
CCR9/CXCR5 Co-Expressing CD4 T Cells Are Increased in Primary Sjögren’s Syndrome and Are Enriched in PD-1/ICOS-Expressing Effector T Cells
 ,
,
Abstract
:1. Introduction
2. Results
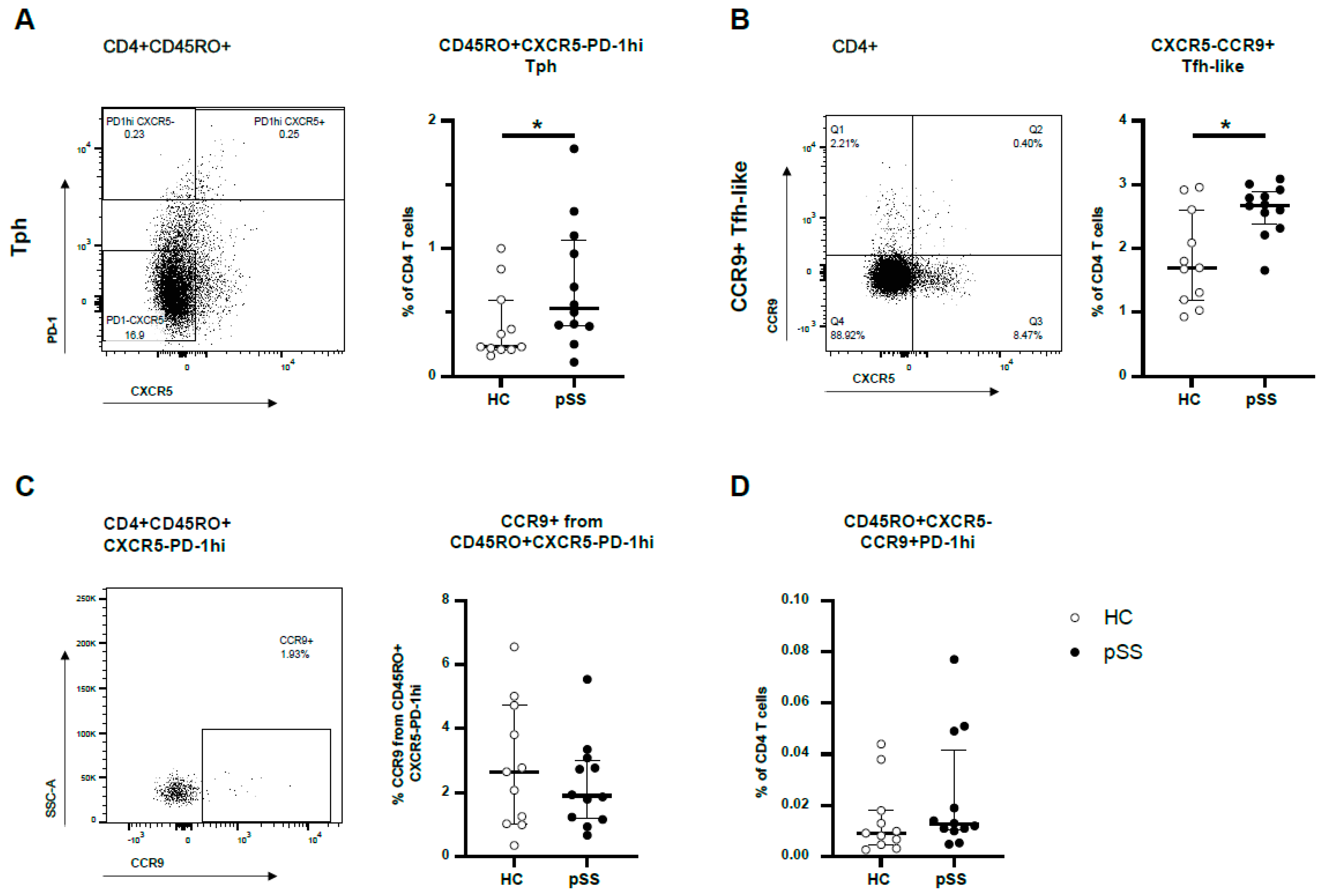
2.1. CXCR5-PD-1hi Tph Cells and CXCR5-CCR9+ Tfh-like Cells Are More Abundant in pSS Patients but These Cell Types Show Little Overlap in Blood
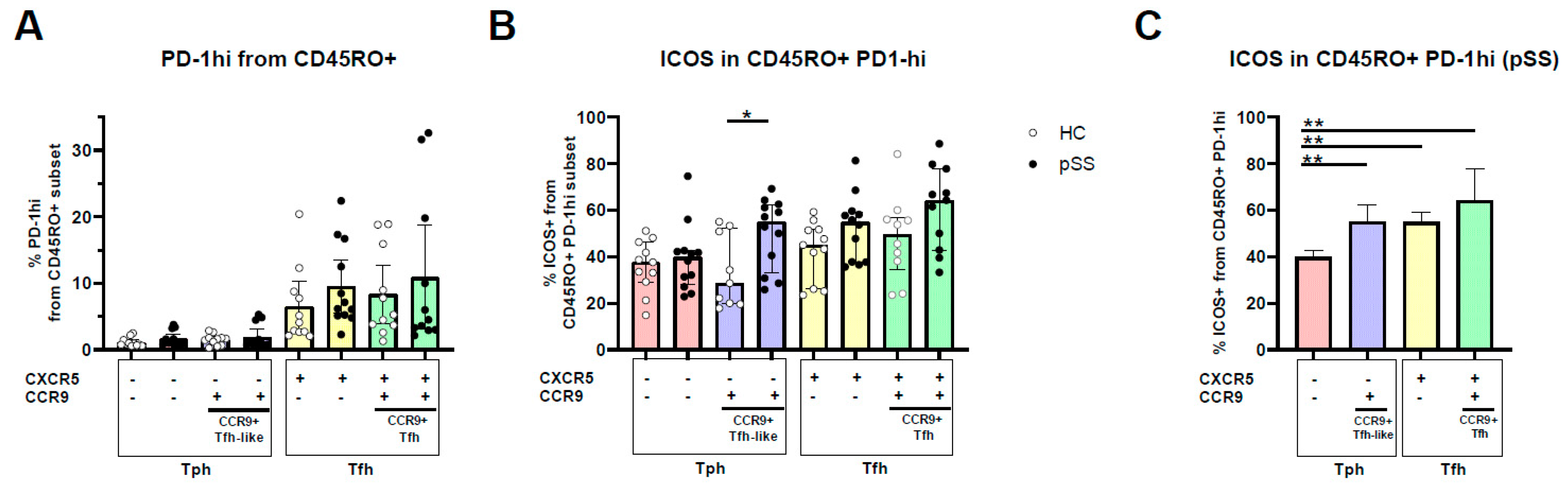
2.2. PD-1hi Expression Is Highest in CXCR5+ Memory Cells, CCR9 and CXCR5/CCR9 Co-Expressing Memory PD-1hi Cells Show Increased ICOS Expression
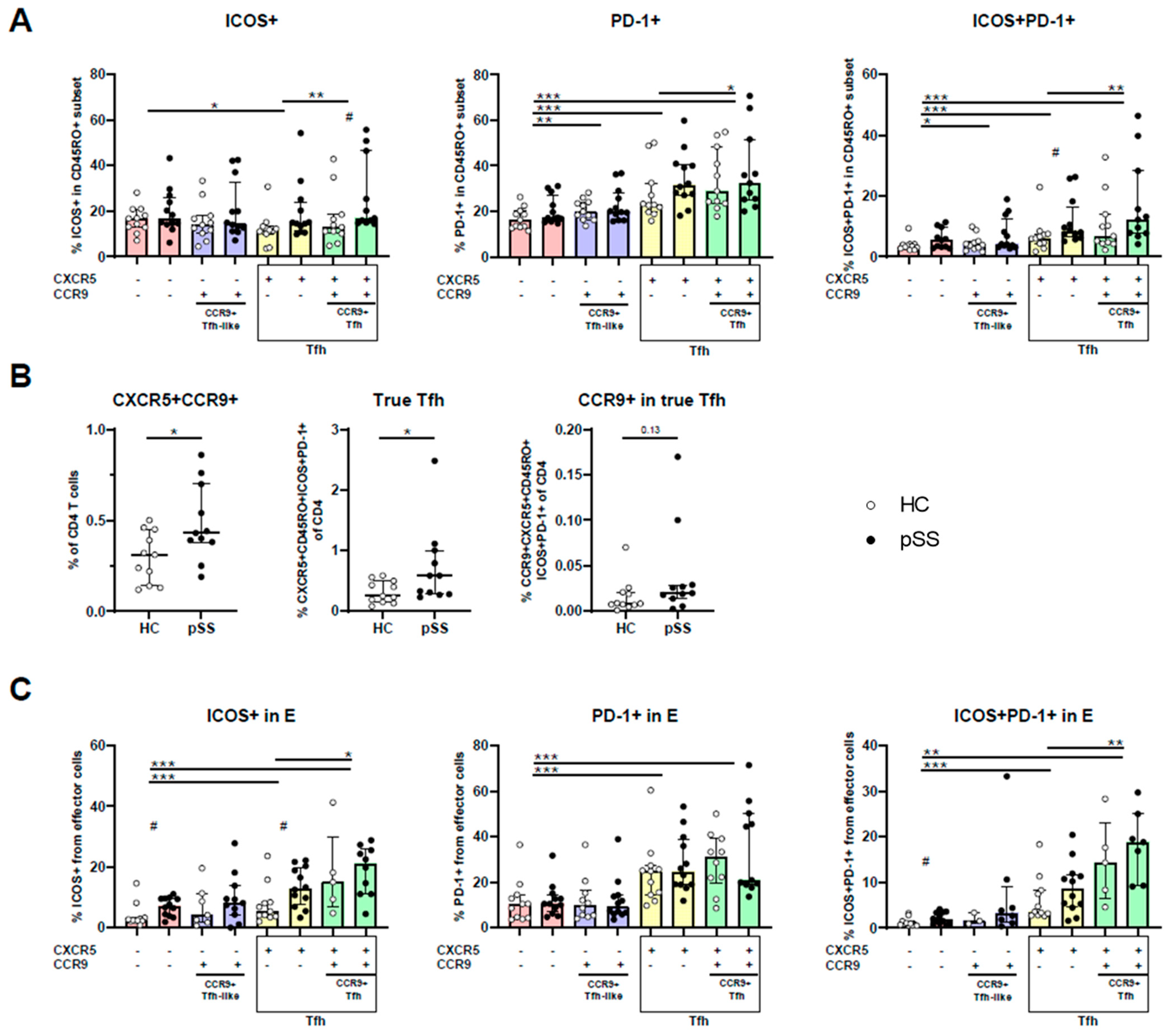
2.3. CXCR5/CCR9 Co-Expressing Memory and Effector Cells Are Enriched for ICOS+PD-1+ Cells and Are Enriched in pSS
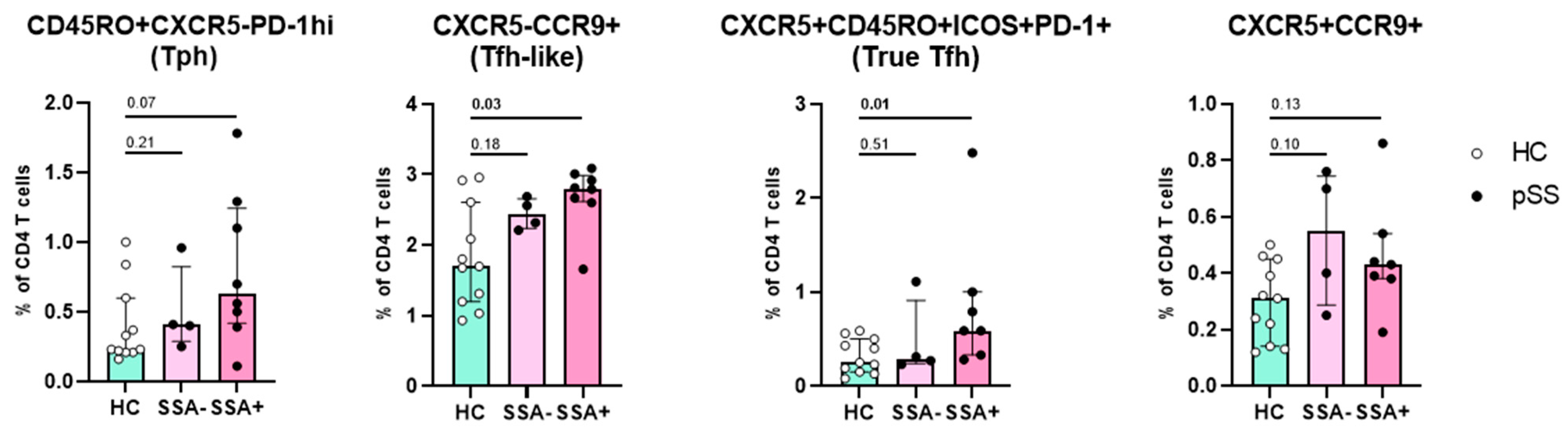
2.4. Increased Numbers of CCR9+ Tfh-like Cells and True Tfh Cells Are Associated with Autoimmunity
3. Discussion
4. Materials and Methods
4.1. Participant Inclusion
4.2. Flow Cytometry
4.3. Gating of PD-1hi Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fox, R.I. Sjogren’s syndrome. Lancet 2005, 366, 321–331. [Google Scholar] [CrossRef]
- Brito-Zerón, P.; Baldini, C.; Bootsma, H.; Bowman, S.J.; Jonsson, R.; Mariette, X.; Sivils, K.; Theander, E.; Tzioufas, A.; Ramos-Casals, M. Sjogren syndrome. Nat. Rev. Dis. Prim. 2016, 2, 16047. [Google Scholar] [CrossRef] [PubMed]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Gottenberg, J.E.; Seror, R.; Miceli-Richard, C.; Benessiano, J.; Devauchelle-Pensec, V.; Dieude, P.; Dubost, J.J.; Fauchais, A.L.; Goeb, V.; Hachulla, E.; et al. Serum Levels of Beta2-Microglobulin and Free Light Chains of Immunoglobulins Are Associated with Systemic Disease Activity in Primary Sjögren’s Syndrome. Data at Enrollment in the Prospective ASSESS Cohort. PLoS ONE 2013, 8, e59868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, K.; Papp, G.; Barath, S.; Szanto, A.; Zeher, M. Follicular helper T cells may play an important role in the severity of primary Sjögren’s syndrome. Clin. Immunol. 