
The Role and Mechanism of MicroRNA 21 in Osteogenesis: An Update
 , , and
, , and
Abstract
:1. Introduction
1.1. Bone Remodeling and Homeostasis
1.2. The Coupling Mechanism between Osteoblasts and Osteoclasts
1.3. Role of MicroRNA in Bone Homeostasis
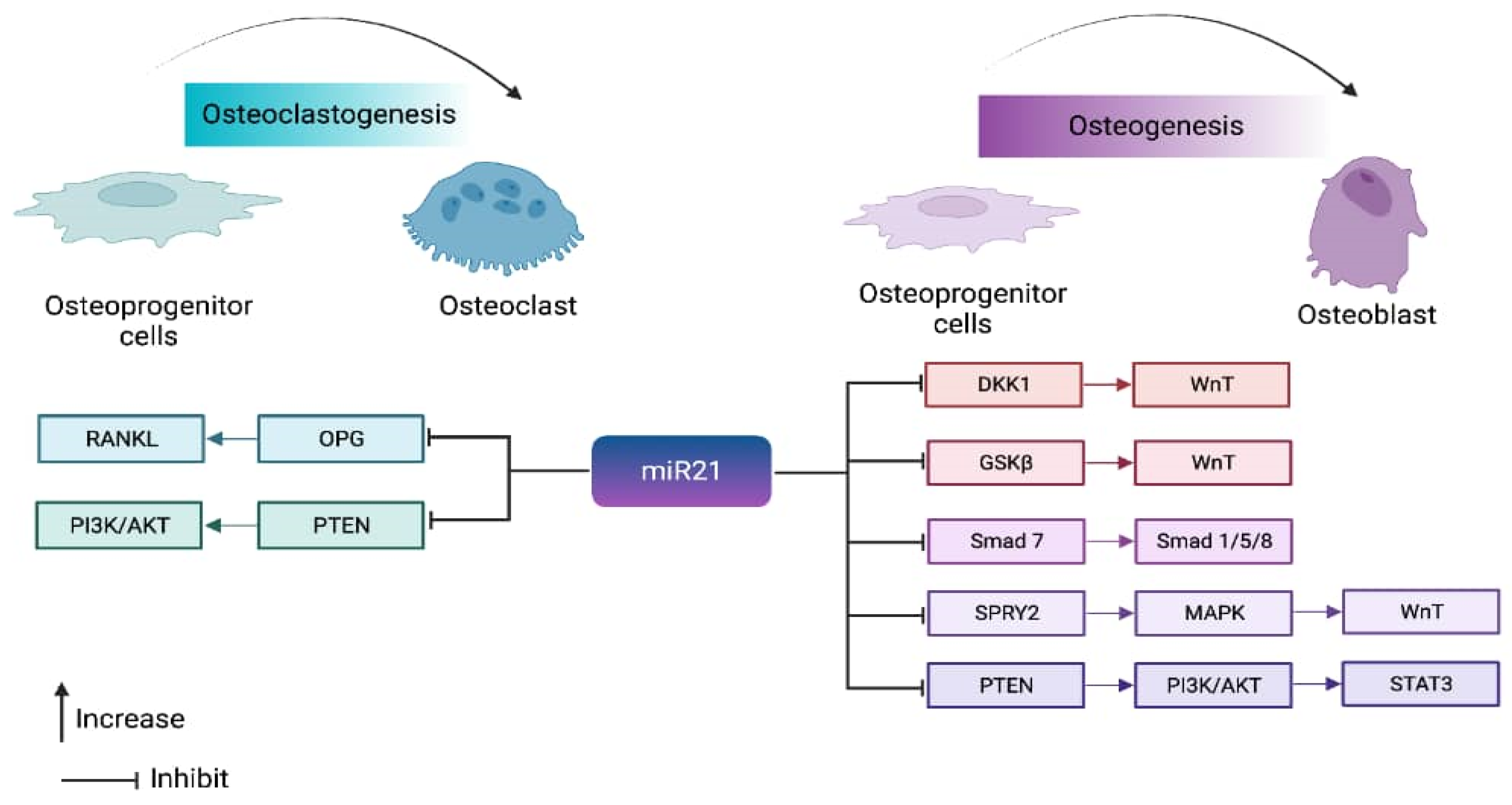
2. Role of MicroRNA 21 in Bone Homeostasis
3. Empirical Evidence of MicroRNA-21 in Regulating Bone Homeostasis
3.1. In Vitro Studies
3.2. In Vivo Studies
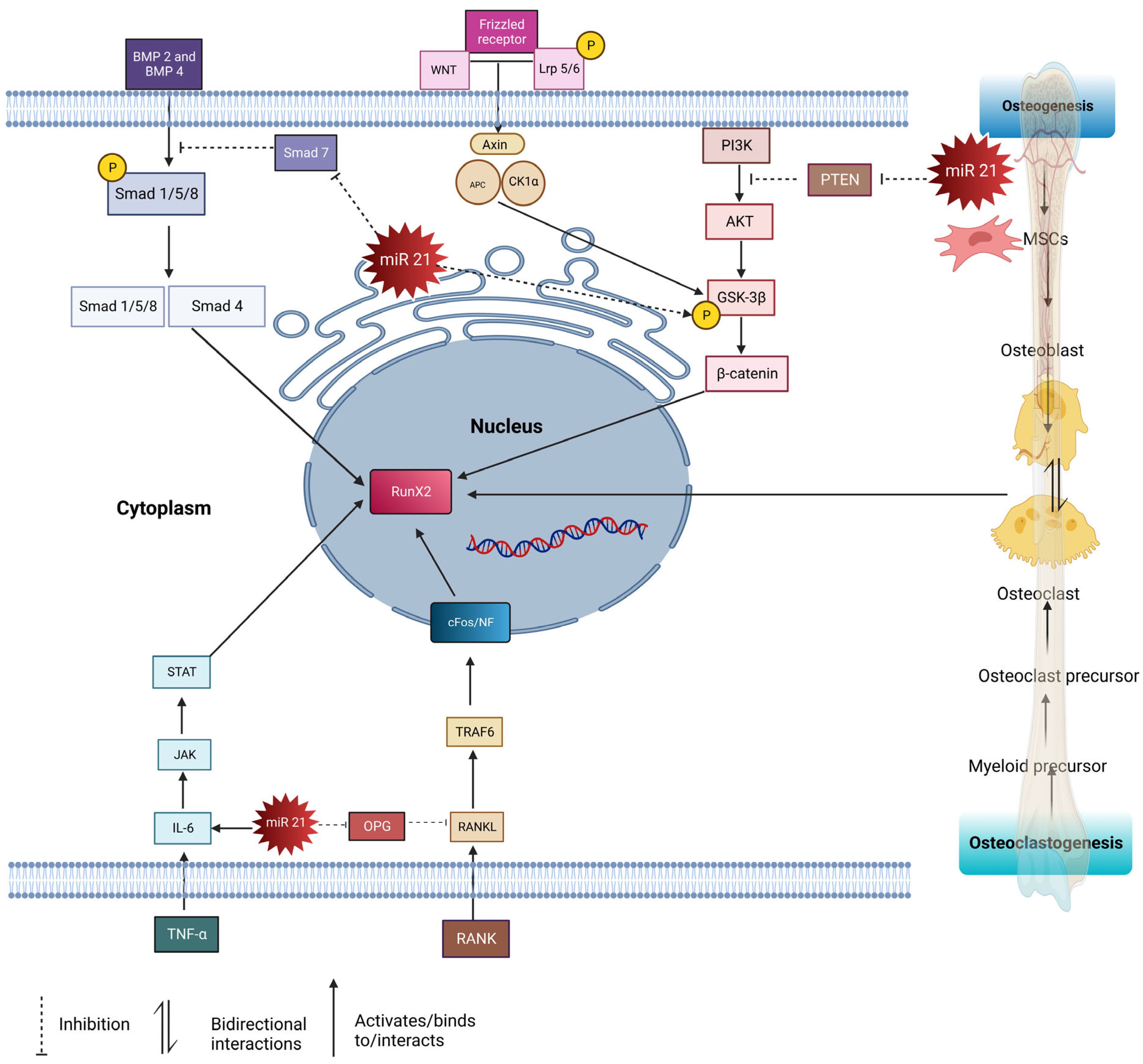
4. Pathways Regulated by miR-21 in Bone Homeostasis
4.1. Smad Pathway
4.2. RANKL/RANK/OPG Pathway
4.3. STAT3 Pathway
4.4. PTEN-PI3K/Akt Signaling Pathway
4.5. WNT/β- Catenin Pathway
5. miR21 in Therapeutic Applications
6. miR-21 in the Coupling Mechanism
7. Conclusions
8. Knowledge Gap and Future Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Rooij, E. The Art of MicroRNA Research. Circ. Res. 2011, 108, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.R.; Burr, D.B. Bone Modeling and Remodeling. In Basic and Applied Bone Biology; Academic Press: Cambridge, MA, USA, 2013; pp. 75–90. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. The RANKL/RANK/OPG Pathway. Epidemiol. Pathophysiol. Accessory 2007, 5, 98–194. [Google Scholar] [CrossRef]
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Kenkre, J.S.; Bassett, J.H.D. The bone remodelling cycle. Ann. Clin. Biochem. 2018, 55, 308–327. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. 2020. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Zhao, C.; Irie, N.; Takada, Y.; Shimoda, K.; Miyamoto, T.; Nishiwaki, T.; Suda, T.; Matsuo, K. Bidirectional ephrinB2-EphB4 signaling controls bone homeostasis. Cell Metab. 2006, 4, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Liu, Z.; Chen, Y. Regulation of TGF- b signaling by Smad7 Overview of TGF- b Signaling Pathways. Acta Biochim. Biophys. Sin. 2009, 41, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, M.; Nakashima, T.; Taniguchi, M.; Kodama, T.; Kumanogoh, A.; Takayanagi, H. Osteoprotection by semaphorin 3A. Nature 2012, 485, 69–74. [Google Scholar] [CrossRef]
- Mizuno, A.; Kanno, T.; Hoshi, M.; Shibata, O.; Yano, K.; Fujise, N.; Kinosaki, M.; Yamaguchi, K.; Tsuda, E.; Murakami, A.; et al. Transgenic mice overexpressing soluble osteoclast differentiation factor (sODF) exhibit severe osteoporosis. J. Bone Miner. Metab. 2002, 20, 337–344. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-Osteoclast Interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef]
- Sriram, M.; Sainitya, R.; Kalyanaraman, V.; Dhivya, S.; Selvamurugan, N. Biomaterials mediated microRNA delivery for bone tissue engineering. Int. J. Biol. Macromol. 2015, 74, 404–412. [Google Scholar] [CrossRef]
- Wildman, B.J.; Godfrey, T.C.; Rehan, M.; Chen, Y.; Afreen, L.H.; Hassan, Q. MICROmanagement of Runx2 Function in Skeletal Cells. Curr. Mol. Biol. Rep. 2019, 5, 55–64. [Google Scholar] [CrossRef]
- Mitchelson, K.R. Roles of the canonical myomiRs miR-1, -133 and -206 in cell development and disease. World J. Biol. Chem. 2015, 6, 162–208. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, X.; Qi, H.; Hou, X.; Zhao, J.; Yuan, X.; Ma, X. MiR-21 nanocapsules promote early bone repair of osteoporotic fractures by stimulating the osteogenic differentiation of bone marrow mesenchymal stem cells. J. Orthop. Transl. 2020, 24, 76–87. [Google Scholar] [CrossRef]
- Li, X.; Guo, L.; Liu, Y.; Su, Y.; Xie, Y.; Du, J.; Zhou, J.; Ding, G.; Wang, H.; Bai, Y.; et al. MicroRNA-21 promotes osteogenesis of bone marrow mesenchymal stem cells via the Smad7-Smad1/5/8-Runx2 pathway. Biochem. Biophys. Res. Commun. 2017, 493, 928–933. [Google Scholar] [CrossRef]
