
Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms


 , , , and
, , , and 
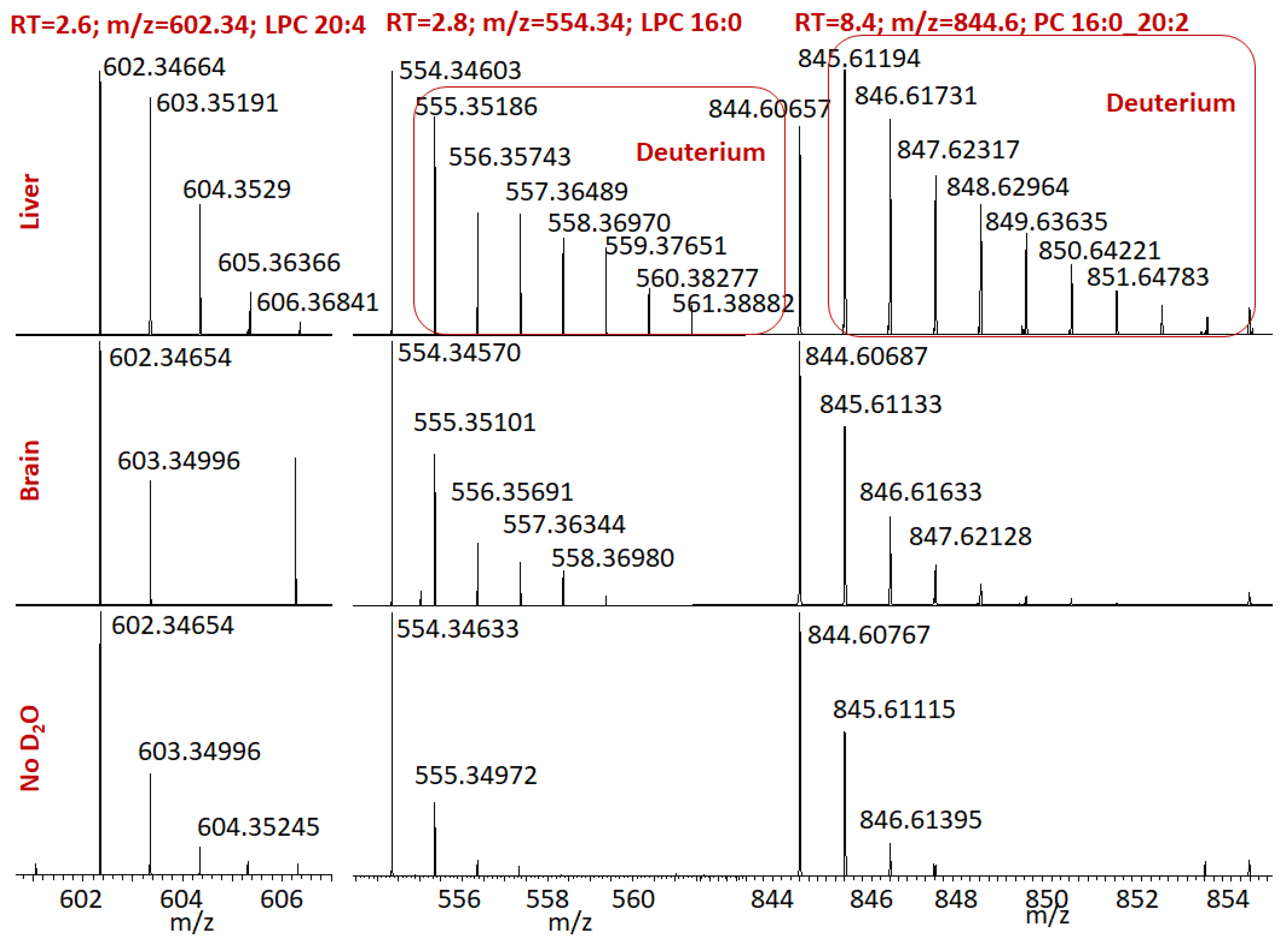{kind=link}
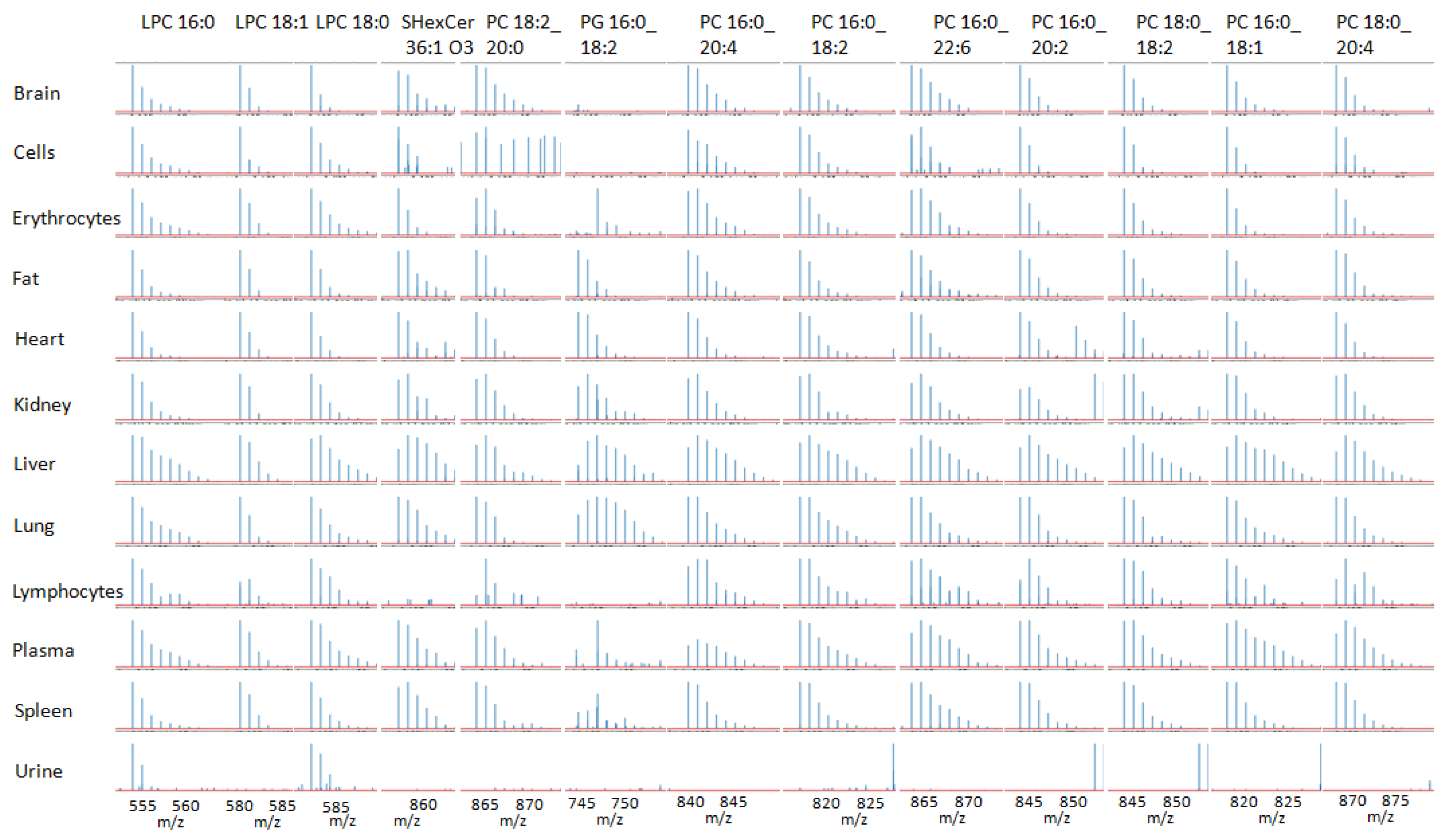{kind=link}
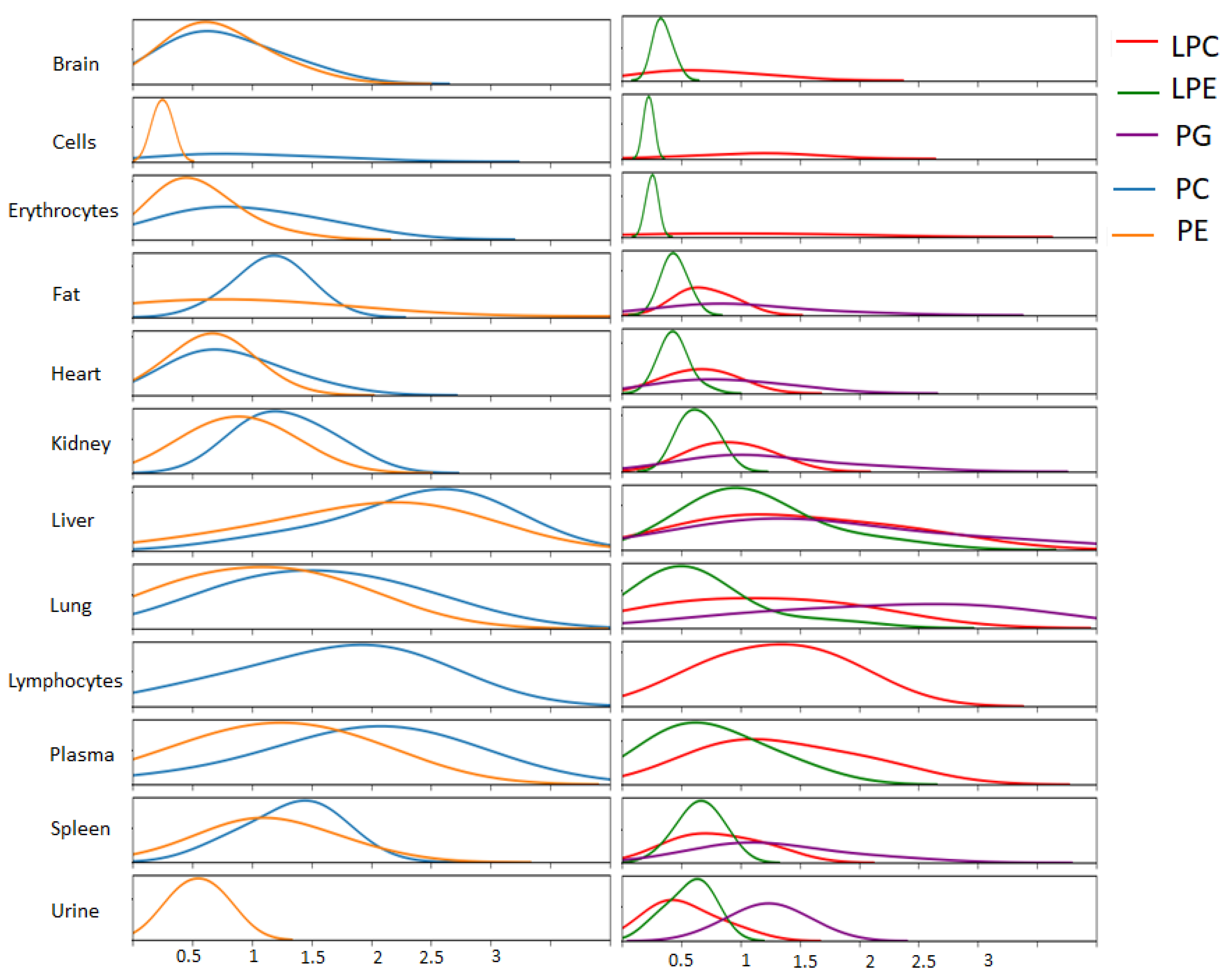{kind=link}
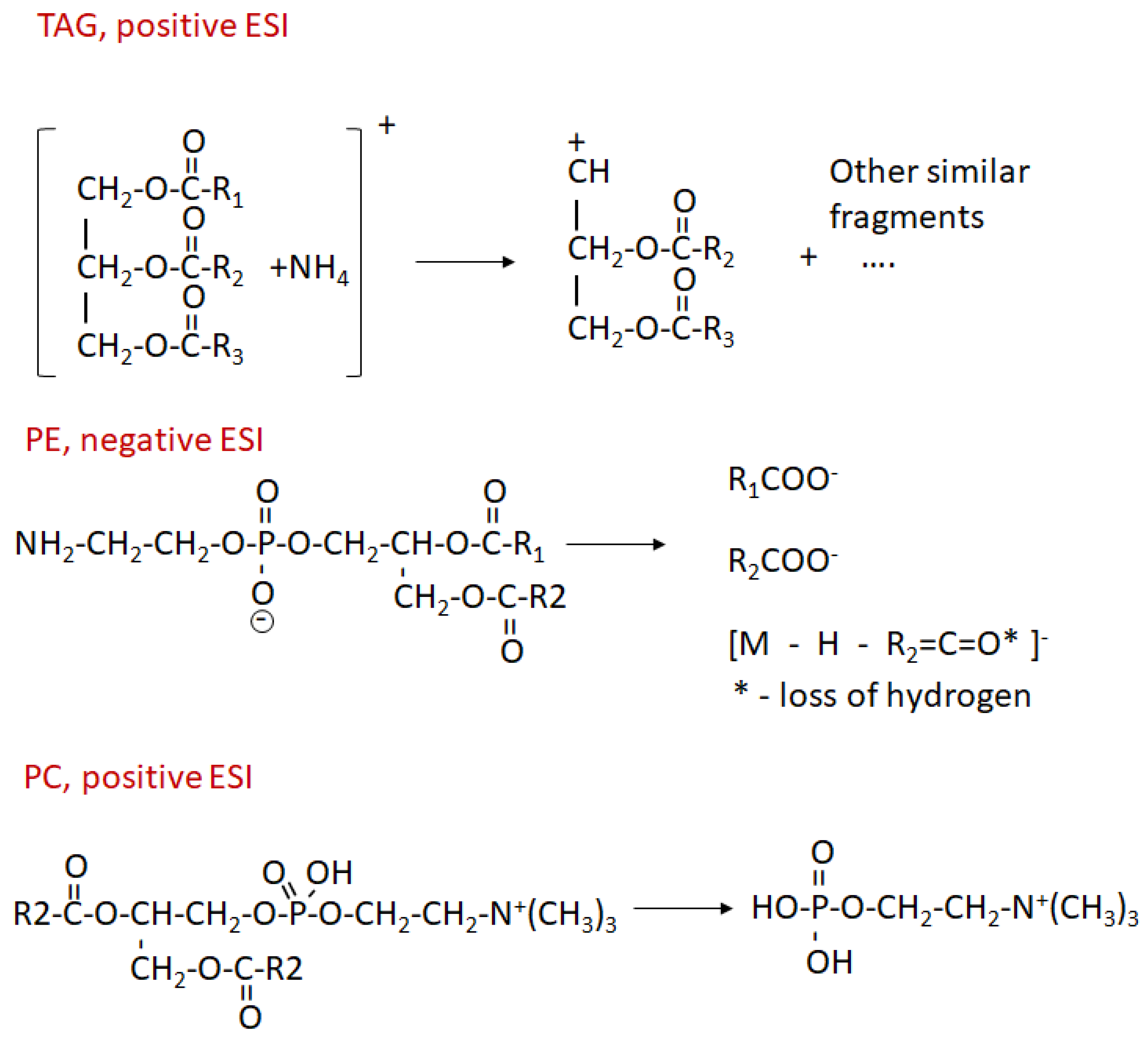{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
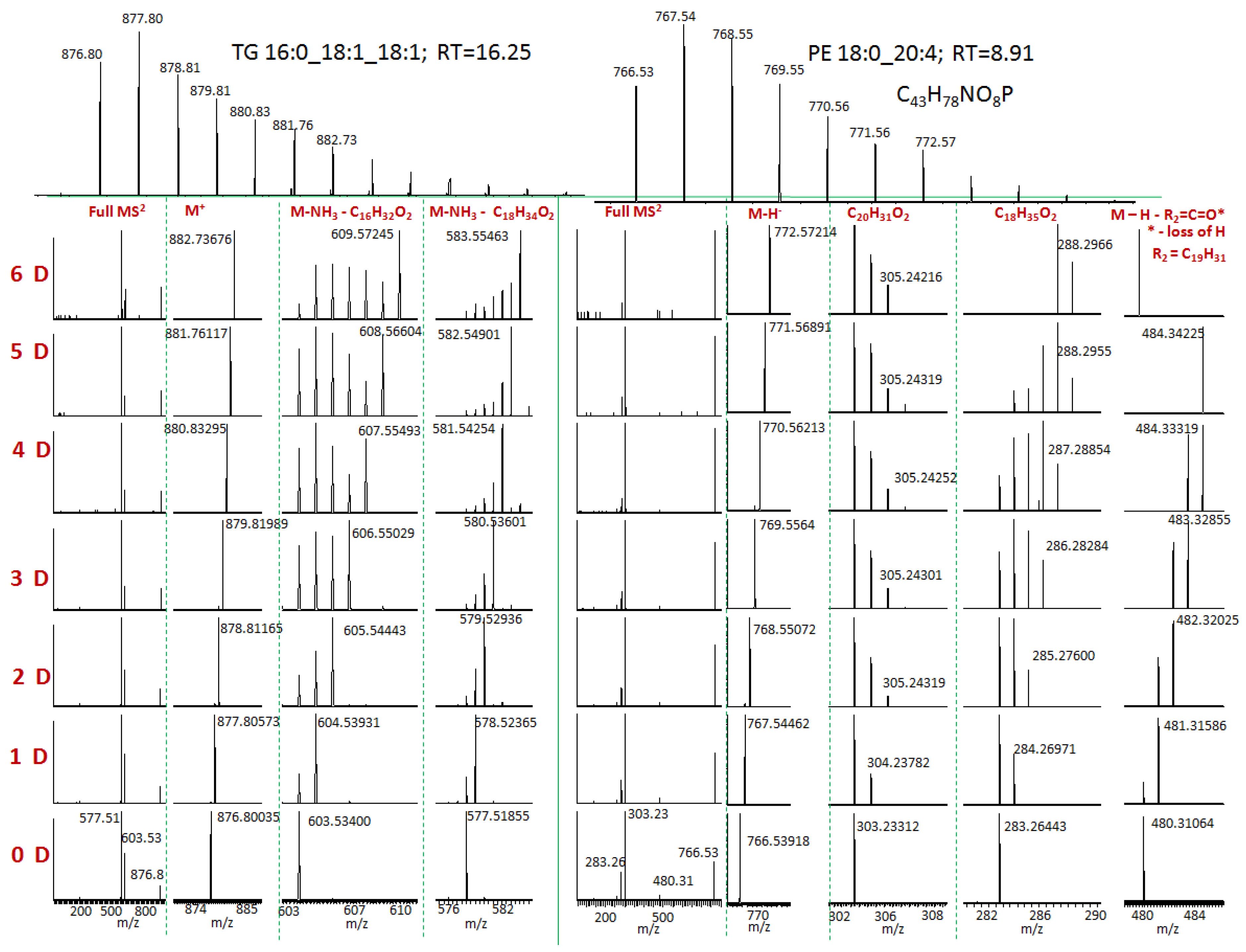
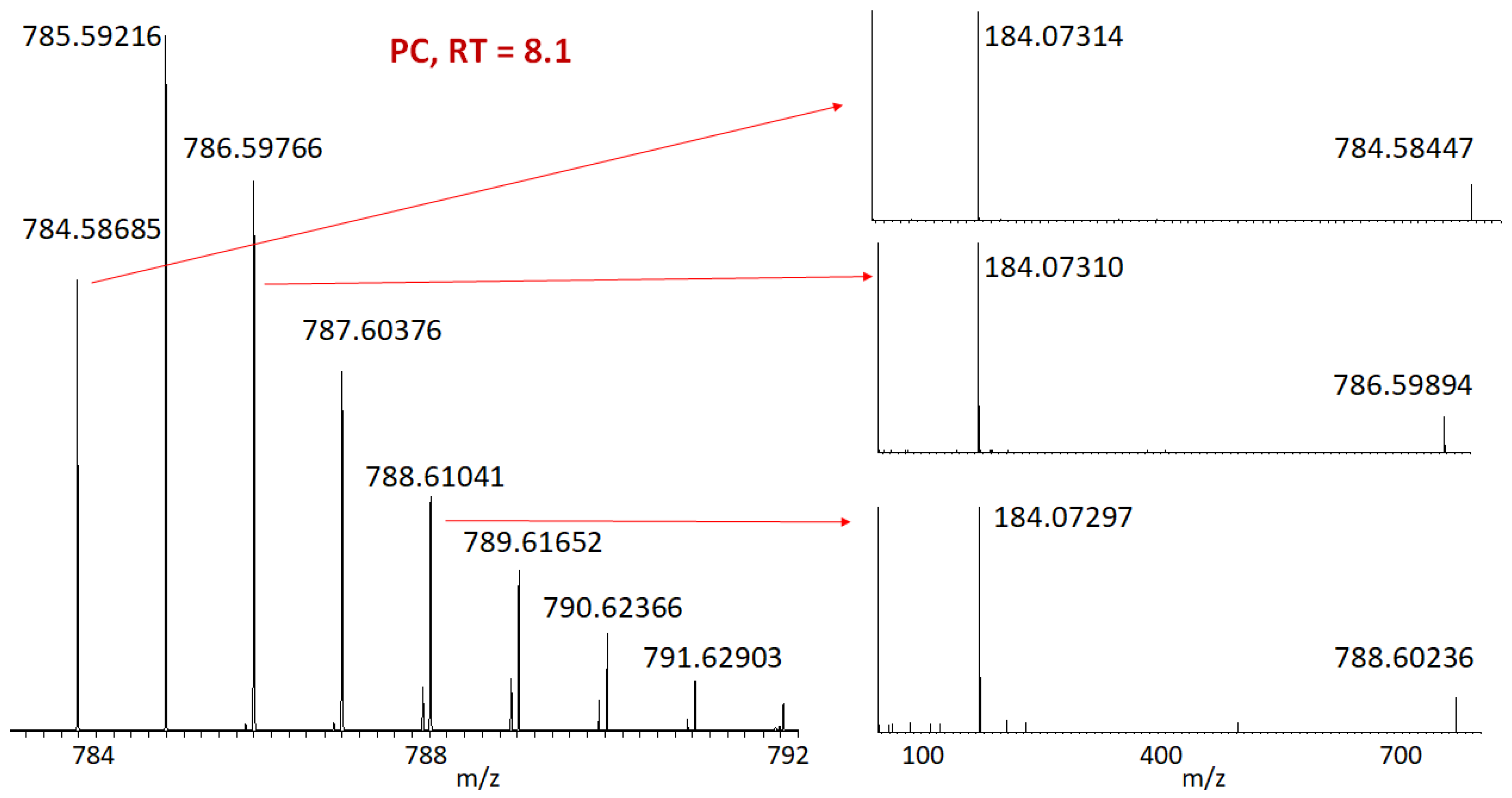
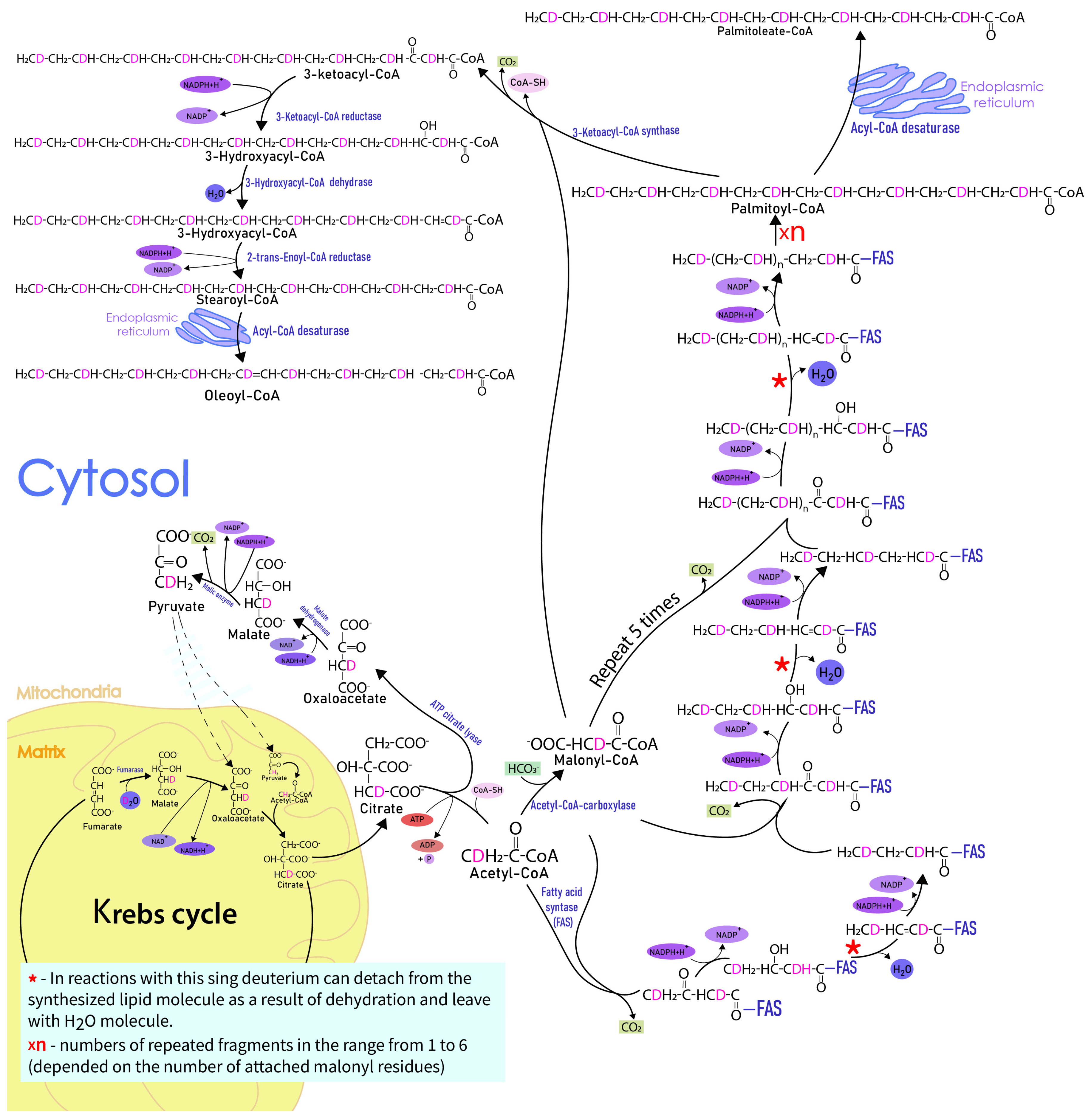
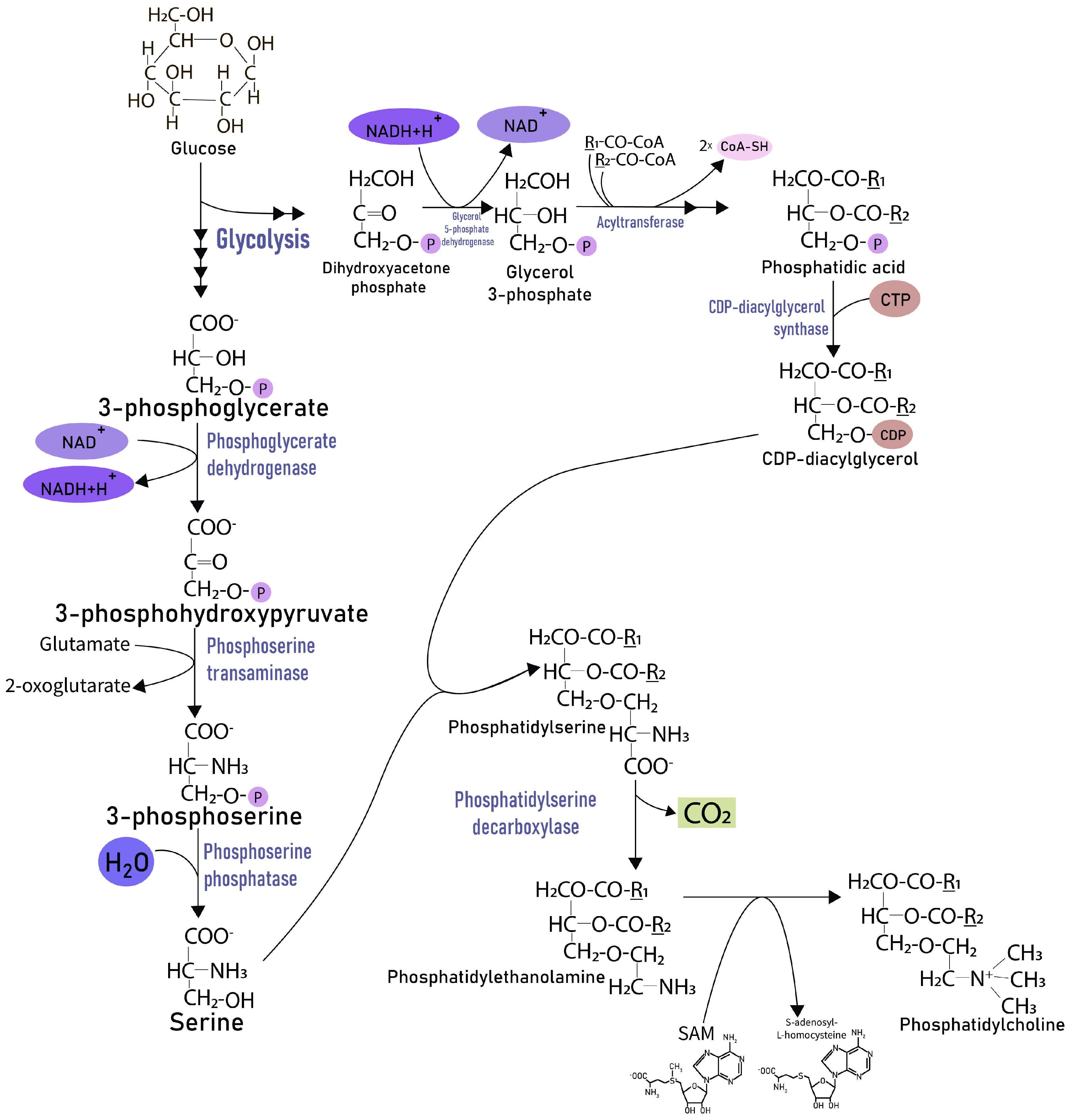
2. Results
3. Discussion
4. Materials and Methods
- A total of 300 mL of cold methanol was added to 40 mL of aliquots of the sample and vigorously shaken on a shaker for 1 min;
- Then, 1 mL of cold MTBE was added, and the mixture was treated with ultrasound for 10 min and incubated for 40 min at 4 °C with stirring;
- A total of 250 mL of water was added to the extract to separate the phases. The extract was shaken for 1 min at 4 °C, then centrifuged for 10 min at 13,000 rpm at 4 °C;
- An aliquot of 1000 µL of the upper layer containing nonpolar components was collected in a separate vial;