2013, 147, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.-Z.; Nititham, J.; Taylor, K.; Miceli-Richard, C.; Sordet, C.; Wachsmann, D.; Bahram, S.; Georgel, P.; Criswell, L.A.; Sibilia, J.; et al. Differentiation of follicular helper T cells by salivary gland epithelial cells in primary Sjögren’s syndrome. J. Autoimmun. 2014, 51, 57–66. [Google Scholar] [CrossRef]
- Jin, L.; Yu, D.; Li, X.; Yu, N.; Li, X.; Wang, Y.; Wang, Y. CD4+CXCR5+ follicular helper T cells in salivary gland promote B cells maturation in patients with primary Sjogren’s syndrome. Int. J. Clin. Exp. Pathol. 2014, 7, 1988–1996. [Google Scholar] [PubMed]
- Li, X.Y.; Wu, Z.B.; Ding, J.; Zheng, Z.H.; Li, X.Y.; Chen, L.N.; Zhu, P. Role of the frequency of blood CD4(+) CXCR5(+) CCR6(+) T cells in autoimmunity in patients with Sjogren’s syndrome. Biochem. Biophys. Res. Commun. 2012, 422, 238–244. [Google Scholar] [CrossRef]
- Rao, D.A.; Gurish, M.F.; Goodman, V.P.B.S.M.; Pernis, V.P.B.S.M.G.A.B.; Marshall, J.L.; Ivashkiv, L.T.D.A.B.P.L.B.; Karlson, E.W.; Nigrovic, L.A.H.P.A.; Filer, A.; Buckley, J.L.M.A.F.C.D.; et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature 2017, 542, 110–114. [Google Scholar] [CrossRef] [Green Version]
- Blokland, S.L.; Hillen, M.R.; Kruize, A.A.; Meller, S.; Homey, B.; Smithson, G.M.; Radstake, T.R.; van Roon, J.A. Increased CCL25 and T Helper Cells Expressing CCR9 in the Salivary Glands of Patients With Primary Sjögren’s Syndrome: Potential New Axis in Lymphoid Neogenesis. Arthritis Rheumatol. 2017, 69, 2038–2051. [Google Scholar] [CrossRef] [Green Version]
