

Genome Identification and Evolutionary Analysis of LBD Genes and Response to Environmental Factors in Phoebe bournei

 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Identification of LBD Family Members and Their Physicochemical Properties
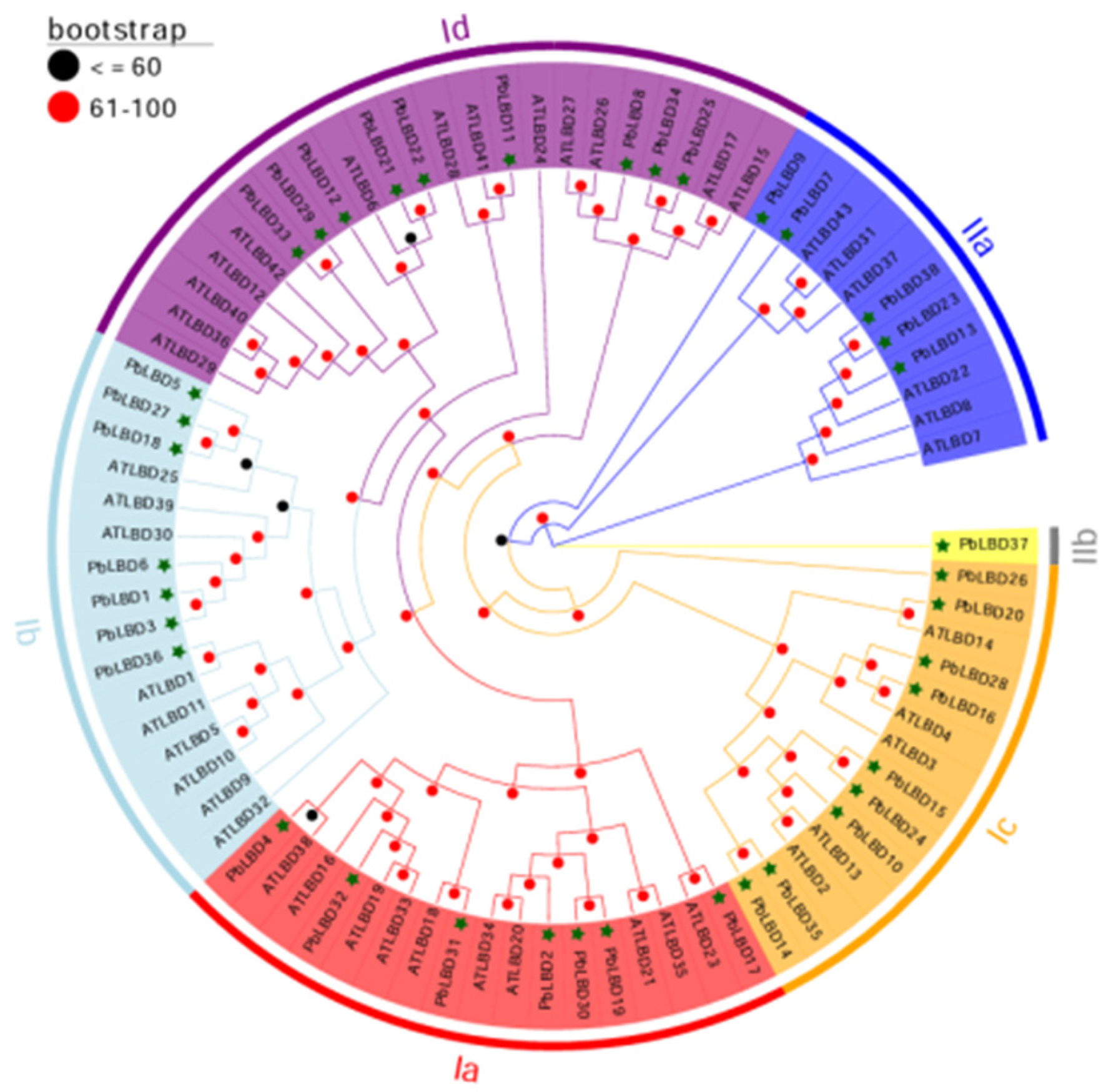
2.2. Phylogenetic Analysis of the LBD Family Members in P. bournei
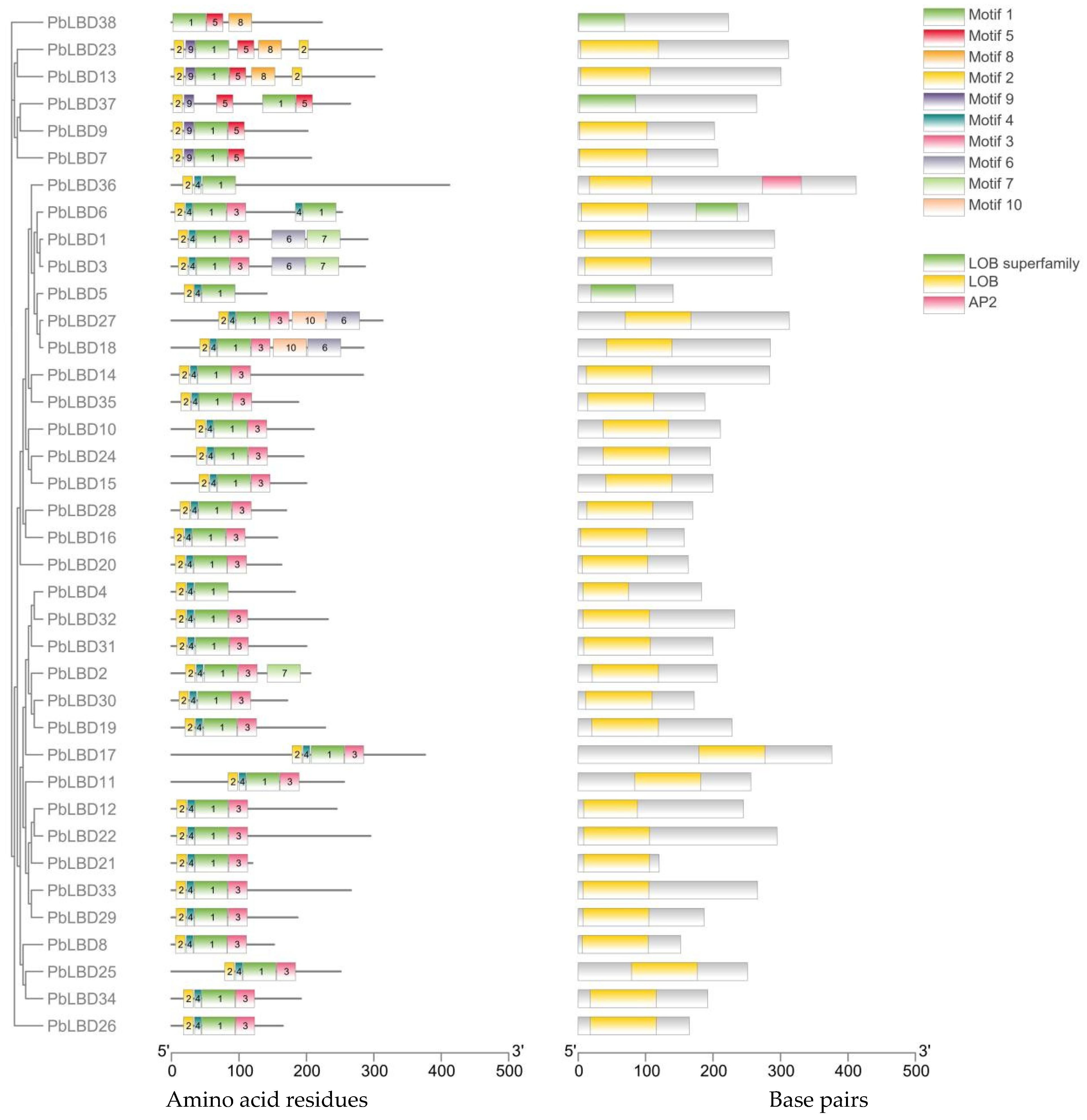
2.3. Analysis of Conserved Motifs and Protein Structure of PbLBD
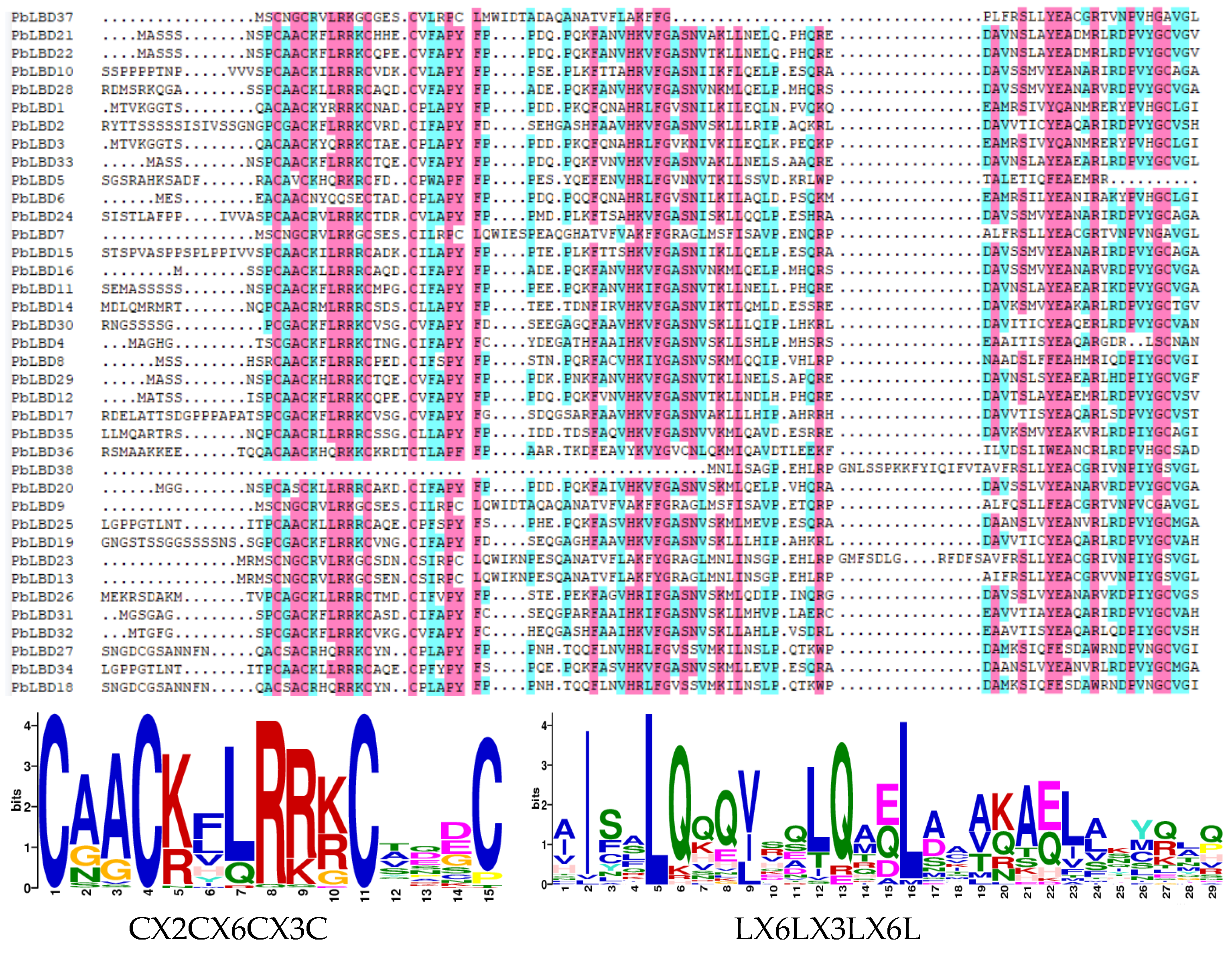
2.4. LBD Protein Sequence Comparison and Their Tertiary Structure
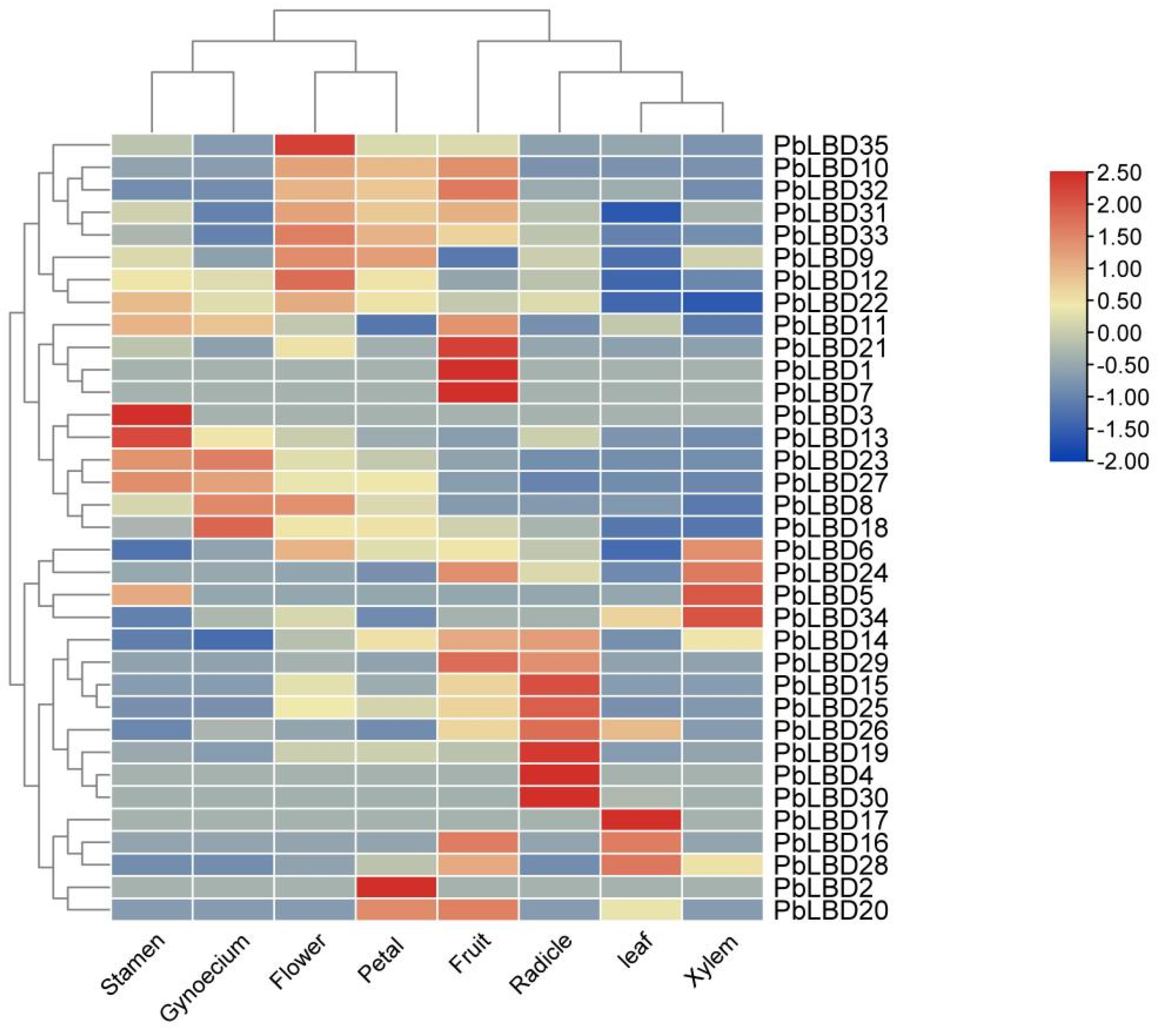
2.5. The Variance of LBD Gene Expression Level in Different Tissues in P. bournei
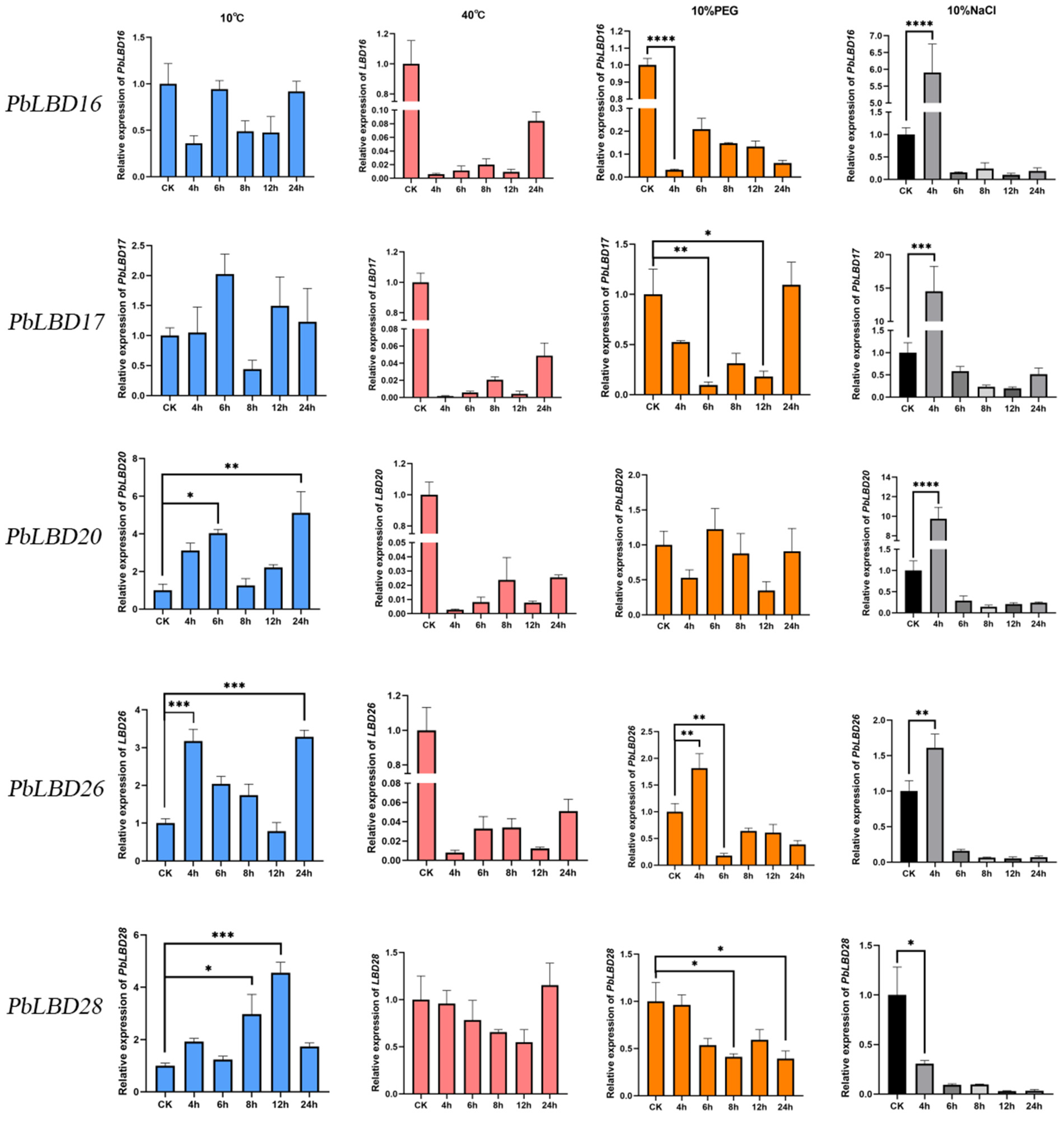
2.6. The Induced Expression of PbLBD Genes Associated with Stress Treatment
3. Discussion
3.1. The Role of the Transcription Factor for Lateral Organ Boundary Domains in Phoebe bournei and Its Significance for Bioinformatics Research
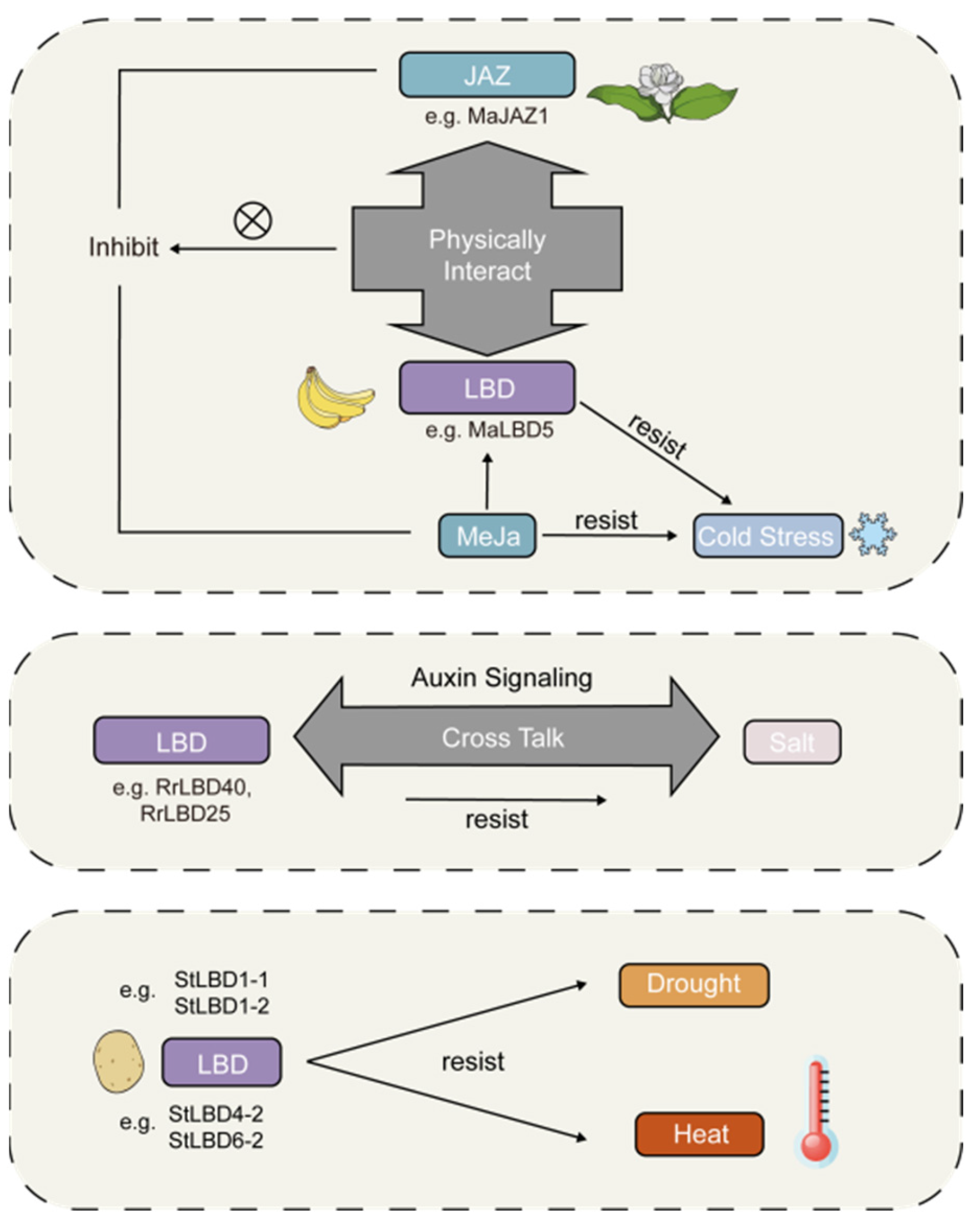
3.2. The Effects of Environmental Factors on LBD Genes in P. bournei
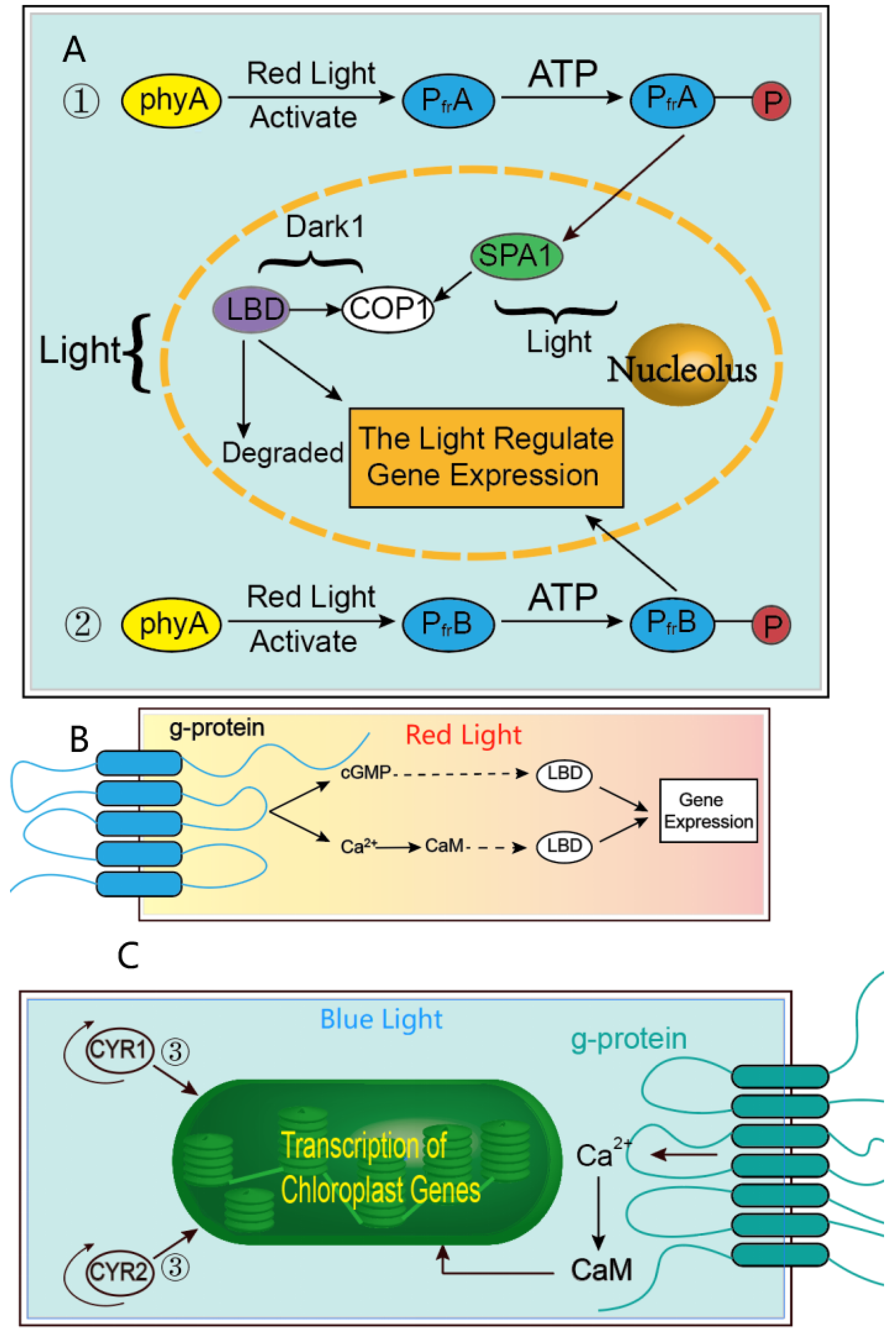
3.2.1. The Consequences and Mechanism of How the Transcription Factors Respond to Varying Light Quality
3.2.2. The Response of Genes under Cold, Heat, Salt, and Drought Stress
4. Materials and Methods
4.1. Materials
4.1.1. Plant Materials and Treatment
4.1.2. Test Setup, Parameter Settings, and Processing Instructions
4.2. Methods
4.2.1. Identification of the LBD Gene Family in P. bournei
4.2.2. Determination of Physicochemical Properties of PbLBD Genes and Protein Prediction
4.2.3. Phylogenetic Analysis of the LBD Gene Family in P. bournei
4.2.4. Analysis of the Conserved Motifs, Gene Structures, and Characteristic Domains of the LBD Gene Family in P. bournei
4.2.5. Expression Patterns of PbLBD Gene
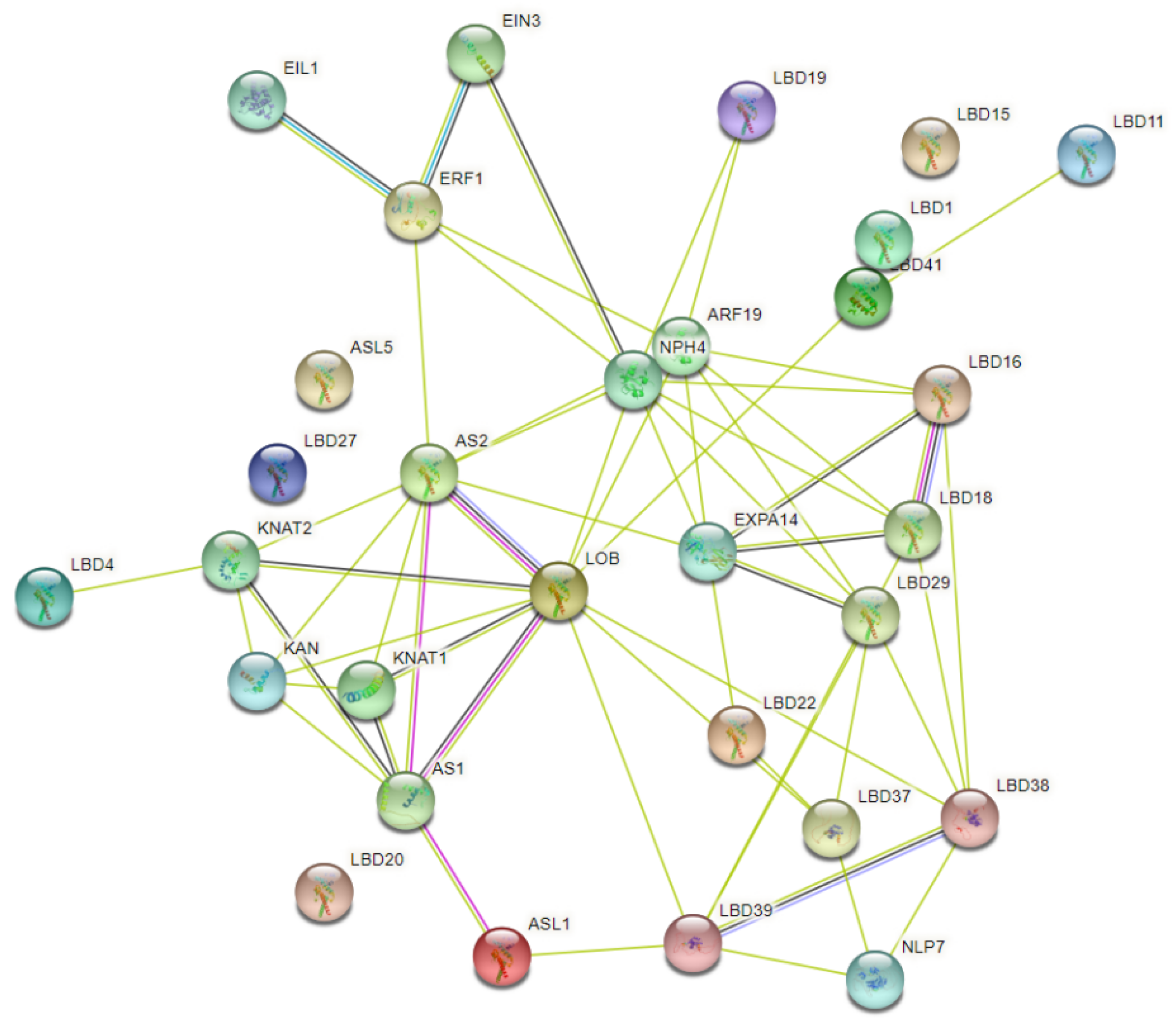
4.2.6. Protein Interaction Network Analysis of PbLBD Proteins
4.2.7. Plant Material and Stress Treatment with RNA Extraction and qRT–PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Danmei, W.; Jiping, L.; Tao, T. Determining the optimal density of Phoebe bournei plantations based on dynamic programming under close-to-nature management measures. Sustainability 2022, 14, 847. [Google Scholar]
