
Chronological Changes in the Expression and Localization of Sox9 between Achilles Tendon Injury and Functional Recovery in Mice

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Functional Torque Test
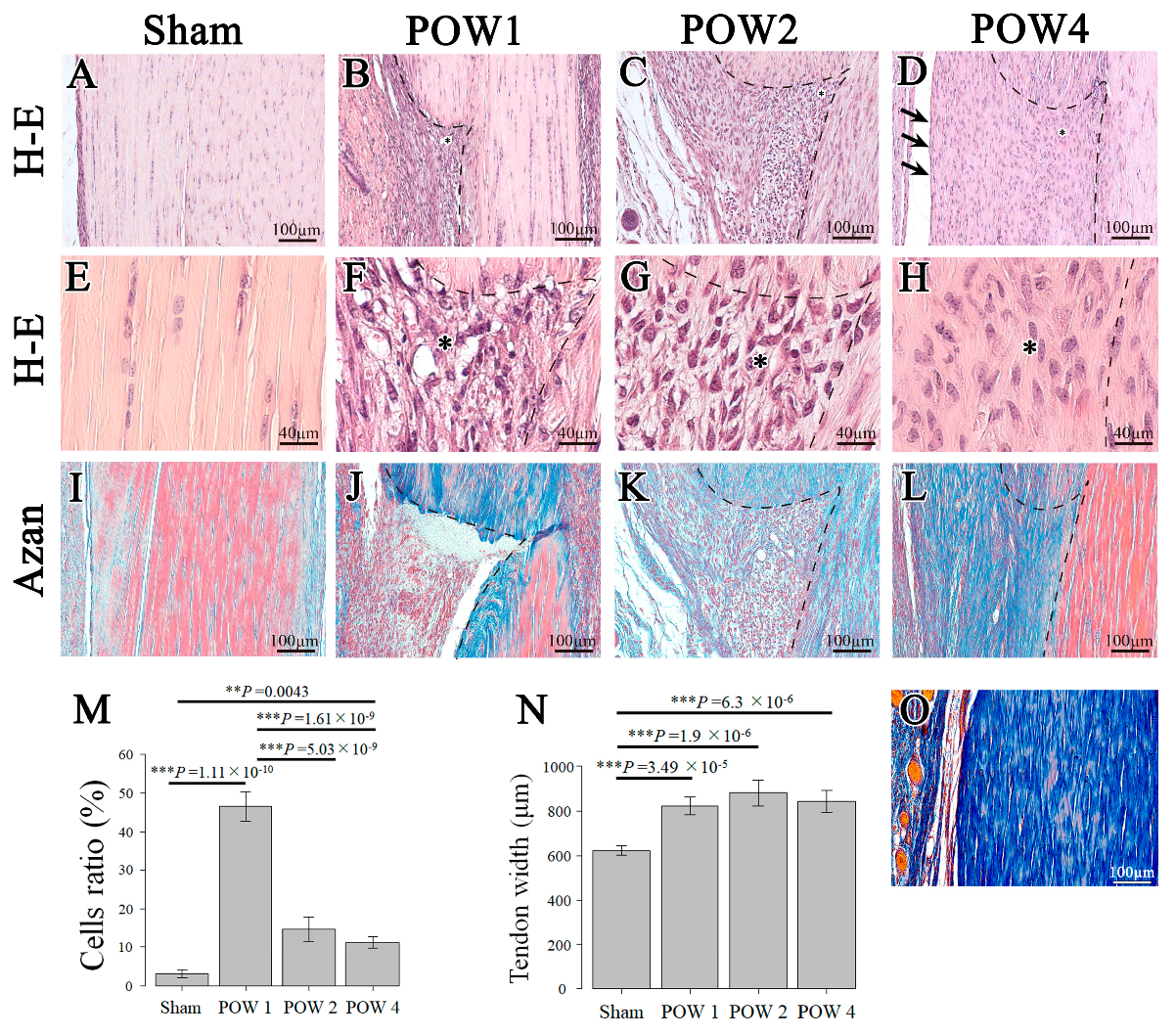
2.2. Morphological Analysis
2.3. mRNA Expression Triggered by Tendon Injury
2.4. Immunohistochemical Staining of Tendon Injury Samples
2.5. Lineage Tracing of Sox9-Expressing Cells
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Tendon Injury Mouse Model
4.3. Physiological Functional Measurements
4.4. Morphological Examination
4.5. RT-PCR
4.6. Immunohistochemical Analysis
4.7. Lineage Tracing Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lantto, I.; Heikkinen, J.; Flinkkilä, T.; Ohtonen, P.; Leppilahti, J. Epidemiology of Achilles tendon ruptures: Increasing incidence over a 33-year period. Scand. J. Med. Sci. Sports 2015, 25, e133–e138. [Google Scholar] [CrossRef] [PubMed]
- Lemme, N.J.; Li, N.Y.; DeFroda, S.F.; Kleiner, J.; Owens, B.D. Epidemiology of achilles tendon ruptures in the united states: Athletic and nonathletic injuries from 2012 to 2016. Orthop. J. Sports Med. 2018, 6, 2325967118808238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Bone and Joint Initiative. The Burden of Musculoskeletal Disease in the United States, 2nd ed.; American Academy of Orthopaedic Surgeons: Rosemont, IL, USA, 2011. [Google Scholar]
- Ceravolo, M.L.; Gaida, J.E.; Keegan, R.J. Quality-of-life in achilles tendinopathy: An exploratory study. Clin. J. Sport Med. 2020, 30, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Nourissat, G.; Berenbaum, F.; Duprez, D. Tendon injury: From biology to tendon repair. Nat. Rev. Rheumatol. 2015, 11, 223–233. [Google Scholar] [CrossRef]
- Lang, J. Über das gleitgewebe der Sehnen, Muskeln, fascien und Gefäße. Zeitschrift fur anatomie und entwicklungsgeschichte. Geshicht 1960, 122, 197–231. [Google Scholar]
- Lang, J. Concerning the blood vessels of the synovial sheath. Acta Anat. 1963, 54, 273–309. [Google Scholar] [CrossRef]
- Jozsa, L.; Kannus, P. Human tendons: Anatomy, physiology and pathology. J. Athl. Train. 1998, 33, 185–186. [Google Scholar]
- Webster, K.E.; Feller, J.A.; Leigh, W.B.; Richmond, A.K. Younger patients are at increased risk for graft rupture and contralateral injury after anterior cruciate ligament reconstruction. Am. J. Sports Med. 2014, 42, 641–647. [Google Scholar] [CrossRef]
- He, P.; Ruan, D.; Huang, Z.; Wang, C.; Xu, Y.; Cai, H.; Liu, H.; Fei, Y.; Heng, B.C.; Chen, W.; et al. Comparison of tendon development versus tendon healing and regeneration. Front. Cell Dev. Biol. 2022, 10, 821667. [Google Scholar] [CrossRef]
- Schweitzer, R.; Chyung, J.H.; Murtaugh, L.C.; Brent, A.E.; Rosen, V.; Olson, E.N.; Lassar, A.; Tabin, C.J. Analysis of the tendon cell fate using scleraxis, a specific marker for tendons and ligaments. Development 2001, 128, 3855–3866. [Google Scholar] [CrossRef]