- Bocharnikov, A.V.; Keegan, J.; Wacleche, V.S.; Cao, Y.; Fonseka, C.Y.; Wang, G.; Muise, E.S.; Zhang, K.X.; Arazi, A.; Keras, G.; et al. PD-1hiCXCR5–T peripheral helper cells promote B cell responses in lupus via MAF and IL-21. JCI Insight 2019, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Crotty, S. Bcl6-Mediated Transcriptional Regulation of Follicular Helper T cells (TFH). Trends Immunol. 2021, 42, 336–349. [Google Scholar] [CrossRef]
- Chen, W.; Yang, F.; Xu, G.; Ma, J.; Lin, J. Follicular helper T cells and follicular regulatory T cells in the immunopathology of primary Sjögren’s syndrome. J. Leukoc. Biol. 2021, 109, 437–447. [Google Scholar] [CrossRef]
- Brokstad, K.A.; Fredriksen, M.; Zhou, F.; Bergum, B.; Brun, J.G.; Cox, R.J.; Skarstein, K. T follicular-like helper cells in the peripheral blood of patients with primary Sjögren’s syndrome. Scand. J. Immunol. 2018, 88, e12679. [Google Scholar] [CrossRef] [Green Version]
- Pontarini, E.; Murray-Brown, W.J.; Croia, C.; Lucchesi, D.; Conway, J.; Rivellese, F.; Fossati-Jimack, L.; Astorri, E.; Prediletto, E.; Corsiero, E.; et al. Unique expansion of IL-21+ Tfh and Tph cells under control of ICOS identifies Sjögren’s syndrome with ectopic germinal centres and MALT lymphoma. Ann. Rheum. Dis. 2020, 79, 1588–1599. [Google Scholar] [CrossRef]
- Verstappen, G.M.; Nakshbandi, U.; Mossel, E.; Haacke, E.A.; van der Vegt, B.; Vissink, A.; Bootsma, H.; Kroese, F.G. Is the T Follicular Regulatory: Follicular Helper T Cell Ratio in Blood a Biomarker for Ectopic Lymphoid Structure Formation in Sjögren’s Syndrome? Comment on the Article by Fonseca et al. Arthritis Rheumatol. 2018, 70, 1354–1355. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, H.; Liang, Z.; Feng, M.; Wu, Y.; Qin, Y.; Zhao, X.; Gao, C.; Liu, G.; Luo, J. Sirolimus therapy restores the PD-1+ICOS+Tfh:CD45RA-Foxp3high activated Tfr cell balance in primary Sjögren’s syndrome. Mol. Immunol. 2022, 147, 90–100. [Google Scholar] [CrossRef]
