
Mechanism of Social Stress-Related Erectile Dysfunction in Mice: Impaired Parasympathetic Neurotransmission and Ketamine

 , , and
, , and 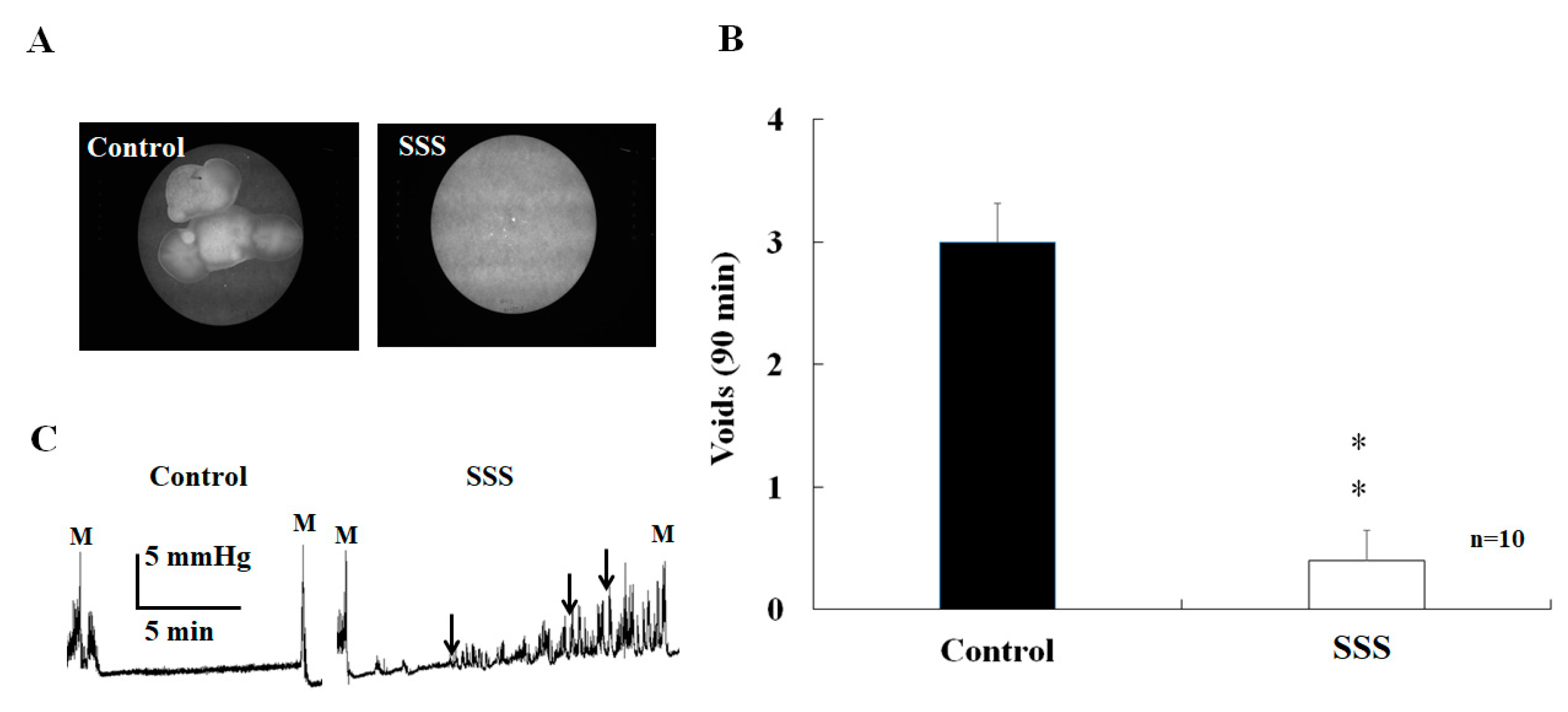{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of SS on Voiding
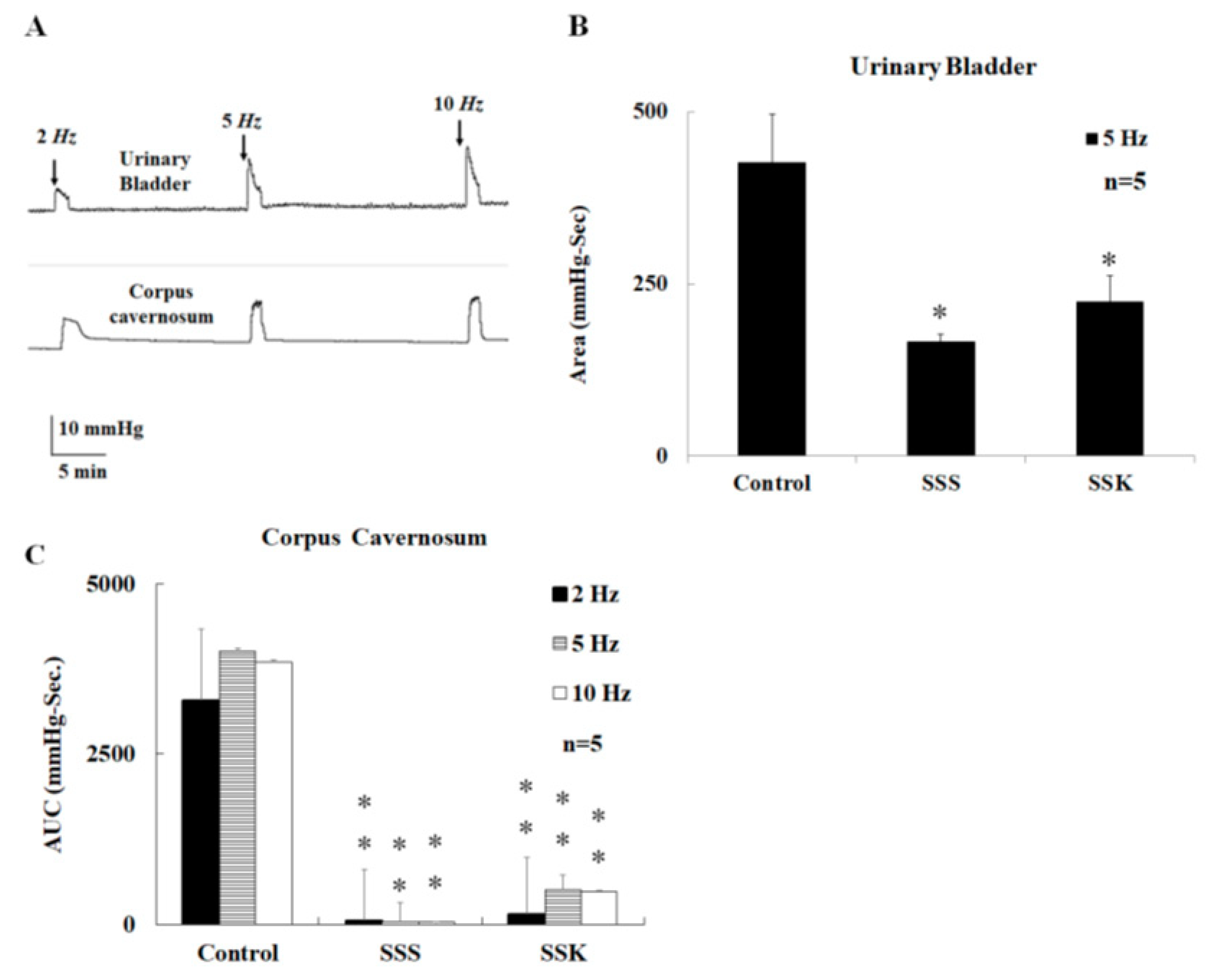
2.2. Effects of SS on Urinary Bladder Pressure and ICP
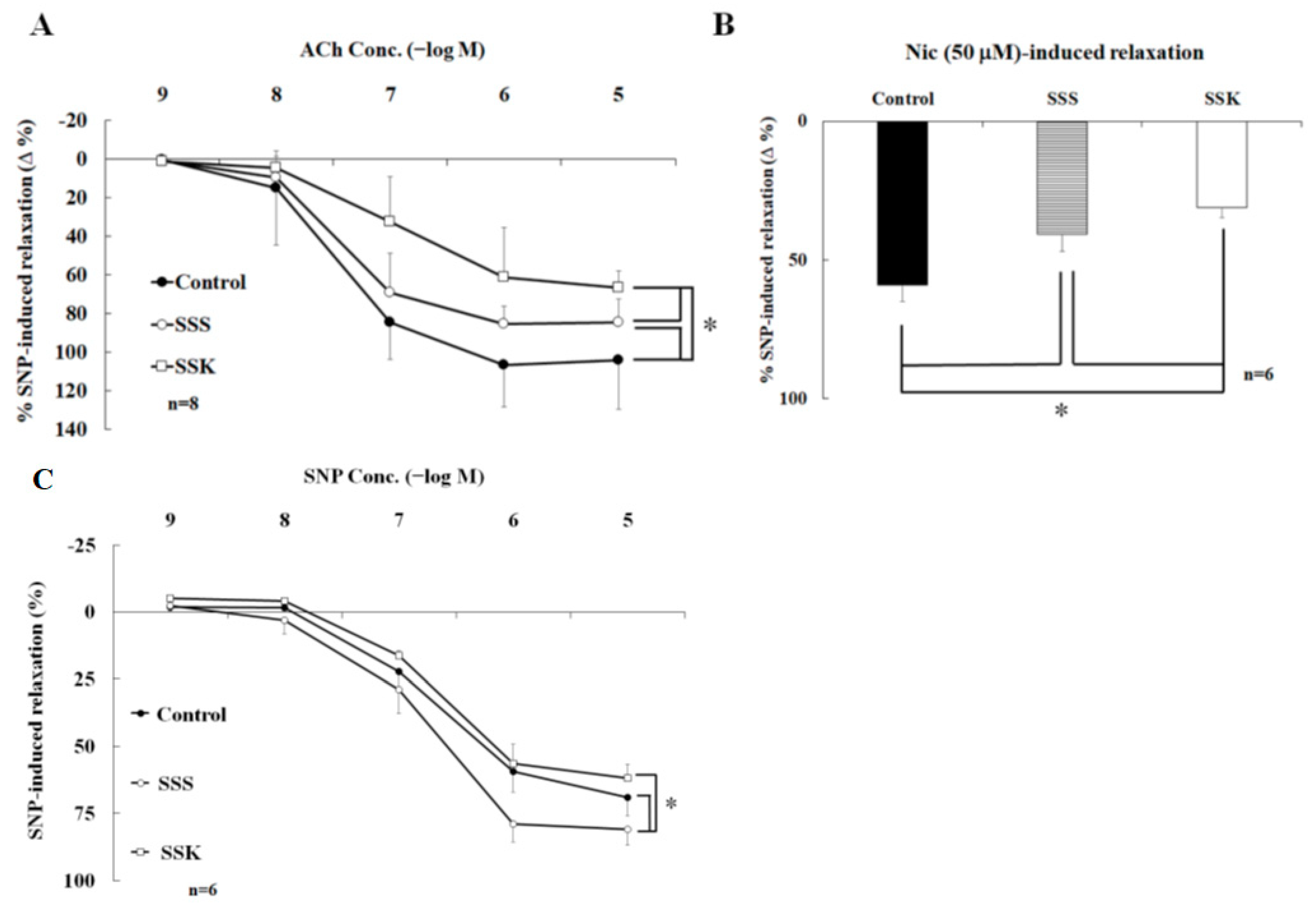
2.3. Effects of SS on the Relaxation of Corpus Cavernosum Strips
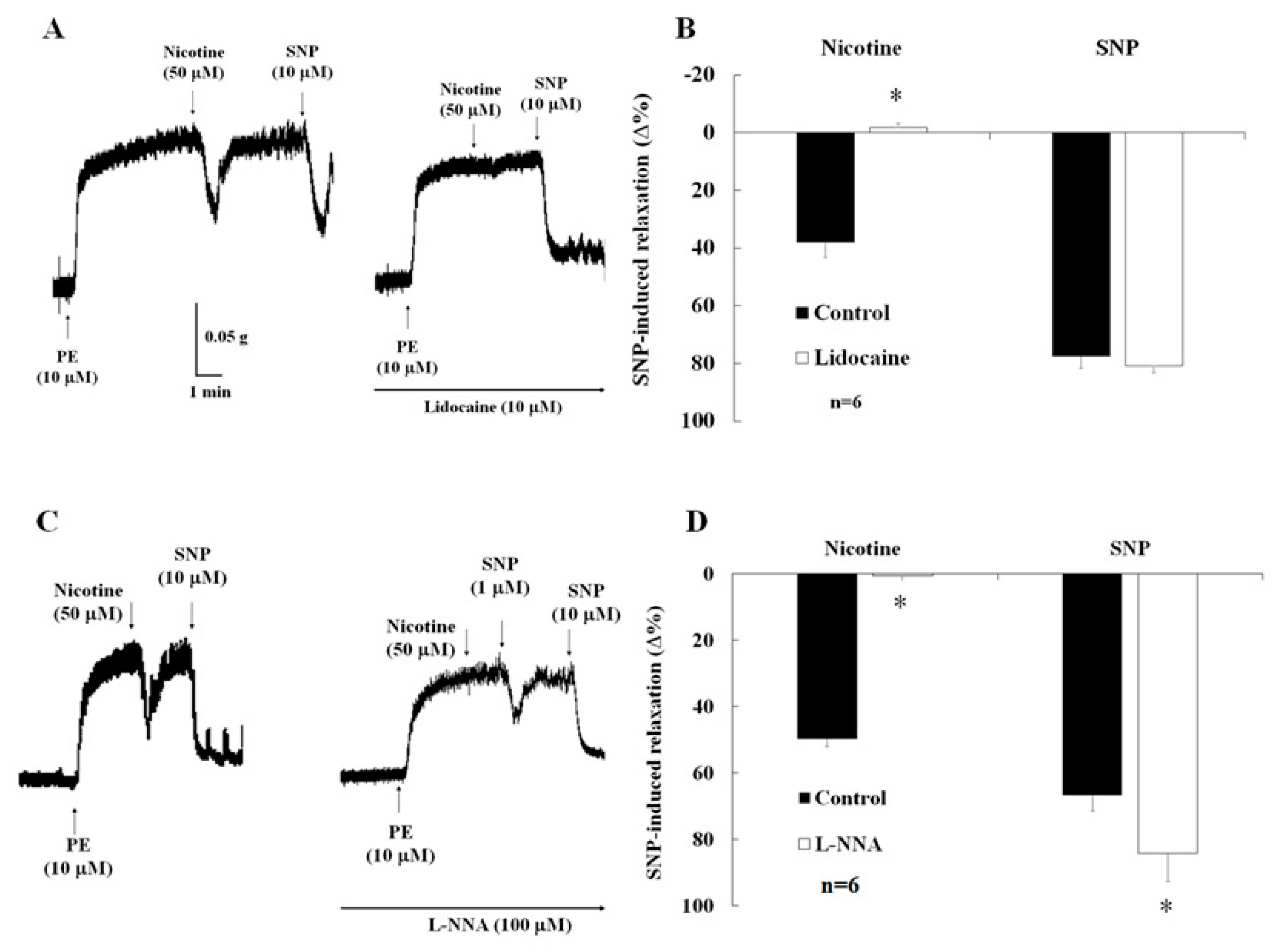
2.4. Effects of Lidocaine on Nicotine-Induced Relaxation
2.5. Effects of NO Synthase Inhibitor on Nicotine-Induced Relaxation and ICP
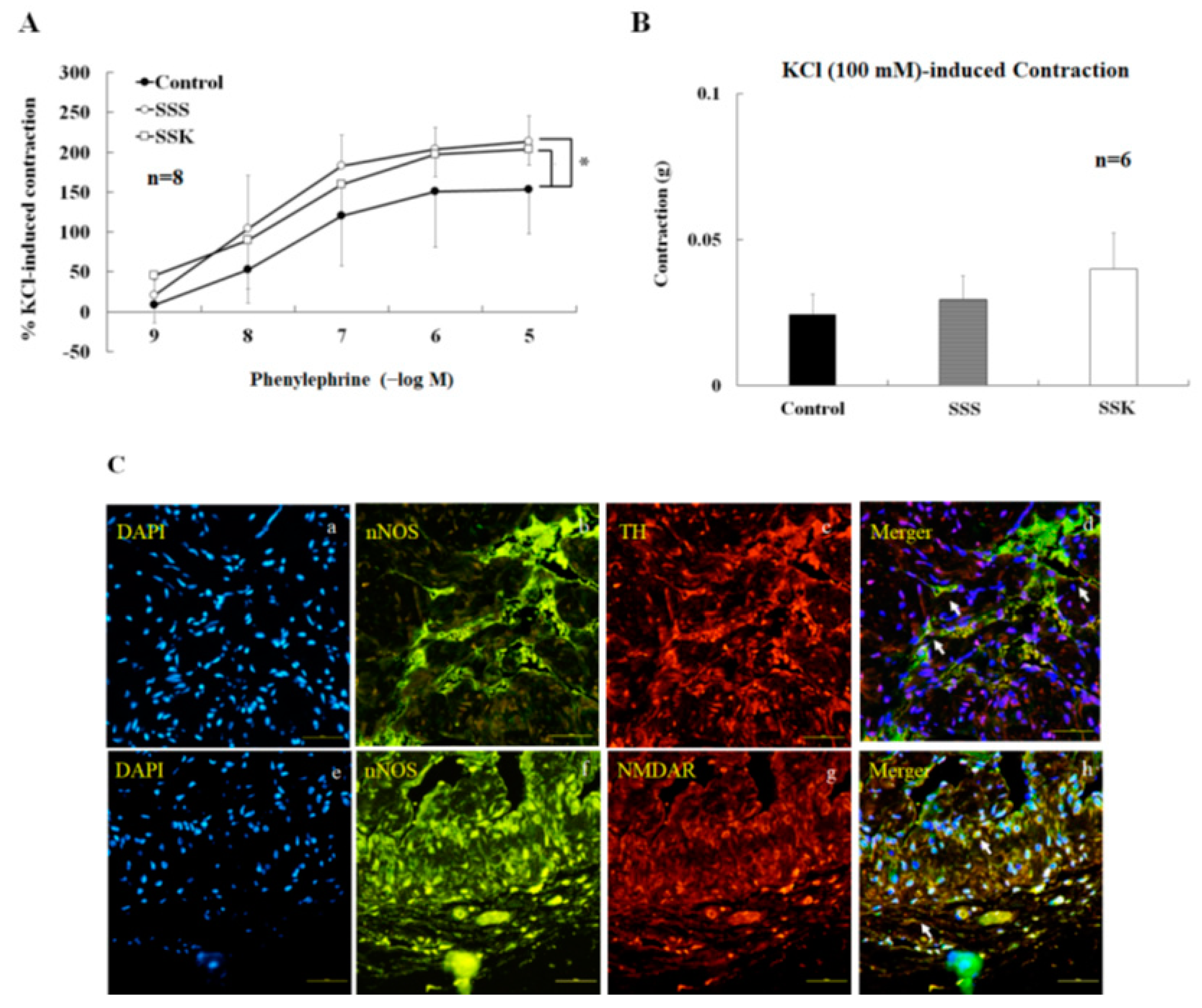
2.6. Effects of SS on the Contraction of Corpus Cavernosum Strips
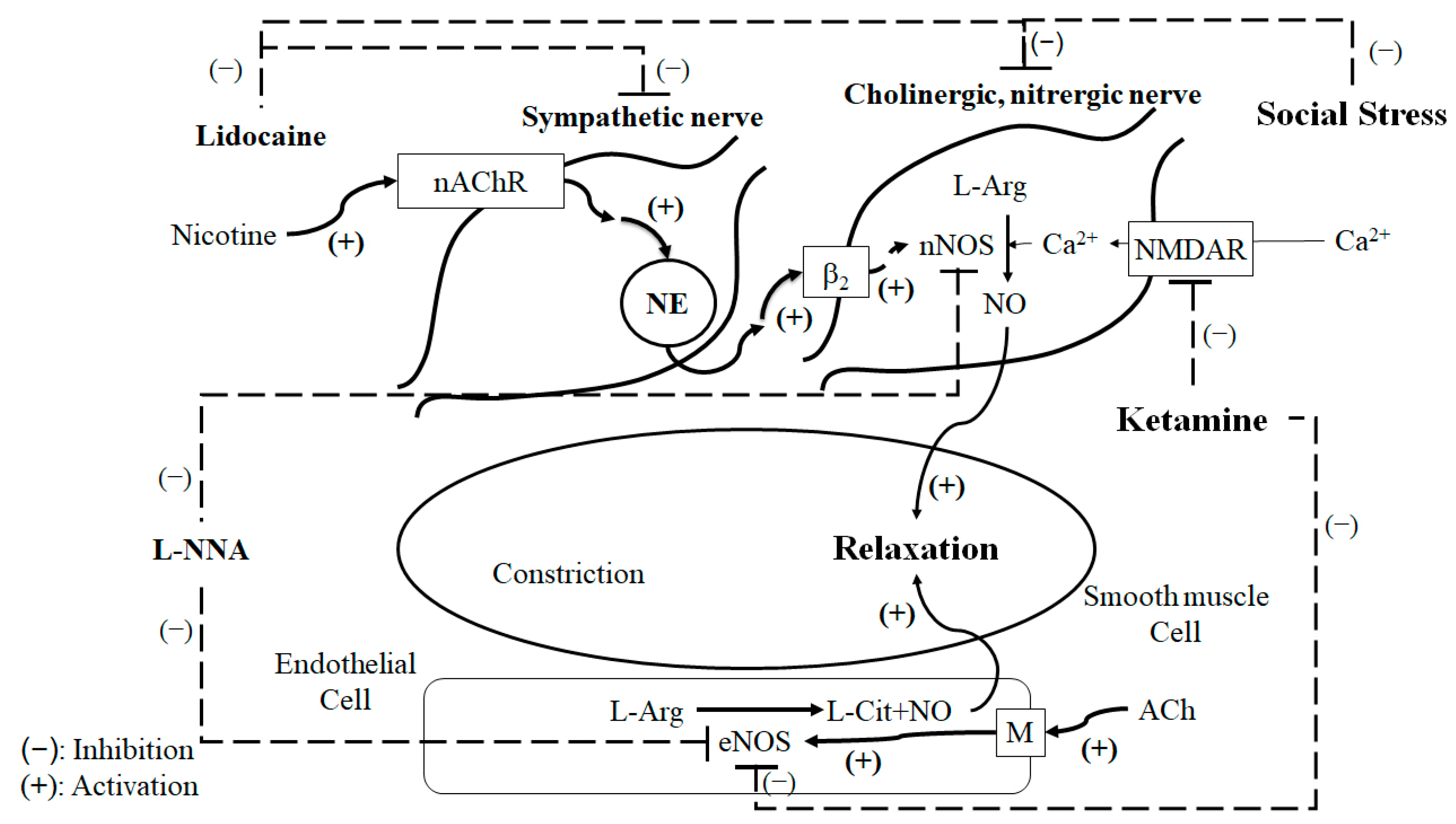
3. Discussion
4. Materials and Methods
4.1. SS Model
4.2. Ketamine Administration Procedures
4.3. Urinary Bladder Pressure
4.4. ICP Measure
4.5. Tissue Preparation
4.6. Tissue Bath Wire Myography
4.7. Double-Labeled Immunohistochemistry
4.8. Drugs Used and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mingin, G.C.; Peterson, A.; Erickson, C.S.; Nelson, M.T.; Vizzard, M.A. Social stress induces changes in urinary bladder function, bladder NGF content, and generalized bladder inflammation in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R893–R900. [Google Scholar] [PubMed] [Green Version]
- Wood, S.K.; Baez, M.A.; Bhatnagar, S.; Valentino, R.J. Social stress-induced bladder dysfunction: Potential role of corticotropin-releasing factor. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1671–R1678. [Google Scholar] [PubMed] [Green Version]