- Shimazu, J.; Wei, J.; Karsenty, G. Smurf1 Inhibits Osteoblast Differentiation, Bone Formation, and Glucose Homeostasis. Cell Rep. 2016, 15, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Kumarswamy, R.; Volkmann, I.; Thum, T. Regulation and function of miRNA-21 in health and disease. RNA Biol. 2011, 8, 706–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.; Tran, N. Global miRNA to miRNA Interactions: Impacts for miR-21. Trends Cell Biol. 2021, 31, 3–5. [Google Scholar] [CrossRef]
- Meng, Y.-B.; Li, X.; Li, Z.-Y.; Zhao, J.; Yuan, X.-B.; Ren, Y.; Cui, Z.-D.; Liu, Y.-D.; Yang, X.-J. microRNA-21 promotes osteogenic differentiation of mesenchymal stem cells by the PI3K/β-catenin pathway. J. Orthop. Res. 2015, 33, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, F.; Wang, Z.; Fu, Q.; Liang, A. MicroRNA-21 promotes osteogenic differentiation by targeting small mothers against decapentaplegic 7. Mol. Med. Rep. 2012, 12, 1561–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheedy, F.J. Turning 21: Induction of miR-21 as a Key Switch in the Inflammatory Response. Front. Immunol. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.-H.; Tsao, C.-J. Emerging role of microRNA-21 in cancer (Review). Biomed. Rep. 2016, 5, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Van Wijnen, A.J.; van de Peppel, J.; van Leeuwen, J.P.; Lian, J.B.; Stein, G.S.; Westendorf, J.J.; Oursler, M.-J.; Im, H.-J.; Taipaleenmäki, H.; Hesse, E.; et al. MicroRNA Functions in Osteogenesis and Dysfunctions in Osteoporosis. Curr. Osteoporos. Rep. 2013, 11, 72–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Xiao, J.; Zheng, K.; Feng, X.; Zhang, J.; Song, D.; Wang, C.; Shen, X.; Zhao, X.; Wei, C.; et al. MiR-21/STAT3 Signal Is Involved in Odontoblast Differentiation of Human Dental Pulp Stem Cells Mediated by TNF-α. Cell. Reprogram. 2018, 20, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, Z.; Gu, X.; Jin, Y.; Feng, C.; Yang, S.; Wei, F. MicroRNA-21 affects mechanical force–induced midpalatal suture remodelling. Cell Prolif. 2020, 53, e12697. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, Z.; Wang, J.; Ji, X.; Yao, Z.; Wang, X. miR-21 promotes osteoclastogenesis through activation of PI3K/Akt signaling by targeting Pten in RAW264.7 cells. Mol. Med. Rep. 2020, 21, 1125–1132. [Google Scholar] [CrossRef] [Green Version]
- Geng, Z.; Wang, X.; Zhao, J.; Li, Z.; Ma, L.; Zhu, S.; Liang, Y.; Cui, Z.; He, H.; Yang, X. The synergistic effect of strontium-substituted hydroxyapatite and microRNA-21 on improving bone remodeling and osseointegration. Biomater. Sci. 2018, 6, 2694–2703. [Google Scholar] [CrossRef]
- Yang, C.; Liu, X.; Zhao, K.; Zhu, Y.; Hu, B.; Zhou, Y.; Wang, M.; Wu, Y.; Zhang, C.; Xu, J.; et al. miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1α pathway and enhances bone regeneration in critical size defects. Stem Cell Res. Ther. 2019, 10, 65. [Google Scholar] [CrossRef]
- Wu, L.; Su, Y.; Lin, F.; Zhu, S.; Wang, J.; Hou, Y.; Du, J.; Liu, Y.; Guo, L. MicroRNA-21 promotes orthodontic tooth movement by modulating the RANKL/OPG balance in T cells. Oral Dis. 2020, 26, 370–380. [Google Scholar] [CrossRef]
- Smieszek, A.; Marcinkowska, K.; Pielok, A.; Sikora, M. The Role of miR-21 in Osteoblasts—Osteoclasts. Cells 2020, 9, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.-L.; Chen, Z.-H.; Teng, Y.-Y.; Liu, S.-Y.; Jia, Y.; Zhang, K.-W.; Sun, Z.-L.; Wu, J.-J.; Yuan, Z.-D.; Feng, Y.; et al. The Smad Dependent TGF-β and BMP Signaling Pathway in Bone Remodeling and Therapies. Front. Mol. Biosci. 2021, 8, 593310. [Google Scholar] [CrossRef] [PubMed]
- Al-Bari, A.A.; Al Mamun, A. Current advances in regulation of bone homeostasis. FASEB BioAdvance 2020, 2, 668–679. [Google Scholar] [CrossRef]