- Szabó, K.; Papp, G.; Szántó, A.; Tarr, T.; Zeher, M. A comprehensive investigation on the distribution of circulating follicular T helper cells and B cell subsets in primary Sjögren’s syndrome and systemic lupus erythematosus. Clin. Exp. Immunol. 2015, 183, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Szabo, K.; Papp, G.; Dezso, B.; Zeher, M. The histopathology of labial salivary glands in primary sjögren’s syndrome: Focusing on follicular helper t cells in the inflammatory infiltrates. Mediat. Inflamm. 2014, 2014, 631787. [Google Scholar] [CrossRef] [Green Version]
- Maehara, T.; Moriyama, M.; Hayashida, J.N.; Tanaka, A.; Shinozaki, S.; Kubo, Y.; Matsumura, K.; Nakamura, S. Selective localization of T helper subsets in labial salivary glands from primary Sjögren’s syndrome patients. Clin. Exp. Immunol. 2012, 169, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, K.Y.; Kim, H.O.; Kwok, S.K.; Ju, J.H.; Park, K.S.; Sun, D.I.; Jhun, J.Y.; Oh, H.J.; Park, S.H.; Kim, H.Y. Impact of interleukin-21 in the pathogenesis of primary Sjögren’s syndrome: Increased serum levels of interleukin-21 and its expression in the labial salivary glands. Arthritis Res. Ther. 2011, 13, R179. [Google Scholar] [CrossRef] [Green Version]
- Amft, N.; Curnow, S.J.; Scheel-Toellner, D.; Devadas, A.; Oates, J.; Crocker, J.; Hamburger, J.; Ainsworth, J.; Mathews, J.; Salmon, M.; et al. Ectopic expression of the B cell–attracting chemokine BCA-1 (CXCL13) on endothelial cells and within lymphoid follicles contributes to the establishment of germinal center–like structures in Sjögren’s syndrome. Arthritis Rheum. 2001, 44, 2633–2641. [Google Scholar] [CrossRef]