- Mingin, G.C.; Heppner, T.J.; Tykocki, N.R.; Erickson, C.S.; Vizzard, M.A.; Nelson, M.T. Social stress in mice induces urinary bladder overactivity and increases TRPV1 channel-dependent afferent nerve activity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R629–R638. [Google Scholar]
- Wood, S.K.; McFadden, K.; Griffin, T.; Wolfe, J.H.; Zderic, S.; Valentino, R.J. A corticotropin-releasing factor receptor antagonist improves urodynamic dysfunction produced by social stress or partial bladder outlet obstruction in male rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R940–R950. [Google Scholar] [PubMed] [Green Version]
- Yoshimura, H.; Kimura, N. Ethopharmacology of copulatory disorder induced by chronic social conflict in male mice. Neurosci. Biobehav. Rev. 1991, 15, 497–500. [Google Scholar]
- Sugiura, K.; Yoshimura, H.; Yokoyama, M. An animal model of copulatory disorder induced by social stress in male mice: Effects of apomorphine and L-dopa. Psychopharmacology 1997, 133, 249–255. [Google Scholar]
- Mizuno, T.; Yotsuyanagi, S.; Nagasaka, Y.; Namiki, M. Dehydroepiandrosterone alleviates copulatory disorder induced by social stress in male rats. J. Sex. Med. 2006, 3, 612–618. [Google Scholar]
- Menard, C.; Pfau, M.L.; Hodes, G.E.; Kana, V.; Wang, V.X.; Bouchard, S.; Takahashi, A.; Flanigan, M.E.; Aleyasin, H.; LeClair, K.B.; et al. Social stress induces neurovascular pathology promoting depression. Nat. Neurosci. 2017, 20, 1752–1760. [Google Scholar]
