

IL-21/IL-21R Promotes the Pro-Inflammatory Effects of Macrophages during C. muridarum Respiratory Infection

Abstract
:1. Introduction
2. Results
2.1. IL-21/IL-21R Promotes Pulmonary Macrophage Infiltration following C.muridarum Respiratory Infection
2.2. Transcriptome Analysis Suggests the Participation of IL-21/IL-21R in M1 Polarization and Inflammatory Responses of Macrophages during Chlamydial Respiratory Infection
2.3. IL-21/IL-21R Is Required for Lung Macrophage Polarization towards M1 Phenotypes during C. muridarum Respiratory Infection
2.4. IL-21/IL-21R Promotes Pro-Inflammatory Cytokine Production from Pulmonary Macrophages during C.muridarum Respiratory Infection
2.5. IL-21/IL-21R Could Enhance the Function of M1-Type Macrophages during C.muridarum Infection
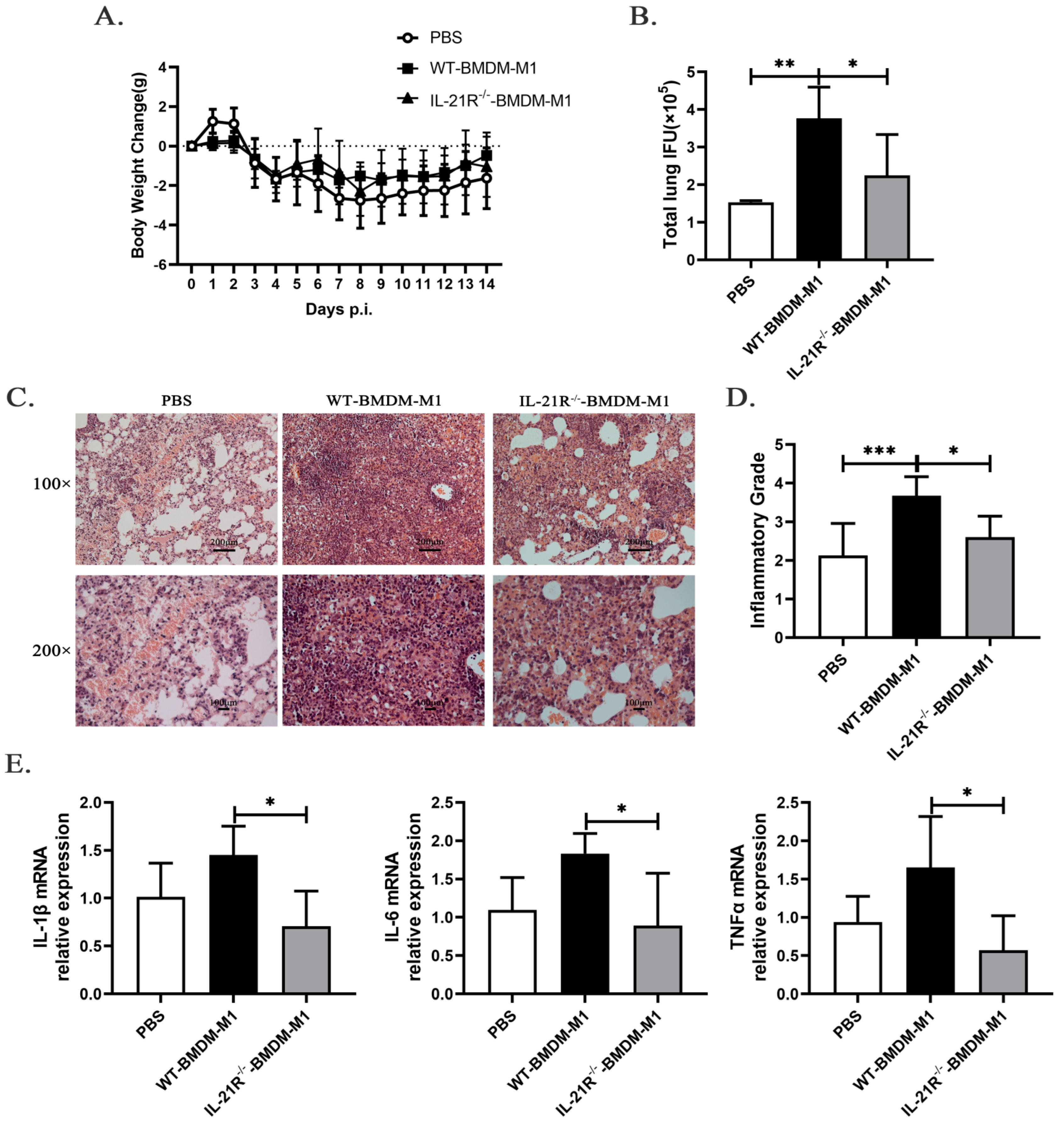
2.6. IL-21/IL-21R Might Aggravate M1-Mediated Pulmonary Inflammation following C. muridarum Infection
3. Materials and Methods
3.1. Mice
3.2. C.muridarum Respiratory Tract Infection Models
3.3. Lung Single Cells Preparation
3.4. RAW 264.7 Culture and Treatment
3.5. Flow Cytometry
3.6. Immunofluorescence Staining
3.7. Quantitative Real-Time PCR(qPCR)
3.8. NO Analysis
3.9. ROS Analysis
3.10. Administration of rmIL-21
3.11. Induction and Adoptive Transfer of M1-Type Bone Marrow-Derived Macrophages (BMDM-M1)
3.12. Pulmonary Chlamydia Loads and Histopathological Analysis
3.13. Bioinformatic Analysis
3.14. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collingro, A.; Köstlbacher, S.; Horn, M. Chlamydiae in the Environment. Trends Microbiol. 2020, 28, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Borel, N.; Polkinghorne, A.; Pospischil, A. A Review on Chlamydial Diseases in Animals: Still a Challenge for Pathologists? Vet. Pathol. 2018, 55, 374–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, G.; Straube, E.; Essig, A.; Reinhold, P.; Sachse, K. Chlamydial zoonoses. Dtsch. Arztebl. Int. 2010, 107, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, R.K.; Gupta, K.; Jordan, S.J.; Chi, X.; Lensing, S.Y.; Press, C.G.; Geisler, W.M. An Adaptive Chlamydia trachomatis-Specific IFN-gamma-Producing CD4(+) T Cell Response Is Associated with Protection against Chlamydia Reinfection in Women. Front. Immunol. 2018, 9, 1981. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; HayGlass, K.; Brunham, R.C. Genetically determined differences in IL-10 and IFN-gamma responses correlate with clearance of Chlamydia trachomatis mouse pneumonitis infection. J. Immunol. 1996, 156, 4338–4344. [Google Scholar] [CrossRef]
- Bai, H.; Cheng, J.; Gao, X.; Joyee, A.G.; Fan, Y.; Wang, S.; Jiao, L.; Yao, Z.; Yang, X. IL-17/Th17 promotes type 1 T cell immunity against pulmonary intracellular bacterial infection through modulating dendritic cell function. J. Immunol. 2009, 183, 5886–5895. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Yang, J.; Qiu, H.; Wang, S.; Fan, Y.; Han, X.; Xie, S.; Yang, X. Intranasal inoculation of Chlamydia trachomatis mouse pneumonitis agent induces significant neutrophil infiltration which is not efficient in controlling the infection in mice. Immunology 2005, 114, 246–254. [Google Scholar] [CrossRef]
