

Trypanosoma cruzi Secreted Cyclophilin TcCyP19 as an Early Marker for Trypanocidal Treatment Efficiency

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
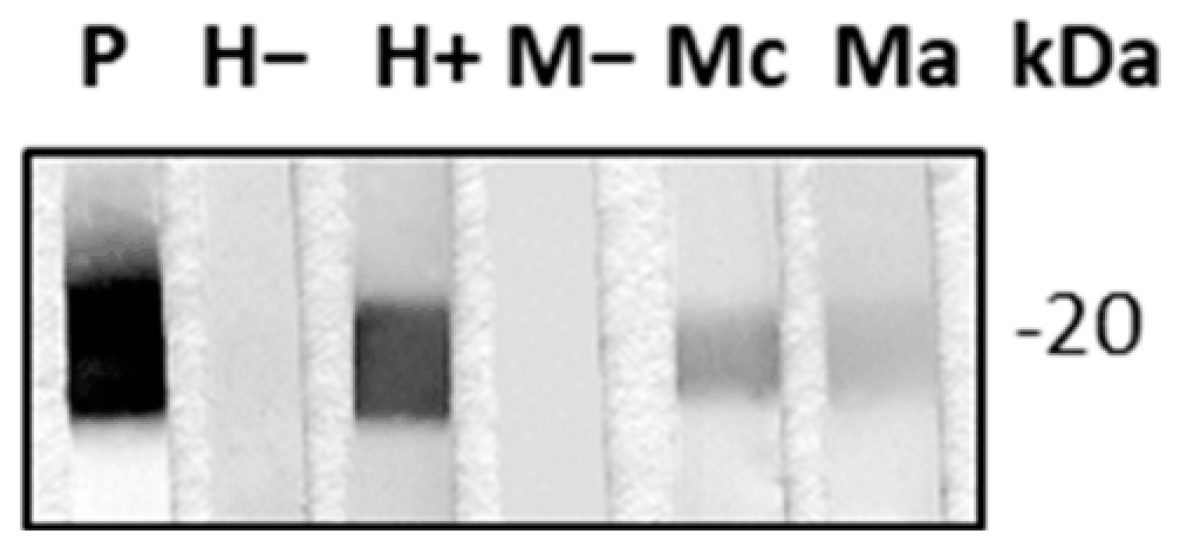
2.1. TcCyP19 Cyclophilin Is Secreted in the Extracellular Environment
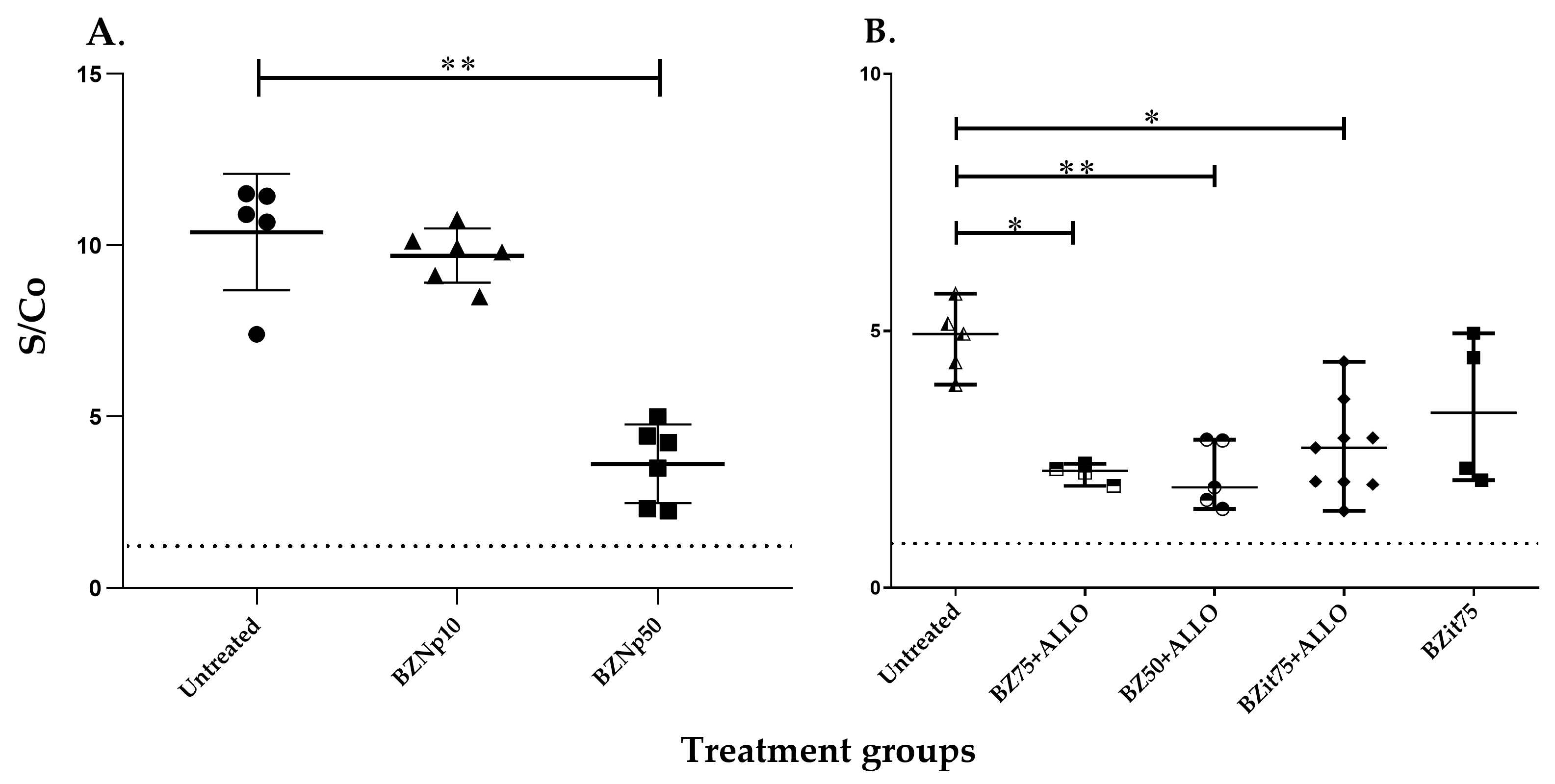
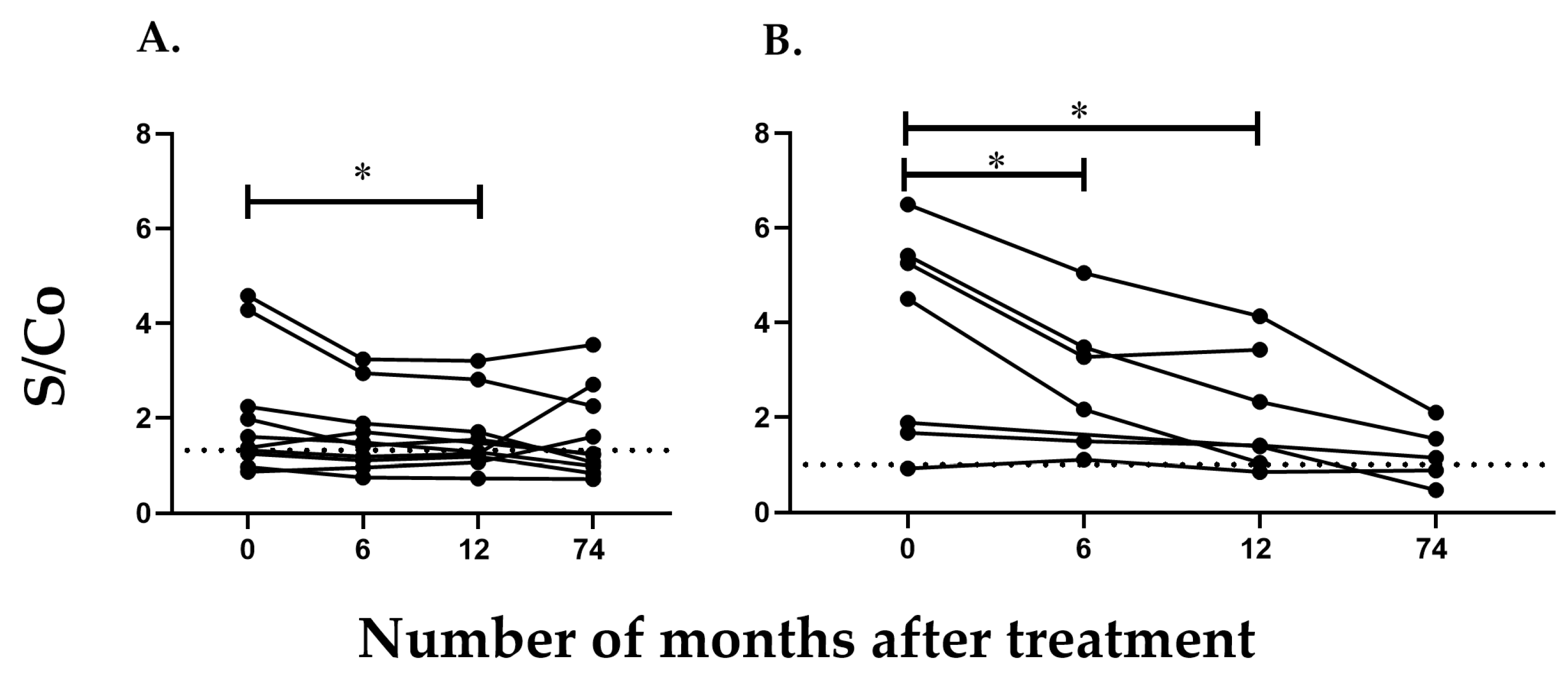
2.2. Detection of Antibodies against TcCyP19 Protein in T. cruzi-Infected Mice