- Pitsikas, N. The role of nitric oxide (NO) donors in anxiety. Lights Shad. Nitric Oxide 2018, 77, 6–11. [Google Scholar]
- Baranyi, A.; Amouzadeh-Ghadikolai, O.; Rothenhäusler, H.B.; Theokas, S.; Robier, C.; Baranyi, M.; Koppitz, M.; Reicht, G.; Hlade, P.; Meinitzer, A. Nitric Oxide-Related Biological Pathways in Patients with Major Depression. PLoS ONE 2015, 10, e0143397. [Google Scholar]
- Joca, S.R.L.; Sartim, A.G.; Roncalho, A.L.; Diniz, C.F.A.; Wegener, G. Nitric oxide signalling and antidepressant action revisited. Cell Tissue Res. 2019, 377, 45–58. [Google Scholar]
- Chang, H.H.; Lee, Y.C.; Chen, M.F.; Kuo, J.S.; Lee, T.J. Sympathetic activation increases basilar arterial blood flow in normotensive but not hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1123–H1130. [Google Scholar]
- Lee, T.J.; Saito, A. Altered cerebral vessel innervation in the spontaneously hypertensive rat. Circ. Res. 1984, 55, 392–403. [Google Scholar] [PubMed]
- Burnett, A.L.; Lowenstein, C.J.; Bredt, D.S.; Chang, T.S.; Snyder, S.H. Nitric oxide: A physiologic mediator of penile erection. Science 1992, 257, 401–403. [Google Scholar]
- Ozkara, H.; Alan, C.; Atukeren, P.; Uyaner, I.; Demirci, C.; Gümüştaş, M.K.; Alici, B. Changes of nitric oxide synthase-containing nerve fibers and parameters for oxidative stress after unilateral cavernous nerve resection or manuplation in rat penis. Chin. J. Physiol. 2006, 49, 160–166. [Google Scholar]
