
AhRR and PPP1R3C: Potential Prognostic Biomarkers for Serous Ovarian Cancer

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Ovarian Cancer Spheroids Are Characterized by Stemness Markers’ Expression, Clonogenic Nature, and Peculiar Pathways Activation
2.2. AHRR, GALNT10, and PPP1R3C Expression Correlates with Patients’ Overall Survival
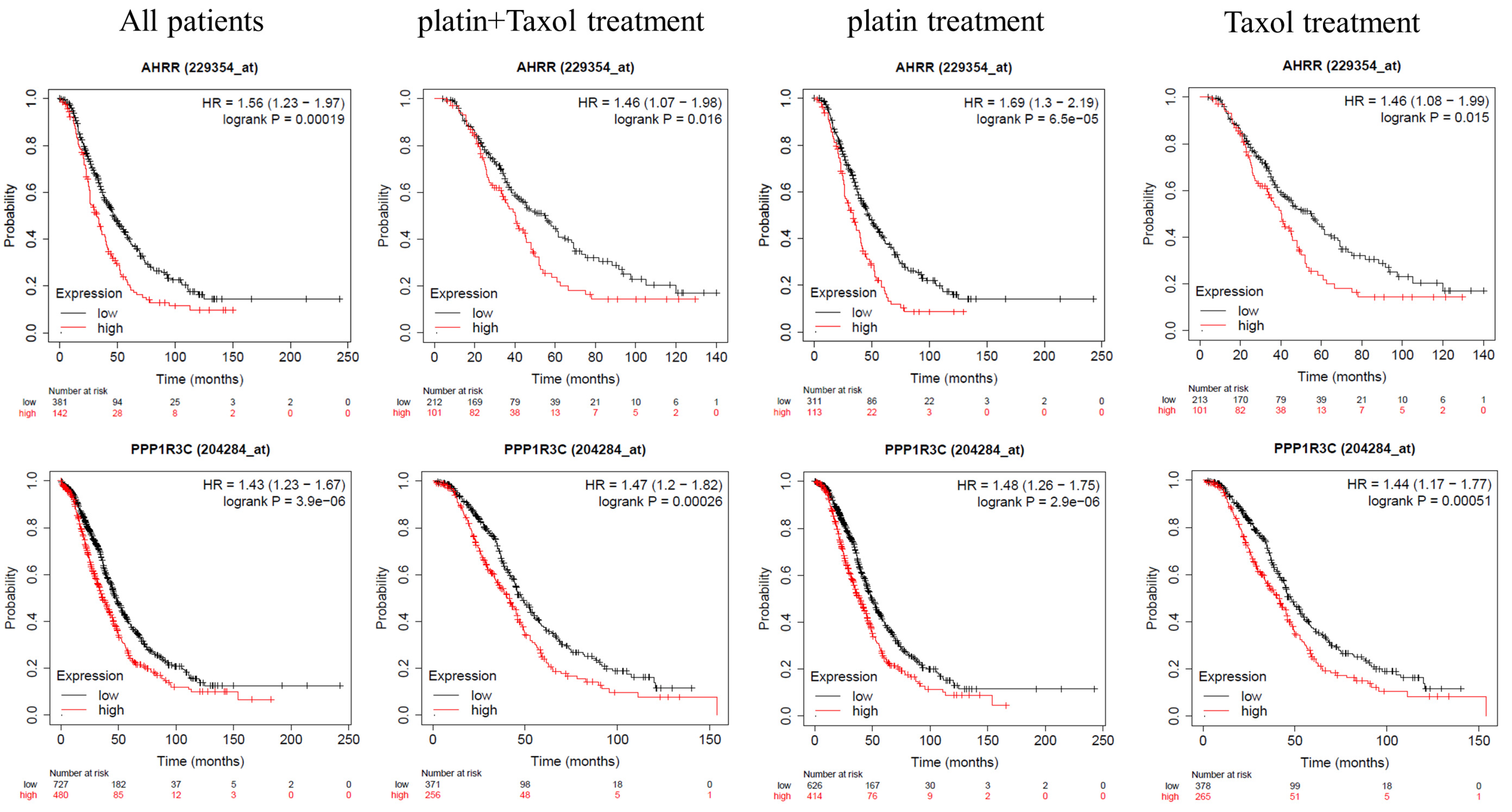
2.3. AhRR and PPP3R1C Expression Correlates with Patients’ Worse Prognoses
2.4. AhRR and PPP1R3C Expression Correlates with Prognosis in Other Cancers
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Ovarian Cancer Spheroids
4.3. PKH Assay
4.4. RNA Extraction and Real Time-PCR
4.5. Array-Comparative Genomic Hybridization (Array-CGH)
4.6. Bioinformatic Analyses
4.6.1. Analysis of Genes Involved in Copy Number Alterations and Their Respective Pathways
4.6.2. Analysis of AhRR, GALNT10 and PPP1R3C Expression in Ovarian Cancer
4.6.3. Correlation Analysis of AhRR and PPP1R3C with Stemness Markers
4.6.4. Correlation between CNAs and mRNA Expression for AHRR and PPP1R3C
4.6.5. Analysis of AHRR and PPP1R3C Expression in Other Cancers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisio, M.-A.; Fu, L.; Goyeneche, A.; Gao, Z.-H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Webb, P.M.; Jordan, S.J. Epidemiology of epithelial ovarian cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 41, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Kossaï, M.; Leary, A.; Scoazec, J.-Y.; Genestie, C. Ovarian Cancer: A Heterogeneous Disease. Pathobiology 2018, 85, 41–49. [Google Scholar] [CrossRef]
- Kroeger, P.T., Jr.; Drapkin, R. Pathogenesis and heterogeneity of ovarian cancer. Curr. Opin. Obs. Gynecol. 2017, 29, 26–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroki, L.; Guntupalli, S.R. Treatment of epithelial ovarian cancer. BMJ 2020, 371, m3773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Cheng, S.; Jin, Y.; Zhao, Y.; Wang, Y. Roles of CA125 in diagnosis, prediction, and oncogenesis of ovarian cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188503. [Google Scholar] [CrossRef]
- Najafi, M.; Mortezaee, K.; Majidpoor, J. Cancer stem cell (CSC) resistance drivers. Life Sci. 2019, 234, 116781. [Google Scholar] [CrossRef] [PubMed]
- Walcher, L.; Kistenmacher, A.-K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.-R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells—Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Yang, Y.; Liu, J.; Gao, Z.; Xu, T.; Chai, J.; Xu, J.; Fan, Z.; Xiao, T.; Jia, Q.; et al. Cancer stem cells in esophageal squamous cell carcinoma. Pathol. Res. Pract. 2022, 237, 154043. [Google Scholar] [CrossRef] [PubMed]
- Eun, K.; Ham, S.W.; Kim, H. Cancer stem cell heterogeneity: Origin and new perspectives on CSC targeting. BMB Rep. 2017, 50, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Conconi, D.; Jemma, A.; Giambra, M.; Redaelli, S.; Croci, G.A.; Dalprà, L.; Lavitrano, M.; Bentivegna, A. Analysis of copy number alterations in bladder cancer stem cells revealed a prognostic role of LRP1B. World J. Urol. 2022, 40, 2267–2273. [Google Scholar] [CrossRef]