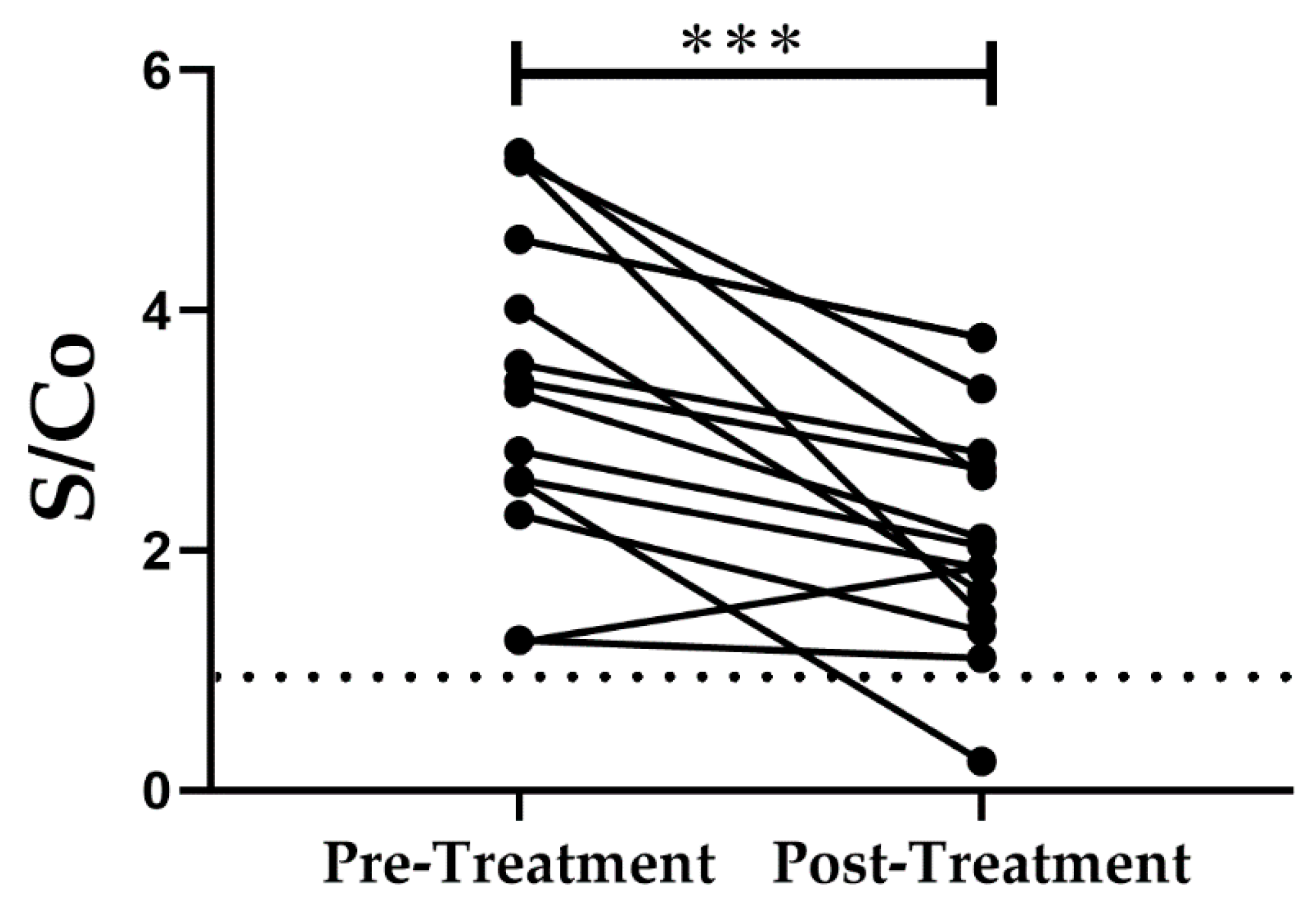
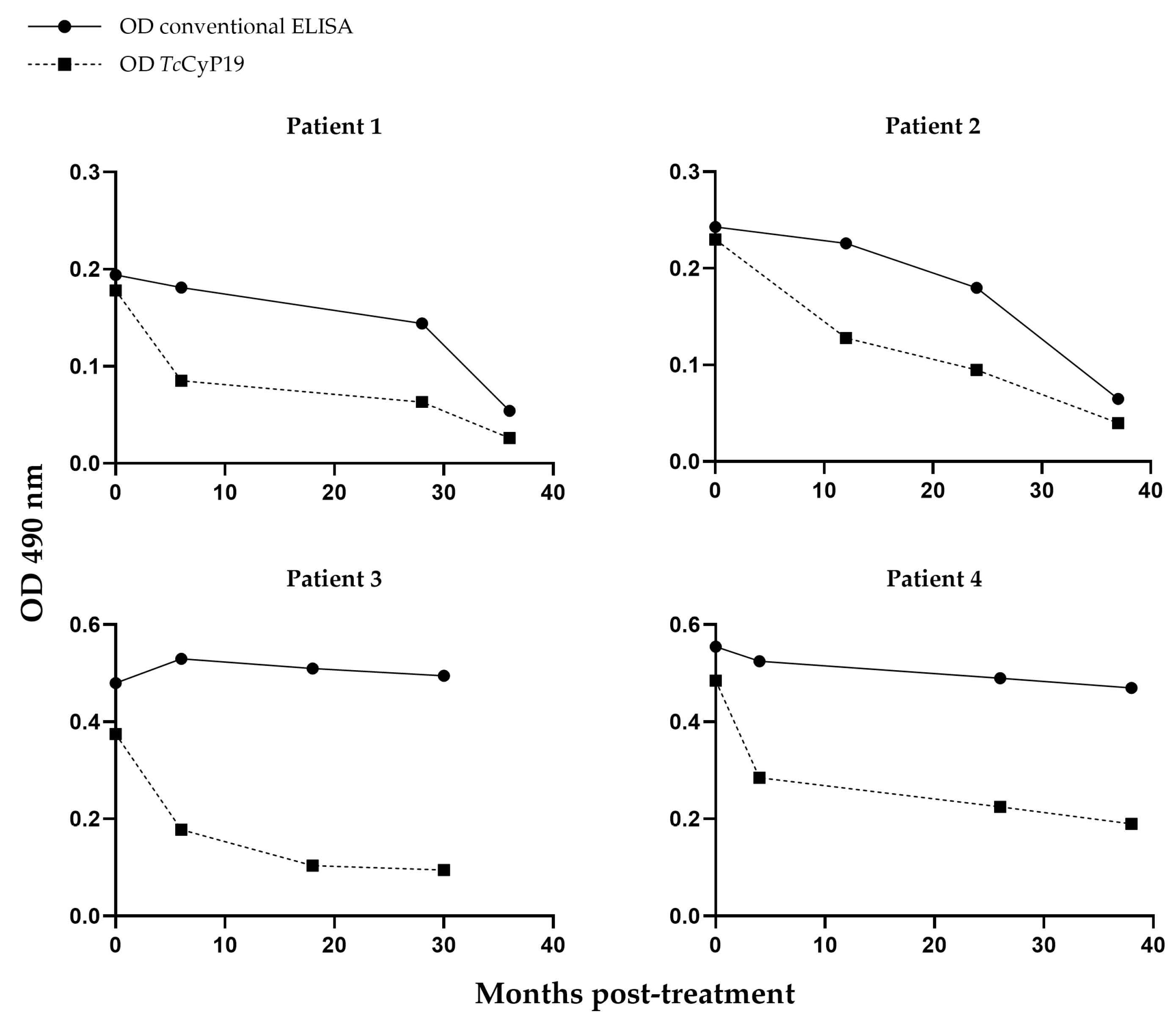
2.3. Detection of Antibodies against TcCyP19 Protein in Chronically Infected T. cruzi Patients
3. Discussion
4. Materials and Methods
4.1. Parasites
4.2. Expression and Purification of T. cruzi CyP19 Recombinant Protein
4.3. Obtention and Purification of Polyclonal Antibodies against TcCyP19
4.4. Sera from Mice Infected with T. cruzi and Treated in the Acute Phase
4.5. Sera from Mice Infected with T. cruzi and Treated in the Chronic Phase
4.6. Sera from Patients Infected with T. cruzi and Treated in the Chronic Phase
4.7. Immuno-Enzymatic Analysis of Proteins Electrotransferred to Nitrocellulose Membranes (Western Blot)
4.8. In-House ELISA for Evaluation of Anti-TcCyP19 Antibodies (ELISA-TcCyP19)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar]
- Bern, C.; Verastegui, M.; Gilman, R.H.; Lafuente, C.; Galdos-Cardenas, G.; Calderon, M.; Pacori, J.; Del Carmen Abastoflor, M.; Aparicio, H.; Brady, M.F.; et al. Congenital Trypanosoma cruzi transmission in Santa Cruz, Bolivia. Clin. Infect. Dis. 2009, 49, 1667–1674. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Chagas Disease (American Trypanosomiasis). Available online: https://www.who.int/health-topics/chagas-disease (accessed on 20 May 2023).
