
L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
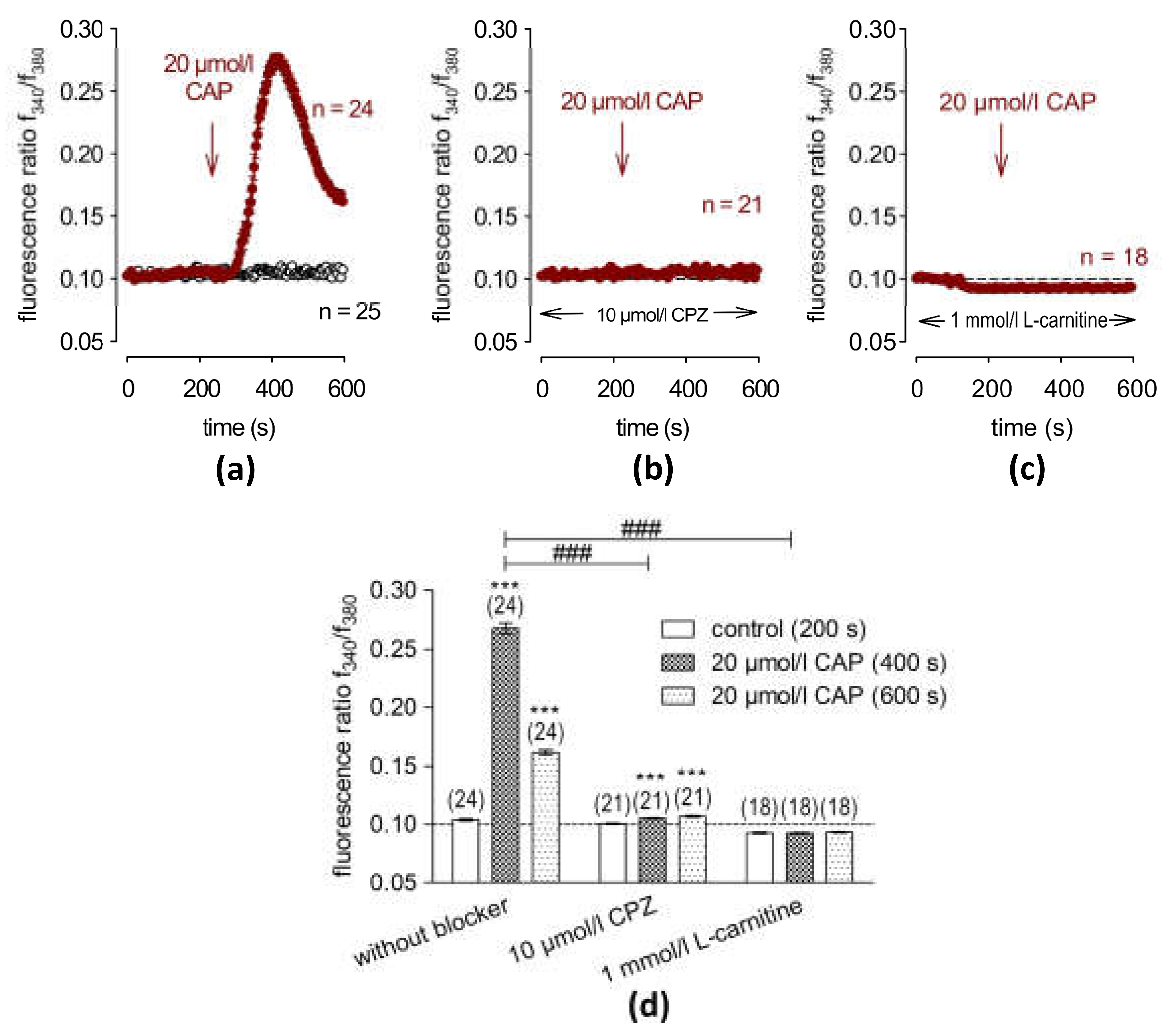
2.1. Functional Validation of TRPV1 Expression
2.2. L-carnitine Suppresses CAP-Induced Increases in Ca2+ and Whole-Cell Currents
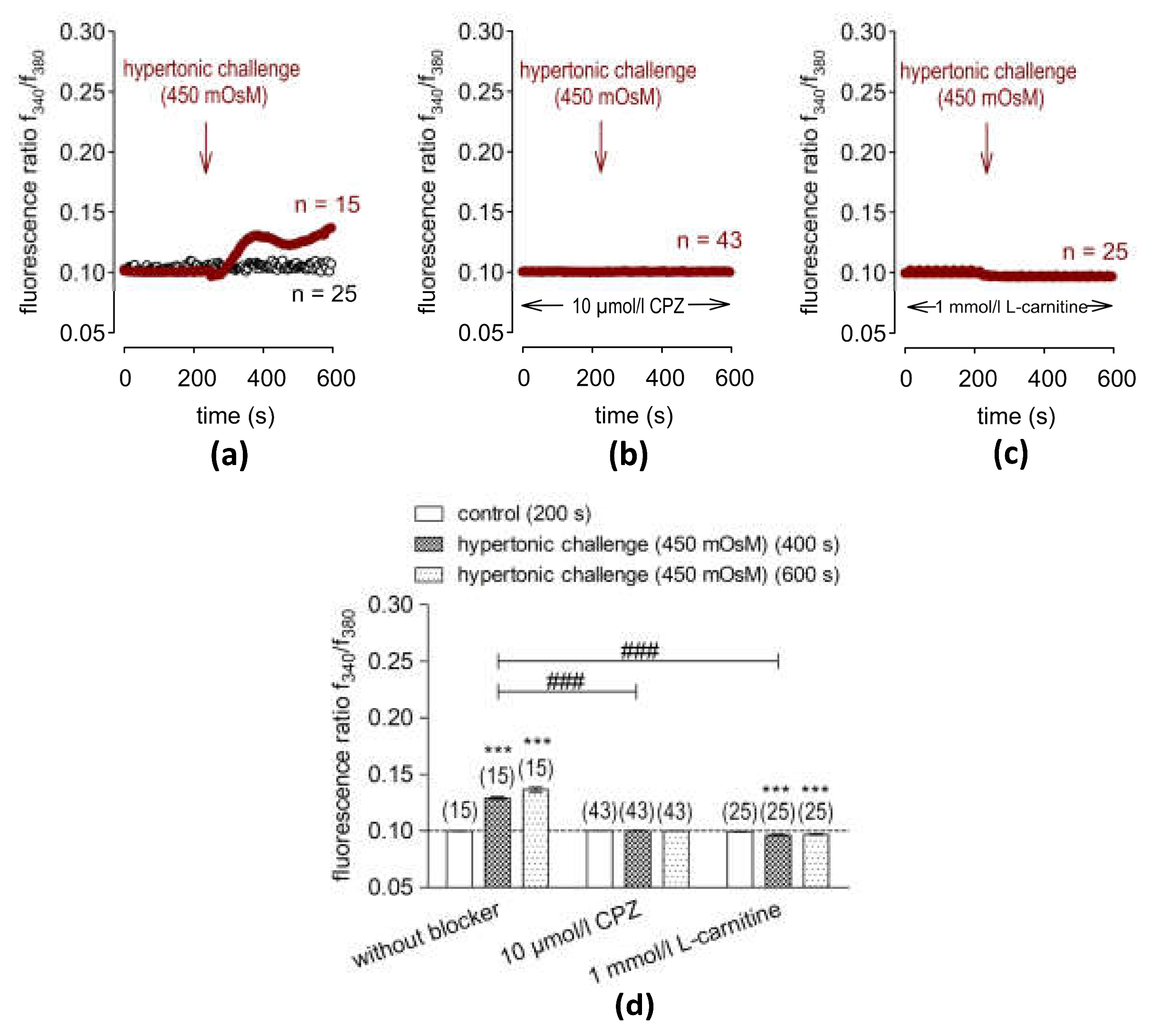
2.3. Hypertonic Challenge Activates TRPV1
2.4. L-Carnitine Suppressed Hypertonicity-Induced Increases in Intracellular Ca2+
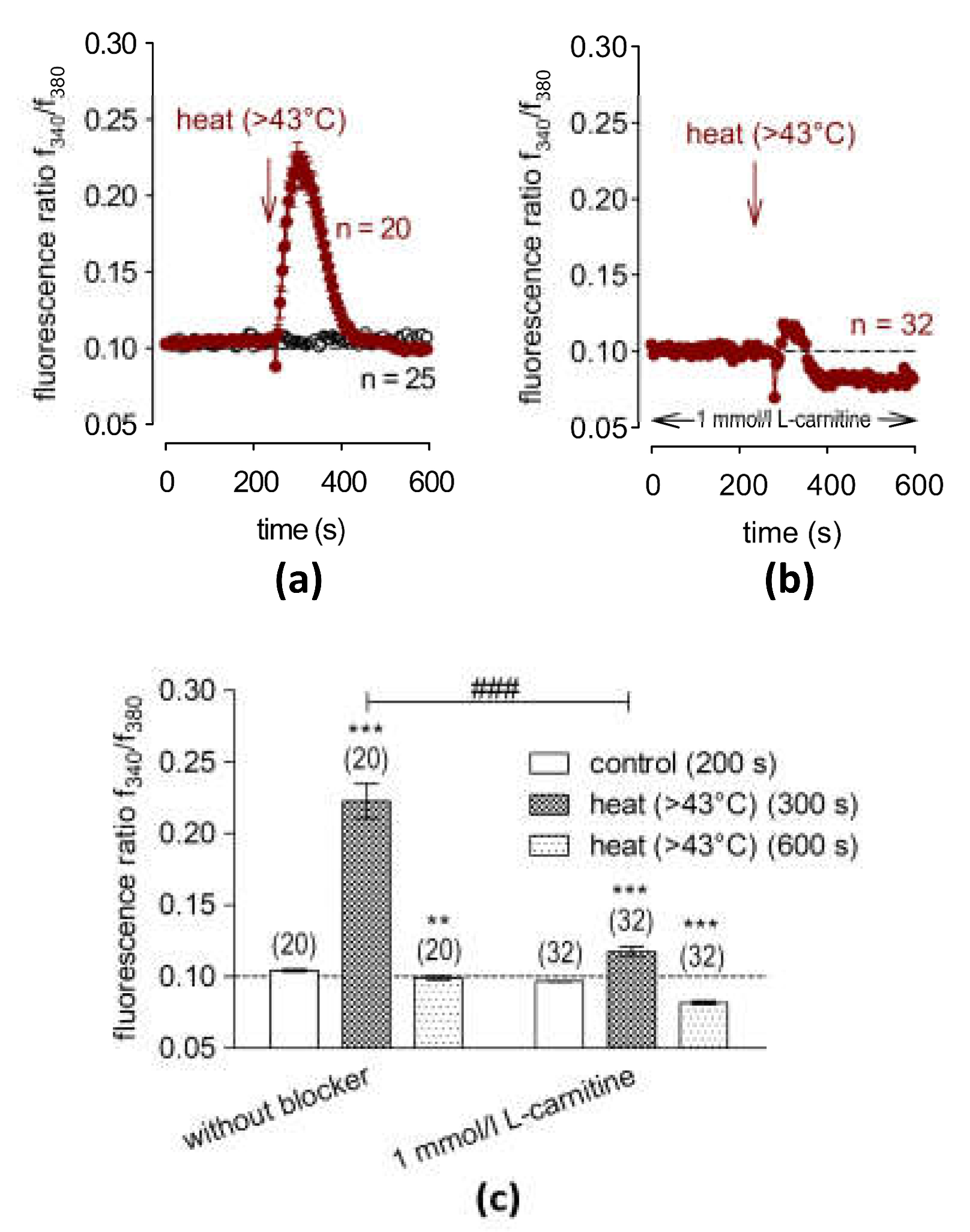
2.5. L-Carnitine Suppresses Heat-Induced Ca2+ Transients
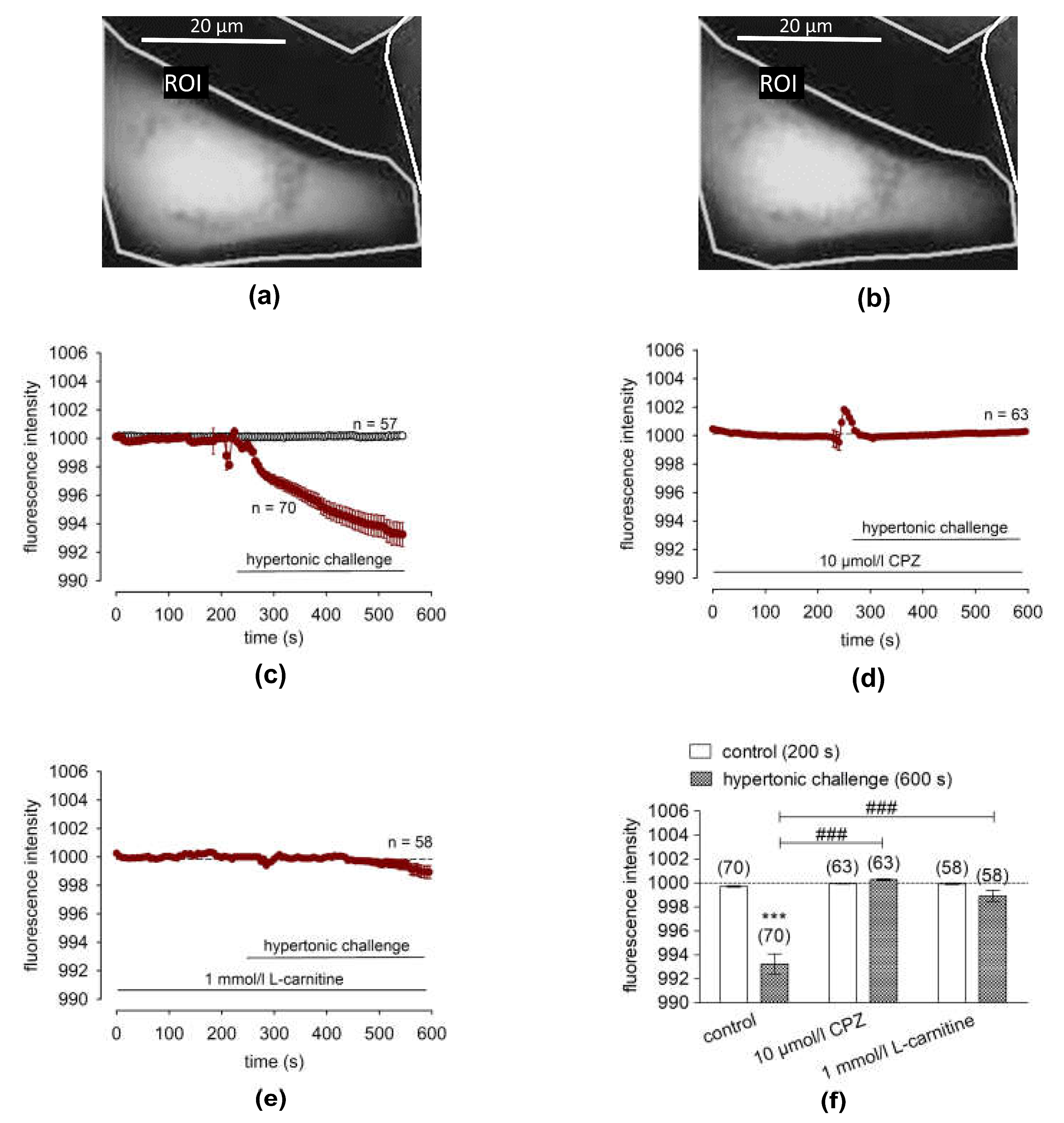
2.6. L-Carnitine Blocks Hypertonicity-Induced Corneal Epithelial Cell Shrinkage
3. Discussion
3.1. Modes of Suppression of TRPV1 Activation by L-Carnitine
3.2. L-Carnitine Suppresses Hypertonicity-Induced Cell Shrinkage and TRPV1 Activation
3.3. L-Carnitine Suppresses Heat-Induced TRPV1 Activation
3.4. L-Carnitine Suppresses Hypertonicity Induced Cell Volume Shrinkage
3.5. Clinical Impact
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Fluorescence Calcium Imaging
4.4. Apparent Cell Volume Measurements
4.5. Statistical Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Methods Concerning Supplementary Materials
Photomultiplier Fluorescence Calcium Recording
Planar Patch-Clamp Recordings
References
- Shanti, Y.; Shehada, R.; Bakkar, M.M.; Qaddumi, J. Prevalence and associated risk factors of dry eye disease in 16 northern West bank towns in Palestine: A cross-sectional study. BMC Ophthalmol. 2020, 20, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouen, P.A.; White, M.L. Dry Eye Disease: Prevalence, Assessment, and Management. Home Healthc. Now 2018, 36, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef]
- Abdulmannan, D.M.; Naser, A.Y.; Ibrahim, O.K.; Mahmood, A.S.; Alyoussef Alkrad, J.; Sweiss, K.; Alrawashdeh, H.M.; Kautsar, A.P. Visual health and prevalence of dry eye syndrome among university students in Iraq and Jordan. BMC Ophthalmol. 2022, 22, 265. [Google Scholar] [CrossRef]
