
Antarctic Soil Metabolomics: A Pilot Study


 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
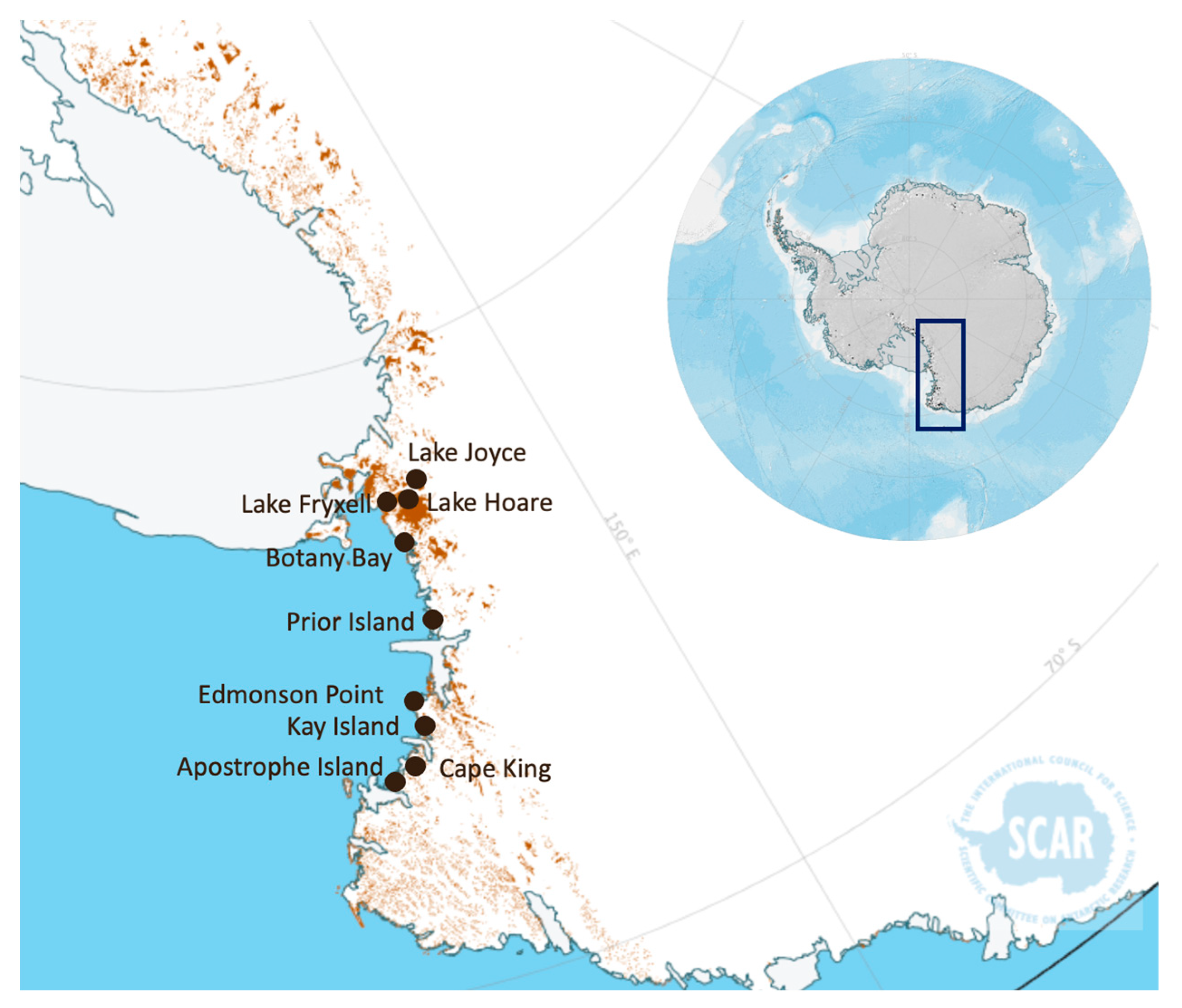
2.1. Antarctic Soil Samples’ Selection
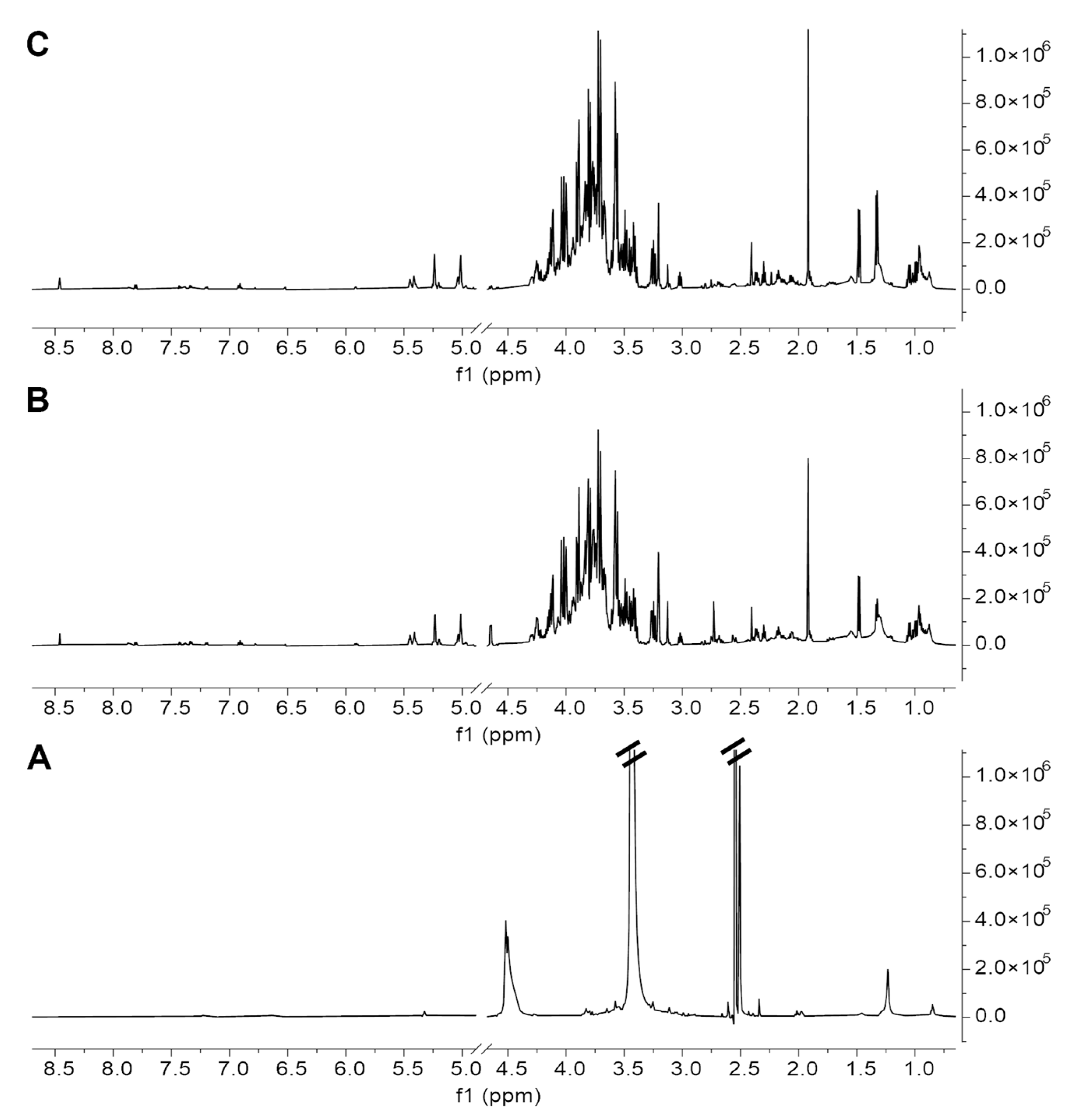
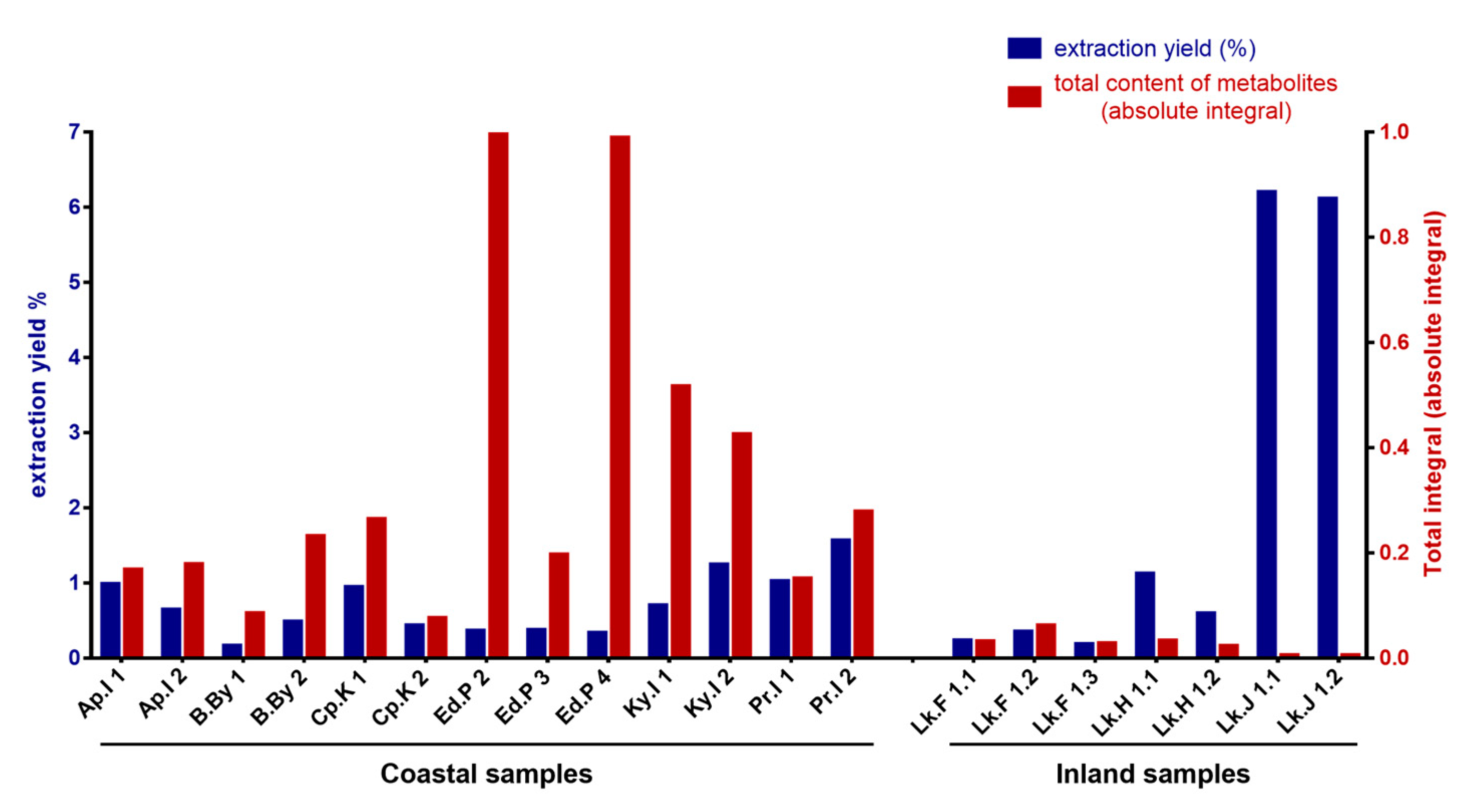
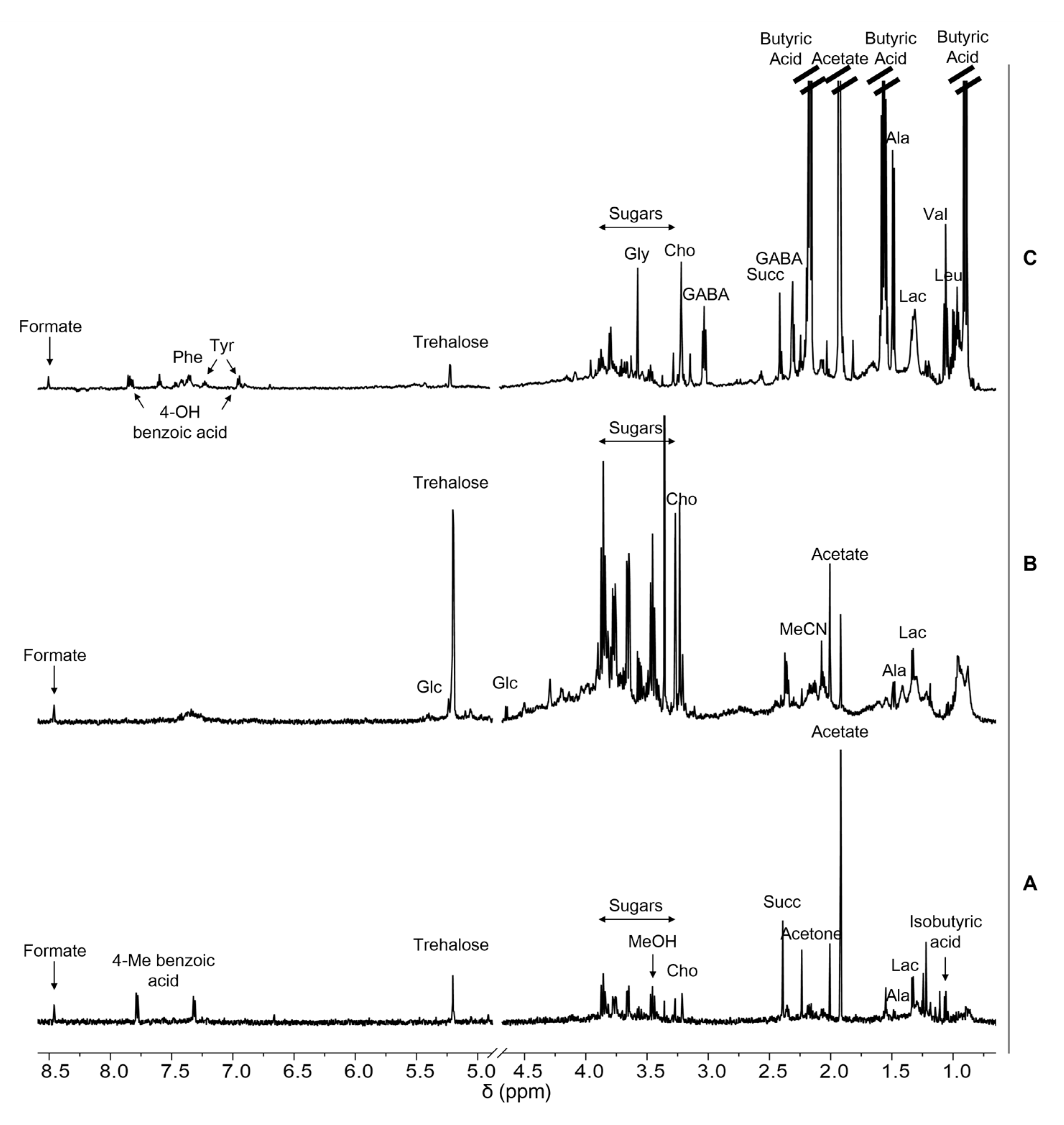
2.2. Antarctic Soil Extract Preparation and NMR-Based Metabolic Profiling
2.3. UPLC/HR-MS Metabolic Profiling of Selected Soil Extracts
3. Discussion
4. Materials and Methods
4.1. Soil Sampling and Storage
4.2. Metabolomics Analyses
4.2.1. Materials
4.2.2. Soil Sample Extractions for NMR and MS Analyses
Extraction in DMSO
Extraction in H2O/CH3OH 1:1
Extraction in H2O/CH3CN 7:3
4.2.3. NMR Metabolic Profiling of Soil Extracts
4.2.4. MS Metabolic Profiling of Selected Soil Extracts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Convey, P.; Chown, S.L.; Clarke, A.; Barnes, D.K.A.; Bokhorst, S.; Cummings, V.; Ducklow, H.W.; Frati, F.; Green, T.G.A.; Gordon, S.; et al. The Spatial Structure of Antarctic Biodiversity. Ecol. Monogr. 2014, 84, 203–244. [Google Scholar] [CrossRef] [Green Version]
- Roudier, P.; O’Neill, T.A.; Almond, P.; Poirot, C. Soil Priorities for Antarctica. Geoderma Reg. 2022, 29, e00499. [Google Scholar] [CrossRef]
- Dennis, P.G.; Newsham, K.K.; Rushton, S.P.; O’Donnell, A.G.; Hopkins, D.W. Soil Bacterial Diversity Is Positively Associated with Air Temperature in the Maritime Antarctic. Sci. Rep. 2019, 9, 2686. [Google Scholar] [CrossRef] [Green Version]
- Newsham, K.K.; Davey, M.L.; Hopkins, D.W.; Dennis, P.G. Regional Diversity of Maritime Antarctic Soil Fungi and Predicted Responses of Guilds and Growth Forms to Climate Change. Front. Microbiol. 2021, 11, 615659. [Google Scholar] [CrossRef]
- Ball, B.A.; Convey, P.; Feeser, K.L.; Nielsen, U.N.; Horn, D.V. Habitat Severity Characteristics Structure Soil Communities at Regional and Local Spatial Scales along the Antarctica Peninsula. Antarct. Sci. 2023, 35, 103–119. [Google Scholar] [CrossRef]
- Kim, M.; Cho, A.; Lim, H.S.; Hong, S.G.; Kim, J.H.; Lee, J.; Choi, T.; Ahn, T.S.; Kim, O.-S. Highly Heterogeneous Soil Bacterial Communities around Terra Nova Bay of Northern Victoria Land, Antarctica. PLoS ONE 2015, 10, e0119966. [Google Scholar] [CrossRef]
- Canini, F.; Geml, J.; D’Acqui, L.P.; Selbmann, L.; Onofri, S.; Ventura, S.; Zucconi, L. Exchangeable Cations and PH Drive Diversity and Functionality of Fungal Communities in Biological Soil Crusts from Coastal Sites of Victoria Land, Antarctica. Fungal Ecol. 2020, 45, 100923. [Google Scholar] [CrossRef]
- Canini, F.; Geml, J.; Buzzini, P.; Turchetti, B.; Onofri, S.; D’Acqui, L.P.; Ripa, C.; Zucconi, L. Growth Forms and Functional Guilds Distribution of Soil Fungi in Coastal Versus Inland Sites of Victoria Land, Antarctica. Biology 2021, 10, 320. [Google Scholar] [CrossRef] [PubMed]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA Is Abundant in Soil and Obscures Estimates of Soil Microbial Diversity. Nat. Microbiol. 2016, 2, 16242. [Google Scholar] [CrossRef] [PubMed]