- Yang, Z.J.; Wu, X.H.; Grossnickle, S.C.; Chen, L.H.; Yu, X.X.; El-Kassaby, Y.A.; Feng, J.L. Formula Fertilization Promotes Phoebe bournei Robust Seedling Cultivation. Forests 2020, 11, 781. [Google Scholar] [CrossRef]
- Chen, Z.; Cao, J.; Yu, H.; Shang, H. Effects of elevated ozone levels on photosynthesis, biomass and non-structural carbohydrates of Phoebe bournei and Phoebe zhennan in subtropical China. Front. Plant Sci. 2018, 9, 1764. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef]
- Iwakawa, H.; Takahashi, H.; Machida, Y.; Machida, C. Roles of ASYMMETRIC LEAVES2 (AS2) and Nucleolar Proteins in the Adaxial-Abaxial Polarity Specification at the Perinucleolar Region in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 7314. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Gan, S. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant J. 2006, 46, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Husbands, A.; Bell, E.M.; Shuai, B.; Smith, H.M.; Springer, P.S. LATERAL ORGAN BOUNDARIES defines a new family of DNA-binding transcription factors and can interact with specific bHLH proteins. Nucleic Acids Res. 2007, 35, 6663–6671. [Google Scholar] [CrossRef]
- Cubas, P.; Lauter, N.; Doebley, J.; Coen, E. The TCP domain: A motif found in proteins regulating plant growth and development. Plant J. 2002, 18, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, J.; Xu, Q.; Dong, J.; Song, L.; Wang, W.; Shen, F. Long noncoding RNA lncRNA354 functions as a competing endogenous RNA of miR160b to regulate ARF genes in response to salt stress in upland cotton. Plant Cell Environ. 2021, 44, 3302–3321. [Google Scholar] [CrossRef]
- Jia, R.; Li, C.; Wang, Y.; Qin, X.; Meng, L.; Sun, X. Genome-Wide Analysis of LBD Transcription Factor Genes in Dendrobium catenatum. Int. J. Mol. Sci. 2022, 23, 2089. [Google Scholar] [CrossRef]
- Majer, C.; Hochholdinger, F. Defining the boundaries: Structure and function of LOB domain proteins. Trends Plant Sci. 2011, 16, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wang, K.; Du, L.; Song, Y.; Li, H.; Ye, X. Genome-Wide Identification and Expression Profiling Analysis of WOX Family Protein-Encoded Genes in Triticeae Species. Int. J. Mol. Sci. 2021, 22, 9325. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Wu, B.; Zhang, J.; Ma, S.; Weng, H.; Wang, G.; Jia, C. Genome-wide analysis of LBD transcription factors reveals PvLBD12 expression profile involved in salt stress in switchgrass (Panicum virgatum L.). Res. Sq. 2022. [Google Scholar] [CrossRef]
