

In Silico Discovery of Potential Inhibitors Targeting the RNA Binding Loop of ADAR2 and 5-HT2CR from Traditional Chinese Natural Compounds

 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Selecting Binding Site
- IHP binding site [35,51] comprising of residues Ala389, Leu390, Asn391, Asp392, Ile397, Arg400, Arg401, Leu404, Tyr408, Gln500, Leu512, Thr513, Met514, Lys519, Arg522, Trp523, Val526, Gly527, Ile528, Gln529, Gly530, Ser531, Leu532, Leu533, Lys629, Leu632, Tyr658, His659, Lys662, Leu663, Tyr668, Gln669, Lys672, Phe676, Trp687, Val688, Glu689, Lys690, Pro691, Thr692, Gln694, and Asp695.
- RNA binding loop [35,39] comprising of residues Lys350, Val351, Gly374, Thr375, Lys376, Cys377, Ile378, Asn379, His394, Ala395, Glu396, Ile446, Thr448, Ser449, Pro450, Cys451, Gly452, Arg455, Ile456, Pro459, Lys483, Ile484, Glu485, Ser486, Gly487, Gln488, Gly489, Thr490, Leu511, Thr513, Cys516, Arg590, Lys594, and Ala595.
- A third plausible binding site lined by residues Ser458, His460, Glu461, Pro462, Ile463, Glu466, Pro467, Ala468, Asp469, Arg470, His471, His552, Asp554, and His555.
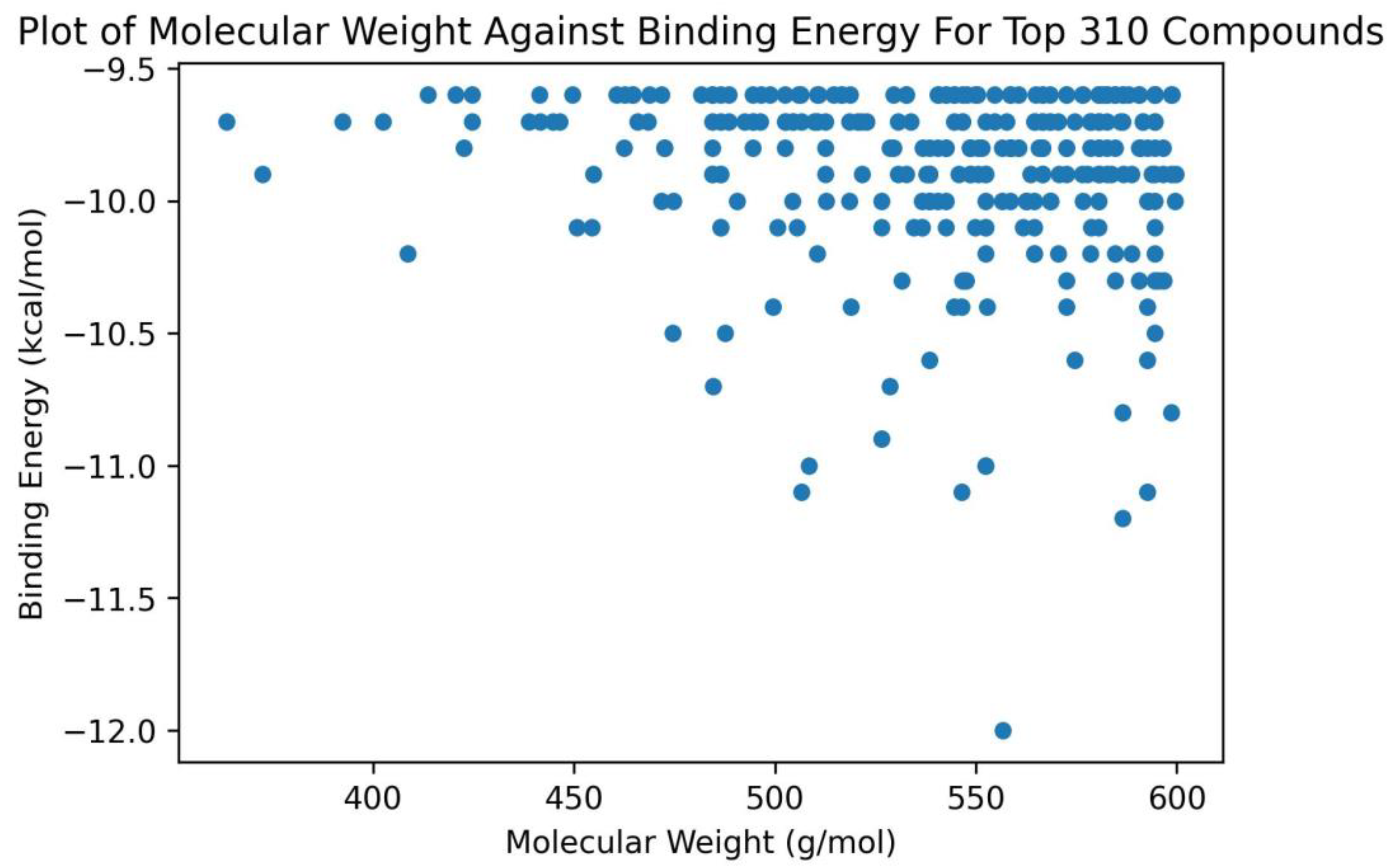
2.2. Molecular Docking of ADAR2
2.3. ADMET Prediction
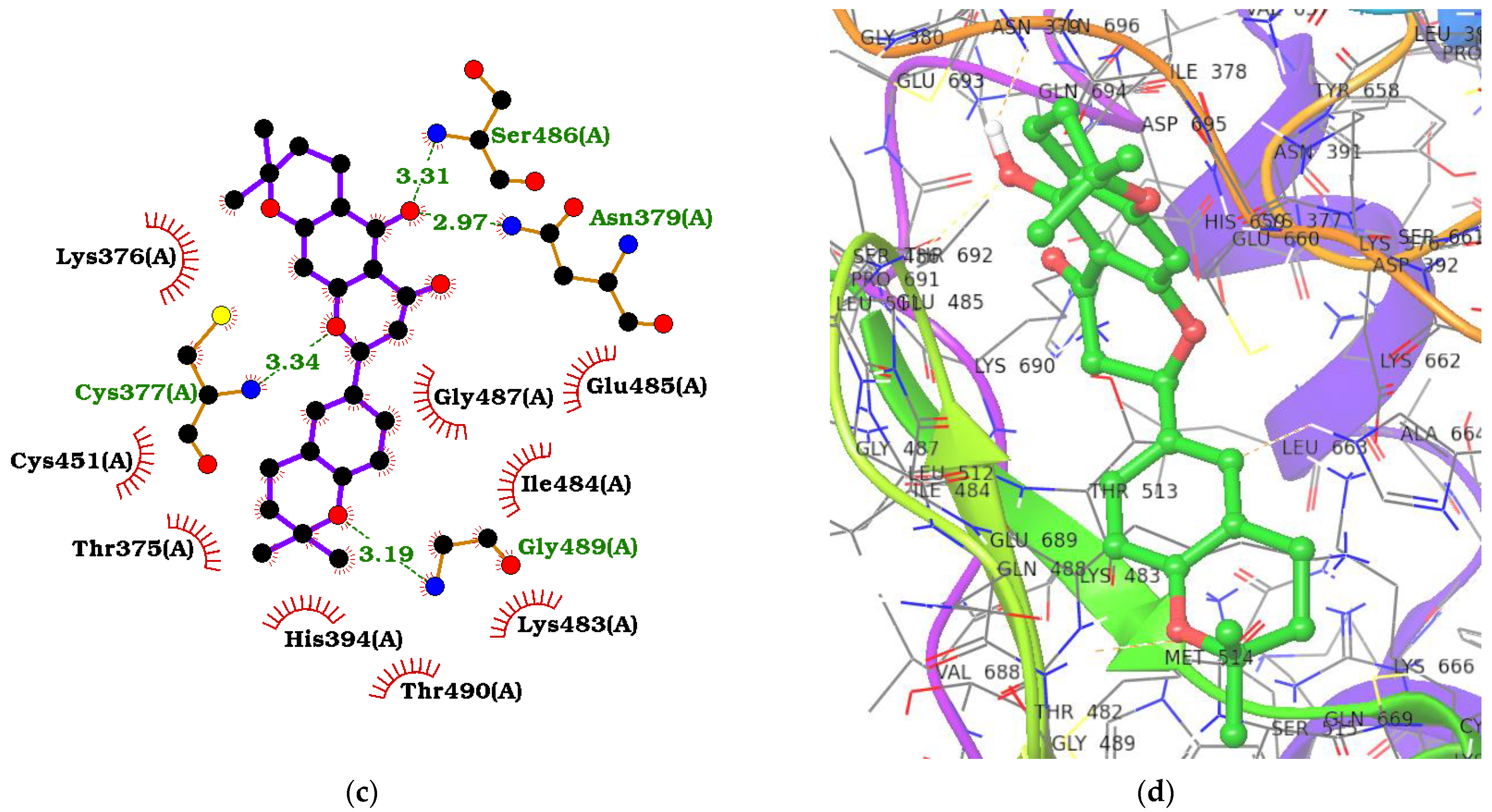
2.4. ADAR2–Ligand Interaction Profiling
2.5. Prediction of Biological Activity of the Selected Hit Compounds