- Sanpaolo, E.R.; Rotondo, C.; Cici, D.; Corrado, A.; Cantatore, F.P. JAK/STAT pathway and molecular mechanism in bone remodeling. Mol. Biol. Rep. 2020, 47, 9087–9096. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Yang, L.; Li, C.; Qing, W. Role of Inflammatory Factors in Regulation of Osteogenesis in Tissue-Engineered Bone. In Osteogenesis and Bone Regeneration; IntechOpen: Rijeka, Croatia, 2019. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Song, Y.; Xiong, Y.; Wang, X.; Xu, K.; Han, B.; Bai, Y.; Liming, Z.; Zhang, Y.; Zhou, L. MicroRNA-21 (Mir-21) Promotes Cell Growth and Invasion by Repressing Tumor Suppressor PTEN in Colorectal Cancer. Cell. Physiol. Biochem. 2017, 43, 945–958. [Google Scholar] [CrossRef]
- Blanco-Aparicio, C.; Renner, O.; Leal, J.F.; Carnero, A. PTEN, more than the AKT pathway. Carcinogenesis 2007, 28, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Bisson, S.-K.; Ung, R.-V.; Mac-Way, F. Role of the Wnt/β-Catenin Pathway in Renal Osteodystrophy. Int. J. Endocrinol. 2018, 2018, 5893514. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Xu, L.; Yang, B.; Ai, J. MicroRNA transport: A new way in cell communication. J. Cell. Physiol. 2013, 228, 1713–1719. [Google Scholar] [CrossRef]
- Huang, C.; Geng, J.; Jiang, S. MicroRNAs in regulation of osteogenic differentiation of mesenchymal stem cells. Cell Tissue Res. 2016, 368, 229–238. [Google Scholar] [CrossRef]
- Kong, L.; Smith, W.; Hao, D. Overview of RAW264.7 for osteoclastogensis study: Phenotype and stimuli. J. Cell. Mol. Med. 2019, 23, 3077–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugatani, T.; Vacher, J.; Hruska, K.A. A microRNA expression signature of osteoclastogenesis. Blood 2011, 117, 3648–3657. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reference | Aim | In Vitro Study | In Vivo Study | Carrier | Findings | Signaling Molecules/ Pathways | Conclusion |
|---|---|---|---|---|---|---|---|
| Li et al., 2017 [16] | To investigate the effects of miR21 on the osteogenic differentiation of BMMSCs and the signaling pathways that influence the biological behaviors of BMMSCs | BMMSCs from the bone cavities of wild-type (WT) and microRNA-21 knock-out (miR21-KO) mice. These BMSCs were osteoinduced and assessed for miR21 expression. BMSCs from WT mice were transfected with exogenous miR21 mimic. All cells were then analyzed through RT-PCR, alizarin and ALP staining, and Western blot for miR21 expression. | 5-week-old female C57BL/6J wild and knock-out mice. n = 5 (20–22 g). The calvarial bone defect model was bilaterally established in the WT and miR21-KO mice. The defective bones were analyzed after 2 months using a stereomicroscope and H&E staining. BMMSCs of WT were transfected with miR21, and BMMSCs of miR21-KO mice were transfected with Smad7 siRNA. | 1. RT-PCR and Western blot analyses showed that the expression of key transcription factors that regulate osteogenesis were significantly upregulated in the WT mice BMSCs compared to miR21-KO mice BMSCs. 2. BMMSCs of WT mice transfected with exogeneous miR21 resulted in enhanced mineralized nodule formation and increased osteogenic marker expressions. BMMSCs from miR21-KO mice showed much lower ALP activity compared to those from WT mice. 3. Western blot showed increased expression of ALP, Runx2, and Smad1/5/8 expression in BMMSCs of miR21-KO mice transfected with Smad7 siRNA. | miR21 directly inhibits Smad7, upregulates the Smad1/5/8-Runx2 pathway, and promotes osteogenesis. | miR21 is indispensable in the osteogenic differentiation of BMMSCs, and miR21 regulates osteoblastic bone via the Smad7-Smad1/5/8-Runx2 pathway. | |