- Swenson, T.L.; Karaoz, U.; Swenson, J.M.; Bowen, B.P.; Northen, T.R. Linking Soil Biology and Chemistry in Biological Soil Crust Using Isolate Exometabolomics. Nat. Commun. 2018, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Withers, E.; Hill, P.W.; Chadwick, D.R.; Jones, D.L. Use of Untargeted Metabolomics for Assessing Soil Quality and Microbial Function. Soil Biol. Biochem. 2020, 143, 107758. [Google Scholar] [CrossRef]
- White, D.; Beyer, L. Pyrolysis-Gas Chromatography/Mass Spectrometry and Pyrolysis-Gas Chromatography/Flame Ionization Detection Analysis of Three Antarctic Soils. J. Anal. Appl. Pyrolysis 1999, 50, 63–76. [Google Scholar] [CrossRef]
- Coleine, C.; Gevi, F.; Fanelli, G.; Onofri, S.; Timperio, A.M.; Selbmann, L. Specific Adaptations Are Selected in Opposite Sun Exposed Antarctic Cryptoendolithic Communities as Revealed by Untargeted Metabolomics. PLoS ONE 2020, 15, e0233805. [Google Scholar] [CrossRef]
- Fanelli, G.; Coleine, C.; Gevi, F.; Onofri, S.; Selbmann, L.; Timperio, A.M. Metabolomics of Dry Versus Reanimated Antarctic Lichen-Dominated Endolithic Communities. Life 2021, 11, 96. [Google Scholar] [CrossRef]
- Rochfort, S.; Ezernieks, V.; Mele, P.; Kitching, M. NMR Metabolomics for Soil Analysis Provide Complementary, Orthogonal Data to MIR and Traditional Soil Chemistry Approaches—A Land Use Study. Magn. Reson. Chem. 2015, 53, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Johns, C.W.; Lee, A.B.; Springer, T.I.; Rosskopf, E.N.; Hong, J.C.; Turechek, W.; Kokalis-Burelle, N.; Finley, N.L. Using NMR-Based Metabolomics to Monitor the Biochemical Composition of Agricultural Soils: A Pilot Study. Eur. J. Soil Biol. 2017, 83, 98–105. [Google Scholar] [CrossRef]
- Soucémarianadin, L.N.; Erhagen, B.; Nilsson, M.B.; Öquist, M.G.; Immerzeel, P.; Schleucher, J. Two Dimensional NMR Spectroscopy for Molecular Characterization of Soil Organic Matter: Application to Boreal Soils and Litter. Org. Geochem. 2017, 113, 184–195. [Google Scholar] [CrossRef]
- Abakumov, E.; Yaneva, R.; Polyakov, V.; Zhiyanski, M. Characterization of Humic Acids Isolated from Selected Soils of Livingston Island by CP/MAS 13C NMR and ESR Spectroscopy. Appl. Environ. Soil Sci. 2022, 2022, e7540077. [Google Scholar] [CrossRef]
- Abakumov, E. Characterisation of Humic Acids, Isolated from Selected Sub-Antarctic Soils by 13C-NMR Spectroscopy. Czech Polar Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Abakumov, E.; Alekseev, I. Stability of Soil Organic Matter in Cryosols of the Maritime Antarctic: Insights from 13C NMR and Electron Spin Resonance Spectroscopy. Solid Earth 2018, 9, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Severgnini, M.; Canini, F.; Consolandi, C.; Camboni, T.; Paolo D’Acqui, L.; Mascalchi, C.; Ventura, S.; Zucconi, L. Highly Differentiated Soil Bacterial Communities in Victoria Land Macro-Areas (Antarctica). FEMS Microbiol. Ecol. 2021, 97, fiab087. [Google Scholar] [CrossRef]
