, 9, 25. [Google Scholar] [CrossRef] [PubMed]
- Calonge, M.; Pinto-Fraga, J.; González-García, M.J.; Enríquez-De-Salamanca, A.; la Rosa, A.L.-D.; Fernández, I.; López-Miguel, A. Effects of the External Environment on Dry Eye Disease. Int. Ophthalmol. Clin. 2017, 57, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.J.; Mehta, J.S.; Tong, L. Effects of environment pollution on the ocular surface. Ocul. Surf. 2018, 16, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Novaes, H.M.D.; Gouveia, N.; de Medeiros, A.P.P. Perinatal mortality and traffic-related air pollution. Rev. Bras. Ginecol. Obstet. Rev. Fed. Bras. Soc. Ginecol. Obstet. 2010, 32, 471–475. [Google Scholar]
- Malerbi, F.K.; Martins, L.C.; Saldiva, P.H.N.; Braga, A.L.F. Ambient levels of air pollution induce clinical worsening of blepharitis. Environ. Res. 2012, 112, 199–203. [Google Scholar] [CrossRef]
- Hwang, S.H.; Choi, Y.H.; Paik, H.J.; Wee, W.R.; Kim, M.K.; Kim, D.H. Potential Importance of Ozone in the Association Between Outdoor Air Pollution and Dry Eye Disease in South Korea. JAMA Ophthalmol. 2016, 134, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Ho, R.C.; Zhang, M.W.; Ho, C.S.; Pan, F.; Lu, Y.; Sharma, V.K. Impact of 2013 south Asian haze crisis: Study of physical and psychological symptoms and perceived dangerousness of pollution level. BMC Psychiatry 2014, 14, 81. [Google Scholar] [CrossRef] [Green Version]
- Wolkoff, P. Ocular discomfort by environmental and personal risk factors altering the precorneal tear film. Toxicol. Lett. 2010, 199, 203–212. [Google Scholar] [CrossRef]
- Novaes, P.; Saldiva, P.H.D.N.; Kara-José, N.; Macchione, M.; Matsuda, M.; Racca, L.; Berra, A. Ambient Levels of Air Pollution Induce Goblet-Cell Hyperplasia in Human Conjunctival Epithelium. Environ. Heal. Perspect. 2007, 115, 1753–1756. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, E.K.; Kang, S.W.; Kim, J.H.; Hwang, H.J.; Kim, T.-I. Effects of ozone exposure on the ocular surface. Free Radic. Biol. Med. 2013, 63, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.J.; Schwartz, J.; Milton, D.K.; Burge, H.A. The work environment and workers’ health in four large office buildings. Environ. Health Perspect. 2003, 111, 1242–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolkoff, P. “Healthy” eye in office-like environments. Environ. Int. 2008, 34, 1204–1214. [Google Scholar] [CrossRef]
- Kojima, T. Contact Lens-Associated Dry Eye Disease: Recent Advances Worldwide and in Japan. Investig. Ophthalmol. Vis. Sci. 2018, 59, DES102–DES108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchino, M.; Schaumberg, D.A.; Dogru, M.; Uchino, Y.; Fukagawa, K.; Shimmura, S.; Satoh, T.; Takebayashi, T.; Tsubota, K. Prevalence of Dry Eye Disease among Japanese Visual Display Terminal Users. Ophthalmology 2008, 115, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- International Dry Eye WorkShop Study Group. The Epidemiology of Dry Eye Disease: Report of the Epidemiology Subcommittee of the International Dry Eye WorkShop (2007). Ocul. Surf. 2007, 5, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Abengózar-Vela, A.; Pinto, F.J.; González-Méijome, J.M.; Ralló, M.; Serés, C.; Calonge, M.; González-García, M.J. Contact Lens Case Cleaning Procedures Affect Storage Solution pH and Osmolality. Optom. Vis. Sci. 2011, 88, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.; Efron, N.; Bandamwar, K.; Barnett, M.; Jacobs, D.S.; Jalbert, I.; Pult, H.; Rhee, M.K.; Sheardown, H.; Shovlin, J.P.; et al. TFOS Lifestyle: Impact of contact lenses on the ocular surface. Ocul. Surf. 2023, 29, 175–219. [Google Scholar] [CrossRef]