- Giambra, M.; Di Cristofori, A.; Conconi, D.; Marzorati, M.; Redaelli, S.; Zambuto, M.; Rocca, A.; Roumy, L.; Carrabba, G.; Lavitrano, M.; et al. Insights into the Peritumoural Brain Zone of Glioblastoma: CDK4 and EXT2 May Be Potential Drivers of Malignancy. Int. J. Mol. Sci. 2023, 24, 2835. [Google Scholar] [CrossRef]
- Giambra, M.; Messuti, E.; Di Cristofori, A.; Cavandoli, C.; Bruno, R.; Buonanno, R.; Marzorati, M.; Zambuto, M.; Rodriguez-Menendez, V.; Redaelli, S.; et al. Characterizing the Genomic Profile in High-Grade Gliomas: From Tumor Core to Peritumoral Brain Zone, Passing through Glioma-Derived Tumorspheres. Biology 2021, 10, 1157. [Google Scholar] [CrossRef]
- Doran, C.G.; Pennington, S.R. Copy number alteration signatures as biomarkers in cancer: A review. Biomark. Med. 2022, 16, 371–386. [Google Scholar] [CrossRef]
- Levine, D.A. The Cancer Genome Atlas Research Network. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macintyre, G.; Goranova, T.E.; De Silva, D.; Ennis, D.; Piskorz, A.M.; Eldridge, M.; Sie, D.; Lewsley, L.-A.; Hanif, A.; Wilson, C.; et al. Copy number signatures and mutational processes in ovarian carcinoma. Nat. Genet. 2018, 50, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liang, H.; Xiao, W.; Wei, P.; Chen, H.; Chen, Z.; Yang, R.; Jiang, H.; Zhang, Y. Whole-exome mutational landscape and molecular marker study in mucinous and clear cell ovarian cancer cell lines 3AO and ES2. BMC Cancer 2023, 23, 321. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Yo, Y.T.; Lee, H.Y.; Liao, Y.P.; Chao, T.K.; Su, P.H.; Lai, H.C. ALDH1-bright epithelial ovarian cancer cells are associated with CD44 expression, drug resistance, and poor clinical outcome. Am. J. Pathol. 2012, 180, 1159–1169. [Google Scholar] [CrossRef]
- Qin, S.; Li, Y.; Cao, X.; Du, J.; Huang, X. NANOG regulates epithelial–mesenchymal transition and chemoresistance in ovarian cancer. Biosci. Rep. 2017, 37, BSR20160247. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Hou, Y.; Huang, Z.; Cai, J.; Wang, Z. SOX2 is required to maintain cancer stem cells in ovarian cancer. Cancer Sci. 2017, 108, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zou, Z.; Tang, J.; Zheng, Y.; Liu, Y.; Luo, Y.; Liu, Q.; Wang, Y. NOS1 upregulates ABCG2 expression contributing to DDP chemoresistance in ovarian cancer cells. Oncol. Lett. 2019, 17, 1595–1602. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.-W.; Wu, J.-H.; Jiang, C.-P. ABCG2: A potential marker of stem cells and novel target in stem cell and cancer therapy. Life Sci. 2010, 86, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.-Y.; Zhang, Q.; Wang, J.-M.; Jiang, J.-Y.; Huyan, L.-Y.; Liu, B.-Q.; Yan, J.; Li, C.; Wang, H.-Q. BAG3 epigenetically regulates GALNT10 expression via WDR5 and facilitates the stem cell-like properties of platin-resistant ovarian cancer cells. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119077. [Google Scholar] [CrossRef]