- Zeng, J.; Xu, Y.; Tan, L.; Zha, X.; Yang, S.; Zhang, H.; Tuo, Y.; Sun, R.; Niu, W.; Pang, G.; et al. IL-21/IL-21R Regulates the Neutrophil-Mediated Pathologic Immune Response during Chlamydial Respiratory Infection. Mediat. Inflamm. 2022, 2022, 4322092. [Google Scholar] [CrossRef]
- Li, S.N.; Wang, W.; Fu, S.P.; Wang, J.F.; Liu, H.M.; Xie, S.S.; Liu, B.R.; Li, Y.; Lv, Q.K.; Li, Z.Q.; et al. IL-21 modulates release of proinflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Mediat. Inflamm. 2013, 2013, 548073. [Google Scholar] [CrossRef] [Green Version]
- Pidwill, G.R.; Gibson, J.F.; Cole, J.; Renshaw, S.A.; Foster, S.J. The Role of Macrophages in Staphylococcus aureus Infection. Front. Immunol. 2020, 11, 620339. [Google Scholar] [CrossRef]
- Hussell, T.; Bell, T.J. Alveolar macrophages: Plasticity in a tissue-specific context. Nat. Rev. Immunol. 2014, 14, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Franke-Ullmann, G.; Pförtner, C.; Walter, P.; Steinmüller, C.; Lohmann-Matthes, M.L.; Kobzik, L. Characterization of murine lung interstitial macrophages in comparison with alveolar macrophages in vitro. J. Immunol. 1996, 157, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Hou, F.; Xiao, K.; Tang, L.; Xie, L. Diversity of Macrophages in Lung Homeostasis and Diseases. Front. Immunol. 2021, 12, 753940. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wu, X.; Zhang, X.; Liu, X.; Deng, G. Heme oxygenase-1 modulates ferroptosis by fine-tuning levels of intracellular iron and reactive oxygen species of macrophages in response to Bacillus Calmette-Guerin infection. Front. Cell. Infect. Microbiol. 2022, 12, 1004148. [Google Scholar] [CrossRef] [PubMed]
- Chandra, P.; Grigsby, S.J.; Philips, J.A. Immune evasion and provocation by Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2022, 20, 750–766. [Google Scholar] [CrossRef] [PubMed]
- Surewaard, B.G.; Deniset, J.F.; Zemp, F.J.; Amrein, M.; Otto, M.; Conly, J.; Omri, A.; Yates, R.M.; Kubes, P. Identification and treatment of the Staphylococcus aureus reservoir in vivo. J. Exp. Med. 2016, 213, 1141–1151. [Google Scholar] [CrossRef]
- Jupelli, M.; Shimada, K.; Chiba, N.; Slepenkin, A.; Alsabeh, R.; Jones, H.D.; Peterson, E.; Chen, S.; Arditi, M.; Crother, T.R. Chlamydia pneumoniae infection in mice induces chronic lung inflammation, iBALT formation, and fibrosis. PLoS ONE 2013, 8, e77447. [Google Scholar] [CrossRef]
- Huang, L.; Nazarova, E.V.; Russell, D.G. Mycobacterium tuberculosis: Bacterial Fitness within the Host Macrophage. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef]