| Xu et al., 2018 [26] | To analyze the regulation of miR21/STAT3 and odontoblast differentiation of DPSCs in the inflammatory microenvironment induced by TNF-alpha. | Human-impacted third molars collected from patients (18–28 years of age). n = 15. DPSC cells from 3rd passage were treated in a TNF-alpha induced inflammatory microenvironment (1–100 ng/mL). Cells were also transfected with STAT3 siRNA, control siRNA, and miRNA inhibitor as (−) control. MTT assay, ALP and alizarin staining, Western blot, and RT- PCR were carried out. | NIL | 1. The expressions of (dentin matrix acidic phosphoprotein) DMP1 and (dental sialophosphoprotein) DSPP, stimulated by TNF-alpha, at low concentrations (10 ng/mL). 2. Increased expression of STAT3 and miR21 was observed in low concentrations of TNF-alpha. 3. PCR showed increased DMP1, DSPP, miR21, and STAT3 in the nucleus due to stimulation by low concentrations of TNF-alpha. 4. Chromatin was precipitated with STAT3 antibodies and was significantly enriched for the miR21 promoter sequence. 5. Expressions of DMP1 and DSPP significantly decreased when transfected with siSTAT3 or treated with Cucurbitacin I (Cuc I). Besides that, pri miR21 also increased under this condition. This shows STAT3 is directly involved in the expression of miR21 during odontoblast differentiation. | In the TNF-alpha-induced microenvironment, STAT3 protein became activated (p-STAT3) and increased bone morphogenetic protein receptor 2 (BMPR2) expression by stimulating the maturation of miR21. | STAT3 was activated by TNF-alpha, stimulated miR21 maturation in DPSCs, and mediated odontoblast differentiation. | |
| M. Li et al., 2020 [27] | To elucidate the effects of miR-21 on the mid-palatal suture-induced bone remodeling by expanding the palatal suture. | BMSCs from miR-21−/− and WT mice were cultured Cell migration and cytotoxicity assays were carried out | Wild-type and KO C57BL/6 mice. 6 weeks old, male. 19–21 g. They were used to establish animal models of rapid maxillary expansion (RME). 4 groups were established: (i) WT (control); (ii) WT + RME; (iii) miR21-/1; (iv) miR21−/− + RME. The maxillae were harvested by euthanizing 3 mice every 1, 3, 7, 14, and 28 days. (PCR) genotyping for WT and miR21−/− mice was performed. miR-21−/− mice with expansion force received agomir-21, and controls were treated with PBS (0.2 mL) through intraperitoneal injection every 3 days for 2 consecutive weeks Histology, histochemical, and immunohistochemical studies were conducted. | 1. Histological analysis showed new bone formation in WT mice to be more rapid than in the miR-21−/− mice. 2. Alizarin and Calcein complex markers showed stronger expression in WT mice compared to miR21−/− mice 3. Expression of ALP and OCN revealed increased expression in WT mice compared to miR-21−/− mice 4. TRAP staining indicated increased RANKl and decreased OPG expression in miR21−/− (RME) mice. 5. Increased osteoprotegerin (Opg) expression and decreased Rankl expression in miR21−/− mice were observed using immunohistochemical staining and immunofluorescence. 6. miR21−/− mice received agomir-21, and PCR showed increased expression of ALP, OCN, and OPG to RANK ratio. | miR-21 affected bone remodeling by coupling reaction through RANKL/OPG. miR-21 regulated the proliferation and migration of BMSCs and bone formation during RME in WT mice. | Findings showed that overexpression of miR21 can promote osteogenic differentiation and accelerate fracture healing. | |