- Xie, T.; Zeng, L.; Chen, X.; Rong, H.; Wu, J.; Batley, J.; Jiang, J.; Wang, Y. Genome-wide analysis of the lateral organ boundaries domain gene family in Brassica napus. Genes 2020, 11, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.Q.; Li, T.; Wang, X.; Sun, W.J.; Zhang, T.T.; You, C.X.; Wang, X.F. Identification and characterization of apple MdNLP7 transcription factor in the nitrate response. Plant Sci. 2022, 316, 111158. [Google Scholar] [CrossRef]
- Liu, H.; Cao, M.; Chen, X.; Ye, M.; Zhao, P.; Nan, Y.; Li, W.; Zhang, C.; Kong, L.; Kong, N.; et al. Genome-wide analysis of the lateral organ boundaries domain (LBD) gene family in Solanum tuberosum. Int. J. Mol. Sci. 2019, 20, 5360. [Google Scholar] [CrossRef] [Green Version]
- Gombos, M.; Zombori, Z.; Szécsényi, M.; Sándor, G.; Kovács, H.; Györgyey, J. Characterization of the LBD gene family in Brachypodium: A phylogenetic and transcriptional study. Plant Cell Rep. 2017, 36, 61–79. [Google Scholar] [CrossRef]
- Ba, L.J.; Kuang, J.F.; Chen, J.Y.; Lu, W.J. MaJAZ1 attenuates the MaLBD5-mediated transcriptional activation of jasmonate biosynthesis gene MaAOC2 in regulating cold tolerance of banana fruit. J. Agric. Food Chem. 2016, 64, 738–745. [Google Scholar] [CrossRef]
- Lu, Q.; Shao, F.; Macmillan, C.; Wilson, I.W.; van der Merwe, K.; Hussey, S.G.; Myburg, A.A.; Dong, X.; Qiu, D. Genomewide analysis of the lateral organ boundaries domain gene family in Eucalyptus grandis reveals members that differentially impact secondary growth. Plant Biotechnol. J. 2017, 16, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Hou, Z.; Liao, J.; Qin, Y.; Wang, L.; Wang, X.; Su, W.; Cai, Z.; Fang, Y.; Aslam, M.; et al. Genome-wide identification and expression analysis of LBD transcription factor genes in passion fruit (Passiflora edulis). Int. J. Mol. Sci. 2022, 23, 4700. [Google Scholar] [CrossRef]