- Qin, S.; Li, J.; Zhou, C.; Privratsky, B.; Schettler, J.; Deng, X.; Xia, Z.; Zeng, Y.; Wu, H.; Wu, M. SHIP-1 Regulates Phagocytosis and M2 Polarization through the PI3K/Akt-STAT5-Trib1 Circuit in Pseudomonas aeruginosa Infection. Front. Immunol. 2020, 11, 307. [Google Scholar] [CrossRef] [Green Version]
- Mohamed Elashiry, M.; Tian, F.; Elashiry, M.; Zeitoun, R.; Elsayed, R.; Andrews, M.L.; Bergeon, B.E.; Cutler, C.; Tay, F. Enterococcus faecalis shifts macrophage polarization toward M1-like phenotype with an altered cytokine profile. J. Oral Microbiol. 2021, 13, 1868152. [Google Scholar] [CrossRef]
- Beam, J.E.; Maiocchi, S.; Cartaya, A.; Rowe, S.E.; Bahnson, E.S.; Conlon, B.P. The Use of Acute Immunosuppressive Therapy to Improve Antibiotic Efficacy against Intracellular Staphylococcus aureus. Microbiol. Spectr. 2022, 10, e0085822. [Google Scholar] [CrossRef]
- Hashimoto-Kataoka, T.; Hosen, N.; Sonobe, T.; Arita, Y.; Yasui, T.; Masaki, T.; Minami, M.; Inagaki, T.; Miyagawa, S.; Sawa, Y.; et al. Interleukin-6/interleukin-21 signaling axis is critical in the pathogenesis of pulmonary arterial hypertension. Proc. Natl. Acad. Sci. USA 2015, 112, E2677–E2686. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.M.; King, C. IL-21-producing Th cells in immunity and autoimmunity. J. Immunol. 2013, 191, 3501–3506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Jiang, X.; Zhu, J.; Yue, D.; Zhang, X.; Wang, X.; You, Y.; Wang, B.; Xu, Y.; Lu, C.; et al. IL-21/IL-21R signaling suppresses intestinal inflammation induced by DSS through regulation of Th responses in lamina propria in mice. Sci. Rep. 2016, 6, 31881. [Google Scholar] [CrossRef] [Green Version]
- Vallieres, F.; Girard, D. IL-21 enhances phagocytosis in mononuclear phagocyte cells: Identification of spleen tyrosine kinase as a novel molecular target of IL-21. J. Immunol. 2013, 190, 2904–2912. [Google Scholar] [CrossRef] [PubMed]
- Booty, M.G.; Barreira-Silva, P.; Carpenter, S.M.; Nunes-Alves, C.; Jacques, M.K.; Stowell, B.L.; Jayaraman, P.; Beamer, G.; Behar, S.M. IL-21 signaling is essential for optimal host resistance against Mycobacterium tuberculosis infection. Sci. Rep. 2016, 6, 36720. [Google Scholar] [CrossRef] [Green Version]
- Wanford, J.J.; Hames, R.G.; Carreno, D.; Jasiunaite, Z.; Chung, W.Y.; Arena, F.; Di Pilato, V.; Straatman, K.; West, K.; Farzand, R.; et al. Interaction of Klebsiella pneumoniae with tissue macrophages in a mouse infection model and ex-vivo pig organ perfusions: An exploratory investigation. Lancet Microbe 2021, 2, e695–e703. [Google Scholar] [CrossRef]
- Dodd, J.S.; Clark, D.; Muir, R.; Korpis, C.; Openshaw, P.J.M. Endogenous IL-21 regulates pathogenic mucosal CD4 T-cell responses during enhanced RSV disease in mice. Mucosal Immunol. 2013, 6, 704–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spolski, R.; West, E.E.; Li, P.; Veenbergen, S.; Yung, S.; Kazemian, M.; Oh, J.; Yu, Z.X.; Freeman, A.F.; Holland, S.M.; et al. IL-21/type I interferon interplay regulates neutrophil-dependent innate immune responses to Staphylococcus aureus. eLife 2019, 8, e45501. [Google Scholar] [CrossRef]