- Vigo, L.; Senni, C.; Pellegrini, M.; Vagge, A.; Ferro Desideri, L.; Carones, F.; Scorcia, V.; Giannaccare, G. Effects of a New Formulation of Multiple-Action Tear Substitute on Objective Ocular Surface Parameters and Ocular Discomfort Symptoms in Patients with Dry Eye Disease. Ophthalmol. Ther. 2022, 11, 1441–1447. [Google Scholar] [CrossRef]
- de Paiva, C.S.; Pflugfelder, S.C.; Ng, S.M.; Akpek, E.K. Topical cyclosporine A therapy for dry eye syndrome. Cochrane Database Syst. Rev. 2019, 9, CD010051. [Google Scholar] [CrossRef]
- Messmer, E.M. Osmoprotection as a new therapeutic principle. Ophthalmologe 2007, 104, 987–990. [Google Scholar] [CrossRef]
- Garrett, Q.; Zhang, X.; Wang, Y.; Simmons, P.; Vehige, J.; Li, J.Y.; Chen, W. Topical administration of L-carnitine on prevention and treatment of murine dry eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, 921. [Google Scholar] [CrossRef] [Green Version]
- Kaercher, T.; Thelen, U.; Brief, G.; Morgan-Warren, R.J.; Leaback, R. A prospective, multicenter, noninterventional study of Optive Plus((R)) in the treatment of patients with dry eye: The prolipid study. Clin. Ophthalmol. 2014, 8, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Deng, R.; Li, J.; Chi, W.; Su, Z.; Lin, J.; Pflugfelder, S.C.; Li, D.Q. Protective Effects of L-Carnitine Against Oxidative Injury by Hyperosmolarity in Human Corneal Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5503–5511. [Google Scholar] [CrossRef] [Green Version]
- Bron, A.J.; Tomlinson, A.; Foulks, G.N.; Pepose, J.S.; Baudouin, C.; Geerling, G.; Nichols, K.K.; Lemp, M.A. Rethinking dry eye disease: A perspective on clinical implications. Ocul. Surf. 2014, 12, S1–S31. [Google Scholar] [CrossRef] [PubMed]