- Lee, H.W.; Cho, C.; Pandey, S.K.; Park, Y.; Kim, M.J.; Kim, J. LBD16 and LBD18 acting downstream of ARF7 and ARF19 are involved in adventitious root formation in Arabidopsis. BMC Plant Biol. 2019, 19, 46. [Google Scholar] [CrossRef] [Green Version]
- Xiong, H.; Shi, A.; Wu, D.; Weng, Y.; Qin, J.; Ravelombola, W.S.; Shu, X.; Zhou, W. Genome-Wide identification, classification and evolutionary expansion of KNOX gene family in rice (Oryza sativa) and Populus (Populustrichocarpa). Am. J. Plant Sci. 2018, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wang, D.; Zhang, C.; Ye, M.; Kong, N.; Ma, H.; Chen, Q. Comprehensive analysis and expression profiling of PIN, AUX/LAX, and ABCB auxin transporter gene families in Solanum tuberosum under Phytohormone stimuli and abiotic stresses. Biology 2021, 10, 127. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Liu, C.Y.; Liu, C.X.; Zhao, Y.L.; Xu, R.R. Genomewide analysis of the lateral organ boundaries domain gene family in Vitis vinifera. J. Genet. 2016, 95, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Zhang, S.Z.; Zheng, C.C. Genomewide analysis of LATERAL ORGAN BOUNDARIES domain gene family in Zea mays. J. Genet. 2014, 93, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, W.; Cheng, Y.; Feng, L. Genome-wide identification of LATERAL ORGAN BOUNDARIES DOMAIN (LBD) transcription factors and screening of salt stress candidates of Rosa rugosa thunb. Biology 2021, 10, 992. [Google Scholar] [CrossRef]
- Shuai, B.; Reynaga-Peña, C.G.; Springer, P.S. The lateral organ boundaries gene defines a novel, plant-specific gene family. Plant Physiol. 2002, 129, 747–761. [Google Scholar] [CrossRef] [Green Version]
- Willige, B.C.; Zander, M.; Yoo, C.Y.; Phan, A.; Garza, R.M.; Trigg, S.A.; Yupeng, H.; Joseph, R.N.; Huaming, C.; Meng, C.; et al. PHYTOCHROME-INTERACTING FACTORs trigger environmentally responsive chromatin dynamics in plants. Nat. Genet. 2021, 53, 955–961. [Google Scholar] [CrossRef]
- Sharma, R. Phytochrome: A serine kinase illuminates the nucleus! Curr. Sci. 2001, 80, 178–188. [Google Scholar]
- Liu, Y.; Ye, Y.; Wang, Y.; Jiang, L.; Yue, M.; Tang, L.; Jin, M.; Zhang, Y.; Lin, Y.; Tang, H. B-Box transcription factor FaBBX22 promotes light-induced anthocyanin accumulation in Strawberry (Fragaria × ananassa). Int. J. Mol. Sci. 2022, 23, 7757. [Google Scholar] [CrossRef]