- Rassi, A., Jr.; Rassi, A.; Marcondes de Rezende, J. American trypanosomiasis (Chagas disease). Lancet 2012, 26, 275–291. [Google Scholar]
- Zacks, M.A.; Wen, J.J.; Vyatkina, G.; Bhatia, V.; Garg, N. An overview of chagasic cardiomyopathy: Pathogenic importance of oxidative stress. Acad. Bras. Cienc. 2005, 77, 695–715. [Google Scholar]
- Viotti, R.; Vigliano, C.; Álvarez, M.G.; Lococo, B.; Petti, M.; Bertocchi, G.; Armenti, A.; De Rissio, A.M.; Cooley, G.; Tarleton, R.; et al. Impact of Aetiological Treatment on Conventional and Multiplex Serology in Chronic Chagas Disease. PLoS Negl. Trop. Dis. 2011, 5, e1314. [Google Scholar] [CrossRef]
- Altcheh, J.; Castro, L.; Dib, J.C.; Grossmann, U.; Huang, E.; Moscatelli, G.; Rocha, J.J.P.; Ramírez, T.E.; on behalf of the CHICO Study Group. Prospective, historically controlled study to evaluate the efficacy and safety of a new paediatric formulation of nifurtimox in children aged 0 to 17 years with Chagas disease one year after treatment (CHICO). PLoS Negl. Trop. Dis. 2021, 15, e0008912. [Google Scholar] [CrossRef]
- Pan American Health Organization. Guidelines for the Diagnosis and Treatment of Chagas Disease. Available online: http://iris.paho.org/xmlui/bitstream/handle/123456789/49653/9789275120439_eng.pdf?sequence=6&isAllowed=y (accessed on 20 May 2023).
- Morillo, C.A.; Marin-Neto, J.A.; Avezum, A.; Sosa-Estani, S.; Rassi, A., Jr.; Rosas, F.; Villena, E.; Quiroz, R.; Bonilla, R.; Britto, C.; et al. Randomized Trial of Benznidazole for Chronic Chagas’ Cardiomyopathy. N. Engl. J. Med. 2015, 373, 1295–1306. [Google Scholar]
- Pinazo, M.-J.; Guerrero, L.; Posada, E.; Rodríguez, E.; Soy, D.; Gascon, J. Benznidazole-Related Adverse Drug Reactions and Their Relationship to Serum Drug Concentrations in Patients with Chronic Chagas Disease. Antimicrob. Agents Chemother. 2013, 57, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Docampo, R.; Moreno, S.N.J. Free Radical Metabolites in the Mode of Action of Chemotherapeutic Agents and Phagocytic Cells on Trypanosoma cruzi. Clin. Infect. Dis. 1984, 6, 223–238. [Google Scholar] [CrossRef]
- Viotti, R.; Vigliano, C.; Lococo, B.; Alvarez, M.G.; Petti, M.; Bertocchi, G.; Armenti, A. Side effects of benznidazole as treatment in chronic Chagas disease: Fears and realities. Expert. Rev. Anti-Infect. Ther. 2009, 7, 157–163. [Google Scholar] [CrossRef]
- Andrade, A.L.; Martelli, C.M.; Oliveira, R.M.; Silva, S.A.; Aires, A.I.; Soussumi, L.M.; Covas, D.T.; Silva, L.S.; Andrade, J.G.; Travassos, L.R.; et al. Short report: Benznidazole efficacy among Trypanosoma cruzi-infected adolescents after a six-year follow-up. Am. J. Trop. Med. Hyg. 2004, 71, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Sosa Estani, S.; Segura, E.L.; Ruiz, A.M.; Velazquez, E.; Porcel, B.M.; Yampotis, C. Efficacy of chemotherapy with benznidazole in children in the indeterminate phase of Chagas’ disease. Am. J. Trop. Med. Hyg. 1998, 59, 526–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albareda, M.C.; Natale, M.A.; De Rissio, A.M.; Fernandez, M.; Serjan, A.; Alvarez, M.G.; Cooley, G.; Shen, H.; Viotti, R.; Bua, J.; et al. Distinct Treatment Outcomes of Antiparasitic Therapy in Trypanosoma cruzi-Infected Children Is Associated With Early Changes in Cytokines, Chemokines, and T-Cell Phenotypes. Front. Immunol. 2018, 9, 1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viotti, R.; Vigliano, C.; Lococo, B.; Bertocchi, G.; Petti, M.; Alvarez, M.G.; Postan, M.; Armenti, A. Long-term cardiac outcomes of treating chronic Chagas disease with benznidazole versus no treatment: A nonrandomized trial. Ann. Intern. Med. 2006, 144, 724–734. [Google Scholar] [CrossRef]