- Messmer, E.M.; Bulgen, M.; Kampik, A. Hyperosmolarity of the tear film in dry eye syndrome. Dev. Ophthalmol. 2010, 45, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Begley, C.; Chen, M.; Bradley, A.; Bonanno, J.; McNamara, N.A.; Nelson, J.D.; Simpson, T. A link between tear instability and hyperosmolarity in dry eye. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3671–3679. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Reinach, P.S.; Paulsen, F.; Pleyer, U. Thermosensitive transient receptor potential channels in human corneal epithelial cells. J. Cell. Physiol. 2011, 226, 1828–1842. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, H.; Wang, Z.; Mergler, S.; Wolosin, J.M.; Reinach, P.S. Functional TRPV1 expression in human corneal fibroblasts. Exp. Eye Res. 2013, 107, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Lyras, E.M.; Reinach, P.S.; Dwarakanath, A.; Paulsen, F.; Pleyer, U. Calcium regulation by thermo- and osmosensing transient receptor potential vanilloid channels (TRPVs) in human conjunctival epithelial cells. Histochem. Cell Biol. 2012, 137, 743–761. [Google Scholar] [CrossRef]
- Nishihara, E.; Hiyama, T.Y.; Noda, M. Osmosensitivity of transient receptor potential vanilloid 1 is synergistically enhanced by distinct activating stimuli such as temperature and protons. PLoS ONE 2011, 6, e22246. [Google Scholar] [CrossRef] [Green Version]
- Sharif Naeini, R.; Witty, M.F.; Seguela, P.; Bourque, C.W. An N-terminal variant of Trpv1 channel is required for osmosensory transduction. Nat. Neurosci. 2006, 9, 93–98. [Google Scholar] [CrossRef]
- Voets, T.; Droogmans, G.; Wissenbach, U.; Janssens, A.; Flockerzi, V.; Nilius, B. The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature 2004, 430, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Dhaka, A.; Uzzell, V.; Dubin, A.E.; Mathur, J.; Petrus, M.; Bandell, M.; Patapoutian, A. TRPV1 is activated by both acidic and basic pH. J. Neurosci. 