- Ciaramelli, C.; Palmioli, A.; Airoldi, C. Coffee Variety, Origin and Extraction Procedure: Implications for Coffee Beneficial Effects on Human Health. Food Chem. 2019, 278, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Palmioli, A.; Alberici, D.; Ciaramelli, C.; Airoldi, C. Metabolomic Profiling of Beers: Combining 1H NMR Spectroscopy and Chemometric Approaches to Discriminate Craft and Industrial Products. Food Chem. 2020, 327, 127025. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Orlando, F.; Palmioli, A.; Alali, S.; Airoldi, C.; De Noni, I.; Vaglia, V.; Bocchi, S.; Iriti, M. Different Phytotoxic Effect of Lolium Multiflorum Lam. Leaves against Echinochloa Oryzoides (Ard.) Fritsch and Oriza Sativa L. Environ. Sci. Pollut. Res. 2020, 27, 33204–33214. [Google Scholar] [CrossRef] [PubMed]
- Patti, F.; Palmioli, A.; Vitalini, S.; Bertazza, L.; Redaelli, M.; Zorzan, M.; Rubin, B.; Mian, C.; Bertolini, C.; Iacobone, M.; et al. Anticancer Effects of Wild Mountain Mentha Longifolia Extract in Adrenocortical Tumor Cell Models. Front. Pharmacol. 2020, 10, 1647. [Google Scholar] [CrossRef] [Green Version]
- Ciaramelli, C.; Palmioli, A.; De Luigi, A.; Colombo, L.; Sala, G.; Salmona, M.; Airoldi, C. NMR-Based Lavado Cocoa Chemical Characterization and Comparison with Fermented Cocoa Varieties: Insights on Cocoa’s Anti-Amyloidogenic Activity. Food Chem. 2021, 341, 128249. [Google Scholar] [CrossRef] [PubMed]
- Ciaramelli, C.; Palmioli, A.; Angotti, I.; Colombo, L.; De Luigi, A.; Sala, G.; Salmona, M.; Airoldi, C. NMR-Driven Identification of Cinnamon Bud and Bark Components With Anti-Aβ Activity. Front. Chem. 2022, 10, 896253. [Google Scholar] [CrossRef]
- de Divitiis, M.; Ami, D.; Pessina, A.; Palmioli, A.; Sciandrone, B.; Airoldi, C.; Regonesi, M.E.; Brambilla, L.; Lotti, M.; Natalello, A.; et al. Cheese-Whey Permeate Improves the Fitness of Escherichia Coli Cells during Recombinant Protein Production. Biotechnol. Biofuels Bioprod. 2023, 16, 30. [Google Scholar] [CrossRef]
- Airoldi, C.; Ciaramelli, C.; Palmioli, A. SMA Libraries for Metabolite Identification and Quantification in Antarctic Soils. Bicocca Open Archive Research Data, 2023, V1, doi:10.17632/h2pbhgv9wc.1. Available online: https://board.unimib.it/datasets/h2pbhgv9wc/1 (accessed on 1 July 2023).
- Greer, B.; Chevallier, O.; Quinn, B.; Botana, L.M.; Elliott, C.T. Redefining Dilute and Shoot: The Evolution of the Technique and Its Application in the Analysis of Foods and Biological Matrices by Liquid Chromatography Mass Spectrometry. TrAC Trends Anal. Chem. 2021, 141, 116284. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- RoyChowdhury, T.; Bramer, L.M.; Brown, J.; Kim, Y.-M.; Zink, E.; Metz, T.O.; McCue, L.A.; Diefenderfer, H.L.; Bailey, V. Soil Metabolomics Predict Microbial Taxa as Biomarkers of Moisture Status in Soils from a Tidal Wetland. Microorganisms 2022, 10, 1653. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.M.; Tfaily, M.M.; Kolton, M.; Johnston, E.R.; Petro, C.; Zalman, C.A.; Hanson, P.J.; Heyman, H.M.; Kyle, J.E.; Hoyt, D.W.; et al. Soil Metabolome Response to Whole-Ecosystem Warming at the Spruce and Peatland Responses under Changing Environments Experiment. Proc. Natl. Acad. Sci. USA 2021, 118, e2004192118. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.-H.M. The Strengths and Weaknesses of NMR Spectroscopy and Mass Spectrometry with Particular Focus on Metabolomics Research. In Metabonomics: Methods and Protocols; Bjerrum, J.T., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 161–193. ISBN 978-1-4939-2377-9. [Google Scholar]
- Zeki, Ö.C.; Eylem, C.C.; Reçber, T.; Kır, S.; Nemutlu, E. Integration of GC–MS and LC–MS for Untargeted Metabolomics Profiling. J. Pharm. Biomed. Anal. 2020, 190, 113509. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Intini, N.; Mastrorilli, P.; Latronico, M.; Scapicchio, P.; Triggiani, M.; Bevilacqua, V.; Fanizzi, P.; Acquotti, D.; Airoldi, C.; et al. Performance Assessment in Fingerprinting and Multi Component Quantitative NMR Analyses. Anal. Chem. 2015, 87, 6709–6717. [Google Scholar] [CrossRef]
- Gallo, V.; Ragone, R.; Musio, B.; Todisco, S.; Rizzuti, A.; Mastrorilli, P.; Pontrelli, S.; Intini, N.; Scapicchio, P.; Triggiani, M.; et al. A Contribution to the Harmonization of Non-Targeted NMR Methods for Data-Driven Food Authenticity Assessment. Food Anal. Methods 2020, 13, 530–541. [Google Scholar] [CrossRef]
- Musio, B.; Ragone, R.; Todisco, S.; Rizzuti, A.; Latronico, M.; Mastrorilli, P.; Pontrelli, S.; Intini, N.; Scapicchio, P.; Triggiani, M.; et al. A Community-Built Calibration System: The Case Study of Quantification of Metabolites in Grape Juice by QNMR Spectroscopy. Talanta 2020, 214, 120855. [Google Scholar] [CrossRef]
- Thevelein, J.M. Regulation of Trehalose Mobilization in Fungi. Microbiol. Rev. 1984, 48, 42–59. [Google Scholar] [CrossRef]
- Marquis, B.J.; Louks, H.P.; Bose, C.; Wolfe, R.R.; Singh, S.P. A New Derivatization Reagent for HPLC–MS Analysis of Biological Organic Acids. Chromatographia 2017, 80, 1723–1732. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Li, L. Chemical Derivatization in LC-MS-Based Metabolomics Study. TrAC Trends Anal. Chem. 2020, 131, 115988. [Google Scholar] [CrossRef]
- Willacey, C.C.W.; Naaktgeboren, M.; Lucumi Moreno, E.; Wegrzyn, A.B.; van der Es, D.; Karu, N.; Fleming, R.M.T.; Harms, A.C.; Hankemeier, T. LC–MS/MS Analysis of the Central Energy and Carbon Metabolites in Biological Samples Following Derivatization by Dimethylaminophenacyl Bromide. J. Chromatogr. A 2019, 1608, 460413. [Google Scholar] [CrossRef]
- Airoldi, C.; Ciaramelli, C.; Palmioli, A. SMA Libraries for Metabolite Identification and Quantification in Coffee Extracts. Bicocca Open Archive Research Data, 2018, V1. doi:10.17632/fs3vf7jbg5.1. Available online: https://board.unimib.it/datasets/fs3vf7jbg5/1 (accessed on 1 July 2023).