- Priviero, F.B.; Jin, L.M.; Ying, Z.; Teixeira, C.E.; Webb, R.C. Up-regulation of the RhoA/Rho-kinase signaling pathway in corpus cavernosum from endothelial nitric-oxide synthase (NOS), but not neuronal NOS, null mice. J. Pharmacol. Exp. Ther. 2010, 333, 184–192. [Google Scholar] [PubMed] [Green Version]
- Dashwood, M.R.; Crump, A.; Shi-Wen, X.; Loesch, A. Identification of neuronal nitric oxide synthase (nNOS) in human penis: A potential role of reduced neuronally-derived nitric oxide in erectile dysfunction. Curr. Pharm. Biotechnol. 2011, 12, 1316–1321. [Google Scholar] [PubMed]
- Chen, C.H.; Lee, M.H.; Chen, Y.C.; Lin, M.F. Ketamine-snorting associated cystitis. J. Formos. Med. Assoc. 2011, 110, 787–791. [Google Scholar]
- Kuiken, S.D.; van den Berg, S.J.; Tytgat, G.N.; Boeckxstaens, G.E. Oral S(+)-ketamine does not change visceral perception in health. Dig. Dis. Sci. 2004, 49, 1745–1751. [Google Scholar]
- Franceschelli, A.; Sens, J.; Herchick, S.; Thelen, C.; Pitychoutis, P.M. Sex differences in the rapid and the sustained antidepressant-like effects of ketamine in stress-naïve and “depressed” mice exposed to chronic mild stress. Neuroscience 2015, 290, 49–60. [Google Scholar]
- Brachman, R.A.; McGowan, J.C.; Perusini, J.N.; Lim, S.C.; Pham, T.H.; Faye, C.; Gardier, A.M.; Mendez-David, I.; David, D.J.; Hen, R.; et al. Ketamine as a Prophylactic Against Stress-Induced Depressive-like Behavior. Biol. Psychiatry 2016, 79, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Hashimoto, K. Comparison of ketamine, 7,8-dihydroxyflavone, and ANA-12 antidepressant effects in the social defeat stress model of depression. Psychopharmacology 2015, 232, 4325–4335. [Google Scholar] [CrossRef]