- Magliulo, G.; Plateroti, R.; Plateroti, A.M. Gastroesophageal reflux disease and the presence of pepsin in the tears. Med. Hypotheses 2013, 80, 129–130. [Google Scholar] [CrossRef]
- Owji, N.; Abtahi, S.M. Does gastroesophageal reflux contribute to development of acquired nasolacrimal duct obstruction? Med. Hypotheses 2010, 74, 455–456. [Google Scholar] [CrossRef]
- Bonini, S.; Labetoulle, M.; Messmer, E.; Aragona, P.; Castillo, J.M.B.; Ciprandi, G.; Damiani, V.; Irkec, M.; Baudouin, C.; Rolando, M. Association of Dry Eye with Laryngopharyngeal Reflux in Clinical Practice. Curr. Eye Res. 2021, 47, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Mazzacane, D.; Damiani, V.; Silvestri, M.; Ciprandi, G.; Marino, P. Eye reflux: An ocular extraesophageal manifestation of gastric reflux. Int. J. Ophthalmol. 2018, 11, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Iannella, G.; Di Nardo, G.; Plateroti, R.; Rossi, P.; Plateroti, A.M.; Mariani, P.; Magliulo, G. Investigation of pepsin in tears of children with laryngopharyngeal reflux disease. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 2312–2315. [Google Scholar] [CrossRef] [PubMed]
- Lechien, J.R.; De Vos, N.; Everard, A.; Saussez, S. Laryngopharyngeal reflux: The microbiota theory. Med. Hypotheses 2021, 146, 110460. [Google Scholar] [CrossRef]
- Mayo-Yáñez, M.; Viña-Vázquez, S.; Lechien, J.R.; Chiesa-Estomba, C.M.; Calvo-Henríquez, C.; González-Torres, L. Involvement of Laryngopharyngeal Reflux in Ocular Diseases: A State-of-the-Art Review. J. Voice 2021. [Google Scholar] [CrossRef]
- Plateroti, R.; Sacchetti, M.; Magliulo, G.; Plateroti, A.M.; Pace, A.; Moramarco, A.; Lambiase, A.; Bruscolini, A. Evidence of Pepsin-Related Ocular Surface Damage and Dry Eye (PROD Syndrome) in Patients with Laryngopharyngeal Reflux. Life 2020, 10, 202. [Google Scholar] [CrossRef]
- Allen, A.; Pearson, J.P.; Blackburn, A.; Coan, R.M.; Hutton, D.A.; Mall, A.S. Pepsins and the Mucus Barrier in Peptic Ulcer Disease. Scand. J. Gastroenterol. 1988, 23, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Periman, L.M.; Perez, V.L.; Saban, D.R.; Lin, M.C.; Neri, P. The Immunological Basis of Dry Eye Disease and Current Topical Treatment Options. J. Ocul. Pharmacol. Ther. 2020, 36, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Ono, S.J.; Abelson, M.B. Allergic conjunctivitis: Update on pathophysiology and prospects for future treatment. J. Allergy Clin. Immunol. 2005, 115, 118–122. [Google Scholar] [CrossRef]
- Rodrigues, J.; Kuruvilla, M.E.; Vanijcharoenkarn, K.; Patel, N.; Hom, M.M.; Wallace, D.V. The spectrum of allergic ocular diseases. Ann. Allergy Asthma Immunol. 2021, 126, 240–254. [Google Scholar] [CrossRef]
- Fujishima, H.; Okada, N.; Matsumoto, K.; Fukagawa, K.; Igarashi, A.; Matsuda, A.; Ono, J.; Ohta, S.; Mukai, H.; Yoshikawa, M.; et al. The usefulness of measuring tear periostin for the diagnosis and management of ocular allergic diseases. J. Allergy Clin. Immunol. 2016, 138, 459–467.e2. [Google Scholar] [CrossRef] [Green Version]
- Backman, H.; Räisänen, P.; Hedman, L.; Stridsman, C.; Andersson, M.; Lindberg, A.; Lundbäck, B.; Rönmark, E. Increased prevalence of allergic asthma from 1996 to 2006 and further to 2016-results from three population surveys. Clin. Exp. Allergy 2017, 47, 1426–1435. [Google Scholar] [CrossRef] [Green Version]