| Wang et al., 2020 [28] | To investigate the effects of miR21 on osteoclastogenesis and bone resorption, as well as the potential molecular mechanisms underlying the Pten-PI3K/Akt signaling pathway in osteoclasts. | The murine macrophage cell line RA W264.7 was cultured with RANKL to induce osteoclastogenesis. RA W264.7 cells were transfected with: miR NC; miR21 mimic; miR21 inhibitor; miR21 mimic + LY29400 (P13K inhibitor). Trap staining, Western Blot, and RT PCR were carried out. Bone resorption assay: Bovine bone slices were treated with the transfected cells/controls, and the bone resorption was calculated as the pit area/total bone area of each slice. | 1. TRAP analysis indicated that miR21 was progressively upregulated during osteoclastogenesis in vitro. 2. RT-PCR Western blot analyses showed decreased Pten protein expression in the miR21 mimic group, thus suggesting that miR21 negatively regulated PTEN. A high-level miR21 was detected during osteoclastogenesis. 3. The percentage of bone resorption was significantly higher in the miR21 mimic group compared to NC and inhibitor. 4. The miR21 mimic group decreased the protein expression levels of Pten, but upregulated p-A kt and NFATc1 expression. 5. The bone resorption assay resulted as significantly higher in the mimic group compared to the other groups. | PTEN regulates osteoclastogenesis in the RANKL-induced RA W264.7 osteoclast cell line by activating the P13K/Akt signaling pathway. | miR 21 was upregulated in osteoclastogenesis and promoted bone resorption by activating the PI3K/Akt signaling pathway via targeting PTEN. | ||
| Geng et al., 2018 [29] | To evaluate biodegradable SrHA and miR21 composite coatings on Ti implants, and the effects on bone formation and osseointegration by promoting osteogenesis and inhibiting osteoclastic activity. | MG63 osteoblast-like cell line was used Cell morphology, density, cell proliferation, ALP activity, and cell distribution were studied | Mature New Zealand white rabbits. n = 30. 2.5–3 kg. Rabbits were implanted with four of the Tis coated with SrHA and miR21 rods at the distal femur and tibial plateau of the hind legs | miR21 was encapsulated in acrylamide polymer. Ti-SrHA was synthesized miR21 was coated on Ti-SrHA Encapsulated miR21 was observed under TEM Ti-SrHA-21 was observed through confocal imaging and X-ray photoelectron spectroscopy (XPS) | 1. The miR21 nanocapsules, synthesized by an in situ polymerization method, showed a spherical structure with a uniform diameter of ~30 nm. 2. Confocal images suggested that the miR21 nanocapsules were distributed evenly on the surfaces of Ti-21 and Ti-SrHA-21. 3. H&E and Masson Trichome staining results indicated that SrHA and miR21 had synergistic effects on bone remodeling and new bone mineralization. 4. Fluorescence microscopy images of the MG63 cells incorporated with Ti-SrHA-21 exhibited the best cell-spreading property and highest cell density compared to other groups. 5. Ti-SrHA-21 significantly increased the CD31 expression in the early stages of surgery. CD31 is an important endothelial marker that could contribute to the development of new blood vessels and, thus, plays an important role in bone formation and increased osteogenesis-related gene expression (including COL-I, RUNX2, OCN, and OPN). 6. The X-ray and CT scan showed that all samples exhibited good osseointegration, especially Ti-SrHA-21, which showed remarkable osteoconductivity and osteoinductivity. 7. Raman spectra results indicated that the degree of new bone mineralization increased with the healing time. | miR21 promotes stem cells’ survival and osteogenesis, presumably via the Smad7-Smad1/5/8-Runx2 and Akt pathways. | SrHA and miR21 synergistically promote angiogenesis, osteogenesis, anti-osteoclastic, osseointegration, bone mineralization, and bone–implant bonding strength. |