- Docheva, D.; Hunziker, E.B.; Fässler, R.; Brandau, O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol. Cell Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onizuka, N.; Ito, Y.; Inagawa, M.; Nakahara, H.; Takada, S.; Lotz, M.; Toyama, Y.; Asahara, H. The Mohawk homeobox transcription factor regulates the differentiation of tendons and volar plates. J. Orthop. Sci. 2014, 19, 172–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagakura, R.; Yamamoto, M.; Jeong, J.; Hinata, N.; Katori, Y.; Chang, W.J.; Abe, S. Switching of Sox9 expression during musculoskeletal system development. Sci. Rep. 2020, 10, 8425. [Google Scholar] [CrossRef] [PubMed]
- Soeda, T.; Deng, J.M.; de Crombrugghe, B.D.; Behringer, R.R.; Nakamura, T.; Akiyama, H. Sox9-expressing precursors are the cellular origin of the cruciate ligament of the knee joint and the limb tendons. Genesis 2010, 48, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Sakabe, T.; Sakai, K.; Maeda, T.; Sunaga, A.; Furuta, N.; Schweitzer, R.; Sasaki, T.; Sakai, T. Transcription factor scleraxis vitally contributes to progenitor lineage direction in wound healing of adult tendon in mice. J. Biol. Chem. 2018, 293, 5766–5780. [Google Scholar] [CrossRef] [Green Version]
- Dyment, N.A.; Hagiwara, Y.; Matthews, B.G.; Li, Y.; Kalajzic, I.; Rowe, D.W. Lineage tracing of resident tendon progenitor cells during growth and natural healing. PLoS ONE 2014, 9, e96113. [Google Scholar] [CrossRef] [Green Version]
- Delgado Caceres, M.D.; Angerpointner, K.; Galler, M.; Lin, D.; Michel, P.A.; Brochhausen, C.; Lu, X.; Varadarajan, A.R.; Warfsmann, J.; Stange, R.; et al. Tenomodulin knockout mice exhibit worse late healing outcomes with augmented trauma-induced heterotopic ossification of Achilles tendon. Cell Death Dis. 2021, 12, 1049. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, S.; Luo, D.; He, D.; Shi, C.; Zhu, L.; Guan, B.; Li, Z.; Zhang, T.; Zhou, Y.; et al. Functional regeneration and repair of tendons using biomimetic scaffolds loaded with recombinant periostin. Nat. Commun. 2021, 12, 1293. [Google Scholar] [CrossRef]
- Schwartz, A.G.; Galatz, L.M.; Thomopoulos, S. Enthesis regeneration: A role for Gli1+ progenitor cells. Development 2017, 144, 1159–1164. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.; Lu, H.H.; Schweitzer, R. Molecular regulation of tendon cell fate during development. J. Orthop. Res. 2015, 33, 800–812. [Google Scholar] [CrossRef]
- Shintaku, Y.; Murakami, T.; Yanagita, T.; Kawanabe, N.; Fukunaga, T.; Matsuzaki, K.; Uematsu, S.; Yoshida, Y.; Kamioka, H.; Takano-Yamamoto, T.; et al. Sox9 expression during fracture repair. Cells Tissues Organs 2011, 194, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, S.T.; Serowoky, M.A.; Vakhshori, V.; Tripuraneni, N.; Hegde, N.V.; Lieberman, J.R.; Crump, J.G.; Mariani, F.V. Sox9+ messenger cells orchestrate large-scale skeletal regeneration in the mammalian rib. Elife 2019, 8, e40715. [Google Scholar] [CrossRef]