- Bertelsen, R.J.; Brantsæter, A.L.; Magnus, M.C.; Haugen, M.; Myhre, R.; Jacobsson, B.; Longnecker, M.P.; Meltzer, H.M.; London, S.J. Probiotic milk consumption in pregnancy and infancy and subsequent childhood allergic diseases. J. Allergy Clin. Immunol. 2013, 133, 165–171.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemter, A.M.; Nagler, C.R. Influences on allergic mechanisms through gut, lung, and skin microbiome exposures. J. Clin. Investig. 2019, 129, 1483–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, A.; Castegnaro, A.; Valerio, A.L.; Lazzarini, D. Epidemiology of allergic conjunctivitis: Clinical appearance and treatment patterns in a population-based study. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 482–488. [Google Scholar] [CrossRef]
- Rosario, N.; Bielory, L. Epidemiology of allergic conjunctivitis. Curr. Opin. Allergy Clin. Immunol. 2011, 11, 471–476. [Google Scholar] [CrossRef]
- Dzidic, M.; Abrahamsson, T.R.; Artacho, A.; Collado, M.C.; Mira, A.; Jenmalm, M.C. Oral microbiota maturation during the first 7 years of life in relation to allergy development. Allergy 2018, 73, 2000–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, D.-W.; Min, H.J.; Kim, M.-S.; Whon, T.W.; Shin, N.-R.; Kim, P.S.; Kim, H.S.; Lee, J.Y.; Kang, W.; Choi, A.M.K.; et al. Dysbiosis of Inferior Turbinate Microbiota Is Associated with High Total IgE Levels in Patients with Allergic Rhinitis. Infect. Immun. 2018, 86, e00934-17. [Google Scholar] [CrossRef] [Green Version]
- Alkotob, S.S.; Cannedy, C.; Harter, K.; Movassagh, H.; Paudel, B.; Prunicki, M.; Sampath, V.; Schikowski, T.; Smith, E.; Zhao, Q.; et al. Advances and novel developments in environmental influences on the development of atopic diseases. Allergy 2020, 75, 3077–3086. [Google Scholar] [CrossRef]
- Leonardi, A.; Di Stefano, A.; Motterle, L.; Zavan, B.; Abatangelo, G.; Brun, P. Transforming growth factor-β/Smad—Signalling pathway and conjunctival remodelling in vernal keratoconjunctivitis. Clin. Exp. Allergy 2011, 41, 52–60. [Google Scholar] [CrossRef]
- Brun, P.; Tarricone, E.; Di Stefano, A.; Mattiuzzo, E.; Mehrbod, P.; Ghavami, S.; Leonardi, A. The regulatory activity of autophagy in conjunctival fibroblasts and its possible role in vernal keratoconjunctivitis. J. Allergy Clin. Immunol. 2020, 146, 1210–1213.e9. [Google Scholar] [CrossRef] [PubMed]
- Iovieno, A.; Lambiase, A.; Sacchetti, M.; Stampachiacchiere, B.; Micera, A.; Bonini, S. Preliminary evidence of the efficacy of probiotic eye-drop treatment in patients with vernal keratoconjunctivitis. Graefe’s Arch. Clin. Exp. Ophthalmol. 2008, 246, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Andrés, S.; García, M.L.; Espina, M.; Valero, J.; Valls, O. Tear pH, air pollution, and contact lenses. Am. J. Optom. Physiol. Opt. 1988, 65, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, A.A.M.; Novaes, P.; Matsuda, M.; Braga, A.; Saldiva, P.H.N.; Alves, M.R.; Monteiro, M.L.R. Correlation between signs and symptoms of ocular surface dysfunction and tear osmolarity with ambient levels of air pollution in a large metropolitan area. Cornea 2013, 32, e11–e15. [Google Scholar] [CrossRef]
- Huang, S.K.; Zhang, Q.; Qiu, Z.; Chung, K.F. Mechanistic impact of outdoor air pollution on asthma and allergic diseases. J. Thorac. Dis. 2015, 7, 23–33. [Google Scholar] [CrossRef]
- Fujishima, H.; Satake, Y.; Okada, N.; Kawashima, S.; Matsumoto, K.; Saito, H. Effects of diesel exhaust particles on primary cultured healthy human conjunctival epithelium. Ann. Allergy Asthma Immunol. 2013, 110, 39–43. [Google Scholar] [CrossRef]