- Airoldi, C.; Palmioli, A.; Ciaramelli, C.; Alberici, D. SMA Libraries for Metabolite Identification and Quantification in Beers. Mendeley Data, 2019, V2, doi:10.17632/wvr6mccwwx.2. Available online: https://data.mendeley.com/datasets/wvr6mccwwx/2 (accessed on 1 July 2023).
- Palmioli, A.; Airoldi, C. SMA Libraries for Metabolite Identification and Quantification in Mentha Longifolia Extracts. Bicocca Open Archive Research Data, 2019, V1. doi:10.17632/hz2yt8khf8.1. Available online: https://board.unimib.it/datasets/hz2yt8khf8/1 (accessed on 1 July 2023).
- Palmioli, A.; Airoldi, C. SMA Libraries for Metabolite Identification and Quantification in Lolium Multiflorum Extracts. Bicocca Open Archive Research Data, 2019, V1. doi:10.17632/pxyk95g5j6.1. Available online: https://board.unimib.it/datasets/pxyk95g5j6/1 (accessed on 1 July 2023).
- Airoldi, C.; Ciaramelli, C.; Palmioli, A. SMA Libraries for Metabolite Identification and Quantification in Cocoa Extracts. Bicocca Open Archive Research Data, 2020, V1. doi:10.17632/z2k7z5wtg8.1. Available online: https://board.unimib.it/datasets/z2k7z5wtg8/1 (accessed on 1 July 2023).
- Airoldi, C.; Ciaramelli, C.; Palmioli, A. SMA Libraries for Metabolite NMR-Based Identification and Quantification in Cinnamon Extracts. Bicocca Open Archive Research Data, 2021, V1, doi:10.17632/v3kj6zn9tf.1. Available online: https://board.unimib.it/datasets/v3kj6zn9tf/1 (accessed on 1 July 2023).
- Palmioli, A.; Airoldi, C. SMA Library for Metabolites Identification and Quantification in Cheese Whey Permeate (CWP). Bicocca Open Archive Research Data, 2022, V1. doi:10.17632/xn5ntwbtjz.1. Available online: https://board.unimib.it/datasets/xn5ntwbtjz/1 (accessed on 1 July 2023).
- Airoldi, C.; Ciaramelli, C.; Fumagalli, M.; Bussei, R.; Mazzoni, V.; Viglio, S.; Iadarola, P.; Stolk, J. 1H NMR To Explore the Metabolome of Exhaled Breath Condensate in A1-Antitrypsin Deficient Patients: A Pilot Study. J. Proteome Res. 2016, 15, 4569–4578. [Google Scholar] [CrossRef] [PubMed]
- Ciaramelli, C.; Fumagalli, M.; Viglio, S.; Bardoni, A.M.; Piloni, D.; Meloni, F.; Iadarola, P.; Airoldi, C. 1H NMR To Evaluate the Metabolome of Bronchoalveolar Lavage Fluid (BALf) in Bronchiolitis Obliterans Syndrome (BOS): Toward the Development of a New Approach for Biomarker Identification. J. Proteome Res. 2017, 16, 1669–1682. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Geographic Name | Coordinates | Type of Crust |
|---|---|---|---|
| Ap.I 1 | Apostrophe Island site 1 | CS—73°31′09.5″ S 167°25′55.3″ E—41 m asl | Grey crust with few mosses |
| Ap.I 2 | Apostrophe Island site 2 | CS—73°31′10.5″ S 167°25′55.3″ E—41 m asl | Brown thin crust with mosses and lichens |
| B.By 1 | Botany Bay site 1 | CS—77°00′26.0″ S 162°32′39.4″ E—115 m asl | Black thin crust disconnected from the below soil |
| B.By 2 | Botany Bay site 2 | CS—77°00′26.7″ S 162°32′40.5″ E—94 m asl | Black highly developed crust |
| Cp.K 1 | Cape King site 1 | CS—73°35′08.2″ S 166°37′19.2″ E—144 m asl | Black highly developed crust with mosses |
| Cp.K 2 | Cape King site 2 | CS—73°35′08.9″ S 166°37′3.5″ E—124 m asl | Dry thin and black crust with very few mosses |
| Ed.P 2 | Edmonson Point site 2 | CS—74°19′44.9″ S 165°07′39.9″ E—29 m asl | Thin signs of biological colonisation |
| Ed.P 3 | Edmonson Point site 3 | CS—74°19′45.2″ S 165°07′35.8″ E—31 m asl | Medium developed crust |
| Ed.P 4 | Edmonson Point site 4 | CS—74°19′45.1″ S 165°07′38.7″ E—30 m asl | Well-developed crust completely covered by mosses and lichens |
| Ky.I 1 | Kay Island site 1 | CS—74°04′12.6″ S 165°18′59.5″ E—190 m asl | Low developed crust with some mosses |
| Ky.I 2 | Kay Island site 2 | CS—74°04′11.8″ S 165°18′58.7″ E—61 m asl | Well-developed crust with more mosses |
| Pr.I 1 | Prior Island site 1 | CS—75°40′52.9″ S 162°53′38.3″ E—102 m asl | Grey thin and low developed crust |
| Pr.I 2 | Prior Island site 2 | CS—75°40′54.5″ S 162°53′45.8″ E—98 m asl | Wet highly developed crust dominated by mosses |
| Lk.F 1.1 | Lake Fryxell site 1 sample 1 | IS—77°36′7.2″ S 163°16′5.4″ E—28 m asl | Diffuse superficial crust, with saline efflorescence |