- Spolski, R.; Wang, L.; Wan, C.K.; Bonville, C.A.; Domachowske, J.B.; Kim, H.P.; Yu, Z.; Leonard, W.J. IL-21 promotes the pathologic immune response to pneumovirus infection. J. Immunol. 2012, 188, 1924–1932. [Google Scholar] [CrossRef] [Green Version]
- Niu, W.; Xu, Y.; Zha, X.; Zeng, J.; Qiao, S.; Yang, S.; Zhang, H.; Tan, L.; Sun, L.; Pang, G.; et al. IL-21/IL-21R Signaling Aggravated Respiratory Inflammation Induced by Intracellular Bacteria through Regulation of CD4(+) T Cell Subset Responses. J. Immunol. 2021, 206, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, Mechanisms, and Significance of Macrophage Plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Kees, T.; Almeida, A.S.; Liu, B.; He, X.-Y.; Ng, D.; Han, X.; Spector, D.L.; McNeish, I.A.; Gimotty, P.; et al. Activating a collaborative innate-adaptive immune response to control metastasis. Cancer Cell 2021, 39, 1361–1374.e9. [Google Scholar] [CrossRef]
- Rajaram, K.; Nelson, D.E. Chlamydia muridarum infection of macrophages elicits bactericidal nitric oxide production via reactive oxygen species and cathepsin B. Infect. Immun. 2015, 83, 3164–3175. [Google Scholar] [CrossRef]
- Lin, K.; Baritaki, S.; Vivarelli, S.; Falzone, L.; Scalisi, A.; Libra, M.; Bonavida, B. The Breast Cancer Protooncogenes HER2, BRCA1 and BRCA2 and Their Regulation by the iNOS/NOS2 Axis. Antioxidants 2022, 11, 1195. [Google Scholar] [CrossRef]
- Shekhar, S.; Peng, Y.; Wang, S.; Yang, X. CD103+ lung dendritic cells (LDCs) induce stronger Th1/Th17 immunity to a bacterial lung infection than CD11b(hi) LDCs. Cell. Mol. Immunol. 2018, 15, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zha, X.; Yang, S.; Niu, W.; Tan, L.; Xu, Y.; Zeng, J.; Tang, Y.; Sun, L.; Pang, G.; Qiao, S.; et al. IL-27/IL-27R Mediates Protective Immunity against Chlamydial Infection by Suppressing Excessive Th17 Responses and Reducing Neutrophil Inflammation. J. Immunol. 2021, 206, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Yang, S.; Sun, R.; Tuo, Y.; Tan, L.; Zhang, H.; Zhang, Y.; Che, X.; Lu, T.; Zhang, X.; et al. A Pathogenic Role for FcγRI in the Immune Response against Chlamydial Respiratory Infection. Microorganisms 2023, 11, 39. [Google Scholar] [CrossRef]
- Jian, L.; Li, C.; Wang, X.; Sun, L.; Ma, Z.; Zhao, J. IL-21 impairs pro-inflammatory activity of M1-like macrophages exerting anti-inflammatory effects on rheumatoid arthritis. Autoimmunity 2022, 55, 75–85. [Google Scholar] [CrossRef]
- Xu, M.; Liu, M.; Du, X.; Li, S.; Li, H.; Li, X.; Li, Y.; Wang, Y.; Qin, Z.; Fu, Y.X.; et al. Intratumoral Delivery of IL-21 Overcomes Anti-Her2/Neu Resistance through Shifting Tumor-Associated Macrophages from M2 to M1 Phenotype. J. Immunol. 2015, 194, 4997–5006. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Baulch, J.E.; Madan, S.; Salah, S.; Cheeks, S.N.; Krattli, R.P.; Subramanian, V.S.; Acharya, M.M.; Agrawal, A. Impact of IL-21-associated peripheral and brain crosstalk on the Alzheimer’s disease neuropathology. Cell. Mol. Life Sci. 2022, 79, 331. [Google Scholar] [CrossRef] [PubMed]