- A total of 400 µL of buffer (MeOH:MTBE:H2O = 3:10:2.5) was added to the lower phase for repeated extraction;
- The sample was shaken and centrifuged for 10 min at 13,000 rpm at 4 °C. The upper fractions were combined (a total of 1300 µL) and evaporated dry in a vacuum concentrator at room temperature;
- The dry residue was re-dissolved in 200 mL of a mixture of acetonitrile: isopropanol cooled to 0 °C (7:3 (v/v));
- The sample was shaken for 10 min, kept in an ice-cooled ultrasonic bath for 10 min, and centrifuged for 5 min at 13,000 rpm;
- Before the HPLC MS analysis, the samples were diluted 1:5 and 1:2 with a mixture of acetonitrile: isopropanol (7:3 (v/v)) for measurements in the registration mode of positively and negatively charged ions, respectively.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boren, J.; Taskinen, M.R.; Olofsson, S.O.; Levin, M. Ectopic lipid storage and insulin resistance: A harmful relationship. J. Intern. Med. 2013, 274, 25–40. [Google Scholar] [CrossRef]
- Guebre-Egziabher, F.; Alix, P.M.; Koppe, L.; Pelletier, C.C.; Kalbacher, E.; Fouque, D.; Soulage, C.O. Ectopic lipid accumulation: A potential cause for metabolic disturbances and a contributor to the alteration of kidney function. Biochimie 2013, 95, 1971–1979. [Google Scholar] [CrossRef]
- Duarte, J.A.; Carvalho, F.; Pearson, M.; Horton, J.D.; Browning, J.D.; Jones, J.G.; Burgess, S.C. A high-fat diet suppresses de novo lipogenesis and desaturation but not elongation and triglyceride synthesis in mice. J. Lipid Res. 2014, 55, 2541–2553. [Google Scholar] [CrossRef] [Green Version]
- Ando, S.; Tanaka, Y.; Toyoda, Y.; Kon, K. Turnover of myelin lipids in aging brain. Neurochem. Res. 2003, 28, 5–13. [Google Scholar] [CrossRef]
- Schoenheimer, R.; Rittenberg, D. The application of isotopes to the study of intermediary metabolism. Science 1938, 87, 221–226. [Google Scholar] [CrossRef]
- Barrett, H.; Best, C.; Ridout, J.H. A study of the source of liver fat using deuterium as an indicator. J. Physiol. 1938, 93, 367. [Google Scholar] [CrossRef] [Green Version]
- Schoenheimer, R.; Rittenberg, D. Deuterium as an indicator in the study of intermediary metabolism. Science 1935, 82, 156–157. [Google Scholar] [CrossRef]
