
APE-1/Ref-1 Inhibition Blocks Malignant Pleural Mesothelioma Cell Proliferation and Migration: Crosstalk between Oxidative Stress and Epithelial Mesenchymal Transition (EMT) in Driving Carcinogenesis and Metastasis


 and
and 
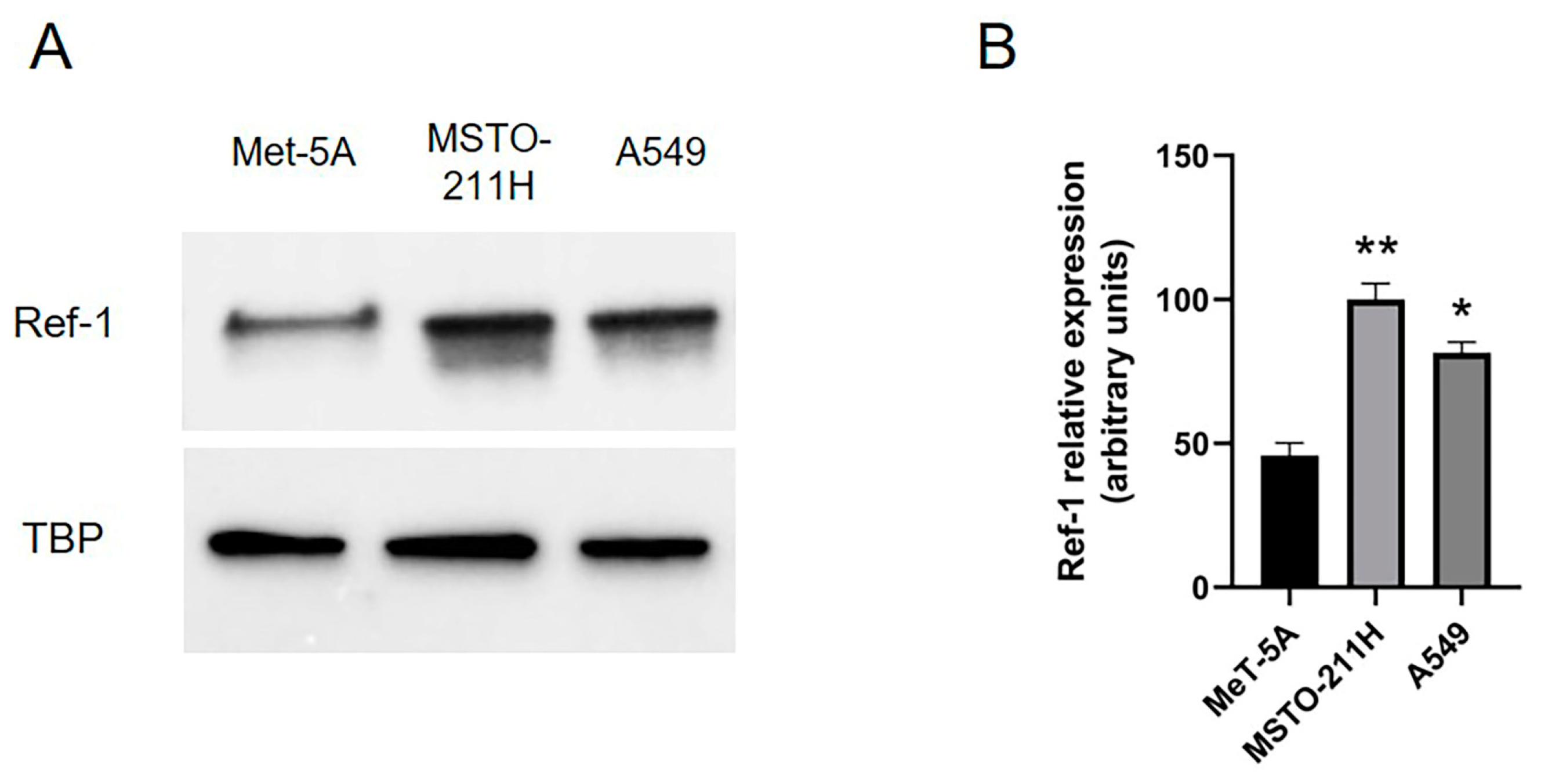{kind=link}
{kind=link}
{kind=link}
{kind=link}
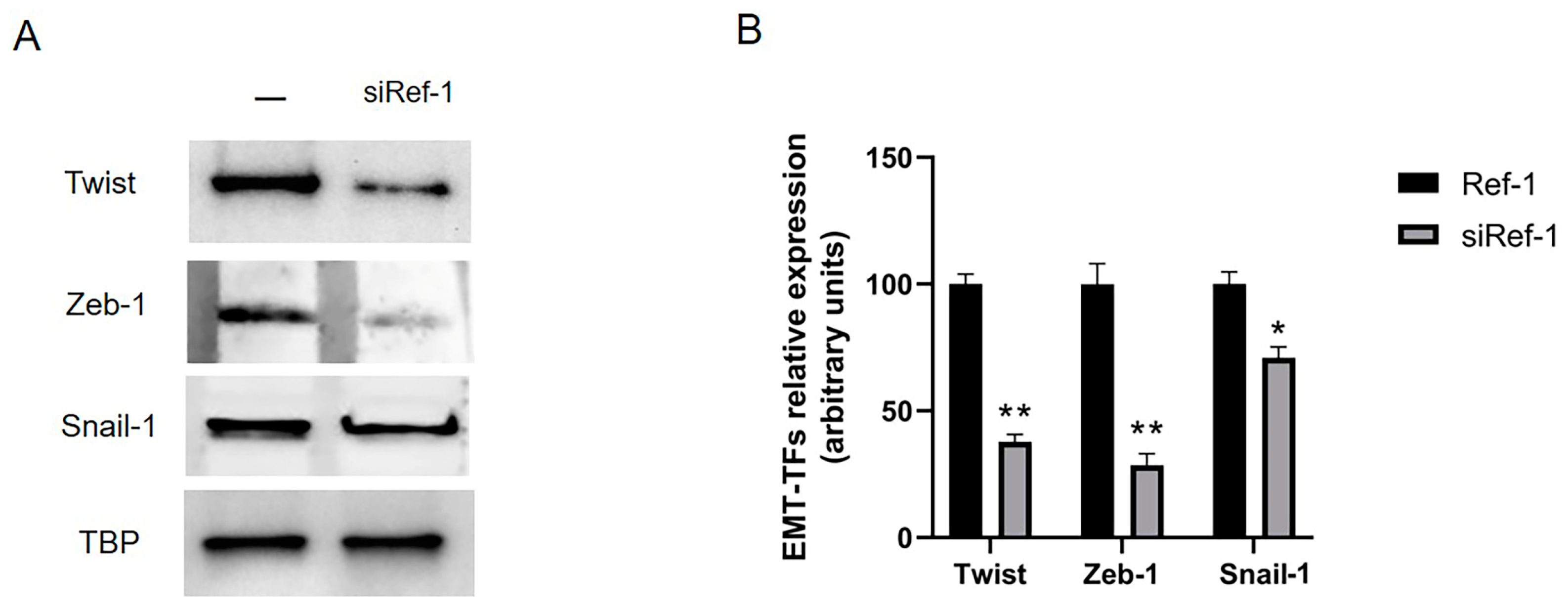{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Overexpressed Ref-1 in MPM Cells Is Downregulated after Co-Incubation with E3330 Specific Ref-1 Inhibitor
2.2. Overexpressed Ref-1 in MPM and A549 Cells Is Downregulated after siRNA Transfection
2.3. Overexpressed EMT-TFs in MPM Are Downregulated after Co-Incubation with Both E3330-Specific Ref-1 Inhibitor and siRNA Ref-1
2.4. Effect of Ref-1 Inhibition/Knockdown on Cellular Proliferation
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. E3330 Ref-1 Inhibitor
4.3. siRNA Ref-1
4.4. Nuclear Protein Extraction
4.5. Western Blot Analysis
4.6. Migration Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicolini, F.; Bocchini, M.; Bronte, G.; Delmonte, A.; Guidoboni, M.; Crinò, L.; Mazza, M. Malignant Pleural Mesothelioma: State-of-the-Art on Current Therapies and Promises for the Future. Front. Oncol. 2020, 9, 1519. [Google Scholar] [CrossRef]
- Brcic, L.; Kern, I. Clinical Significance of Histologic Subtyping of Malignant Pleural Mesothelioma. Transl. Lung Cancer Res. 2020, 9, 924–933. [Google Scholar] [CrossRef]
- Bibby, A.C.; Tsim, S.; Kanellakis, N.; Ball, H.; Talbot, D.C.; Blyth, K.G.; Maskell, N.A.; Psallidas, I. Malignant Pleural Mesothelioma: An Update on Investigation, Diagnosis and Treatment. Eur. Respir. Rev. 2016, 25, 472–486. [Google Scholar] [CrossRef]
- Kamp, D.W. Asbestos-Induced Lung Diseases: An Update. Transl. Res. 2009, 153, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Ramundo, V.; Zanirato, G.; Aldieri, E. The Epithelial-to-Mesenchymal Transition (EMT) in the Development and Metastasis of Malignant Pleural Mesothelioma. IJMS 2021, 22, 12216. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.H.; Toyokuni, S. Malignant Mesothelioma as an Oxidative Stress-Induced Cancer: An Update. Free Radic. Biol. Med. 2015, 86, 166–178. [Google Scholar] [CrossRef]