- Van Belle, T.L.; Nierkens, S.; Arens, R.; von Herrath, M.G. Interleukin-21 receptor-mediated signals control autoreactive T cell infiltration in pancreatic islets. Immunity 2012, 36, 1060–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, R.; Fina, D.; Peluso, I.; Stolfi, C.; Fantini, M.C.; Gioia, V.; Caprioli, F.; Blanco, G.D.V.; Paoluzi, O.A.; MacDonald, T.T.; et al. A functional role for interleukin-21 in promoting the synthesis of the T-cell chemoattractant, MIP-3alpha, by gut epithelial cells. Gastroenterology 2007, 132, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Yang, S.; Tuo, Y.; Zha, X.; Sun, R.; Lu, T.; Zhang, H.; Tan, L.; Qiao, S.; Bai, H. IL-27 Signaling Promotes Th1 Response by Downregulating IL-10 Production in DCs during Chlamydial Respiratory Infection. Microorganisms 2023, 11, 604. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) |
|---|---|---|
| β-Actin | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT |
| TNFα | CTGAACTTCGGGGTGATCGG | GGCTTGTCACTCGAATTTTGAGA |
| IL-12p40 | TGGTTTGCCATCGTTTTGCTG | ACAGGTGAGGTTCACTGTTTCT |
| TGFβ | AAAACAGGGGCAGTTACTACAAC | TGGCAGATATAGACCATCAGCA |
| IL-10 | CTTACTGACTGGCATGAGGATCA | GCAGCTCTAGGAGCATGTGG |
| iNOS | GTTCTCAGCCCAACAATACAAGA | GTGGACGGGTCGATGTCAC |
| ARG-1 | TTGGGTGGATGCTCACACTG | GTACACGATGTCTTTGGCAGA |
| IL-1β | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG |
| IL-6 | TGAACAACGATGATGCACTTGCAG | TAGCCACTCCTTCTGTGACTCCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Zeng, J.; Hao, W.; Sun, R.; Tuo, Y.; Tan, L.; Zhang, H.; Liu, R.; Bai, H. IL-21/IL-21R Promotes the Pro-Inflammatory Effects of Macrophages during C. muridarum Respiratory Infection. Int. J. Mol. Sci. 2023, 24, 12557. https://doi.org/10.3390/ijms241612557
Yang S, Zeng J, Hao W, Sun R, Tuo Y, Tan L, Zhang H, Liu R, Bai H. IL-21/IL-21R Promotes the Pro-Inflammatory Effects of Macrophages during C. muridarum Respiratory Infection. International Journal of Molecular Sciences. 2023; 24(16):12557. https://doi.org/10.3390/ijms241612557
Chicago/Turabian StyleYang, Shuaini, Jiajia Zeng, Wenlian Hao, Ruoyuan Sun, Yuqing Tuo, Lu Tan, Hong Zhang, Ran Liu, and Hong Bai. 2023. "IL-21/IL-21R Promotes the Pro-Inflammatory Effects of Macrophages during C. muridarum Respiratory Infection" International Journal of Molecular Sciences 24, no. 16: 12557. https://doi.org/10.3390/ijms241612557
APA StyleYang, S., Zeng, J., Hao, W., Sun, R., Tuo, Y., Tan, L., Zhang, H., Liu, R., & Bai, H. (2023). IL-21/IL-21R Promotes the Pro-Inflammatory Effects of Macrophages during C. muridarum Respiratory Infection. International Journal of Molecular Sciences, 24(16), 12557. https://doi.org/10.3390/ijms241612557