- Goh, T.; Joi, S.; Mimura, T.; Fukaki, H. The establishment of asymmetry in Arabidopsis lateral root founder cells is regulated by LBD16/ASL18 and related LBD/ASL proteins. Development 2012, 139, 883–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhang, J.; Xu, J.; Li, Y.; Guo, L.; Wang, Z.; Zhang, X.; Zhao, B.; Guo, Y.D.; Zhang, N. CRISPR/Cas9 targeted mutagenesis of SlLBD40, a Lateral Organ Boundaries Domain transcription factor, enhances drought tolerance in tomato. Plant Sci. 2020, 301, 110683. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Yanagawa, Y.; Munesada, T.; Horii, Y.; Kuriyama, T.; Kawashima, M.; Kondou, Y.; Yoshizumi, T.; Mitsuda, N.; Ohme-Takagi, M.; et al. A collection of inducible transcription factor-GR fusion lines for functional analyses in Arabidopsis thaliana. Plant J. 2022, 111, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Gong, X.; Xie, Z.; Qi, K.; Yuan, K.; Jiao, Y.; Pan, Q.; Zhang, S.; Shiratake, K.; Tao, S. Cryptochrome-mediated blue-light signal contributes to lignin biosynthesis in stone cells in pear fruit. Plant Sci. 2022, 318, 111211. [Google Scholar] [CrossRef]
- Huang, X.; Yan, H.; Liu, Y.; Yi, Y. Genome-wide analysis of LATERAL ORGAN BOUNDARIES DOMAIN-in Physcomitrella patens and stress responses. Genes Genom. 2020, 42, 651–662. [Google Scholar] [CrossRef]
- Guo, Z.; Xu, H.; Lei, Q.; Du, J.; Li, C.; Wang, C.; Yang, Y.; Yang, Y.; Sun, X. Arabidopsis transcription factor LBD15 mediates ABA signaling and water-deficit stress tolerance via regulating ABI4 expression. Plant J. 2020, 104, 510–521. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Slater, G.S.C.; Birney, E. Automated generation of heuristics for biological sequence comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34, W435–W439. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.D.; Handsaker, R.E.; Pulit, S.L.; Nizzari, M.M.; O’Donnell, C.J.; De Bakker, P.I. SNAP: A web-based tool for identification and annotation of proxy SNPs using HapMap. Bioinformatics 2008, 24, 2938–2939. [Google Scholar] [CrossRef] [Green Version]
- Eddy, S.R. Accelerated profifile HMM searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [PubMed] [Green Version]