- Blokland, S.L.M.; Van Vliet-Moret, F.M.; Hillen, M.R.; Pandit, A.; Goldschmeding, R.; Kruize, A.; Bouma, G.; Van Maurik, A.; Olek, S.; Hoffmueller, U.; et al. Epigenetically quantified immune cells in salivary glands of Sjögren’s syndrome patients: A novel tool that detects robust correlations of T follicular helper cells with immunopathology. Rheumatology 2020, 59, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Blokland, S.L.M.; Hillen, M.R.; van Vliet-Moret, F.M.; Bikker, A.; de Jager, W.; Pandit, A.; Kruize, A.; Radstake, T.R.D.J.; Roon, J.A.G.V. Salivary gland secretome: A novel tool towards molecular stratification of patients with primary Sjögren’s syndrome and non-autoimmune sicca. RMD Open 2019, 5, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, F.; Bombardieri, M.; Manzo, A.; Blades, M.C.; Morgan, P.R.; Challacombe, S.; Valesini, G.; Pitzalis, C. Association of CXCL13 and CCL21 expression with the progressive organization of lymphoid-like structures in Sjögren’s syndrome. Arthritis Rheum. 2005, 6, 1773–1784. [Google Scholar] [CrossRef]
- Bombardieri, M.; Barone, F.; Humby, F.; Kelly, S.; McGurk, M.; Morgan, P.; Challacombe, S.; De Vita, S.; Valesini, G.; Spencer, J.; et al. Activation-Induced Cytidine Deaminase Expression in Follicular Dendritic Cell Networks and Interfollicular Large B Cells Supports Functionality of Ectopic Lymphoid Neogenesis in Autoimmune Sialoadenitis and MALT Lymphoma in Sjögren’s Syndrome. J. Immunol. 2007, 7, 4929–4938. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.; Hillen, M.R.; Barone, F. Salivary Gland Pathology in Sjögren’s Syndrome. Rheum. Dis. Clin. N. Am. 2016, 42, 473–483. [Google Scholar] [CrossRef]
- Risselada, A.P.; Looije, M.F.; Kruize, A.A.; Bijlsma, J.W.J.; van Roon, J.A.G. The Role of Ectopic Germinal Centers in the Immunopathology of Primary Sjögren’s Syndrome: A Systematic Review. Semin. Arthritis Rheum. 2013, 42, 368–376. [Google Scholar] [CrossRef]