| Lk.F 1.2 | Lake Fryxell site 1 sample 2 | “ | “ |
| Lk.F 1.3 | Lake Fryxell site 1 sample 3 | “ | “ |
| Lk.H 1.1 | Lake Hoare site 1 sample 1 | IS—77°37′26.2″ S 162°53′27.8″ E—83 m asl | Low developed crust, with saline efflorescence |
| Lk.H 1.2 | Lake Hoare site 1 sample 2 | “ | “ |
| Lk.J 1.1 | Lake Joyce site 1 sample 1 | IS—77°42′21.1″ S 161°34′14.3″ E—448 m asl | Whitish consistent crusts on a sandy soil |
| Lk.J 1.2 | Lake Joyce site 1 sample 2 | “ | “ |
| Metabolite | Assignment | 1H Chemical Shift |
|---|---|---|
| Acetate | CH3 | 1.92 (s) |
| Acetone | 2 × CH3 | 2.23 (s) |
| Acetonitrile | CH3 | 2.07 (s) |
| Alanine | CHα | 3.79 (m) |
| CH3 | 1.48 (d) | |
| Aspartate | CHα | 3.9 (m) |
| CH2β | 2.69 (m)–2.81 (dd) | |
| Betaine | CH | 3.89 (m) |
| 3 × CH3 | 3.25 (m) | |
| Butyric acid | CH2 (1) | 2.16 (t) |
| CH2 (2) | 1.56 (h) | |
| CH3 | 0.90 (t) | |
| Choline | 2 × CH3 | 3.21 (s) |
| Citrate | CH2 (2), CH2 (6) | 2.65 (d)–2.56 (d) |
| Fatty acid | CH2 | 2.2 (t) |
| CH2 (3) | 1.55 (m) | |
| CH2 (ω2) | 1.30 (m) | |
| CH3 (ω0) | 0.90 (m) | |
| Formate | O=C-H | 8.46 (s) |
| GABA (γ-Aminobutyric acid) | CH (1) | 3.00 (t) |
| CH (3) | 2.30 (t) | |
| CH (2) | 1.89 (m) | |
| α-D-Glucose | CH (1) | 5.24 (d) |
| CH (2) | 3.85 (m) | |
| CH2 (6) | 3.82–3.76 (m) | |
| CH (3) | 3.72 (m) | |
| CH (5) | 3.54 (m) | |
| CH (4) | 3.42 (m) | |
| β-D-Glucose | CH (1) | 4.65 (d) |
| CH2 (6) | 3.71–3.89 (m) | |
| CH (3), CH (5) | 3.47 (m) | |
| CH (4) | 3.41 (m) | |
| CH (2) | 3.24 (m) | |
| Glutamate | CHα | 3.76 (m) |
| CH2γ | 2.34 (m) | |
| CH2β | 2.08 (m) | |
| Glycine | CH2 | 3.56 (s) |
| 4-Hydroxybenzoic acid | CH (2, 6) | 7.81 (d) |
| CH (3, 5) | 6.91 (d) | |
| Isobutyric acid | 2 × CH3 | 1.06 (d) |
| CH | 2.39 (m) | |
| Isoleucine | CHα | 3.71 (d) |
| CH | 1.97 (m) | |
| CH2 | 1.25–1.46 (m) | |
| CH3 (7) | 1.00 (d) | |
| CH3 (6) | 0.91 (t) | |
| Lactate | CH | 4.11 (q) |
| CH3 | 1.33 (d) | |
| Leucine | CHα | 3.72 (d) |
| CH2 + CH | 1.71 (m) | |
| 2 × CH3 | 0.95 (t) | |
| Malic acid | CH2 (3) | 2.36 (dd)–2.67 (dd) |
| Methanol | CH3 | 3.36 (s) |
| 4-Methylbenzoic acid | CH (2, 6) | 7.78 (d) |
| CH (3, 5) | 7.32 (d) | |
| CH3 | 2.39 (s) | |
| Phenylalanine | CH (7), CH (9) | 7.43 (d) |
| CH (8) | 7.38 (m) | |
| CH (6), CH (10) | 7.32 (d) | |
| CHα | 3.98 (m) | |
| CH2β | 3.10 (dd)–3.27 (dd) | |
| Succinate | 2 × CH2 | 2.39 (s) |
| Sucrose | CH (7) (Glc) | 5.42 (d) |
| CH (3) (Fru) | 4.22 (d) | |
| CH (4) (Fru) | 4.06 (t) | |
| CH (5) (Fru) | 3.88 (m) | |
| CH (9) (Glc) | 3.83 (m) | |
| CH 17+19 | 3.81 (m) | |
| CH (11) (Glc) | 3.75 (t) | |
| CH (13) (Fru) | 3.67 (s) | |
| CH (12) (Glc) | 3.56 (dd) | |
| CH 1(0) (Glc) | 3.47 (t) | |
| Trehalose | CH (1) | 5.20 (d) |
| CH2 (6) | 3.85 (m) | |
| CH (3), CH (5) | 3.77 (m) | |
| CH (4) | 3.66 (dd) | |
| CH (2) | 3.46 (t) | |
| Tyrosine | CH (7), CH (10) | 7.20 (d) |
| CH (6), CH (11) | 6.90 (d) | |
| CHα | 3.92 (m) | |
| CH2β | 3.04 (dd)–3.18 (dd) | |
| Uridine | CH (6) | 7.86 (m) |
| CH (5) pyrimidine + CH (1′) ribose | 5.92 (m) | |
| CH (2′) Ribose | 4.36 (m) | |
| CH (3′) Ribose | 4.24 (m) | |
| CH (4′) Ribose | 4.12 (m) | |
| Valine | CHα | 3.60 (d) |
| CH (2) | 2.26 (m) | |
| CH3 (6) | 1.04 (d) | |
| CH3 (3) | 0.98 (d) |
| Acetate | Acetone | Alanine | Aspartate | Butyric Acid | Choline | Formate | GABA | Glucose | Glycine | ||
| Ap.I 1 | 11.89 | 2.47 | ND | ND | ND | 2.17 | 2.37 | ND | ND | ND | |
| Ap.I 2 | 7.05 | 4.07 | ND | ND | ND | 1.47 | 2.23 | ND | ND | ND | |
| B.By 1 | 11.77 | 1.48 | ND | ND | ND | 2.08 | 4.88 | ND | ND | ND | |
| B.By 2 | 12.73 | 3.32 | 7.13 | ND | ND | 8.26 | ND | ND | 80.47 | ND | |
| Cp.K 1 | 10.49 | 3.74 | 12.35 | ND | ND | 3.34 | ND | ND | ND | ND | |
| Cp.K 2 | 8.45 | 1.39 | ND | ND | ND | 0.77 | 1.10 | ND | ND | ND | |
| Ed.P 2 | 57.61 | 8.87 | 47.57 | 10.20 | ND | 16.11 | 6.66 | 7.84 | 391.54 | ND | |
| Ed.P 3 | 91.08 | 1.00 | 10.69 | ND | 65.44 | 1.78 | 1.55 | 12.58 | ND | 5.10 | |
| Ed.P 4 | 130.11 | 1.96 | 19.46 | ND | 98.98 | 4.26 | 1.08 | 19.10 | ND | 6.21 | |
| Ky.I 1 | 76.82 | 6.54 | 56.36 | 15.00 | ND | ND | 11.81 | 8.87 | 191.57 | ND | |