2009, 29, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, Y.; Yang, H.; Capo-Aponte, J.E.; Tachado, S.D.; Wolosin, J.M.; Reinach, P.S. NF-kappaB feedback control of JNK1 activation modulates TRPV1-induced increases in IL-6 and IL-8 release by human corneal epithelial cells. Mol. Vis. 2011, 17, 3137–3146. [Google Scholar] [PubMed]
- Zhang, F.; Yang, H.; Wang, Z.; Mergler, S.; Liu, H.; Kawakita, T.; Tachado, S.D.; Pan, Z.; Capo-Aponte, J.E.; Pleyer, U.; et al. Transient receptor potential vanilloid 1 activation induces inflammatory cytokine release in corneal epithelium through MAPK signaling. J. Cell. Physiol. 2007, 213, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Wang, Z.; Yang, H.; Zhang, F.; Reinach, P.S. TRPV1 activation is required for hypertonicity-stimulated inflammatory cytokine release in human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Reinach, P.S.; Shirai, K.; Kitano, A.; Kao, W.W.; Flanders, K.C.; Miyajima, M.; Liu, H.; Zhang, J.; Saika, S. TRPV1 involvement in inflammatory tissue fibrosis in mice. Am. J. Pathol. 2011, 178, 2654–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Transport of L-carnitine in human corneal and conjunctival epithelial cells. Mol. Vis. 2010, 16, 1823–1831. [Google Scholar]
- Hazarbassanov, R.M.; Queiroz-Hazarbassanov, N.G.T.; Barros, J.N.; Gomes, J.A.P. Topical Osmoprotectant for the Management of Postrefractive Surgery-Induced Dry Eye Symptoms: A Randomised Controlled Double-Blind Trial. J. Ophthalmol. 2018, 2018, 4324590. [Google Scholar] [CrossRef]
- Baudouin, C.; Cochener, B.; Pisella, P.J.; Girard, B.; Pouliquen, P.; Cooper, H.; Creuzot-Garcher, C. Randomized, phase III study comparing osmoprotective carboxymethylcellulose with sodium hyaluronate in dry eye disease. Eur. J. Ophthalmol. 2012, 22, 751–761. [Google Scholar] [CrossRef]