- Chaffer, C.L.; San Juan, B.P.; Lim, E.; Weinberg, R.A. EMT, Cell Plasticity and Metastasis. Cancer Metastasis Rev. 2016, 35, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial–Mesenchymal Transition and Implications for Cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Cannito, S.; Novo, E.; di Bonzo, L.V.; Busletta, C.; Colombatto, S.; Parola, M. Epithelial–Mesenchymal Transition: From Molecular Mechanisms, Redox Regulation to Implications in Human Health and Disease. Antioxid. Redox Signal. 2010, 12, 1383–1430. [Google Scholar] [CrossRef]
- Xiao, D.; He, J. Epithelial mesenchymal transition and lung cancer. J. Thorac. Dis. 2010, 2, 154–159. [Google Scholar] [CrossRef]
- Legras, A.; Pécuchet, N.; Imbeaud, S.; Pallier, K.; Didelot, A.; Roussel, H.; Gibault, L.; Fabre, E.; Le Pimpec-Barthes, F.; Laurent-Puig, P.; et al. Epithelial-to-Mesenchymal Transition and MicroRNAs in Lung Cancer. Cancers 2017, 9, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, M.Q.; Ward, C.; Muller, H.K.; Sohal, S.S.; Walters, E.H. Epithelial Mesenchymal Transition (EMT) and Non-Small Cell Lung Cancer (NSCLC): A Mutual Association with Airway Disease. Med. Oncol. 2017, 34, 45. [Google Scholar] [CrossRef] [Green Version]
- Schramm, A.; Opitz, I.; Thies, S.; Seifert, B.; Moch, H.; Weder, W.; Soltermann, A. Prognostic Significance of Epithelial–Mesenchymal Transition in Malignant Pleural Mesothelioma. Eur. J. Cardio-Thorac. Surg. 2010, 37, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Amjad, S.; Yun, H.; Mani, S.; de Perrot, M. A Panel of Emerging EMT Genes Identified in Malignant Mesothelioma. Sci. Rep. 2022, 12, 1007. [Google Scholar] [CrossRef] [PubMed]
- Turini, S.; Bergandi, L.; Gazzano, E.; Prato, M.; Aldieri, E. Epithelial to Mesenchymal Transition in Human Mesothelial Cells Exposed to Asbestos Fibers: Role of TGF-β as Mediator of Malignant Mesothelioma Development or Metastasis via EMT Event. IJMS 2019, 20, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwanami, T.; Uramoto, H.; Nakagawa, M.; Shimokawa, H.; Yamada, S.; Kohno, K.; Tanaka, F. Clinical Significance of Epithelial-Mesenchymal Transition-Associated Markers in Malignant Pleural Mesothelioma. Oncology 2014, 86, 109–116. [Google Scholar] [CrossRef]