- Verstappen, G.M.; Meiners, P.M.; Corneth, O.B.J.; Visser, A.; Arends, S.; Abdulahad, W.H.; Hendriks, R.W.; Vissink, A.; Kroese, F.G.M.; Bootsma, H. Attenuation of Follicular Helper T Cell–Dependent B Cell Hyperactivity by Abatacept Treatment in Primary Sjögren’s Syndrome. Arthritis Rheumatol. 2017, 69, 1850–1861. [Google Scholar] [CrossRef] [Green Version]
- Dupré, A.; Pascaud, J.; Rivière, E.; Paoletti, A.; Ly, B.; Mingueneau, M.; Mariette, X.; Nocturne, G. Association between T follicular helper cells and T peripheral helper cells with B-cell biomarkers and disease activity in primary Sjögren syndrome. RMD Open 2021, 7, e001442. [Google Scholar] [CrossRef]
- Rao, D.A. T cells that help B cells in chronically inflamed tissues. Front. Immunol. 2018, 9, 1924. [Google Scholar] [CrossRef] [PubMed]
- McGuire, H.M.; Vogelzang, A.; Ma, C.S.; Hughes, W.E.; Silveira, P.A.; Tangye, S.G.; Christ, D.; Fulcher, D.; Falcone, M.; King, C. A Subset of Interleukin-21 + Chemokine Receptor CCR9 + T Helper Cells Target Accessory Organs of the Digestive System in Autoimmunity. Immunity 2011, 34, 602–615. [Google Scholar] [CrossRef] [Green Version]
- Hinrichs, A.C.; Blokland, S.L.M.; Lopes, A.P.; Wichers, C.G.K.; Kruize, A.A.; Pandit, A.; Radstake, T.R.D.J.; van Roon, J.A.G. Transcriptome Analysis of CCR9+ T Helper Cells From Primary Sjögren’s Syndrome Patients Identifies CCL5 as a Novel Effector Molecule. Front. Immunol. 2021, 1, 2988. [Google Scholar] [CrossRef]
- Wacleche, V.S.; Wang, R.; Rao, D.A. Identification of T Peripheral Helper (Tph) Cells. Methods Mol. Biol. 2022, 2380, 59–76. [Google Scholar]
- Kunkel, E.J.; Campbell, D.J.; Butcher, E.C. Chemokines in lymphocyte trafficking and intestinal immunity. Microcirculation 2003, 10, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Cuello, C.; Palladinetti, P.; Tedla, N.; Di Girolamo, N.; Lloyd, A.R.; McCluskey, P.J.; Wakefield, D. Chemokine expression and leucocyte infiltration in Sjögren’s syndrome. Br. J. Rheumatol. 