- Yang, S.S.; Jang, M.Y.; Lee, K.H.; Hsu, W.T.; Chen, Y.C.; Chen, W.S.; Chang, S.J. Sexual and bladder dysfunction in male ketamine abusers: A large-scale questionnaire study. PLoS ONE 2018, 13, e0207927. [Google Scholar]
- Li, C.C.; Wu, S.T.; Cha, T.L.; Sun, G.H.; Yu, D.S.; Meng, E. A survey for ketamine abuse and its relation to the lower urinary tract symptoms in Taiwan. Sci. Rep. 2019, 9, 7240. [Google Scholar] [CrossRef] [Green Version]
- Li, M.W.; Chao, T.C.; Lim, L.Y.; Chang, H.H.; Yang, S.S. The Acute Effects and Mechanism of Ketamine on Nicotine-Induced Neurogenic Relaxation of the Corpus Cavernosum in Mice. Int. J. Mol. Sci. 2023, 24, 6976. [Google Scholar] [CrossRef]
- Yang, S.S.; Chang, H.H.; Chang, S.J. Does ketamine ameliorate the social stress-related bladder dysfunction in mice? Neurourol. Urodyn. 2020, 39, 935–944. [Google Scholar] [CrossRef]
- Ekström, J.; Elmér, M. Compensatory increase of responses to nerve stimulation of the partially denervated rat urinary bladder. Acta Physiol. Scand. 1980, 110, 21–29. [Google Scholar]
- Hammarström, M.; Sjöstrand, N.O. Comments on the atropine resistance of the neurogenic contractile response of the rat detrusor muscle. Acta Physiol. Scand. 1984, 122, 475–481. [Google Scholar]