| Yang et al., 2019 [30] | To investigate the role of miR21 in promoting osteogenesis in bone marrow-derived stem cells (BMSCs) in vitro. | BMMSCs from Labrador dogs (approximately 2 years old). BMSCs were transfected with miR21 mimic, miR21 inhibitor, or Lenti-miR21(LacZ) using oligofectamine. | Rat. n = 18. A calvarial bone-defect rat model was created. Lenti-miR21/β-TCP/BMSC scaffolds in a canine mandibular defect model. | Carrier Lenti-miR21(LacZ), miR21 mimic, and inhibitor-transfected BMSCs were seeded into β-TCP scaffolds (200 μL per scaffold, at a density of 1.0 × 107 cells/mL) | 1. RT-PCR for BMSCs infected with Lenti-miR21 showed that BMP-2, Runx2, OCN, and OPN mRNA levels increased. 2. Western blotting revealed that HIF-1α, Akt, and PI3K protein levels increased nearly twofold after miR21 mimic transfection, and, in contrast PTEN decreased. 3. Histological examination showed that more new bone formed in the Lenti-miR21/β-TCP BMSCs scaffold implantation group compared with the other two scaffolds (Lenti-LacZ β-TCP and Lenti-LacZ β-TCP BMSC) 4. Micro-CT images revealed new calvarial bone formation in the Lenti miR21/β-TCP/BMSC scaffold group. 5. Gieson’s staining showed that bone formation was observed in the Lenti-miR21/β-TCP/BMSC group, and less in the Lenti-LacZ/β-TCP/BMSC group. 6. Bone formation and mineralization were assessed histomorphometrically via tetracycline, calcein, and ARS fluorescence. Data indicated that Lenti-miR21 enhanced ossification in the induced BMSCs, and effectively promoted new bone formation. | miR21 overexpression increased BMSC migration and HIF-1α activity via the PTEN/PI3K/Akt pathway. | miRNA-21 overexpression in BMSCs may offer great therapeutic promise for rapid bone formation during the healing process by modulation of the osteogenesis process via the PTEN/P13K pathway. |
| Wu et al., 2019 [31] | To investigate the effects of microRNA-21 (miR21) on orthodontic tooth movement (OTM). | CD4+ T cells were purified from the spleenocytes of WT or miR21−/− mice. CD4+ T cells from WT mice were transplanted into miR21−/− mice. ELISA, Western blot, and RT-PCR analyses were carried out. | WT C57BL and miR21−/− mice. Male. 20–25 g 8 weeks old n = 6 An orthodontic tooth movement model was established in C57BL mice. The mice were sacrificed on the 14th day after this procedure, after which the maxillae were removed and the blood was collected by retro-orbital bleeding. Micro-computed tomography was used to measure the distance of tooth movement. Histological, histomorphometric, and immunohistochemical analyses were carried out. | 1. Micro-CT scan-indicated OTM distance of miR21−/− mice was markedly less than that of WT mice. 2. H&E staining showed that the width of the periodontal ligament on the pressure side of the WT was lower than that in the miR21−/− mice. 3. The serum concentration of RANKL in miR21−/− mice was less compared to WT mice. 4. RT-PCR demonstrated that RANKL was downregulated in miR-21−/− mice, and WT mice induced the secretion of RANKL in activated T cells. 5. OTM distance revealed that the distance in miR21−/− mice with T cells was greater than that in miR21−/− mice without T cells, and less than that in WT mice. 6. RANKL/OPG ratio was significantly increased after the infusion of T cells in miR21−/− mice. 7. Macroscopic observation showed that the orthodontic tooth distance of miR- 21−/− mice with T cells was increased compared with that of miR21−/− mice without T cells. 8. The amount of OCs expression in miR21−/− infused with T cells was higher compared with that in WT mice, and the fewest OCs were seen in the miR21−/− mice. | miR21 deficiency constitutively suppresses tooth movement due to reduced RANKL circulation, which leads to a reduced number of OCs miR21 improves OCs via the Pdcd4/Cfos pathway, which influences the expression of the RANKL level secreted by T cells, enhances OC differentiation, and guides the osteoclastogenesis process. | miR21 is closely related to CD4+ T cells in immune responses and stimulates the release of RANKL. As a result, it stimulates the differentiation of OCs. | |