- Evangelista, M.; Koverech, A.; Messano, M.; Pescosolido, N. Comparison of three lubricant eye drop solutions in dry eye patients. Optom. Vis. Sci. 2011, 88, 1439–1444. [Google Scholar] [CrossRef]
- Wojtowicz, J.C.; Arciniega, J.C.; McCulley, J.P.; Mootha, V.V. Effect of systane and optive on aqueous tear evaporation in patients with dry eye disease. Eye Contact Lens 2010, 36, 358–360. [Google Scholar] [CrossRef]
- Kaercher, T.; Buchholz, P.; Kimmich, F. Treatment of patients with keratoconjunctivitis sicca with Optive: Results of a multicenter, open-label observational study in Germany. Clin. Ophthalmol. 2009, 3, 33–39. [Google Scholar] [PubMed]
- Hardten, D.R.; Brown, M.J.; Pham-Vang, S. Evaluation of an isotonic tear in combination with topical cyclosporine for the treatment of ocular surface disease. Curr. Med. Res. Opin. 2007, 23, 2083–2091. [Google Scholar] [CrossRef] [PubMed]
- Nebbioso, M.; Evangelista, M.; Librando, A.; Plateroti, A.M.; Pescosolido, N. Iatrogenic dry eye disease: An eledoisin/carnitine and osmolyte drops study. Biomed. Pharmacother. 2013, 67, 659–663. [Google Scholar] [CrossRef]
- Khajavi, N.; Reinach, P.S.; Skrzypski, M.; Lude, A.; Mergler, S. L-carnitine reduces in human conjunctival epithelial cells hypertonic-induced shrinkage through interacting with TRPV1 channels. Cell. Physiol. Biochem. 2014, 34, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Turan, E.; Valtink, M.; Reinach, P.S.; Skupin, A.; Luo, H.; Brockmann, T.; Ba Salem, M.H.O.; Pleyer, U.; Mergler, S. L-carnitine suppresses transient receptor potential vanilloid type 1 activity and myofibroblast transdifferentiation in human corneal keratocytes. Lab. Investig. 2021, 101, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef]
- Chu, Y.; Cohen, B.E.; Chuang, H.H. A single TRPV1 amino acid controls species sensitivity to capsaicin. Sci. Rep. 2020, 10, 8038. [Google Scholar] [CrossRef]