- Liu, R.-Y.; Zeng, Y.; Lei, Z.; Wang, L.; Yang, H.; Liu, Z.; Zhao, J.; Zhang, H.-T. JAK/STAT3 Signaling Is Required for TGF-β-Induced Epithelial-Mesenchymal Transition in Lung Cancer Cells. Int. J. Oncol. 2014, 44, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Bao, J.; Hao, J.; Peng, Y.; Hong, F. HSP70 Inhibits High Glucose-Induced Smad3 Activation and Attenuates Epithelial-to-Mesenchymal Transition of Peritoneal Mesothelial Cells. Mol. Med. Rep. 2014, 10, 1089–1095. [Google Scholar] [CrossRef] [Green Version]
- Hydrogen Peroxide Promotes Epithelial to Mesenchymal Transition and Stemness in Human Malignant Mesothelioma Cells. Asian Pac. J. Cancer Prev. 2013, 14, 3625–3630. [CrossRef] [Green Version]
- Gorowiec, M.R.; Borthwick, L.A.; Parker, S.M.; Kirby, J.A.; Saretzki, G.C.; Fisher, A.J. Free Radical Generation Induces Epithelial-to-Mesenchymal Transition in Lung Epithelium via a TGF-Β1-Dependent Mechanism. Free Radic. Biol. Med. 2012, 52, 1024–1032. [Google Scholar] [CrossRef]
- Schiavello, M.; Gazzano, E.; Bergandi, L.; Silvagno, F.; Libener, R.; Riganti, C.; Aldieri, E. Identification of Redox-Sensitive Transcription Factors as Markers of Malignant Pleural Mesothelioma. Cancers 2021, 13, 1138. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xiang, D.-B.; Yang, X.; Chen, L.-S.; Li, M.-X.; Zhong, Z.-Y.; Zhang, Y.-S. APE1 Overexpression Is Associated with Cisplatin Resistance in Non-Small Cell Lung Cancer and Targeted Inhibition of APE1 Enhances the Activity of Cisplatin in A549 Cells. Lung Cancer 2009, 66, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Caston, R.A.; Gampala, S.; Armstrong, L.; Messmann, R.A.; Fishel, M.L.; Kelley, M.R. The Multifunctional APE1 DNA Repair–Redox Signaling Protein as a Drug Target in Human Disease. Drug Discov. Today 2021, 26, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.; Logsdon, D.; Messmann, R.A.; Fehrenbacher, J.C.; Fishel, M.L.; Kelley, M.R. Exploiting the Ref-1-APE1 Node in Cancer Signaling and Other Diseases: From Bench to Clinic. NPJ Precis. Oncol. 2017, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Luo, M.; Kelley, M.R. Human Apurinic Endonuclease 1 (APE1) Expression and Prognostic Significance in Osteosarcoma: Enhanced Sensitivity of Osteosarcoma to DNA Damaging Agents Using Silencing RNA APE1 Expression Inhibition. Mol. Cancer Ther. 2004, 3, 679–686. [Google Scholar] [CrossRef]