1998, 37, 779–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kwok, S.K.; Jung, S.M.; Lee, J.; Lee, J.S.; Baek, S.Y.; Kim, E.K.; Ju, J.H.; Park, S.H.; Kim, H.Y. Role of Fractalkine in the Pathogenesis of Primary Sjögren Syndrome: Increased Serum Levels of Fractalkine, Its Expression in Labial Salivary Glands, and the Association with Clinical Manifestations. J. Rheumatol. 2014, 41, 2425–2438. [Google Scholar] [CrossRef]
- Astorri, E.; Scrivo, R.; Bombardieri, M.; Picarelli, G.; Pecorella, I.; Porzia, A.; Valesini, G.; Priori, R. CX3CL1 and CX3CR1 expression in tertiary lymphoid structures in salivary gland infiltrates: Fractalkine contribution to lymphoid neogenesis in Sjögren’s syndrome. Rheumatology 2014, 53, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S. Follicular Helper CD4 T Cells. Annu. Rev. Immunol. 2011, 29, 621–663. [Google Scholar] [CrossRef]
- Choi, Y.S.; Kageyama, R.; Eto, D.; Escobar, T.C.; Johnston, R.J.; Monticelli, L.; Lao, C.; Crotty, S. ICOS Receptor Instructs T Follicular Helper Cell versus Effector Cell Differentiation via Induction of the Transcriptional Repressor Bcl6. Immunity 2011, 34, 932–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, J.S.; Bertino, S.A.; Hernandez, S.G.; Poholek, A.C.; Teplitzky, T.B.; Nowyhed, H.N.; Craft, J. B Cells in T Follicular Helper Cell Development and Function: Separable Roles in Delivery of ICOS Ligand and Antigen. J. Immunol. 2014, 192, 3166–3179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, C.; Bowman, S.J.; et al. 2016 American College of Rheumatology/European League Against Rheumatism Classification Criteria for Primary Sjögren’s Syndrome: A Consensus and Data-Driven Methodology Involving Three International Patient Cohorts. Arthritis Rheumatol. 2017, 69, 35–45. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 11) | pSS (n = 12) | |
|---|---|---|
| Female, n (%) | 11 (100) | 12 (100) |