- Sullivan, C.; Lee, J.; Bushey, W.; Demers, D.; Dinsdale, S.; Lowe, K.; Olmeda, J.; Meng, I.D. Evidence for a phenotypic switch in corneal afferents after lacrimal gland excision. Exp. Eye Res. 2022, 218, 109005. [Google Scholar] [CrossRef]
- Wan, S.J.; Datta, A.; Flandrin, O.; Metruccio, M.M.E.; Ma, S.; Nieto, V.; Kroken, A.R.; Hill, R.Z.; Bautista, D.M.; Evans, D.J.; et al. Nerve-associated transient receptor potential ion channels can contribute to intrinsic resistance to bacterial adhesion in vivo. FASEB J. 2021, 35, e21899. [Google Scholar] [CrossRef]
- Jiao, H.; Ivanusic, J.J.; McMenamin, P.G.; Chinnery, H.R. Distribution of Corneal TRPV1 and Its Association With Immune Cells During Homeostasis and Injury. Investig. Ophthalmol. Vis. Sci. 2021, 62, 6. [Google Scholar] [CrossRef]
- Pan, Z.; Yang, H.; Mergler, S.; Liu, H.; Tachado, S.D.; Zhang, F.; Kao, W.W.; Koziel, H.; Pleyer, U.; Reinach, P.S. Dependence of regulatory volume decrease on transient receptor potential vanilloid 4 (TRPV4) expression in human corneal epithelial cells. Cell Calcium. 2008, 44, 374–385. [Google Scholar] [CrossRef] [Green Version]
- Khandekar, N.; Willcox, M.D.; Shih, S.; Simmons, P.; Vehige, J.; Garrett, Q. Decrease in hyperosmotic stress-induced corneal epithelial cell apoptosis by L-carnitine. Mol. Vis. 2013, 19, 1945–1956. [Google Scholar] [PubMed]
- Huo, J.; Xie, H.P.; Li, J.; Zhou, D. A primary study of L-carnitine protective effect on corneal and conjunctival epithelium of mouse dry eye model induced by hyperosmolar saline. Zhonghua Yan Ke Za Zhi 2012, 48, 330–336. [Google Scholar] [PubMed]
- Tominaga, M.; Caterina, M.J. Thermosensation and pain. J. Neurobiol. 