- Tan, J.; Liu, X.; Zhou, M.; Wang, F.; Ma, L.; Tang, H.; He, G.; Kang, X.; Bian, X.; Tang, K. Effect of treadmill training on fibrocartilage complex repair in tendon-bone insertion healing in the postinflammatory stage. Bone Joint Res. 2023, 12, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Ideo, K.; Tokunaga, T.; Shukunami, C.; Takimoto, A.; Yoshimoto, Y.; Yonemitsu, R.; Karasugi, T.; Mizuta, H.; Hiraki, Y.; Miyamoto, T. Role of Scx+/Sox9+ cells as potential progenitor cells for postnatal supraspinatus enthesis formation and healing after injury in mice. PLoS ONE 2020, 15, e0242286. [Google Scholar] [CrossRef] [PubMed]
- DiFelice, G.S.; List, J.P. Clinical Outcomes of Arthroscopic Primary Repair of Proximal Anterior Cruciate Ligament Tears Are Maintained at Mid-term Follow-up. Arthroscopy 2018, 34, 1085–1093. [Google Scholar] [CrossRef]
- Taylor, S.H.; Al-Youha, S.; Van Agtmael, T.; Lu, Y.; Wong, J.; McGrouther, D.A.; Kadler, K.E. Tendon is covered by a basement membrane epithelium that is required for cell retention and the prevention of adhesion formation. PLoS ONE 2011, 6, e16337. [Google Scholar] [CrossRef] [Green Version]
- Dyment, N.A.; Liu, C.F.; Kazemi, N.; Aschbacher-Smith, L.E.; Kenter, K.; Breidenbach, A.P.; Shearn, J.T.; Wylie, C.; Rowe, D.W.; Butler, D.L. The paratenon contributes to scleraxis-expressing cells during patellar tendon healing. PLoS ONE 2013, 8, e59944. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.; Chien, C.; Bell, R.; Laudier, D.; Tufa, S.F.; Keene, D.R.; Andarawis-Puri, N.; Huang, A.H. Novel model of tendon regeneration reveals distinct cell mechanisms underlying regenerative and fibrotic tendon healing. Sci. Rep. 2017, 7, 45238. [Google Scholar] [CrossRef] [Green Version]
- Weiler, A.; Unterhauser, F.N.; Bail, H.J.; Hüning, M.; Haas, N.P. Alpha-smooth muscle actin is expressed by fibroblastic cells of the ovine anterior cruciate ligament and its free tendon graft during remodeling. J. Orthop. Res. 2002, 20, 310–317. [Google Scholar] [CrossRef]
- Ito, N.; Miyagoe-Suzuki, Y.; Takeda, S.; Kudo, A. Periostin is required for the maintenance of muscle fibers during muscle regeneration. Int. J. Mol. Sci. 2021, 22, 3627. [Google Scholar] [CrossRef]
- Ackerman, J.E.; Studentsova, V.; Myers, M.; Buckley, M.R.; Richards, M.S.; Loiselle, A.E. Non-invasive ultrasound quantification of scar tissue volume identifies early functional changes during tendon healing. J. Orthop. Res. 2019, 37, 2476–2485. [Google Scholar] [CrossRef]
- Lee, J.Y.; Zhou, Z.; Taub, P.J.; Ramcharan, M.; Li, Y.; Akinbiyi, T.; Maharam, E.R.; Leong, D.J.; Laudier, D.M.; Ruike, T.; et al. BMP-12 treatment of adult mesenchymal stem cells in vitro augments tendon-like tissue formation and defect repair in vivo. PLoS ONE 2011, 6, e17531. [Google Scholar] [CrossRef] [PubMed]
- Katzel, E.B.; Wolenski, M.; Loiselle, A.E.; Basile, P.; Flick, L.M.; Langstein, H.N.; Hilton, M.J.; Awad, H.A.; Hammert, W.C.; O’Keefe, R.J. Impact of Smad3 loss of function on scarring and adhesion formation during tendon healing. J. Orthop. Res. 2011, 29, 684–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Galatz, L.M.; Das, R.; Havlioglu, N.; Rothermich, S.Y.; Thomopoulos, S. The role of transforming growth factor beta isoforms in tendon-to-bone healing. Connect. Tissue Res. 2011, 52, 87–98. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Takimoto, A.; Akiyama, H.; Kist, R.; Scherer, G.; Nakamura, T.; Hiraki, Y.; Shukunami, C. Scx+/Sox9+ progenitors contribute to the establishment of the junction between cartilage and tendon/ligament. Development 2013, 140, 2280–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blitz, E.; Sharir, A.; Akiyama, H.; Zelzer, E.I. Tendon-bone attachment unit is formed modularly by a distinct pool of Scx- and Sox9-positive progenitors. Development 2013, 140, 2680–2690. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, J.E.; Best, K.T.; Muscat, S.N.; Pritchett, E.M.; Nichols, A.E.C.; Wu, C.L.; Loiselle, A.E. Defining the spatial-molecular map of fibrotic tendon healing and the drivers of scleraxis-lineage cell fate and function. Cell Rep. 2022, 41, 111706. [Google Scholar] [CrossRef]
- Ishizuka, S.; Yamamoto, M.; Hirouchi, H.; Yotsuya, M.; Ohkubo, M.; Sato, M.; Abe, S. Muscle-bone relationship in temporo mandibular joint disorders after partial discectomy. J. Oral. Biosci. 2021, 63, 436–443. [Google Scholar] [CrossRef]
- Abe, S.; Yamamoto, M. Factors involved in morphogenesis in the muscle-tendon-bone complex. Int. J. Mol. Sci. 2021, 22, 6365. [Google Scholar] [CrossRef]
- Yamamoto, M.; Sakiyama, K.; Kitamura, K.; Yamamoto, Y.; Takagi, T.; Sekiya, S.; Watanabe, G.; Taniguchi, S.; Ogawa, Y.; Ishizuka, S.; et al. Development and Regeneration of Muscle, Tendon, and Myotendinous Junctions in Striated Skeletal Muscle. Int. J. Mol. Sci. 2022, 23, 3006. [Google Scholar] [CrossRef]
- Kanehira, C.; Yamamoto, M.; Hirouchi, H.; Ishuzuka, S.; Sakiyama, K.; Higa, K.; Murakami, G.; Abe, S. Tendinous annulus of zinn for a common origin of the extraocular rectus muscles: A histological study of the orbital apex from donated elderly cadavers. Anat. Sci. Int. 2022, 97, 369–379. [Google Scholar] [CrossRef]
- Takimoto, A.; Oro, M.; Hiraki, Y.; Shukunami, C. Direct conversion of tenocytes into chondrocytes by Sox9. Exp. Cell Res. 2012, 318, 1492–1507. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ito, Y.; Shinohara, M.; Yamashita, S.; Ichinose, S.; Kishida, A.; Oyaizu, T.; Kayama, T.; Nakamichi, R.; Koda, N.; et al. Gene targeting of the transcription factor Mohawk in rats causes heterotopic ossification of Achilles tendon via failed tenogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, 7840–7845. [Google Scholar] [CrossRef] [PubMed]
- Wren, T.A.; Beaupré, G.S.; Carter, D.R. Mechanobiology of tendon adaptation to compressive loading through fibrocartilaginous metaplasia. J. Rehabil. Res. Dev. 2000, 37, 135–143. [Google Scholar] [PubMed]
- Gillard, G.C.; Reilly, H.C.; Bell-Booth, P.G.; Flint, M.H. The influence of mechanical forces on the glycosaminoglycan content of the rabbit flexor digitorum profundus tendon. Connect. Tissue Res. 1979, 7, 37–46. [Google Scholar] [CrossRef]
- He, X.; Bougioukli, S.; Ortega, B.; Arevalo, E.; Lieberman, J.R.; McMahon, A.P. Sox9 positive periosteal cells in fracture repair of the adult mammalian long bone. Bone 2017, 103, 12–19. [Google Scholar] [CrossRef]