- Zhang, G.; Lu, J.; Yang, M.; Wang, Y.; Liu, H.; Xu, C. Elevated GALNT10 expression identifies immunosuppressive microenvironment and dismal prognosis of patients with high grade serous ovarian cancer. Cancer Immunol. Immunother. 2020, 69, 175–187. [Google Scholar] [CrossRef]
- Kousoulidou, L.; Sismani, C.; Patsalis, P.C. Multiplex Amplifiable Probe Hybridization (MAPH) Methodology as an Alternative to Comparative Genomic Hybridization (CGH). Methods Mol. Biol. 2010, 653, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Kumar, N.; Baudis, M. arrayMap: A Reference Resource for Genomic Copy Number Imbalances in Human Malignancies. PLoS ONE 2012, 7, e36944. [Google Scholar] [CrossRef]
- Trasanidis, N.; Katsarou, A.; Ponnusamy, K.; Shen, Y.-A.; Kostopoulos, I.V.; Bergonia, B.; Keren, K.; Reema, P.; Xiao, X.; Szydlo, R.M.; et al. Systems medicine dissection of chr1q-amp reveals a novel PBX1-FOXM1 axis for targeted therapy in multiple myeloma. Blood 2022, 139, 1939–1953. [Google Scholar] [CrossRef]
- Garcia-Mayea, Y.; Mir, C.; Masson, F.; Paciucci, R.; Lleonart, M. Insights into new mechanisms and models of cancer stem cell multidrug resistance. Semin. Cancer Biol. 2020, 60, 166–180. [Google Scholar] [CrossRef]
- Akhtar, S.; Hourani, S.; Therachiyil, L.; Al-Dhfyan, A.; Agouni, A.; Zeidan, A.; Uddin, S.; Korashy, H.M. Epigenetic Regulation of Cancer Stem Cells by the Aryl Hydrocarbon Receptor Pathway. Semin. Cancer Biol. 2022, 83, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Zudaire, E.; Cuesta, N.; Murty, V.; Woodson, K.; Adams, L.; Gonzalez, N.; Martínez, A.; Narayan, G.; Kirsch, I.; Franklin, W.; et al. The aryl hydrocarbon receptor repressor is a putative tumor suppressor gene in multiple human cancers. J. Clin. Investig. 2008, 118, 640–650. [Google Scholar] [CrossRef]
- Hu, A.; Zhang, J.W.; Yang, L.Y.; Qiao, P.P.; Lu, D. AHRR contributes to inflammatory lymphangiogenesis by activating the EPAS1-VEGFD signaling axis in head and neck cancer. Am. J. Cancer Res. 2022, 12, 537–548. [Google Scholar]
- Zhou, Y.; Zhou, S.-X.; Gao, L.; Li, X.-A. Regulation of CD40 signaling in colon cancer cells and its implications in clinical tissues. Cancer Immunol. Immunother. 2016, 65, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; Yun, J.H.; Jung, J.; Yang, Y.; Kim, B.-J.; Lee, S.-J.; Yoon, J.H.; Moon, Y.; Kim, J.-M.; Kwon, Y.-I. Identification of differentially-expressed genes by DNA methylation in cervical cancer. Oncol. Lett. 2015, 9, 1691–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Hurk, K.V.D.; Moerkerk, P.T.; Goeman, J.J.; Beck, S.; Gruis, N.A.; Oord, J.J.V.D.; Winnepenninckx, V.J.; van Engeland, M.; van Doorn, R. Promoter CpG Island Hypermethylation in Dysplastic Nevus and Melanoma: CLDN11 as an Epigenetic Biomarker for Malignancy. J. Investig. Dermatol. 2014, 134, 2957–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Ye, G.; Wang, Y.; Luo, D. MiR-4461 Inhibits Tumorigenesis of Renal Cell Carcinoma by Targeting PPP1R3C. Cancer Biother. Radiopharm. 2022, 37, 503–514. [Google Scholar] [CrossRef]