- Scalise, M.L.; Arrúa, E.C.; Rial, M.S.; Esteva, M.I.; Salomon, C.J.; Fichera, L.E. Promising Efficacy of Benznidazole Nanoparticles in Acute Trypanosoma cruzi Murine Model: In-Vitro and In-Vivo Studies. Am. J. Trop. Med. Hyg. 2016, 95, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Rial, M.S.; Scalise, M.L.; Arrúa EEsteva, M.I.; Salomon, C.J.; Fichera, L.E. Elucidating the impact of low doses of nano-formulated benznidazole in acute experimental Chagas disease. PLoS Negl. Trop. Dis. 2017, 11, e0006119. [Google Scholar] [CrossRef] [Green Version]
- Rial, M.S.; Scalise, M.L.; López Alarcón, M.; Esteva, M.I.; Bua, J.; Benatar, A.F.; Prado, N.G.; Riarte, A.R.; Fichera, L.E. Experimental combination therapy using low doses of benznidazole and allopurinol in mouse models of Trypanosoma cruzi chronic infection. Parasitology 2019, 146, 305–313. [Google Scholar] [CrossRef]
- Rial, M.S.; Seremeta, K.P.; Esteva, M.I.; Búa, J.; Salomon, C.J.; Fichera, L.E. In vitro studies and preclinical evaluation of benznidazole microparticles in the acute Trypanosoma cruzi murine model. Parasitology 2021, 148, 566–575. [Google Scholar] [CrossRef]
- Handschumacher, R.E.; Harding, M.W.; Rice, J.; Drugge, R.J.; Speicher, D.W. Cyclophilin: A specific cytosolic binding protein for cyclosporin A. Science 1984, 226, 544–547. [Google Scholar] [CrossRef]
- Takahashi, N.; Hayano, T.; Suzuki, M. Peptidyl-prolyl cis-trans isomerase is the cyclosporin A-binding protein cyclophilin. Nature 1989, 337, 473–475. [Google Scholar] [CrossRef]
- Galat, A.; Bua, J. Molecular aspects of cyclophilins mediating therapeutic actions of their ligands. Cell Mol. Life Sci. 2010, 67, 3467–3488. [Google Scholar] [CrossRef] [PubMed]
- Potenza, M.; Galat, A.; Minning, T.A.; Ruiz, A.M.; Duran, R.; Tarleton, R.L.; Marín, M.; Fichera, L.E.; Bua, J. Analysis of the Trypanosoma cruzi cyclophilin gene family and identification of Cyclosporin A binding proteins. Parasitology 2006, 132 Pt 6, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Bua, J.; Aslund, L.; Pereyra, N.; García, G.A.; Bontempi, E.J.; Ruiz, A.M. Characterisation of a cyclophilin isoform in Trypanos. Cruzi. FEMS Microbiol. Lett. 2001, 200, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bua, J.; Ruiz, A.M.; Potenza, M.; Fichera, L.E. In vitro anti-parasitic activity of Cyclosporin A analogs on Trypanosoma cruzi. Bioorg Med. Chem. Lett. 2004, 14, 4633–4637. [Google Scholar] [CrossRef]
- Bua, J.; Fichera, L.E.; Fuchs, A.G.; Potenza, M.; Dubin, M.; Wenger, R.O.; Moretti, G.; Scabone, C.M.; Ruiz, A.M. Anti-Trypanosoma cruzi effects of cyclosporin A derivatives: Possible role of a P-glycoprotein and parasite cyclophilins. Parasitology 2008, 135, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Bayer-Santos, E.; Aguilar-Bonavides, C.; Rodrigues, S.P.; Cordero, E.M.; Marques, A.F.; Varela-Ramirez, A.; Choi, H.; Yoshida, N.; da Silveira, J.F.; Almeida, I.C. Proteomic analysis of Trypanosoma cruzi secretome: Characterization of two populations of extracellular vesicles and soluble proteins. J. Proteome Res. 2013, 12, 883–897. [Google Scholar] [CrossRef]
- Kulkarni, M.M.; Karafova, A.; Kamysz, W.; Schenkman, S.; Pelle, R.; McGwire, B.S. Secreted trypanosome cyclophilin inactivates lytic insect defense peptides and induces parasite calcineurin activation and infectivity. J. Biol. Chem. 2013, 288, 8772–8784. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, G.P.; Abukawa, F.M.; Souza-Melo, N.; Alcântara, L.M.; Bittencourt-Cunha, P.; Moraes, C.B.; Jha, B.K.; McGwire, B.S.; Moretti, N.S.; Schenkman, S. Cyclophilin 19 secreted in the host cell cytosol by Trypanosoma cruzi promotes ROS production required for parasite growth. Cell Microbiol. 2021, 23, e13295. [Google Scholar]