- Xiang, D.-B.; Chen, Z.-T.; Wang, D.; Li, M.-X.; Xie, J.-Y.; Zhang, Y.-S.; Qing, Y.; Li, Z.-P.; Xie, J. Chimeric Adenoviral Vector Ad5/F35-Mediated APE1 SiRNA Enhances Sensitivity of Human Colorectal Cancer Cells to Radiotherapy in vitro and in vivo. Cancer Gene Ther. 2008, 15, 625–635. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Chen, X.-H.; Wang, D. Experimental Study Enhancing the Chemosensitivity of Multiple Myeloma to Melphalan by Using a Tissue-Specific APE1-Silencing RNA Expression Vector. Clin. Lymphoma Myeloma 2007, 7, 296–304. [Google Scholar] [CrossRef]
- Fassina, A.; Cappellesso, R.; Guzzardo, V.; Dalla Via, L.; Piccolo, S.; Ventura, L.; Fassan, M. Epithelial–Mesenchymal Transition in Malignant Mesothelioma. Mod. Pathol. 2012, 25, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Merikallio, H.; Pääkkö, P.; Salmenkivi, K.; Kinnula, V.; Harju, T.; Soini, Y. Expression of Snail, Twist, and Zeb1 in Malignant Mesothelioma. APMIS 2013, 121, 1–10. [Google Scholar] [CrossRef]
- Mehta, A.; Dobersch, S.; Romero-Olmedo, A.J.; Barreto, G. Epigenetics in Lung Cancer Diagnosis and Therapy. Cancer Metastasis Rev. 2015, 34, 229–241. [Google Scholar] [CrossRef]
- Cheung, E.C.; Vousden, K.H. The Role of ROS in Tumour Development and Progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Li, Y.; Kong, J.; Li, C. Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Could Serve as a Potential Serological Biomarker for the Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J. Oral. Maxillofac. Surg. 2019, 77, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Gampala, S.; Shah, F.; Lu, X.; Moon, H.; Babb, O.; Umesh Ganesh, N.; Sandusky, G.; Hulsey, E.; Armstrong, L.; Mosely, A.L.; et al. Ref-1 Redox Activity Alters Cancer Cell Metabolism in Pancreatic Cancer: Exploiting This Novel Finding as a Potential Target. J. Exp. Clin. Cancer Res. 2021, 40, 251. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhen, P.; Niu, X.; Dai, Y.; Wang, Y.; Zhou, M. APE1 Promotes Proliferation and Migration of Cutaneous Squamous Cell Carcinoma. J. Dermatol. Sci. 2020, 100, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Peng, Y.; Jiang, X.; Lu, X.; Duan, W.; Zhang, S.; Dai, N.; Shan, J.; Feng, Y.; Li, X.; et al. The Regulatory Role of APE1 in Epithelial-to-Mesenchymal Transition and in Determining EGFR-TKI Responsiveness in Non-Small-Cell Lung Cancer. Cancer Med. 2018, 7, 4406–4419. [Google Scholar] [CrossRef]