- Schoenheimer, R.; Rittenberg, D. Deuterium as an indicator in the study of intermediary metabolism. 9. The conversion of stearic acid into palmitic acid in the organism. J. Biol. Chem. 1937, 120, 155–165. [Google Scholar] [CrossRef]
- Smith, M.E. The turnover of myelin in the adult rat. Biochim. Biophys. Acta (BBA) Lipids Lipid Metab. 1968, 164, 285–293. [Google Scholar] [CrossRef]
- Jungalwala, F.B. The turnover of myelin phosphatidylcholine and sphingomyelin in the adult rat brain. Brain Res. 1974, 78, 99–108. [Google Scholar] [CrossRef]
- Jungalwala, F.; Dawson, R. The turnover of myelin phospholipids in the adult and developing rat brain. Biochem. J. 1971, 123, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Brook, M.S.; Wilkinson, D.J.; Atherton, P.J.; Smith, K. Recent developments in deuterium oxide tracer approaches to measure rates of substrate turnover: Implications for protein, lipid, and nucleic acid research. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Allister, C.A.; Liu, L.-f.; Lamendola, C.A.; Craig, C.M.; Cushman, S.W.; Hellerstein, M.K.; McLaughlin, T.L. In vivo 2H2O administration reveals impaired triglyceride storage in adipose tissue of insulin-resistant humans1. J. Lipid Res. 2015, 56, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Bassilian, S.; Ajie, H.; Schoeller, D.A.; Edmond, J.; Bergner, E.A.; Byerley, L.O. In vivo measurement of fatty acids and cholesterol synthesis using D2O and mass isotopomer analysis. Am. J. Physiol. Endocrinol. Metab. 1994, 266, E699–E708. [Google Scholar] [CrossRef]
- Fu, X.; Deja, S.; Fletcher, J.A.; Anderson, N.N.; Mizerska, M.; Vale, G.; Browning, J.D.; Horton, J.D.; McDonald, J.G.; Mitsche, M.A.; et al. Measurement of lipogenic flux by deuterium resolved mass spectrometry. Nat. Commun. 2021, 12, 3756. [Google Scholar] [CrossRef]
- Leitch, C.A.; Jones, P.J. Measurement of triglyceride synthesis in humans using deuterium oxide and isotope ratio mass spectrometry. Biol. Mass Spectrom. 1991, 20, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Scrimgeour, C.; Begley, I.; Thomason, M. Measurement of deuterium incorporation into fatty acids by gas chromatography/isotope ratio mass spectrometry. Rapid Commun. Mass Spectrom. 1999, 13, 271–274. [Google Scholar] [CrossRef]