2004, 61, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Yu, H.S.; Lin, H.C.; Wu, K.Y.; Yang, R.C.; Kuo, P.L. Heat shock induces apoptosis through reactive oxygen species involving mitochondrial and death receptor pathways in corneal cells. Exp. Eye Res. 2011, 93, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Numata, T.; Shimizu, T.; Okada, Y. TRPM7 is a stretch- and swelling-activated cation channel involved in volume regulation in human epithelial cells. Am. J. Physiol. Cell Physiol. 2007, 292, C460–C467. [Google Scholar] [CrossRef] [Green Version]
- Benfenati, V.; Caprini, M.; Dovizio, M.; Mylonakou, M.N.; Ferroni, S.; Ottersen, O.P.; Amiry-Moghaddam, M. An aquaporin-4/transient receptor potential vanilloid 4 (AQP4/TRPV4) complex is essential for cell-volume control in astrocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 2563–2568. [Google Scholar] [CrossRef]
- Baudouin, C.; Aragona, P.; Messmer, E.M.; Tomlinson, A.; Calonge, M.; Boboridis, K.G.; Akova, Y.A.; Geerling, G.; Labetoulle, M.; Rolando, M. Role of hyperosmolarity in the pathogenesis and management of dry eye disease: Proceedings of the OCEAN group meeting. Ocul. Surf. 2013, 11, 246–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versura, P.; Profazio, V.; Campos, E.C. Performance of tear osmolarity compared to previous diagnostic tests for dry eye diseases. Curr. Eye Res. 2010, 35, 553–564. [Google Scholar] [CrossRef]
- Pescosolido, N.; Imperatrice, B.; Koverech, A.; Messano, M. L-carnitine and short chain ester in tears from patients with dry eye. Optom. Vis. Sci. 2009, 86, E132–E138. [Google Scholar] [CrossRef]
- Messmer, E.M. The pathophysiology, diagnosis, and treatment of dry eye disease. Dtsch. Arztebl. Int. 2015, 112, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, F.A.; Chaudhry, I.A.; Boulton, M.E.; Al-Rajhi, A.A. L-carnitine protects human retinal pigment epithelial cells from oxidative damage. Curr. Eye Res. 2007, 32, 575–584. [Google Scholar] [CrossRef]