- Menju, T.; Date, H. Lung Cancer and Epithelial-Mesenchymal Transition. Gen. Thorac. Cardiovasc. Surg. 2021, 69, 781–789. [Google Scholar] [CrossRef]
- Fishel, M.L.; Xia, H.; McGeown, J.; McIlwain, D.W.; Elbanna, M.; Craft, A.A.; Kaimakliotis, H.Z.; Sandusky, G.E.; Zhang, C.; Pili, R.; et al. Antitumor Activity and Mechanistic Characterization of APE1/Ref-1 Inhibitors in Bladder Cancer. Mol. Cancer Ther. 2019, 18, 1947–1960. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramundo, V.; Zanirato, G.; Palazzo, M.L.; Riganti, C.; Aldieri, E. APE-1/Ref-1 Inhibition Blocks Malignant Pleural Mesothelioma Cell Proliferation and Migration: Crosstalk between Oxidative Stress and Epithelial Mesenchymal Transition (EMT) in Driving Carcinogenesis and Metastasis. Int. J. Mol. Sci. 2023, 24, 12570. https://doi.org/10.3390/ijms241612570
Ramundo V, Zanirato G, Palazzo ML, Riganti C, Aldieri E. APE-1/Ref-1 Inhibition Blocks Malignant Pleural Mesothelioma Cell Proliferation and Migration: Crosstalk between Oxidative Stress and Epithelial Mesenchymal Transition (EMT) in Driving Carcinogenesis and Metastasis. International Journal of Molecular Sciences. 2023; 24(16):12570. https://doi.org/10.3390/ijms241612570
Chicago/Turabian StyleRamundo, Valeria, Giada Zanirato, Maria Luisa Palazzo, Chiara Riganti, and Elisabetta Aldieri. 2023. "APE-1/Ref-1 Inhibition Blocks Malignant Pleural Mesothelioma Cell Proliferation and Migration: Crosstalk between Oxidative Stress and Epithelial Mesenchymal Transition (EMT) in Driving Carcinogenesis and Metastasis" International Journal of Molecular Sciences 24, no. 16: 12570. https://doi.org/10.3390/ijms241612570
APA StyleRamundo, V., Zanirato, G., Palazzo, M. L., Riganti, C., & Aldieri, E. (2023). APE-1/Ref-1 Inhibition Blocks Malignant Pleural Mesothelioma Cell Proliferation and Migration: Crosstalk between Oxidative Stress and Epithelial Mesenchymal Transition (EMT) in Driving Carcinogenesis and Metastasis. International Journal of Molecular Sciences, 24(16), 12570. https://doi.org/10.3390/ijms241612570