- Rumiantseva, L.; Osipenko, S.; Zharikov, A.; Kireev, A.; Nikolaev, E.N.; Kostyukevich, Y. Analysis of 16O/18O and H/D Exchange Reactions between Carbohydrates and Heavy Water Using High-Resolution Mass Spectrometry. Int. J. Mol. Sci. 2022, 23, 3585. [Google Scholar] [CrossRef]
- Rumiantseva, L.; Osipenko, S.; Podolskiy, I.I.; Burmykin, D.A.; Kovaleva, O.; Nikolaev, E.N.; Kostyukevich, Y. Increasing the reliability of compound identification in biological samples using 16O/18O-exchange mass spectrometry. Anal. Bioanal. Chem. 2022, 414, 2537–2543. [Google Scholar] [CrossRef]
- Osipenko, S.; Zherebker, A.; Rumiantseva, L.; Kovaleva, O.; Nikolaev, E.N.; Kostyukevich, Y. Oxygen Isotope Exchange Reaction for Untargeted LC–MS Analysis. J. Am. Soc. Mass. Spectrom. 2022, 33, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Osipenko, S.; Nikolaev, E.; Kostyukevich, Y. Amine additives for improved in-ESI H/D exchange. Analyst 2022, 147, 3180–3185. [Google Scholar] [CrossRef] [PubMed]