- Chen, S.P.; Sun, W.H.; Xiong, Y.F.; Jiang, Y.T.; Liu, X.D.; Liao, X.Y.; Zhang, D.Y.; Jiang, S.Z.; Li, Y.; Liu, B.; et al. The Phoebe genome sheds light on the evolution of magnoliids. Hortic. Res. 2020, 7, 146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, Y.; Pan, Y.; Huang, H.; Li, C.; Li, G.; Tong, Z. Transcriptomic profiling and identification of candidate genes in two Phoebe bournei ecotypes with contrasting cold stress responses. Trees 2018, 32, 1315–1333. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Amino Acid Number | Molecular Weight (Da) | Theoretical Isoelectric Point | Total Mean Hydrophilic Value | Subcellular Localization | α Helix | Extended Strand | β Turn | Random Coil | Secondary Structure Prediction |
|---|---|---|---|---|---|---|---|---|---|---|
| PbLBD1 | 291 | 32,503.12 | 5.97 | −0.592 | Mitochondrial Matrix | 39.52% | 12.37% | 3.44% | 44.67% | Random coil |
| PbLBD2 | 206 | 22,586.44 | 8.18 | −0.169 | Chloroplast Stroma | 50.00% | 5.83% | 1.94% | 42.23% | α helix |
| PbLBD3 | 287 | 32,240.86 | 5.85 | −0.666 | Cytoplasmic Matrix | 37.98% | 12.89% | 3.48% | 45.64% | Random coil |
| PbLBD4 | 183 | 19,998.55 | 5.67 | −0.17 | Cytoplasmic Matrix | 48.09% | 7.10% | 7.10% | 37.70% | α helix |
| PbLBD5 | 141 | 16,394.41 | 5.92 | −0.594 | Peroxisome | 55.32% | 4.96% | 4.26% | 35.46% | α helix |
| PbLBD6 | 253 | 28,464.98 | 7.02 | −0.22 | Chloroplast Stroma | 53.36% | 7.51% | 3.95% | 35.18% | α helix |
| PbLBD7 | 207 | 22,321.56 | 6.73 | −0.066 | Mitochondrial Matrix | 29.95% | 12.08% | 6.76% | 51.21% | Random coil |
| PbLBD8 | 152 | 17,386.88 | 8.72 | −0.369 | Chloroplast Stroma | 51.32% | 2.63% | 1.97% | 44.08% | α helix |
| PbLBD9 | 202 | 21,241.49 | 7.46 | 0.075 | Mitochondrial Matrix | 24.26% | 11.88% | 6.93% | 56.93% | Random coil |
| PbLBD10 | 211 | 23,136.05 | 5.32 | −0.261 | Chloroplast Stroma | 43.60% | 8.06% | 1.90% | 46.45% | Random coil |
| PbLBD11 | 256 | 28,282.9 | 9.49 | −0.758 | Cell Nucleus | 32.03% | 5.86% | 3.52% | 58.59% | Random coil |
| PbLBD12 | 245 | 27,093.06 | 9.33 | −0.312 | Mitochondrial Matrix | 43.27% | 8.57% | 4.08% | 44.08% | Random coil |
| PbLBD13 | 301 | 32,864.15 | 5.33 | −0.385 | Mitochondrial Matrix | 30.90% | 13.62% | 4.32% | 51.16% | Random coil |