| Age, years | 55 (49–60) | 59 (54–64) |
| Anti-Ro/SSA positive, n (%) | 8 (67) | |
| Anti-La/SSB positive, n (%) | 4 (33) | |
| ANA positive, n (%) | 7 (58) | |
| Lymphocytic focus score (foci/4 mm2) | 1.8 (1.1–2.4) | |
| IgA-positive plasma cells (%) | 65 (40–81) | |
| Schirmer (mm/5 min) | 0 (0–1) | |
| Serum IgG (g/L) | 13.6 (11.4–15.2) | |
| ESSDAI score (0–123) | 4 (2–5) | |
| ESSPRI score (0–10) | 6.3 (5.3–7.3) | |
| Immunosuppressants, n | 3 | |
| Hydroxychloroquine, n | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinrichs, A.C.; Kruize, A.A.; Lafeber, F.P.J.G.; Leavis, H.L.; van Roon, J.A.G. CCR9/CXCR5 Co-Expressing CD4 T Cells Are Increased in Primary Sjögren’s Syndrome and Are Enriched in PD-1/ICOS-Expressing Effector T Cells. Int. J. Mol. Sci. 2023, 24, 11952. https://doi.org/10.3390/ijms241511952
Hinrichs AC, Kruize AA, Lafeber FPJG, Leavis HL, van Roon JAG. CCR9/CXCR5 Co-Expressing CD4 T Cells Are Increased in Primary Sjögren’s Syndrome and Are Enriched in PD-1/ICOS-Expressing Effector T Cells. International Journal of Molecular Sciences. 2023; 24(15):11952. https://doi.org/10.3390/ijms241511952
Chicago/Turabian StyleHinrichs, Anneline C., Aike A. Kruize, Floris P. J. G. Lafeber, Helen L. Leavis, and Joel A. G. van Roon. 2023. "CCR9/CXCR5 Co-Expressing CD4 T Cells Are Increased in Primary Sjögren’s Syndrome and Are Enriched in PD-1/ICOS-Expressing Effector T Cells" International Journal of Molecular Sciences 24, no. 15: 11952. https://doi.org/10.3390/ijms241511952
APA StyleHinrichs, A. C., Kruize, A. A., Lafeber, F. P. J. G., Leavis, H. L., & van Roon, J. A. G. (2023). CCR9/CXCR5 Co-Expressing CD4 T Cells Are Increased in Primary Sjögren’s Syndrome and Are Enriched in PD-1/ICOS-Expressing Effector T Cells. International Journal of Molecular Sciences, 24(15), 11952. https://doi.org/10.3390/ijms241511952