| Ky.I 2 | 13.17 | 4.03 | 14.91 | ND | ND | 7.15 | 5.06 | ND | 159.86 | 17.32 | |
| Pr.I 1 | 5.56 | 1.78 | 4.99 | ND | ND | 4.55 | 2.78 | ND | ND | 7.76 | |
| Pr.I 2 | 164.50 | 3.71 | 16.10 | ND | ND | 4.99 | ND | 21.66 | 73.50 | 14.81 | |
| Lk.F 1.1 | 6.21 | ND | ND | ND | ND | 2.51 | ND | ND | ND | ND | |
| Lk.F 1.2 | 11.17 | ND | ND | ND | ND | 1.10 | 1.99 | ND | ND | 2.67 | |
| Lk.F 1.3 | 5.36 | 0.73 | ND | ND | ND | 0.40 | 1.11 | ND | ND | ND | |
| Lk.H 1.1 | 6.41 | 1.73 | ND | ND | ND | 3.36 | ND | ND | ND | ND | |
| Lk.H 1.2 | 4.48 | 1.70 | ND | ND | ND | 2.68 | ND | ND | ND | ND | |
| Lk.J 1.1 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Lk.J 1.2 | 0.83 | 0.98 | ND | ND | ND | ND | ND | ND | ND | ND | |
| 4-Hydroxybenzoic Acid | Isobutyric Acid | Lactate | Methanol | 4-Methylbenzoic Acid | Phenylalanine | Succinate | Sucrose | Trehalose | Tyrosine | Valine | |
| Ap.I 1 | ND | 2.01 | 15.19 | 1.04 | 10.98 | ND | 7.66 | ND | 73.02 | ND | ND |
| Ap.I 2 | ND | ND | 16.45 | 3.05 | ND | ND | ND | 30.12 | 109.76 | ND | ND |
| B.By 1 | ND | ND | 10.33 | 7.05 | ND | ND | 1.53 | ND | 46.32 | ND | ND |
| B.By 2 | ND | 1.57 | 13.87 | 1.29 | 12.25 | ND | 10.55 | 99.95 | 221.58 | ND | ND |
| Cp.K 1 | ND | 1.42 | 34.71 | 2.35 | ND | ND | ND | ND | 113.41 | ND | ND |
| Cp.K 2 | ND | 1.30 | 8.77 | 0.38 | 7.13 | ND | 4.87 | ND | 25.56 | ND | ND |
| Ed.P 2 | 10.50 | ND | 86.18 | ND | ND | 24.23 | ND | 138.75 | 114.56 | 11.54 | 29.52 |
| Ed.P 3 | 0.93 | 0.55 | 9.25 | 0.30 | ND | ND | 1.08 | ND | ND | ND | ND |
| Ed.P 4 | 2.38 | 2.54 | 13.92 | 0.21 | ND | 8.33 | 2.58 | 7.57 | 20.50 | 4.07 | ND |
| Ky.I 1 | ND | ND | 68.40 | 3.57 | ND | 83.92 | ND | 133.95 | 153.12 | 35.15 | 59.98 |
| Ky.I 2 | ND | ND | 34.05 | 15.40 | ND | ND | ND | ND | 319.02 | ND | ND |
| Pr.I 1 | ND | ND | 15.79 | 12.69 | ND | ND | ND | 49.75 | 43.81 | ND | ND |
| Pr.I 2 | ND | 5.73 | 16.51 | 1.29 | ND | 28.74 | 4.82 | ND | 92.65 | ND | ND |
| Lk.F 1.1 | ND | ND | 3.07 | 5.68 | ND | ND | 3.21 | ND | 15.38 | ND | ND |
| Lk.F 1.2 | ND | 1.67 | 10.69 | 1.49 | 12.71 | ND | 11.65 | ND | 39.48 | ND | ND |
| Lk.F 1.3 | ND | ND | 4.81 | 6.28 | ND | ND | 1.76 | ND | 12.37 | ND | ND |
| Lk.H 1.1 | ND | ND | 3.77 | 5.87 | ND | ND | 1.36 | ND | 17.34 | ND | ND |
| Lk.H 1.2 | ND | ND | 5.69 | 0.07 | 4.43 | ND | 3.78 | ND | 8.63 | ND | ND |
| Lk.J 1.1 | ND | ND | ND | 0.53 | ND | ND | ND | ND | ND | ND | ND |
| Lk.J 1.2 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| ID | Rt (min) | Average (m/z) | Metabolite Name | Adduct Type | Reference (m/z) | Error (ppm) | Formula | Ontology | INCHIKEY | Sample | Possible Sources a |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2.09 | 268.1044 | Adenosine | [M+H]+ | 268.1046 | −0.45 | C10H13N5O4 | Purine nucleosides | OIRDTQYFTABQOQ-KQYNXXCUSA-N | Ed.P, Ky.I | Ubiquitous |
| 2 | 3.20 | 120.0812 | Phenylethanolamine | [M−H2O+H]+ | 120.0808 | 3.53 | C8H11NO | Aralkylamines | ULSIYEODSMZIPX-UHFFFAOYSA-N | Ed.P, Ky.I | Ubiquitous |
| 3 | 4.00 | 181.0502 | 4-Hydroxyphenyllactic acid | [M−H]− | 181.0506 | −2.39 | C9H10O4 | Benzenoids | HXIPUYVSSGKLFF-UHFFFAOYSA-N | Ed.P | Microbes |
| 4 | 4.27 | 137.0244 | 4-Hydroxybenzoic acid | [MH]− | 137.0244 | 0.00 | C7H6O3 | Hydroxybenzoic acid derivatives | FJKROLUGYXJWQN-UHFFFAOYSA-N | Ed.P | Plants and microbes |
| 5 | 4.31 | 609.1466 | Luteolin 6-C-glucoside 8-C-arabinoside | [M−H]− | 609.1461 | 0.71 | C27H30O16 | Flavonoid 8-C-glycosides | ZLPSOQFIIQIIAX-UHFFFAOYNA-N | Ed.P | Plants |
| 6 | 4.51 | 159.0664 | 3-Methyladipic acid | [M−H]− | 159.0663 | 0.31 | C7H12O4 | Medium-chain fatty acids | SYEOWUNSTUDKGM-UHFFFAOYSA-N | Ed.P | Animals |
| 7 | 4.53 | 593.1517 | Vicenin 2 | [M−H]− | 593.1512 | 0.93 | C27H30O15 | Flavonoid 8-C-glycosides | FIAAVMJLAGNUKW-UHFFFAOYSA-N | Ed.P | Plants |
| 595.1667 | [M−H]+ | 595.1658 | 1.55 | ||||||||
| 8 | 4.62 | 377.14664 | (-)-Riboflavin | [M+H]+ | 377.1461 | 1.46 | C17H20N4O6 | Flavins | AUNGANRZJHBGPY-SCRDCRAPSA-N | Ed.P, Ky.I | Ubiquitous |
| 9 | 4.74 | 213.9642 | 2-Benzothiazolesulfonic acid | [M−H]− | 213.9638 | 1.82 | C7H5NO3S2 | Benzothiazoles | ZCXGMSGCBDSEOY-UHFFFAOYSA-N | Ed.P, Lk.F | Contaminant |