- Kostyukevich, Y.; Sosnin, S.; Osipenko, S.; Kovaleva, O.; Rumiantseva, L.; Kireev, A.; Zherebker, A.; Fedorov, M.; Nikolaev, E.N. PyFragMS─ A Web Tool for the Investigation of the Collision-Induced Fragmentation Pathways. ACS Omega 2022, 7, 9710–9719. [Google Scholar] [CrossRef] [PubMed]
- Tupertsev, B.; Osipenko, S.; Kireev, A.; Nikolaev, E.; Kostyukevich, Y. Simple In Vitro 18O Labeling for Improved Mass Spectrometry-Based Drug Metabolites Identification: Deep Drug Metabolism Study. Int. J. Mol. Sci. 2023, 24, 4569. [Google Scholar] [CrossRef] [PubMed]
- Kostyukevich, Y.; Osipenko, S.; Rindin, K.; Zherebker, A.; Kovaleva, O.; Rumiantseva, L.; Borisova, L.; Borisova, N.; Vlaskin, M.S.; Nikolaev, E. Analysis of the Bio-oil Produced by the Hydrothermal Liquefaction of Biomass Using High-Resolution Mass Spectrometry and Isotope Exchange. Energy Fuels 2021, 35, 12208–12215. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Zherebker, A.; Orlov, A.; Kovaleva, O.; Burykina, T.; Isotov, B.; Nikolaev, E.N. Hydrogen/Deuterium and ¹⁶O/¹⁸O-Exchange Mass Spectrometry Boosting the Reliability of Compound Identification; USDA: Washington, DC, USA, 2020.