| PbLBD14 | 284 | 32,289.83 | 4.94 | −0.187 | Mitochondrial Matrix | 55.99% | 8.10% | 3.52% | 32.39% | α helix |
| PbLBD15 | 200 | 21,392.35 | 6.81 | −0.237 | Chloroplast Stroma | 50.50% | 5.00% | 0.50% | 44.00% | α helix |
| PbLBD16 | 157 | 17,164.56 | 6.94 | −0.166 | Mitochondrial Matrix | 50.32% | 8.92% | 0.64% | 40.13% | α helix |
| PbLBD17 | 376 | 40,985.39 | 9.25 | −0.437 | Cell Nucleus | 26.60% | 17.29% | 3.72% | 52.39% | α helix |
| PbLBD18 | 285 | 33,259.55 | 8.08 | −0.68 | Endoplasmic Reticulum Membrane | 54.74% | 7.72% | 3.16% | 34.39% | α helix |
| PbLBD19 | 228 | 24,386.8 | 8.26 | −0.178 | Chloroplast Stroma | 39.47% | 2.63% | 2.19% | 55.70% | Random coil |
| PbLBD20 | 163 | 18,185.83 | 6.8 | −0.283 | Cytoplasmic Matrix | 46.01% | 6.75% | 1.23% | 46.01% | α helix/Random coil |
| PbLBD21 | 120 | 13,443.3 | 7.64 | −0.384 | Mitochondrial Matrix | 64.17% | 1.67% | 0.83% | 33.33% | α helix |
| PbLBD22 | 295 | 32,025.06 | 5.65 | −0.26 | Mitochondrial Matrix | 46.10% | 9.83% | 4.75% | 39.32% | α helix |
| PbLBD23 | 312 | 33,954.3 | 6.61 | −0.415 | Mitochondrial Matrix | 28.53% | 12.50% | 4.81% | 54.17% | Random coil |
| PbLBD24 | 196 | 21,207.35 | 8.13 | −0.104 | Chloroplast Stroma | 49.49% | 5.61% | 1.53% | 43.37% | α helix |
| PbLBD25 | 251 | 27,364 | 7.71 | −0.277 | Mitochondrial Matrix | 39.84% | 12.35% | 2.39% | 45.42% | Random coil |
| PbLBD26 | 165 | 18,082.88 | 8.58 | −0.106 | Cytoplasmic Matrix | 50.91% | 9.70% | 1.82% | 37.58% | α helix |
| PbLBD27 | 313 | 36,590.37 | 8.08 | −0.703 | Cytoplasmic Matrix | 53.04% | 8.31% | 3.19% | 35.46% | α helix |
| PbLBD28 | 170 | 18,630.24 | 8.59 | −0.265 | Cell Nucleus | 42.94% | 12.94% | 0.59% | 43.53% | Random coil |
| PbLBD29 | 187 | 20,442.35 | 8.29 | −0.369 | Mitochondrial Matrix | 46.52% | 5.88% | 2.14% | 45.45% | α helix |
| PbLBD30 | 172 | 18,557.34 | 8.51 | 0.005 | Mitochondrial Matrix | 49.42% | 3.49% | 2.33% | 44.77% | α helix |
| PbLBD31 | 200 | 21,829.1 | 7.56 | −0.152 | Cytoplasmic Matrix | 45.50% | 9.50% | 1.50% | 43.50% | α helix |
| PbLBD32 | 232 | 25,559.78 | 5.96 | −0.234 | Peroxisome | 39.66% | 4.31% | 3.88% | 52.16% | Random coil |
| PbLBD33 | 266 | 29,410.15 | 6.42 | −0.544 | Mitochondrial Matrix | 40.98% | 6.77% | 1.88% | 50.38% | Random coil |