- Hong, J.; Zhong, T.; Li, H.; Xu, J.; Ye, X.; Mu, Z.; Lu, Y.; Mashaghi, A.; Zhou, Y.; Tan, M.; et al. Ambient air pollution, weather changes, and outpatient visits for allergic conjunctivitis: A retrospective registry study. Sci. Rep. 2016, 6, 23858. [Google Scholar] [CrossRef] [Green Version]
- Riediker, M.; Monn, C.; Koller, T.; Stahel, W.A.; Wüthrich, B. Air pollutants enhance rhinoconjunctivitis symptoms in pollen-allergic individuals. Ann. Allergy Asthma Immunol. 2001, 87, 311–318. [Google Scholar] [CrossRef]
- Miyazaki, D.; Fukagawa, K.; Fukushima, A.; Fujishima, H.; Uchio, E.; Ebihara, N.; Shoji, J.; Takamura, E.; Namba, K.; Ohashi, Y.; et al. Air pollution significantly associated with severe ocular allergic inflammatory diseases. Sci. Rep. 2019, 9, 18205. [Google Scholar] [CrossRef] [Green Version]
- Ellwood, P.; Asher, M.I.; Björkstén, B.; Burr, M.; Pearce, N.; Robertson, C.F. Diet and asthma, allergic rhinoconjunctivitis and atopic eczema symptom prevalence: An ecological analysis of the International Study of Asthma and Allergies in Childhood (ISAAC) data. ISAAC Phase One Study Group. Eur. Respir. J. 2001, 17, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Glatz, M.; Bosshard, P.P.; Hoetzenecker, W.; Schmid-Grendelmeier, P. The Role of Malassezia spp. in Atopic Dermatitis. J. Clin. Med. 2015, 4, 1217–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparber, F.; Ruchti, F.; LeibundGut-Landmann, S. Host Immunity to Malassezia in Health and Disease. Front. Cell Infect. Microbiol. 2020, 10, 198. [Google Scholar] [CrossRef] [PubMed]
- Alnahas, S.; Hagner, S.; Raifer, H.; Kilic, A.; Gasteiger, G.; Mutters, R.; Hellhund, A.; Prinz, I.; Pinkenburg, O.; Visekruna, A.; et al. IL-17 and TNF-alpha Are Key Mediators of Moraxella catarrhalis Triggered Exacerbation of Allergic Airway Inflammation. Front. Immunol. 2017, 8, 1562. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Lin, J.; Lu, F.; Zhang, X.; Zhang, L.; Gandhi, N.B.; de Paiva, C.S.; Pflugfelder, S.C.; Li, D.-Q. Potential autocrine regulation of interleukin-33/ST2 signaling of dendritic cells in allergic inflammation. Mucosal Immunol. 2013, 6, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Leonardi, A.; Daull, P.; Garrigue, J.-S.; Cavarzeran, F.; Docquier, M.; Di Stefano, A.; Tarricone, E.; Brun, P. Conjunctival transcriptome analysis reveals the overexpression of multiple pattern recognition receptors in vernal keratoconjunctivitis. Ocul. Surf. 2021, 19, 241–248. [Google Scholar] [CrossRef]
- Dart, J.K. The 2016 Bowman Lecture Conjunctival curses: Scarring conjunctivitis 30 years on. Eye 2017, 31, 301–332. [Google Scholar] [CrossRef] [Green Version]
- Ong, H.S.; Minassian, D.; Rauz, S.; Mehta, J.S.; Dart, J.K. Validation of a clinical assessment tool for cicatrising conjunctivitis. Ocul. Surf. 2020, 18, 121–129. [Google Scholar] [CrossRef]
- Ong, H.S.; Dart, J.K.; Mehta, J.S. A Review of Clinical Disease Scoring Systems for Cicatricial Diseases of the Conjunctiva. Front. Med. 2021, 8, 664572. [Google Scholar] [CrossRef]
- Radford, C.F.; Rauz, S.; Williams, G.P.; Saw, V.P.; Dart, J.K. Incidence, presenting features, and diagnosis of cicatrising conjunctivitis in the United Kingdom. Eye 2012, 26, 1199–1208. [Google Scholar] [CrossRef] [Green Version]
- Georgoudis, P.; Sabatino, F.; Szentmary, N.; Palioura, S.; Fodor, E.; Hamada, S.; Scholl, H.P.N.; Gatzioufas, Z. Ocular Mucous Membrane Pemphigoid: Current State of Pathophysiology, Diagnostics and Treatment. Ophthalmol. Ther. 2019, 8, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kittipibul, T.; Puangsricharern, V. The Ocular Microbiome in Stevens-Johnson Syndrome. Mini Review. Front. Med. 2021, 8, 645053. [Google Scholar] [CrossRef]