- Li, Z.; Chen, J.; Zhao, S.; Li, Y.; Zhou, J.; Liang, J.; Tang, H. Discovery and validation of novel biomarkers for detection of cervical cancer. Cancer Med. 2021, 10, 2063–2074. [Google Scholar] [CrossRef]
- Lee, S.K.; Moon, J.W.; Lee, Y.W.; Lee, J.O.; Kim, S.J.; Kim, N.; Kim, J.; Kim, H.S.; Park, S.-H. The effect of high glucose levels on the hypermethylation of protein phosphatase 1 regulatory subunit 3C (PPP1R3C) gene in colorectal cancer. J. Genet. 2015, 94, 75–85. [Google Scholar] [CrossRef]
- Ko, C.-I.; Fan, Y.; de Gannes, M.; Wang, Q.; Xia, Y.; Puga, A. Repression of the Aryl Hydrocarbon Receptor Is Required to Maintain Mitotic Progression and Prevent Loss of Pluripotency of Embryonic Stem Cells. Stem Cells 2016, 34, 2825–2839. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, B.; Eisenhauer, E.; Tregear, M.; Booth, C.M. Progression-free survival: It is time for a new name. Lancet Oncol. 2022, 23, 328–330. [Google Scholar] [CrossRef]
- Walters Haygood, C.L.; Arend, R.C.; Straughn, J.M.; Buchsbaum, D.J. Ovarian cancer stem cells: Can targeted therapy lead to improved progression-free survival? World J. Stem Cells 2014, 6, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wang, W.; Li, F.; Songyang, Z.; Feng, X.; Xin, C.; Dai, Z.; Xiong, Y. Pan-cancer analysis identifies telomerase-associated signatures and cancer subtypes. Mol. Cancer 2019, 18, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.D.; Komosa, M.; Nunes, N.M.; Tabori, U. DNA methylation of the TERT promoter and its impact on human cancer. Curr. Opin. Genet. Dev. 2020, 60, 17–24. [Google Scholar] [CrossRef]
- Ishiguro, T.; Ohata, H.; Sato, A.; Yamawaki, K.; Enomoto, T.; Okamoto, K. Tumor-derived spheroids: Relevance to cancer stem cells and clinical applications. Cancer Sci. 2017, 108, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, S.W.; Shaw, C.A.; Scott, D.A.; Patel, A.; Sahoo, T.; Bacino, C.A.; Pursley, A.; Li, J.; Erickson, R.; Gropman, A.L.; et al. Microarray-based CGH detects chromosomal mosaicism not revealed by conventional cytogenetics. Am. J. Med. Genet. Part A 2007, 143A, 1679–1686. [Google Scholar] [CrossRef] [PubMed]
- Conconi, D.; Redaelli, S.; Bovo, G.; Leone, B.E.; Filippi, E.; Ambrosiani, L.; Cerrito, M.G.; Grassilli, E.; Giovannoni, R.; Dalprà, L.; et al. Unexpected frequency of genomic alterations in histologically normal colonic tissue from colon cancer patients. Tumor Biol. 2016, 37, 13831–13842. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Human Protein Atlas | OncoDB | GEPIA2 | Kaplan–Meier Plotter (RNA-Seq Data) | Kaplan–Meier Plotter (Gene CHIP Data) | |
|---|---|---|---|---|---|
| Copy number gains shared among all spheroids and all cell lines | |||||
| ADRA1B | p < 0.01 favorable | ns | ns | p < 0.01 favorable | p < 0.01 favorable |
| AHRR | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable |
| ATP10B | p < 0.05 favorable | ns | ns | p < 0.05 favorable | ns |