| PbLBD34 | 192 | 20,886.83 | 8.87 | −0.231 | Cell Nucleus | 41.67% | 10.94% | 3.12% | 44.27% | Random coil |
| PbLBD35 | 188 | 21,295.17 | 6.41 | −0.314 | Mitochondrial Matrix | 58.51% | 3.19% | 0.00% | 38.30% | α helix |
| PbLBD36 | 412 | 46,401.64 | 7.15 | −0.619 | Cell Nucleus | 51.21% | 8.74% | 6.07% | 33.98% | α helix |
| PbLBD37 | 265 | 28,549.46 | 5.02 | −0.095 | Mitochondrial Matrix | 32.08% | 12.45% | 9.43% | 46.04% | Random coil |
| PbLBD38 | 223 | 24,319.44 | 7.06 | −0.371 | Cytoplasmic Matrix | 27.35% | 13.45% | 4.48% | 54.71% | Random coil |
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| PbLBD16 | CTGCATCTTTGCACCCTATTTC | CAGTTCCTGGAGCATCTTGT |
| PbLBD17 | CAACAGCAGCAACAGAACATAG | GTATCCGTAAGGTCAAAGCTAGAG |
| PbLBD20 | CAACAGGATCAGTCACCCTTAC | ACATTGCTAGAAGAGGCATAGTT |
| PbLBD27 | AGCCAAGTCTCCCAATTACAG | GGAGCTGGCCAAGATAGATAAA |
| PbLBD28 | GAGCCACAGAAGTTTGCTAATG | CACTCTCGCATTTGCTTCATAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Zhong, M.; Li, J.; Jiang, Y.; Zhou, X.; Justice Ijeoma, C.; Tang, X.; Chen, S.; Cao, S. Genome Identification and Evolutionary Analysis of LBD Genes and Response to Environmental Factors in Phoebe bournei. Int. J. Mol. Sci. 2023, 24, 12581. https://doi.org/10.3390/ijms241612581
Ma Y, Zhong M, Li J, Jiang Y, Zhou X, Justice Ijeoma C, Tang X, Chen S, Cao S. Genome Identification and Evolutionary Analysis of LBD Genes and Response to Environmental Factors in Phoebe bournei. International Journal of Molecular Sciences. 2023; 24(16):12581. https://doi.org/10.3390/ijms241612581
Chicago/Turabian StyleMa, Yiming, Minchen Zhong, Jingshu Li, Yiming Jiang, Xuerong Zhou, Chris Justice Ijeoma, Xinghao Tang, Shipin Chen, and Shijiang Cao. 2023. "Genome Identification and Evolutionary Analysis of LBD Genes and Response to Environmental Factors in Phoebe bournei" International Journal of Molecular Sciences 24, no. 16: 12581. https://doi.org/10.3390/ijms241612581
APA StyleMa, Y., Zhong, M., Li, J., Jiang, Y., Zhou, X., Justice Ijeoma, C., Tang, X., Chen, S., & Cao, S. (2023). Genome Identification and Evolutionary Analysis of LBD Genes and Response to Environmental Factors in Phoebe bournei. International Journal of Molecular Sciences, 24(16), 12581. https://doi.org/10.3390/ijms241612581