- Elhusseiny, A.M.; Fakhari, H.; Bishehsari, F.; Kang, K.; Djalilian, A.R. A gut connection in mucous membrane pemphigoid: Insights into the role of the microbiome. Ocul. Surf. 2019, 17, 615–616. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, M.C.; Foo, C.W.; Scholes, C.T.; Zone, J.J. Immunobullous disease and ulcerative colitis: A case series of six patients. Br. J. Dermatol. 2015, 173, 792–796. [Google Scholar] [CrossRef]
- Venugopal, R.; Satpathy, G.; Sangwan, S.; Kapil, A.; Aron, N.; Agarwal, T.; Pushker, N.; Sharma, N. Conjunctival Microbial Flora in Ocular Stevens-Johnson Syndrome Sequelae Patients at a Tertiary Eye Care Center. Cornea 2016, 35, 1117–1121. [Google Scholar] [CrossRef]
- Frizon, L.; Araújo, M.C.; Andrade, L.; Yu, M.C.Z.; Wakamatsu, T.H.; Höfling-Lima, A.L.; Gomes, J.P. Evaluation of conjunctival bacterial flora in patients with Stevens-Johnson Syndrome. Clinics 2014, 69, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, T.H.; dos Santos, M.S.; Barreiro, T.P.; Sant’Anna, A.E.B.P.P.; Murta, F.; da Costa, A.X.; Marculino, L.G.C.; de Alcântara, R.J.A.; de Farias, C.C.; Gomes, J.P. Clinical Aspects of Stevens-Johnson Syndrome and Toxic Epidermal Necrolysis With Severe Ocular Complications in Brazil. Mini Review. Front. Med. 2021, 8, 649369. [Google Scholar] [CrossRef]
- Gomes, J.A.P.; Santos, M.S.; Ventura, Â.S.; Donato, W.B.C.; Cunha, M.C.; Höfling-Lima, A.L. Amniotic Membrane With Living Related Corneal Limbal/Conjunctival Allograft for Ocular Surface Reconstruction in Stevens-Johnson Syndrome. Arch. Ophthalmol. 2003, 121, 1369–1374. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, D.A.; da Costa, A.X.; Del Duca, E.; Doll, T.; Grupcheva, C.N.; Lazreg, S.; Liu, S.-H.; McGee, S.R.; Murthy, R.; Narang, P.; et al. TFOS Lifestyle: Impact of cosmetics on the ocular surface. Ocul. Surf. 2023, 29, 77–130. [Google Scholar] [CrossRef]
- Ding, J.; Kam, W.R.; Dieckow, J.; Sullivan, D.A. The Influence of 13-cisRetinoic Acid on Human Meibomian Gland Epithelial Cells. Investig. Opthalmol. Vis. Sci. 2013, 54, 4341–4350. [Google Scholar] [CrossRef] [Green Version]
- Di Chen, D.; Wang, J.; Sullivan, D.A.M.; Kam, W.R.M.; Liu, Y. Effects of Terpinen-4-ol on Meibomian Gland Epithelial Cells In Vitro. Cornea 2020, 39, 1541–1546. [Google Scholar] [CrossRef]
- O’Dell, L.E.; Sullivan, A.; Periman, L. Beauty does not have to hurt. Adv. Ocul. Care 2016, 7, 42–47. [Google Scholar]
- O’Dell, L.; Sullivan, A.; Periman, L. If I could turn back time. Adv. Ocular. Care 2016, 25–29. [Google Scholar]
- O’Dell, L.E.; Sullivan, A.; Periman, L. Suffering for beauty: Harmful ingredients and trends in cosmetics. Adv. Ocul. Care 2016, 12–16. [Google Scholar]
- O’Dell, L.E.; Sullivan, A.G.; Periman, L. When beauty talk turns ugly. Adv. Ocular. Care 2017, 8, 16–20. [Google Scholar]
- Goldstein, M.H.; Silva, F.Q.; Blender, N.; Tran, T.; Vantipalli, S. Ocular benzalkonium chloride exposure: Problems and solutions. Eye 2022, 36, 361–368. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Kam, W.R.; Li, Y.; Sullivan, D.A. Toxicity of the cosmetic preservatives parabens, phenoxyethanol and chlorphenesin on human meibomian gland epithelial cells. Exp. Eye Res. 2020, 196, 108057. [Google Scholar] [CrossRef]
- Magerl, A.; Pirker, C.; Frosch, P.J. Allergic contact eczema from shellac and 1,3-butylene glycol in an eyeliner. J. Dtsch. Dermatol. Ges. 2003, 1, 300–302. [Google Scholar]