| CREBRF | ns | ns | ns | ns | p < 0.01 unfavorable |
| EBF1 | ns | ns | ns | ns | p < 0.01 unfavorable |
| GFPT2 | p < 0.05 unfavorable | p < 0.05 unfavorable | ns | p < 0.05 unfavorable | p < 0.01 unfavorable |
| GPRIN1 | ns | ns | ns | ns | p < 0.01 unfavorable |
| KCNMB1 | p < 0.05 favorable | ns | ns | ns | ns |
| LSM11 | p < 0.05 unfavorable | ns | ns | p < 0.05 unfavorable | p < 0.05 favorable |
| PROP1 | ns | ns | ns | p < 0.05 unfavorable | p < 0.05 favorable |
| RASGEF1C | ns | ns | ns | ns | p < 0.05 favorable |
| TBC1D9B | ns | ns | ns | ns | p < 0.01 favorable |
| Copy number gains shared among all spheroids and two cell lines | |||||
| ADAM19 | ns | ns | ns | ns | p < 0.01 favorable |
| ADAMTS2 | p < 0.05 unfavorable | ns | ns | p < 0.05 unfavorable | p < 0.01 unfavorable |
| COL23A1 | p < 0.05 unfavorable | ns | ns | ns | p < 0.01 unfavorable |
| CPLX2 | ns | ns | ns | ns | p < 0.05 unfavorable |
| FABP6 | p < 0.05 favorable | ns | ns | p < 0.05 favorable | ns |
| GABRA1 | na | na | na | p < 0.01 unfavorable | ns |
| GABRA6 | na | na | na | p < 0.01 unfavorable | p < 0.05 unfavorable |
| GABRB2 | ns | ns | ns | ns | p < 0.01 unfavorable |
| GABRP | p < 0.01 favorable | ns | ns | p < 0.01 favorable | p < 0.01 favorable |
| GALNT10 | p < 0.01 unfavorable | p < 0.05 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.05 unfavorable |
| GLRA1 | na | ns | na | p < 0.01 unfavorable | p < 0.05 favorable |
| GRIA1 | p < 0.05 unfavorable | ns | p < 0.05 unfavorable | p < 0.05 unfavorable | ns |
| HNRNPAB | p < 0.01 favorable | ns | ns | p < 0.01 favorable | ns |
| KCNIP1 | p < 0.01 favorable | ns | ns | p < 0.01 favorable | ns |
| LARP1 | ns | ns | ns | ns | p < 0.05 favorable |
| MAMDC2 | p < 0.05 unfavorable | ns | ns | p < 0.05 unfavorable | p < 0.05 unfavorable |
| MFAP3 | p < 0.05 unfavorable | ns | p < 0.05 unfavorable | p < 0.05 unfavorable | ns |
| MKRN2 | ns | ns | ns | ns | p < 0.01 unfavorable |
| MIR762HG | na | p < 0.05 favorable | p < 0.01 favorable | na | na |
| NDST1 | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | ns |
| NEURL1 | p < 0.05 unfavorable | ns | ns | p < 0.05 unfavorable | ns |
| NSD1 | ns | ns | ns | ns | p < 0.01 unfavorable |
| PC | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | ns |
| RANBP17 | ns | ns | ns | ns | p < 0.05 favorable |
| RNF130 | ns | ns | p < 0.05 unfavorable | ns | p < 0.05 unfavorable |
| SFXN1 | p < 0.05 favorable | ns | ns | p < 0.05 favorable | p < 0.05 unfavorable |
| SGCD | ns | ns | ns | ns | ns |
| SH3PXD2B | p < 0.05 unfavorable | ns | ns | p < 0.05 unfavorable | p < 0.05 unfavorable |
| SLIT3 | p < 0.01 unfavorable | ns | ns | p < 0.05 unfavorable | ns |
| TENM2 | p < 0.05 favorable | ns | ns | p < 0.05 favorable | ns |
| UBTD2 | ns | ns | ns | ns | p < 0.01 unfavorable |
| UIMC1 | ns | ns | ns | ns | ns |
| ZNF346 | ns | ns | ns | ns | p < 0.05 unfavorable |
| Copy number gains shared among all spheroids and one cell line | |||||
| NLRP12 | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | ns |