- Fey, T.A.; Gopalakrishnan, M.; Strake, J.G.; King, L.L.; Brioni, J.D.; Sullivan, J.P.; Coghlan, M.J.; Brune, M.E. Effects of ATP-sensitive K+ channel openers and tolterodine on involuntary bladder contractions in a pig model of partial bladder outlet obstruction. Neurourol. Urodyn. 2003, 22, 147–155. [Google Scholar] [PubMed]
- Fabiyi, A.C.; Gopalakrishnan, M.; Lynch, J.J., 3rd; Brioni, J.D.; Coghlan, M.J.; Brune, M.E. In vivo evaluation of the potency and bladder-vascular selectivity of the ATP-sensitive potassium channel openers (-)-cromakalim, ZD6169 and WAY-133537 in rats. BJU Int. 2003, 91, 284–290. [Google Scholar]
- Srilatha, B.; Adaikan, P.G. Estrogen and phytoestrogen predispose to erectile dysfunction: Do ER-alpha and ER-beta in the cavernosum play a role? Urology 2004, 63, 382–386. [Google Scholar] [CrossRef]
- Ohmasa, F.; Saito, M.; Tsounapi, P.; Dimitriadis, F.; Inoue, S.; Shomori, K.; Shimizu, S.; Kinoshita, Y.; Satoh, K. Edaravone ameliorates diabetes-induced dysfunction of NO-induced relaxation in corpus cavernosum smooth muscle in the rat. J. Sex. Med. 2011, 8, 1638–1649. [Google Scholar] [CrossRef]
- Fraga-Silva, R.A.; Costa-Fraga, F.P.; Savergnini, S.Q.; De Sousa, F.B.; Montecucco, F.; da Silva, D.; Sinisterra, R.D.; Mach, F.; Stergiopulos, N.; da Silva, R.F.; et al. An oral formulation of angiotensin-(1-7) reverses corpus cavernosum damages induced by hypercholesterolemia. J. Sex. Med. 2013, 10, 2430–2442. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.M.; Chen, T.L.; Lin, Y.L.; Chen, T.G.; Tai, Y.T. Ketamine reduces nitric oxide biosynthesis in human umbilical vein endothelial cells by down-regulating endothelial nitric oxide synthase expression and intracellular calcium levels. Crit. Care Med. 2005, 33, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Akata, T.; Izumi, K.; Nakashima, M. Mechanisms of direct inhibitory action of ketamine on vascular smooth muscle in mesenteric resistance arteries. Anesthesiology 2001, 95, 452–462. [Google Scholar] [CrossRef] [Green Version]
- Bezanilla, F. Voltage-gated ion channels. IEEE Trans. Nanobioscience 2005, 4, 34–48. [Google Scholar] [CrossRef]
- Haeseler, G.; Tetzlaff, D.; Bufler, J.; Dengler, R.; Münte, S.; Hecker, H.; Leuwer, M. Blockade of voltage-operated neuronal and skeletal muscle sodium channels by S(+)- and R(−)-ketamine. Anesth. Analg. 2003, 96, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Irnaten, M.; Wang, J.; Chang, K.S.; Andresen, M.C.; Mendelowitz, D. Ketamine inhibits sodium currents in identified cardiac parasympathetic neurons in nucleus ambiguus. Anesthesiology 2002, 96, 659–666. [Google Scholar] [CrossRef]
- Bozkurt, N.B.; Vural, I.M.; Sarioglu, Y.; Pekiner, C. Nicotine potentiates the nitrergic relaxation responses of rabbit corpus cavernosum tissue via nicotinic acetylcholine receptors. Eur. J. Pharmacol. 2007, 558, 172–178. [Google Scholar] [CrossRef]
- Faghir-Ghanesefat, H.; Rahimi, N.; Yarmohammadi, F.; Mokhtari, T.; Abdollahi, A.R.; Mehr, S.E.; Dehpour, A.R. The expression, localization and function of α7 nicotinic acetylcholine receptor in rat Corpus cavernosum. J. Pharm. Pharmacol. 2017, 69, 1754–1761. [Google Scholar] [CrossRef]