- Couteau, C.; Morin, T.; Diarra, H.; Coiffard, L. Influence of Cosmetic Type and Distribution Channel on the Presence of Regulated Fragrance Allergens: Study of 2044 Commercial Products. Clin. Rev. Allergy Immunol. 2020, 59, 101–108. [Google Scholar] [CrossRef]
- Le Coz, C.-J.; Leclere, J.-M.; Arnoult, E.; Raison-Peyron, N.; Pons-Guiraud, A.; Vigan, M.; Revidal-Gerda, T.M.O. Allergic contact dermatitis from shellac in mascara. Contact Dermat. 2002, 46, 149–152. [Google Scholar] [CrossRef]
- Prabhasawat, P.; Chirapapaisan, C.; Chitkornkijsin, C.; Pinitpuwadol, W.; Saiman, M.; Veeraburinon, A. Eyeliner Induces Tear Film Instability and Meibomian Gland Dysfunction. Cornea 2019, 39, 473–478. [Google Scholar] [CrossRef]
- Arciniega, J.C.; Uchiyama, E.; Butovich, I.A. Disruption and Destabilization of Meibomian Lipid Films Caused by Increasing Amounts of Ceramides and Cholesterol. Investig. Opthalmol. Vis. Sci. 2013, 54, 1352–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miano, F.; Mazzone, M.; Giannetto, A.; Enea, V.; Mc Cauley, P.; Bailey, A.; Winlove, P.C. Interface Properties of Simplified Tear-Like Fluids in Relation to Lipid and Aqueous Layers Composition. Lacrimal Gland. Tear Film. Dry Eye Syndr. 3 Basic Sci. Clin. Relev. Part A B 2002, 506, 405–417. [Google Scholar] [CrossRef]
- Patterson, M.; Vogel, H.J.; Prenner, E.J. The effect of repeated lateral compression and expansions mimicking blinking on selected tear film polar lipid monofilms. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1859, 319–330. [Google Scholar] [CrossRef]
- Olżyńska, A.; Cwiklik, L. Behavior of sphingomyelin and ceramide in a tear film lipid layer model. Ann. Anat.-Anat. Anz. 2017, 210, 128–134. [Google Scholar] [CrossRef]
- Amano, Y.; Sugimoto, Y.; Sugita, M. Ocular Disorders Due to Eyelash Extensions. Cornea 2012, 31, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Perra, M.T.; Serra, A.; Sirigu, P.; Turno, F. Histochemical demonstration of acetylcholinesterase activity in human Meibomian glands. Eur. J. Histochem. EJH 1996, 40, 39–44. [Google Scholar] [PubMed]
- Knop, E.; Knop, N.; Millar, T.; Obata, H.; Sullivan, D.A. The International Workshop on Meibomian Gland Dysfunction: Report of the Subcommittee on Anatomy, Physiology, and Pathophysiology of the Meibomian Gland. Investig. Opthalmol. Vis. Sci. 2011, 52, 1938–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagefi, M.R.; Dragan, L.; Hughes, S.M.; Klippenstein, K.A.; Seiff, S.R.; Woog, J.J. Adverse Reactions to Permanent Eyeliner Tattoo. Ophthalmic Plast. Reconstr. Surg. 2006, 22, 48–51. [Google Scholar] [CrossRef]
- Lee, Y.B.; Kim, J.J.; Hyon, J.Y.; Wee, W.R.; Shin, Y.J. Eyelid Tattooing Induces Meibomian Gland Loss and Tear Film Instability. Cornea 2015, 34, 750–755. [Google Scholar] [CrossRef]
- Naz, S.; Iqtedar, M.; Ain Qu Aftab, K. Incidence of Human Skin Pathogens from Cosmetic Tools used in Beauty Saloons in Different Areas of Lahore, Pakistan. J. Sci. Res. 2012, 4, 523. [Google Scholar] [CrossRef]
- Ong, H.S.; Corbett, M.C. Corneal infections in the 21st century. Postgrad. Med. J. 2015, 91, 565–571. [Google Scholar] [CrossRef]
- Ong, H.S.; Fung, S.S.M.; Macleod, D.; Dart, J.K.G.; Tuft, S.J.; Burton, M.J. Altered Patterns of Fungal Keratitis at a London Ophthalmic Referral Hospital: An Eight-Year Retrospective Observational Study. Am. J. Ophthalmol. 2016, 168, 227–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.; Apel, A.; Stapleton, F. Risk factors and causative organisms in microbial keratitis. Cornea 2008, 27, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Ong, H.S.; Sharma, N.; Phee, L.M.; Mehta, J.S. Atypical microbial keratitis. Ocul. Surf. 2021. [CrossRef] [PubMed]