| 215.9790 | [M+H]+ | 215.9784 | 2.92 | ||||||||
| 10 | 5.22 | 173.0816 | Suberic acid | [M−H]− | 173.0819 | −1.91 | C8H14O4 | Medium-chain fatty acids | TYFQFVWCELRYAO-UHFFFAOYSA-N | Ed.P, Ky.I | - |
| 11 | 5.84 | 271.1550 | C14H24O5 | [M−H]− | 271.1551 | −0.37 | C14H24O5 | TBD b | TBD b | Ed.P | - |
| 12 | 6.54 | 236.0973 | C10H15N5S | [M−H]− | 236.0975 | −0.85 | C10H15N5S | TBD b | TBD b | Ed.P, Lk.F | - |
| 13 | 6.84 | 421.1860 | C22H30O8 | [M−H]− | 421.1868 | −1.90 | C22H30O8 | TBD b | TBD b | Ed.P, Lk.F | - |
| 14 | 7.11 | 195.0300 | Haematommic acid | [M−H]− | 195.0299 | 0.43 | C9H8O5 | Hydroxybenzoic acids | KCOOTJRKKIDHTM-UHFFFAOYSA-N | Ky.I | Lichens: Asahinea chrysantha |
| 15 | 7.19 | 269.0452 | Apigenin | [M−H]− | 269.0450 | 0.82 | C15H10O5 | Flavones | KZNIFHPLKGYRTM-UHFFFAOYSA-N | Ed.P, Ky.I | Plants |
| 271.0608 | [M+H]+ | 271.0601 | 2.58 | ||||||||
| 16 | 7.37 | 299.0564 | Kaempferide | [M−H]− | 299.0561 | 0.94 | C16H12O6 | Flavonols | SQFSKOYWJBQGKQ-UHFFFAOYSA-N | Ed.P | Plants |
| 301.0712 | [M+H]+ | 301.0712 | 0.10 | ||||||||
| 17 | 7.51 | 237.1493 | C14H20O3 | [M+H]+ | 237.1485 | 3.37 | C14H20O3 | TBD b | TBD b | Ed.P | - |
| 235.1342 | [M−H]− | 235.1340 | 0.85 | ||||||||
| 18 | 7.69 | 299.0194 | Emodic acid | [M−H]− | 299.0197 | 0.95 | C15H8O7 | Anthracenecarboxylic acids | ZJXVNNSMRGTDBI-UHFFFAOYSA-N | Ky.I | Lichens and Fungi |
| 19 | 8.15 | 323.0557 | Sterigmatocystin | [M−H]− | 323.0561 | −1.24 | C18H12O6 | Sterigmatocystins | UTSVPXMQSFGQTM-UHFFFAOYNA-N | Ky.I | Fungi: Aspergillus |
| 20 | 8.47 | 371.0401 | Norstictic acid | [M−H]− | 341.0409 | −2.03 | C18H12O9 | Diarylethers | IEVVSJFLBYOUCJ-UHFFFAOYSA-N | Ky.I | Lichens: Buellia, Dimelaena, Usnea |
| 21 | 9.02 | 313.0352 | Parietinic acid | [M−H]− | 313.0350 | 0.61 | C16H10O7 | Anthracenecarboxylic acids | HEULMVKOOVHXME-UHFFFAOYSA-N | Ky.I | Lichens: Xanthoria, Fulgensia |
| 22 | 9.04 | 307.0613 | Pulvinic acid | [M−H]− | 307.0610 | 0.81 | C18H12O5 | Butenolides | CMFBGFRHPQTELQ-JQIJEIRASA-N | Ky.I | Lichens: Letharia, Candelaria. Fungi: Aspergillus |
| 23 | 9.09 | 305.0448 | Calycin | [M−H]− | 305.0450 | −0.69 | C18H10O5 | Benzofurans | CGRCGRBHNKRILW-JQIJEIRASA-N | Ky.I | Lichens: Candelaria, Pseudocyphellaria |
| 24 | 9.70 | 269.0454 | Emodin | [M−H]− | 269.0455 | −0.22 | C15H10O5 | Hydroxyanthraquinones | RHMXXJGYXNZAPX-UHFFFAOYSA-N | Ky.I | Plants, Fungi and crustose lichen |
| 25 | 9.71 | 275.2009 | (9S,13S)-12-Oxophytodienoate | [M−H2O+H]+ | 275.2006 | 1.20 | C18H28O3 | Prostaglandins and related compounds | PMTMAFAPLCGXGK-TTXFDSJOSA-N | Ed.P | Plants |
| 293.2119 | [M+H]+ | 293.2111 | 2.66 | ||||||||
| 26 | 11.11 | 297.2426 | Lichesterylic acid | [M−H]− | 297.2435 | −3.16 | C18H34O3 | Long-chain fatty acids | FYKXUGIGKHKTDH-UHFFFAOYSA-N | Ed.P | Lichens |
| 27 | 11.17 | 374.9593 | 6-O-Methylarthothelin | [M+H]+ | 374.9580 | 3.49 | C15H9Cl3O5 | Xanthones | SMLUHOHPDVBXKH-UHFFFAOYSA-N | Ky.I | Lichen: Lecanora hybocarpa, Dimelaena |
| 372.9445 | [M−H]− | 372.9440 | 1.31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciaramelli, C.; Palmioli, A.; Brioschi, M.; Viglio, S.; D’Amato, M.; Iadarola, P.; Tosi, S.; Zucconi, L.; Airoldi, C. Antarctic Soil Metabolomics: A Pilot Study. Int. J. Mol. Sci. 2023, 24, 12340. https://doi.org/10.3390/ijms241512340
Ciaramelli C, Palmioli A, Brioschi M, Viglio S, D’Amato M, Iadarola P, Tosi S, Zucconi L, Airoldi C. Antarctic Soil Metabolomics: A Pilot Study. International Journal of Molecular Sciences. 2023; 24(15):12340. https://doi.org/10.3390/ijms241512340
Chicago/Turabian StyleCiaramelli, Carlotta, Alessandro Palmioli, Maura Brioschi, Simona Viglio, Maura D’Amato, Paolo Iadarola, Solveig Tosi, Laura Zucconi, and Cristina Airoldi. 2023. "Antarctic Soil Metabolomics: A Pilot Study" International Journal of Molecular Sciences 24, no. 15: 12340. https://doi.org/10.3390/ijms241512340
APA StyleCiaramelli, C., Palmioli, A., Brioschi, M., Viglio, S., D’Amato, M., Iadarola, P., Tosi, S., Zucconi, L., & Airoldi, C. (2023). Antarctic Soil Metabolomics: A Pilot Study. International Journal of Molecular Sciences, 24(15), 12340. https://doi.org/10.3390/ijms241512340