- Kostyukevich, Y.; Zherebker, A.; Vlaskin, M.S.; Roznyatovsky, V.A.; Grishin, Y.K.; Nikolaev, E. Speciation of structural fragments in crude oil by means of isotope exchange in near-critical water and Fourier transform mass spectrometry. Anal. Bioanal. Chem. 2019, 411, 3331–3339. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Vlaskin, M.; Zherebker, A.; Grigorenko, A.; Borisova, L.; Nikolaev, E. High-resolution mass spectrometry study of the bio-oil samples produced by thermal liquefaction of microalgae in different solvents. J. Am. Soc. Mass. Spectrom. 2019, 30, 605–614. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Vladimirov, G.; Stekolschikova, E.; Ivanov, D.G.; Yablokov, A.; Zherebker, A.Y.; Sosnin, S.; Orlov, A.; Fedorov, M.; Khaitovich, P.; et al. Hydrogen/Deuterium Exchange Aiding Compound Identification for LC-MS and MALDI Imaging Lipidomics. Anal. Chem. 2019, 91, 13465–13474. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Zherebker, A.; Vlaskin, M.S.; Borisova, L.; Nikolaev, E. Microprobe for the thermal analysis of crude oil coupled to photoionization Fourier transform mass spectrometry. Anal. Chem. 2018, 90, 8756–8763. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Ovchinnikov, G.; Kononikhin, A.; Popov, I.; Oseledets, I.; Nikolaev, E. Thermal dissociation and H/D exchange of streptavidin tetramers at atmospheric pressure. Int. J. Mass Spectrom. 2018, 427, 100–106. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Nikolaev, E. Ion source multiplexing on a single mass spectrometer. Anal. Chem. 2018, 90, 3576–3583. [Google Scholar] [CrossRef]
- Kostyukevich, Y.; Kononikhin, A.; Popov, I.; Nikolaev, E. Analytical description of the H/D exchange kinetic of macromolecule. Anal. Chem. 2018, 90, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Kostyukevich, Y.; Acter, T.; Zherebker, A.; Ahmed, A.; Kim, S.; Nikolaev, E. Hydrogen/deuterium exchange in mass spectrometry. Mass Spectrom. Rev. 2018, 37, 811–853. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostyukevich, Y.; Stekolshikova, E.; Levashova, A.; Kovalenko, A.; Vishnevskaya, A.; Bashilov, A.; Kireev, A.; Tupertsev, B.; Rumiantseva, L.; Khaitovich, P.; et al. Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms. Int. J. Mol. Sci. 2023, 24, 11725. https://doi.org/10.3390/ijms241411725
Kostyukevich Y, Stekolshikova E, Levashova A, Kovalenko A, Vishnevskaya A, Bashilov A, Kireev A, Tupertsev B, Rumiantseva L, Khaitovich P, et al. Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms. International Journal of Molecular Sciences. 2023; 24(14):11725. https://doi.org/10.3390/ijms241411725
Chicago/Turabian StyleKostyukevich, Yury, Elena Stekolshikova, Anna Levashova, Anna Kovalenko, Anna Vishnevskaya, Anton Bashilov, Albert Kireev, Boris Tupertsev, Lidiia Rumiantseva, Philipp Khaitovich, and et al. 2023. "Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms" International Journal of Molecular Sciences 24, no. 14: 11725. https://doi.org/10.3390/ijms241411725
APA StyleKostyukevich, Y., Stekolshikova, E., Levashova, A., Kovalenko, A., Vishnevskaya, A., Bashilov, A., Kireev, A., Tupertsev, B., Rumiantseva, L., Khaitovich, P., Osipenko, S., & Nikolaev, E. (2023). Untargeted Lipidomics after D2O Administration Reveals the Turnover Rate of Individual Lipids in Various Organs of Living Organisms. International Journal of Molecular Sciences, 24(14), 11725. https://doi.org/10.3390/ijms241411725