- Aragona, P.; Ferreri, G.; Micali, A.; Puzzolo, D. Morphological changes of the conjunctival epithelium in contact lens wearers evaluated by impression cytology. Eye 1998, 12 Pt 3a, 461–466. [Google Scholar] [CrossRef]
- Larkin, D.F.; Leeming, J.P. Quantitative alterations of the commensal eye bacteria in contact lens wear. Eye 1991, 5 Pt 1, 70–74. [Google Scholar] [CrossRef]
- Stapleton, F.; Willcox, M.D.; Fleming, C.M.; Hickson, S.; Sweeney, D.F.; Holden, B.A. Changes to the ocular biota with time in extended- and daily-wear disposable contact lens use. Infect. Immun. 1995, 63, 4501–4505. [Google Scholar] [CrossRef] [Green Version]
- Sankaridurg, P.R.; Markoulli, M.; de la Jara, P.L.; Harmis, N.; Varghese, T.; Willcox, M.; Holden, B.A. Lid and conjunctival micro biota during contact lens wear in children. Optom. Vis. Sci. 2009, 86, 312–317. [Google Scholar] [CrossRef]
- Iskeleli, G.; Bahar, H.; Eroglu, E.; Torun, M.M.; Ozkan, S. Microbial changes in conjunctival flora with 30-day continuous-wear silicone hydrogel contact lenses. Eye Contact Lens. 2005, 31, 124–126. [Google Scholar] [CrossRef]
- Retuerto, M.A.; Szczotka-Flynn, L.; Mukherjee, P.K.; Debanne, S.; Iyengar, S.K.; Richardson, B.; Cameron, M.; Ghannoum, M.A. Diversity of Ocular Surface Bacterial Microbiome Adherent to Worn Contact Lenses and Bacterial Communities Associated with Care Solution Use. Eye Contact Lens: Sci. Clin. Pr. 2019, 45, 331–339. [Google Scholar] [CrossRef]
- Willcox, M.; Sharma, S.; Naduvilath, T.J.; Sankaridurg, P.R.; Gopinathan, U.; Holden, B.A. External ocular surface and lens microbiota in contact lens wearers with corneal infiltrates during extended wear of hydrogel lenses. Eye Contact Lens. 2011, 37, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Sankaridurg, P.R.; Willcox, M.D.; Sharma, S.; Gopinathan, U.; Janakiraman, D.; Hickson, S.; Vuppala, N.; Sweeney, D.F.; Rao, G.N.; A Holden, B. Haemophilus influenzae adherent to contact lenses associated with production of acute ocular inflammation. J. Clin. Microbiol. 1996, 34, 2426–2431. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Ocular Surface | Studies | Study Population (Eyes) | Genus-Level Ocular Surface Microbiome |
|---|---|---|---|
| Disease | (Year) | ||
| Contact lens wear | Shin et al. [23] (2016) | 9 contact lens wearers | Higher abundance *: |
| 11 non-contact lens wearers | Methylobacterium | ||
| Lactobacillus | |||
| Acinetobacter | |||
| Pseudomonas | |||
| Lower abundance *: | |||
| Haemophilus | |||
| Streptococcus | |||
| Staphylococcus | |||
| Corynebacterium | |||
| Zhang et al. [24] (2017) | 20 OKL wearers | OKL wearers | |
| 22 SCL wearers | Lower abundance *: Bacillus, Tatumella, Lactobacillus | ||
| 25 non-contact lens wearers | |||
| SCL wearers | |||
| Higher abundance *: Elizabethkingia | |||
| Lower abundance *: Delftia | |||
| Dry Eye Disease | Graham et al. [25] (2007) | 57 Non-dry eye disease | Higher abundance *: |
| 34 Dry eye disease | Coagulase negative staphylococcus | ||
| Staphylococcus | |||
| Corynebacterium | |||
| Propionibacterium | |||
| Bacillus | |||
| Lee et al. [26] (2012) | 7 Blepharitis | Higher abundance *: | |
| 4 Healthy controls | Staphylococcus | ||
| Streptophyta | |||
| Corynebacterium | |||
| Enhydrobacter | |||
| Lower abundance *: | |||
| Propionibacterium | |||
| Li et al. [27] (2019) | 54 Non-dry eye disease | Dry eye disease | |