| Wang et al., 2020 [28] | To investigate the effect of miR21 on bone reconstruction by inducing maxillary bone defects in wild-type (WT) and miR21 knock-out (miR21-KO) mice, and exploring these mice as maxillary bone defect models. | WT and miR21-KO C57BL/6 mice. Female mice. 20–22 g. A round bone defect with a diameter of 1.2 mm was established at the maxillary bone on the palatal side of the incisors in each mouse. miR21-KO + agomir mice were injected with 100 μL of miR21 agomir intra-peritoneally every three days. The WT and miR21-KO mice received 100 μL PBS injections. Bone histology, histomorphometry, and immunohistochemical analyses were carried out. | 1. The lengths of the maxillary bone defects, as measured by H&E staining, were significantly longer in the miR21-KO group compared with the WT group. 2. Masson staining and the quantitative analyses also showed less new bone staining in the miR-21-KO group compared to WT mice. 3. The miR21-KO group exhibited lower ALP and OCN expressions than the WT group. 4. The value of BV/TV, as calculated by micro-CT, indicated that more new bone was detected in the miR21-KO + agomir group than in the miR21-KO group. 5. H&E staining showed that the length of the maxillary bone defect was decreased in the miR21-KO + agomir mice. | As concluded based on Li X et al., miR21 was found to promote osteogenesis of BMSCs via the Smad7-Smad1/5/8-Runx2 pathway. The results obtained in this study show that miR21 improved bone development. The vast differences can be seen in the miR KO and the miR KO + agomir. | miR21 was crucial for the bone reconstruction of maxillary defects in vivo, which suggests that it could be a therapeutic target for maxillary bone defects. | ||
| Sun et al., 2020 [15] | To explore the effect of nanoencapsulation (miR21) on the healing process of osteoporotic fractures as a new alternative method to accelerate the healing of osteoporotic fractures. | BMSCs were isolated from osteoporotic ovariectomized (OVX) rats. Flow cytometry to detect ALP expression and calcium deposition in BMSCs isolated from osteoporotic ovariectomized (OVX) rats. | Sprague–Dawley (SD) rats Elderly; 14-month-old. A model of female osteoporotic bone defects was generated using Sprague–Dawley rats. n = 30. Groups: A: CMCS/n (miR21); B: CMCS/n (NC-miR); C: saline. 3 months after the procedure, the lumbar bone mineral density and trabecular bone microarchitecture of the lumbar were measured. Histological and immunohistochemical studies were conducted | - miR21 or nanoencapsulated miR21 (acrylamide polymer) and O-carboxymethyl chitosan (CMCS) were mixed until they formed a gel-like material CMCS/n (miR21) - DLS and TEM analyses were carried out | 1. Electrophoresis and zeta potential analyses confirmed that the encapsulation efficacy of the nanocapsules for miR21 was satisfactory. 2. The results of ALP staining, toluidine blue staining, and oil red O staining confirmed that BMSCs derived from the OVX rat model could be differentiated from osteoblasts. 3. The result of confocal microscopy showed that significant green fluorescence was present in the cytoplasm of BMSCs after incubation with n (miR21) labeled with FAM. 4. Alizarin red staining confirmed that BMSCs treated with n (miR21) showed increased mineralization. 