- Itoh, Y.; Murakami, T.; Mori, T.; Agata, N.; Kimura, N.; Inoue-Miyazu, M.; Hayakawa, K.; Hirano, T.; Sokabe, M.; Kawakami, K. Training at non-damaging intensities facilitates recovery from muscle atrophy. Muscle Nerve 2017, 55, 243–253. [Google Scholar] [CrossRef]
- Walia, B.; Huang, A.H. Tendon stem progenitor cells: Understanding the biology to inform therapeutic strategies for tendon repair. J. Orthop. Res. 2019, 37, 1270–1280. [Google Scholar] [CrossRef]
- Kawamoto, T. Use of a new adhesive film for the preparation of multi-purpose fresh-frozen sections from hard tissues, whole-animals, insects and plants. Arch. Histol. Cytol. 2003, 66, 123–143. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Assay ID | Amplicon Length | |
|---|---|---|---|
| Sox9 | transcription factor for differentiated chondrocyte and tendon cells | Mm00448840_m1 | 101 |
| α-SMA | protein belonged actin, a marker of epitenon important during tendon regeneration | Mm00725412_s1 | 95 |
| Postn | protein expressed in the extracellular matrix that supports TSPC growth and tendon formation | Mm00450111_m1 | 79 |
| Scx | transcription factor for differentiated tendon cells | Mm01205675_m1 | 59 |
| Col1a1 | fibroplastic collagen abundant in tendons | Mm00801666_g1 | 89 |
| Col3a1 | early differentiation marker for regenerate injured tendons | Mm00802305_g1 | 80 |
| Fmod | small leucine-rich proteoglycan for regulation in collagen matrix assembly | Mm00491215_m1 | 75 |
| GAPDH | housekeeping genes expressed in most cells are used as an internal control | Mm99999915_g1 | 107 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, G.; Yamamoto, M.; Taniguchi, S.; Sugiyama, Y.; Hirouchi, H.; Ishizuka, S.; Kitamura, K.; Mizoguchi, T.; Takayama, T.; Hayashi, K.; et al. Chronological Changes in the Expression and Localization of Sox9 between Achilles Tendon Injury and Functional Recovery in Mice. Int. J. Mol. Sci. 2023, 24, 11305. https://doi.org/10.3390/ijms241411305
Watanabe G, Yamamoto M, Taniguchi S, Sugiyama Y, Hirouchi H, Ishizuka S, Kitamura K, Mizoguchi T, Takayama T, Hayashi K, et al. Chronological Changes in the Expression and Localization of Sox9 between Achilles Tendon Injury and Functional Recovery in Mice. International Journal of Molecular Sciences. 2023; 24(14):11305. https://doi.org/10.3390/ijms241411305
Chicago/Turabian StyleWatanabe, Genji, Masahito Yamamoto, Shuichirou Taniguchi, Yuki Sugiyama, Hidetomo Hirouchi, Satoshi Ishizuka, Kei Kitamura, Toshihide Mizoguchi, Takashi Takayama, Katsuhiko Hayashi, and et al. 2023. "Chronological Changes in the Expression and Localization of Sox9 between Achilles Tendon Injury and Functional Recovery in Mice" International Journal of Molecular Sciences 24, no. 14: 11305. https://doi.org/10.3390/ijms241411305
APA StyleWatanabe, G., Yamamoto, M., Taniguchi, S., Sugiyama, Y., Hirouchi, H., Ishizuka, S., Kitamura, K., Mizoguchi, T., Takayama, T., Hayashi, K., & Abe, S. (2023). Chronological Changes in the Expression and Localization of Sox9 between Achilles Tendon Injury and Functional Recovery in Mice. International Journal of Molecular Sciences, 24(14), 11305. https://doi.org/10.3390/ijms241411305