| 35 Dry eye disease | Higher abundance *: | ||
| (25 MGD; 10 non-MGD) | Bacteroidia | ||
| Bacteroidetes | |||
| Lower abundance *: | |||
| Pseudomonas | |||
| Protebacteria | |||
| MGD | |||
| Higher abundance †: | |||
| Bacillus | |||
| Lower abundance †: | |||
| Bacteroidetes | |||
| Dong et al. [28] (2019) | 47 MGD | Higher abundance *: | |
| 42 Healthy controls | Staphylococcus | ||
| Sphingomonas | |||
| Lower abundance *: | |||
| Corynebacterium | |||
| Zhao et al. [29] (2020) | 61 MGD | Higher abundance *: | |
| 15 Healthy controls | Rubrobacter | ||
| Novibacillus | |||
| Campylobacter | |||
| Geobacillus | |||
| Sphingomonas | |||
| Corynebacterium | |||
| Sphingobium | |||
| Pedobacter | |||
| Fictibacillus | |||
| Enterococcus | |||
| Allergic Eye Diseases | Liang et al. [30] (2020) | 21 SAC/PAC | SAC/PAC |
| 18 VKC | Higher abundance ‡: Brevibacterium, Staphylococcus, Hymenobacter, Microbacterium | ||
| VKC | |||
| Higher abundance ‡: Streptococcus, Auricoccus, Actinomyces, Campylobacter, Prevotella, Paracoccus, Atopobium, Candida | |||
| Leonardi et al. [31] (2021) | VKC | Higher abundance *: | |
| Heathy controls | Haemophilus | ||
| Rothia | |||
| Corynebacterium | |||
| Prevotella | |||
| Bacillus | |||
| Cicatrising conjunctivitis | Zilliox et al. [32] (2020) | 12 ocular SJS | Higher abundance *: |
| 6 healthy controls | Staphylococcus | ||
| Corynebacterium | |||
| Streptococcus | |||
| Lactobacillus | |||
| Prevotella | |||
| Fusobacterium | |||
| Enterobacteriaceae | |||
| Kittipibul et al. [33] (2021) | 20 ocular SJS | Higher abundance *: | |
| 20 healthy controls | Bacteroides | ||
| Faecalibacterium | |||
| Salinivibrio | |||
| Akkermansia | |||
| Prevotella | |||
| Coprococcus | |||
| Streptococcus | |||
| Lactobacillus | |||
| Fusobacterium | |||
| Bifidobacterium | |||
| Blautia | |||
| Bacillus | |||
| Phascolarctobacterium | |||
| Paraprevotella | |||
| Acinetobacter | |||
| Ruminococcus | |||
| Megamonas | |||
| Odoribacter | |||
| Staphylococcus | |||
| Pseudoalteromonas | |||
| Erwinia | |||
| Pseudomonas | |||
| Collinsella | |||
| Methanobrevibacter | |||
| Veillonella | |||
| Thermomonas | |||
| Roseburia | |||
| Turicibacter | |||
| Lower abundance *: | |||
| Vibrio | |||
| Acrobacter | |||
| Cetobacterium | |||
| Methylophaga | |||
| Tenacibaculum | |||
| Fusibacter | |||
| Clostridium | |||
| Cohaesibacter | |||
| Shewanella | |||
| Ueta et al. [34] (2021) | 37 ocular SJS | Higher abundance *: | |
| 9 healthy controls | Corynebacterium | ||
| Neisseriaceae | |||
| Staphylococcus | |||
| Propionibacterium | |||
| Streptococcus | |||
| Escherichia | |||
| Fusobacterium | |||
| Lawsonella | |||
| Serratia |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, M.; Tong, L.; Mehta, J.S.; Ong, H.S. Impact of Exposomes on Ocular Surface Diseases. Int. J. Mol. Sci. 2023, 24, 11273. https://doi.org/10.3390/ijms241411273
Hong M, Tong L, Mehta JS, Ong HS. Impact of Exposomes on Ocular Surface Diseases. International Journal of Molecular Sciences. 2023; 24(14):11273. https://doi.org/10.3390/ijms241411273
Chicago/Turabian StyleHong, Merrelynn, Louis Tong, Jodhbir S. Mehta, and Hon Shing Ong. 2023. "Impact of Exposomes on Ocular Surface Diseases" International Journal of Molecular Sciences 24, no. 14: 11273. https://doi.org/10.3390/ijms241411273
APA StyleHong, M., Tong, L., Mehta, J. S., & Ong, H. S. (2023). Impact of Exposomes on Ocular Surface Diseases. International Journal of Molecular Sciences, 24(14), 11273. https://doi.org/10.3390/ijms241411273