- Jha, B.K.; Varikuti, S.; Verma, C.; Shivahare, R.; Bishop, N.; Dos Santos, G.P.; McDonald, J.; Sur, A.; Myler, P.J.; Schenkman, S.; et al. Immunization with a Trypanosoma cruzi cyclophilin-19 deletion mutant protects against acute Chagas disease in mice. NPJ Vaccines 2023, 8, 63. [Google Scholar] [CrossRef]
- Hoffmann, H.; Schiene-Fischer, C. Functional aspects of extracellular cyclophilins. Biol. Chem. 2014, 395, 721–735. [Google Scholar] [CrossRef]
- Pellé, R.; McOdimba, F.; Chuma, F.; Wasawo, D.; Pearson, T.W.; Murphy, N.B. The African trypanosome cyclophilin A homologue contains unusual conserved central and N-terminal domains and is developmentally regulated. Gene 2002, 290, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Nagarkatti, R.; Acosta, D.; Acharyya, N.; de Araujo, F.F.; Elói-Santos, S.M.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Debrabant, A. A novel Trypanosoma cruzi secreted antigen as a potential biomarker of Chagas disease. Sci. Rep. 2020, 10, 19591. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.S.; Jung, E.S.; Kim, J.H.; Jung, Y.K.; Kim, J.H.; An, H.; Yim, H.J.; Yeon, J.E.; Byun, K.S.; Kim, C.D.; et al. Significance of anti-HCV signal-to-cutoff ratio in predicting hepatitis C viremia. Korean J. Intern. Med. 2009, 24, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Ballani, N.; Gupta, E. Low signal-to-cutoff ratio (S/Co) in the diagnosis of hepatitis C: A diagnostic dilemma? Indian. J. Gastroenterol. 2015, 34, 413–414. [Google Scholar] [CrossRef] [PubMed]
- White DA, E.; Anderson, E.S.; Basham, K.; Ng, V.L.; Russell, C.; Lyons, M.S.; Powers-Fletcher, M.V.; Giordano, T.P.; Muldrew, K.L.; Siatecka, H.; et al. Clinical Utility of the Signal-to-Cutoff Ratio of Reactive HIV Antigen/Antibody Screening Tests in Guiding Emergency Physician Management. J. Acquir. Immune Defic. Syndr. 2022, 89, 332–339. [Google Scholar] [CrossRef]
- Fabbro, D.L.; Streiger, M.L.; Arias, E.D.; Bizai, M.L.; del Barco, M.; Amicone, N.A. Trypanocide treatment among adults with chronic Chagas disease living in Santa Fe city (Argentina), over a mean follow-up of 21 years: Parasitological, serological and clinical evolution. Rev. Soc. Bras. Med. Trop. 2007, 40, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Machado-de-Assis, G.F.; Diniz, G.A.; Montoya, R.A.; Dias JC, P.; Coura, J.R.; Machado-Coelho GL, L.; Albajar-Viñas, P.; Morais Torres, R.; de Lana, M. A serological, parasitological and clini-cal evaluation of untreated Chagas disease patients and those treated with benznidazole before and thirteen years after intervention. Mem. Inst. Oswaldo Cruz 2013, 108, 873–880. [Google Scholar] [CrossRef]
- Castro, A.M.; Luquetti, A.O.; Rassi, A.; Chiari, E.; da Cunha Galvão, L.M. Detection of parasitemia profiles by blood culture after treatment of human chronic Trypanosoma cruzi infection. Parasitol. Res. 2006, 99, 379–383. [Google Scholar] [CrossRef]
- Pinazo, M.J.; Thomas, M.C.; Bua, J.; Perrone, A.; Schijman, A.G.; Viotti, R.J.; Ramsey, J.M.; Ribeiro, I.; Sosa-Estani, S.; López, M.C.; et al. Biological markers for evaluating therapeutic efficacy in Chagas disease, a systematic review. Expert. Rev. Anti-Infect. Ther. 2014, 12, 479–496. [Google Scholar] [CrossRef]
- Fernandez-Villegas, A.; Pinazo, M.J.; Maranon, C.; Thomas, M.C.; Posada, E.; Carrilero, B.; Segovia, M.; Gascon, J.; Lopez, M.C. Short-term follow-up of chagasic patients after benzonidazole treatment using multiple serological markers. BMC Infect. Dis. 2011, 11, 206. [Google Scholar] [CrossRef] [Green Version]