5. Micro-CT showed that the defect site was significantly bridged with new cancellous bone in the CMCS/n (miR21) group compared with the CMCS/n (NC-miR) and control groups. Quantitative analysis of alizarin red staining showed that the miR21-treated group had higher calcium nodule formation than that of the n (NC-miR) group or the control group. 6. miR21 promoted the expression of ALP and RUNX-2 genes in BMSCs isolated from OVX rats and inhibited the expression of caspase-3, thereby enabling the healing of osteoporotic bone defects. | miR21 accelerates the healing of osteoporotic bone defects by promoting the early expression of the ALP and RUNX-2 genes in BMSCs. | miR21 could promote early bone repair in osteoporotic bone defects by stimulating the osteogenic differentiation of BMSCs. |
| Smieszek et al., 2020 [32] | To investigate the impact of miR21 inhibition on pre-osteoblastic cell differentiation and paracrine signaling towards pre-osteoclasts, using an indirect co-culture model of mouse pre-osteoblast (MC3T3) and pre-osteoclast (4B12) cell lines. | A pre-osteoblastic mouse cell line MC3T3 and an osteoclast precursor cell line 4B12 were used. MC3T3 cells were transfected with miR21 inhibitor and then osteo-induced, followed by co-culturing with osteoclast precursor cell line 4B12. RT PCR, Western blot, SEM, ALP, and immunostaining were performed | 1. MC3T3 transfected with miR21 inhibitor and co-cultured with 4B12 showed a significant increase in Ocl. 2. Decreased mineralization was observed in cultures of MC3T3 with blocked activity of miR21. 3. Images obtained with a confocal microscope confirmed that OPN expression decreased after miR21 inhibition. 4. Increased levels of transcripts for RANK, TRAP, CTSK, CAII, and MMP-9 in 4B12 co-cultured with MC3T3 without an inhibitor were correlated with increased pre-osteoclast activity. 5. Decreased expression of miR21 in MC3T3 pre-osteoblasts cell line resulted in the loss of their bone-forming capability. 6. The inhibition of miR21 expression attenuated the paracrine activity of preosteoblasts, which was associated with increased apoptosis of pre-osteoclasts in the co-culture model. | miR21 regulates the differentiation of progenitor cells into bone-forming cells via the Smad7-Smad1/5/8-Runx2 pathway. | MiR21 may have a dual role in the process of osteoblast–osteoclast coupling, probably because its targets (e.g., RANKL and OPN) are regulated dynamically during the process of osteogenesis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subramaniam, R.; Vijakumaran, U.; Shanmuganantha, L.; Law, J.-X.; Alias, E.; Ng, M.-H. The Role and Mechanism of MicroRNA 21 in Osteogenesis: An Update. Int. J. Mol. Sci. 2023, 24, 11330. https://doi.org/10.3390/ijms241411330
Subramaniam R, Vijakumaran U, Shanmuganantha L, Law J-X, Alias E, Ng M-H. The Role and Mechanism of MicroRNA 21 in Osteogenesis: An Update. International Journal of Molecular Sciences. 2023; 24(14):11330. https://doi.org/10.3390/ijms241411330
Chicago/Turabian StyleSubramaniam, Revatyambigai, Ubashini Vijakumaran, Lohashenpahan Shanmuganantha, Jia-Xian Law, Ekram Alias, and Min-Hwei Ng. 2023. "The Role and Mechanism of MicroRNA 21 in Osteogenesis: An Update" International Journal of Molecular Sciences 24, no. 14: 11330. https://doi.org/10.3390/ijms241411330
APA StyleSubramaniam, R., Vijakumaran, U., Shanmuganantha, L., Law, J.-X., Alias, E., & Ng, M.-H. (2023). The Role and Mechanism of MicroRNA 21 in Osteogenesis: An Update. International Journal of Molecular Sciences, 24(14), 11330. https://doi.org/10.3390/ijms241411330