- Corrales, R.M.; Luo, L.; Chang, E.Y.; Pflugfelder, S.C. Effects of osmoprotectants on hyperosmolar stress in cultured human corneal epithelial cells. Cornea 2008, 27, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Fakih, D.; Guerrero-Moreno, A.; Baudouin, C.; Reaux-Le Goazigo, A.; Parsadaniantz, S.M. Capsazepine decreases corneal pain syndrome in severe dry eye disease. J. Neuroinflamm. 2021, 18, 111. [Google Scholar] [CrossRef] [PubMed]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Su, Z.; Deng, R.; Lin, J.; Li, D.Q.; Pflugfelder, S.C. Effects of L-carnitine, erythritol and betaine on pro-inflammatory markers in primary human corneal epithelial cells exposed to hyperosmotic stress. Curr. Eye Res. 2015, 40, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Capo-Aponte, J.E.; Iserovich, P.; Reinach, P.S. Characterization of regulatory volume behavior by fluorescence quenching in human corneal epithelial cells. J. Membr. Biol. 2005, 207, 11–22. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucius, A.; Chhatwal, S.; Valtink, M.; Reinach, P.S.; Li, A.; Pleyer, U.; Mergler, S. L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 11815. https://doi.org/10.3390/ijms241411815
Lucius A, Chhatwal S, Valtink M, Reinach PS, Li A, Pleyer U, Mergler S. L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells. International Journal of Molecular Sciences. 2023; 24(14):11815. https://doi.org/10.3390/ijms241411815
Chicago/Turabian StyleLucius, Alexander, Sirjan Chhatwal, Monika Valtink, Peter S. Reinach, Aruna Li, Uwe Pleyer, and Stefan Mergler. 2023. "L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells" International Journal of Molecular Sciences 24, no. 14: 11815. https://doi.org/10.3390/ijms241411815
APA StyleLucius, A., Chhatwal, S., Valtink, M., Reinach, P. S., Li, A., Pleyer, U., & Mergler, S. (2023). L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells. International Journal of Molecular Sciences, 24(14), 11815. https://doi.org/10.3390/ijms241411815