- Nangle, M.R.; Cotter, M.A.; Cameron, N.E. An in vitro investigation of aorta and corpus cavernosum from eNOS and nNOS gene-deficient mice. Pflug. Arch. 2004, 448, 139–145. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.-Y.; Chao, T.-C.; Hsu, C.-K.; Chang, H.-H.; Yang, S.S.-D. Mechanism of Social Stress-Related Erectile Dysfunction in Mice: Impaired Parasympathetic Neurotransmission and Ketamine. Int. J. Mol. Sci. 2023, 24, 11973. https://doi.org/10.3390/ijms241511973
Wu S-Y, Chao T-C, Hsu C-K, Chang H-H, Yang SS-D. Mechanism of Social Stress-Related Erectile Dysfunction in Mice: Impaired Parasympathetic Neurotransmission and Ketamine. International Journal of Molecular Sciences. 2023; 24(15):11973. https://doi.org/10.3390/ijms241511973
Chicago/Turabian StyleWu, Shu-Yu, Tze-Chen Chao, Chun-Kai Hsu, His-Hsien Chang, and Stephen Shei-Dei Yang. 2023. "Mechanism of Social Stress-Related Erectile Dysfunction in Mice: Impaired Parasympathetic Neurotransmission and Ketamine" International Journal of Molecular Sciences 24, no. 15: 11973. https://doi.org/10.3390/ijms241511973
APA StyleWu, S. -Y., Chao, T. -C., Hsu, C. -K., Chang, H. -H., & Yang, S. S. -D. (2023). Mechanism of Social Stress-Related Erectile Dysfunction in Mice: Impaired Parasympathetic Neurotransmission and Ketamine. International Journal of Molecular Sciences, 24(15), 11973. https://doi.org/10.3390/ijms241511973