| PPP1R3C | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable | p < 0.01 unfavorable |
| PYROXD2 | ns | ns | ns | ns | p < 0.01 favorable |
| Ovcar5 Line | Ovcar5 Spheroids | Ovcar8 Line | Ovcar8 Spheroids | Caov3 Line | Caov3 Spheroids | ||
|---|---|---|---|---|---|---|---|
| AhRR | CNA | non mosaic gain | non mosaic gain | non mosaic gain | non mosaic gain | non mosaic gain | amplification |
| mRNA | * fc = 53 ± 19 | * fc = 26 ± 4 | * fc = 27 ± 11 | * fc = 44 ± 27 | fc = 0.5 ± 0.03 | fc = 0.4 ± 0.1 | |
| PPP1R3C | CNA | disomy | mosaic gain (43%) | non mosaic gain | non mosaic gain | disomy | non mosaic gain |
| mRNA | * fc = 28 ± 4 | * fc = 24 ± 7 | fc = 0.4 ± 0.2 | fc = 1 ± 0.8 | fc = 1.3 ± 0.15 | fc = 1 ± 0.25 |
| Gene | Cases with CNAs n° | 6 Months Survival | 1 Year Survival | 5 Years Survival | Overall Survival | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gain | Loss | Gain | Loss | Gain | Loss | Gain | Loss | |||||||||||
| Gain | Loss | Alive | Dead | Alive | Dead | Alive | Dead | Alive | Dead | Alive | Dead | Alive | Dead | Alive | Dead | Alive | Dead | |
| AhRR | 301 | 43 | 285/301 94.7% | 16/301 5.3% | 42/43 97.7% | 1/43 2.3% | 275/301 91.4% | 26/301 8.6% | 37/43 86% | 6/43 14% | 154/301 51.2% | 147/301 48.8% | 22/43 51.1% | 21/43 48.8% | 123/301 40.8% | 178/301 59.1% | 20/43 46.5% | 23/43 53.5% |
| PPP1R3C | 94 | 167 | 90/94 95.7% | 4/94 4.3% | 162/167 97% | 5/167 3% | 86/94 91.5% | 8/94 8.5% | 158/167 94.6% | 9/167 5.4% | 45/94 47.9% | 49/94 52.1% | 95/167 56.9% | 72/167 43.1% | 38/94 40.4% | 56/94 59.6% | 76/167 45.5% | 91/167 54.5% |
| GDC all cases (585) | 6 months survival: 555 alive (94.9%), 30 dead (5.1%) | 1 year survival: 531 alive (90.8%), 54 dead (9.2%) | 5 years survival: 293 alive (50.1%), 292 dead (49.9%) | Overall survival: 236 alive (40.3%), 349 dead (59.7%) | ||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ardizzoia, A.; Jemma, A.; Redaelli, S.; Silva, M.; Bentivegna, A.; Lavitrano, M.; Conconi, D. AhRR and PPP1R3C: Potential Prognostic Biomarkers for Serous Ovarian Cancer. Int. J. Mol. Sci. 2023, 24, 11455. https://doi.org/10.3390/ijms241411455
Ardizzoia A, Jemma A, Redaelli S, Silva M, Bentivegna A, Lavitrano M, Conconi D. AhRR and PPP1R3C: Potential Prognostic Biomarkers for Serous Ovarian Cancer. International Journal of Molecular Sciences. 2023; 24(14):11455. https://doi.org/10.3390/ijms241411455
Chicago/Turabian StyleArdizzoia, Alessandra, Andrea Jemma, Serena Redaelli, Marco Silva, Angela Bentivegna, Marialuisa Lavitrano, and Donatella Conconi. 2023. "AhRR and PPP1R3C: Potential Prognostic Biomarkers for Serous Ovarian Cancer" International Journal of Molecular Sciences 24, no. 14: 11455. https://doi.org/10.3390/ijms241411455
APA StyleArdizzoia, A., Jemma, A., Redaelli, S., Silva, M., Bentivegna, A., Lavitrano, M., & Conconi, D. (2023). AhRR and PPP1R3C: Potential Prognostic Biomarkers for Serous Ovarian Cancer. International Journal of Molecular Sciences, 24(14), 11455. https://doi.org/10.3390/ijms241411455