- Floridia-Yapur, N.; Monje-Rumi, M.; Ragone, P.; Lauthier, J.J.; Tomasini, N.; Alberti D′Amato, A.; Diosque, P.; Cimino, R.; Gil, J.F.; Sanchez, D.O.; et al. TcTASV Antigens of Trypanosoma cruzi: Utility for Diagnosis and High Accuracy as Biomarkers of Treatment Efficacy in Pediatric Patients. Am. J. Trop. Med. Hyg. 2019, 101, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Pinazo Delgado, M.J.; Gascón, J. (Eds.) Chagas Disease; Springer Nature: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Fabbro, D.; Velazquez, E.; Bizai, M.L.; Denner, S.; Olivera, V.; Arias, E.; Pravia, C.; Ruiz, A.M. Evaluation of the ELISA-F29 test as an early marker of therapeutic efficacy in adults with chronic Chagas disease. Rev. Inst. Med. Trop. Sao Paulo 2013, 55, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Padilla, J.; López, M.C.; Esteva, M.; Zrein, M.; Casellas, A.; Gómez, I.; Granjon, E.; Méndez, S.; Benítez, C.; Ruiz, A.M.; et al. Serological reactivity against T. cruzi-derived antigens: Evaluation of their suitability for the assessment of response to treatment in chronic Chagas disease. Acta Trop. 2021, 221, 105990. [Google Scholar] [CrossRef] [PubMed]
- Rivero, R.; Esteva, M.; Huang, E.; Colmegna, L.; Altcheh, J.; Grossmann, U.; Ruiz, A.M. ELISA F29—A therapeutic efficacy biomarker in Chagas disease: Evaluation in pediatric patients treated with nifurtimox and followed for 4 years post-treatment. PLoS Negl. Trop. Dis. 2023; in press. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Sex (%) | Treatment | Mean Age in Years (Range) | Mean Time (Months) of Sampling after Treatment (Range) | ||||
|---|---|---|---|---|---|---|---|---|
| Female | Male | BZN (%) | Nifurtimox (%) | |||||
| Adults | 16 | 12/16 (75) | 4/16 (25) | 15/16 (94) | 1/16 (6) | 38.9 (22–48) | 11.9 (2–39) | |
| Children | Group A | 10 | 5/10 (50) | 5/10 (50) | 9/10 (90) | 1/10 (10) | 8.3 (6–14) | 73 (48–120) |
| Group B | 7 | 3/7 (43) | 4/7 (57) | 6/7 (86) | 1/7 (14) | 12.6 (1116) | 59.7 (48–73) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perrone, A.E.; Pinillo, M.; Rial, M.S.; Fernández, M.; Milduberger, N.; González, C.; Bustos, P.L.; Fichera, L.E.; Laucella, S.A.; Albareda, M.C.; et al. Trypanosoma cruzi Secreted Cyclophilin TcCyP19 as an Early Marker for Trypanocidal Treatment Efficiency. Int. J. Mol. Sci. 2023, 24, 11875. https://doi.org/10.3390/ijms241511875
Perrone AE, Pinillo M, Rial MS, Fernández M, Milduberger N, González C, Bustos PL, Fichera LE, Laucella SA, Albareda MC, et al. Trypanosoma cruzi Secreted Cyclophilin TcCyP19 as an Early Marker for Trypanocidal Treatment Efficiency. International Journal of Molecular Sciences. 2023; 24(15):11875. https://doi.org/10.3390/ijms241511875
Chicago/Turabian StylePerrone, Alina E., Mariana Pinillo, Marcela S. Rial, Marisa Fernández, Natalia Milduberger, Carolina González, Patricia L. Bustos, Laura E. Fichera, Susana A. Laucella, María Cecilia Albareda, and et al. 2023. "Trypanosoma cruzi Secreted Cyclophilin TcCyP19 as an Early Marker for Trypanocidal Treatment Efficiency" International Journal of Molecular Sciences 24, no. 15: 11875. https://doi.org/10.3390/ijms241511875
APA StylePerrone, A. E., Pinillo, M., Rial, M. S., Fernández, M., Milduberger, N., González, C., Bustos, P. L., Fichera, L. E., Laucella, S. A., Albareda, M. C., & Bua, J. (2023). Trypanosoma cruzi Secreted Cyclophilin TcCyP19 as an Early Marker for Trypanocidal Treatment Efficiency. International Journal of Molecular Sciences, 24(15), 11875. https://doi.org/10.3390/ijms241511875