2.6. Molecular Dynamics Simulations
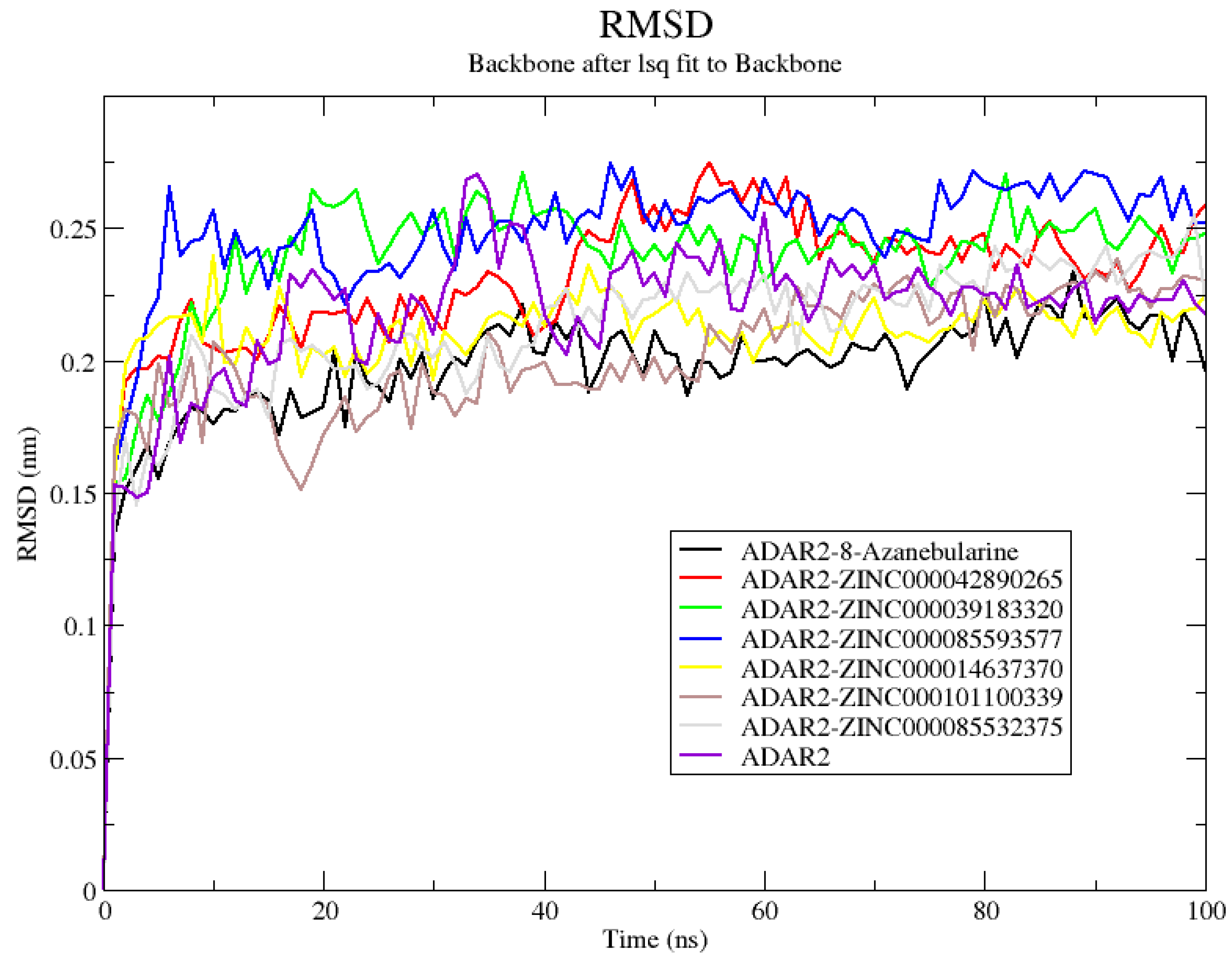
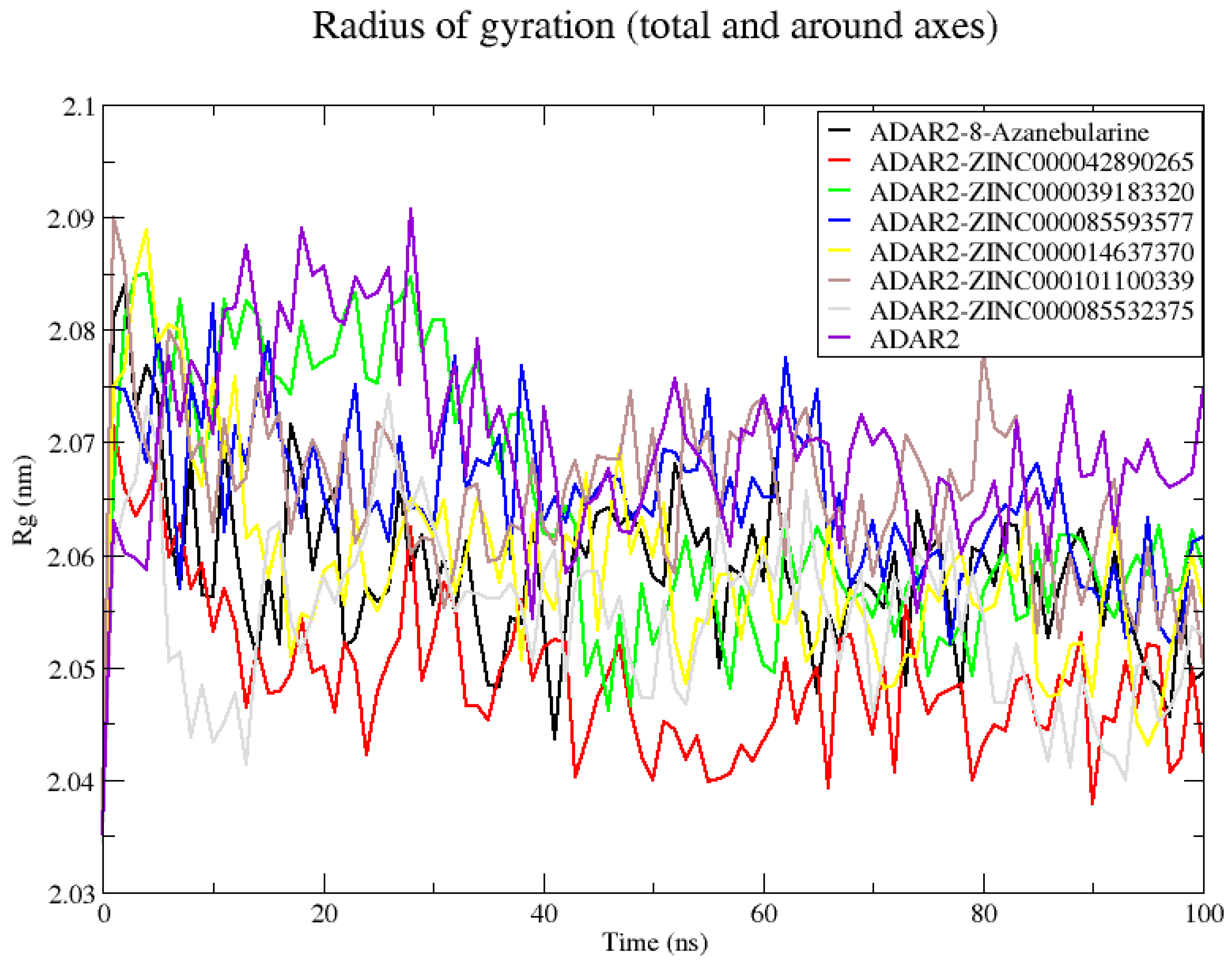
2.6.1. Analyzing RMSD, RMSF, and Rg
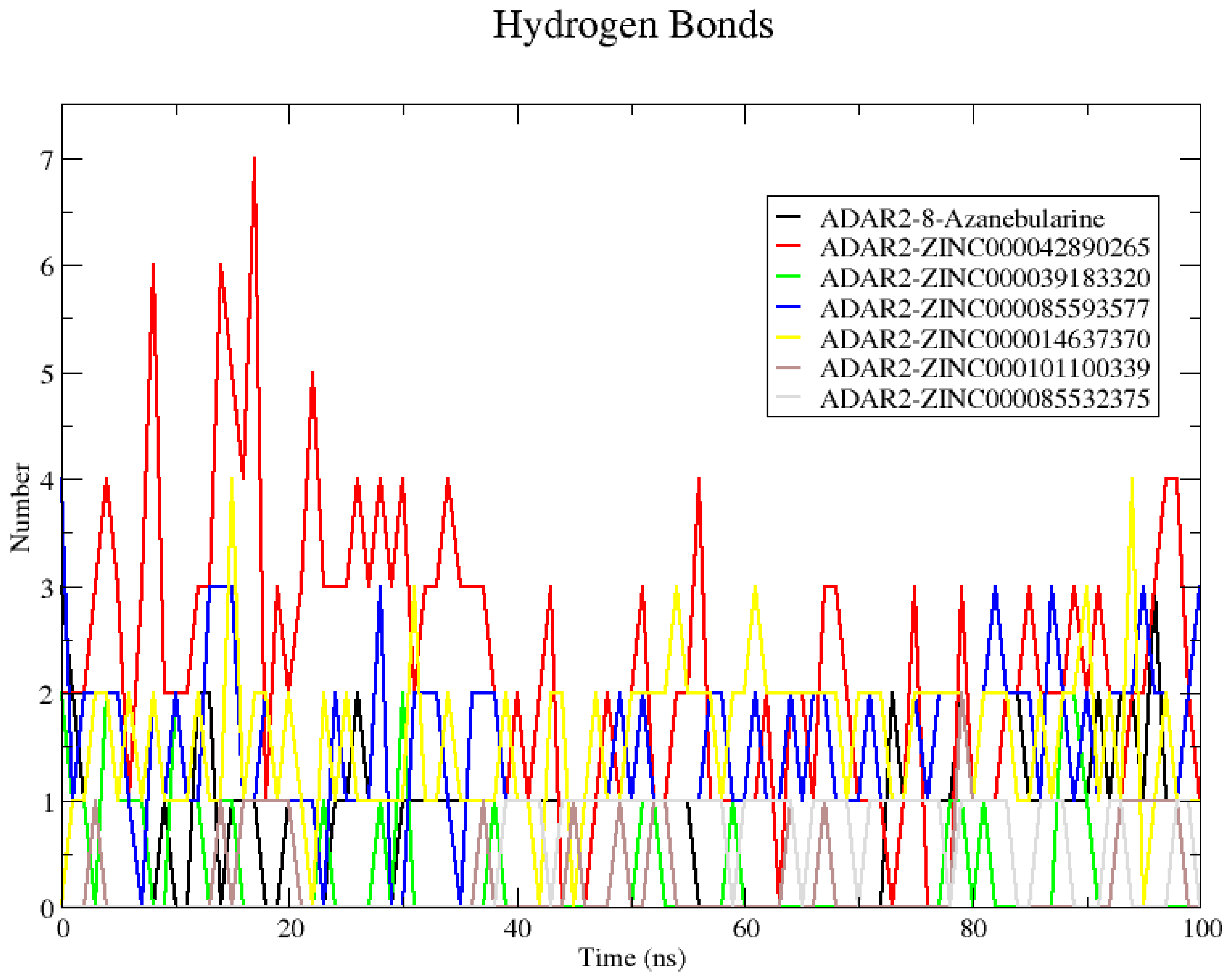
2.6.2. Analyzing Snapshots and Hydrogen Bonds
2.7. MM/PBSA Calculations for the ADAR2–Ligand Complexes
2.7.1. Analyzing Binding Free Energy
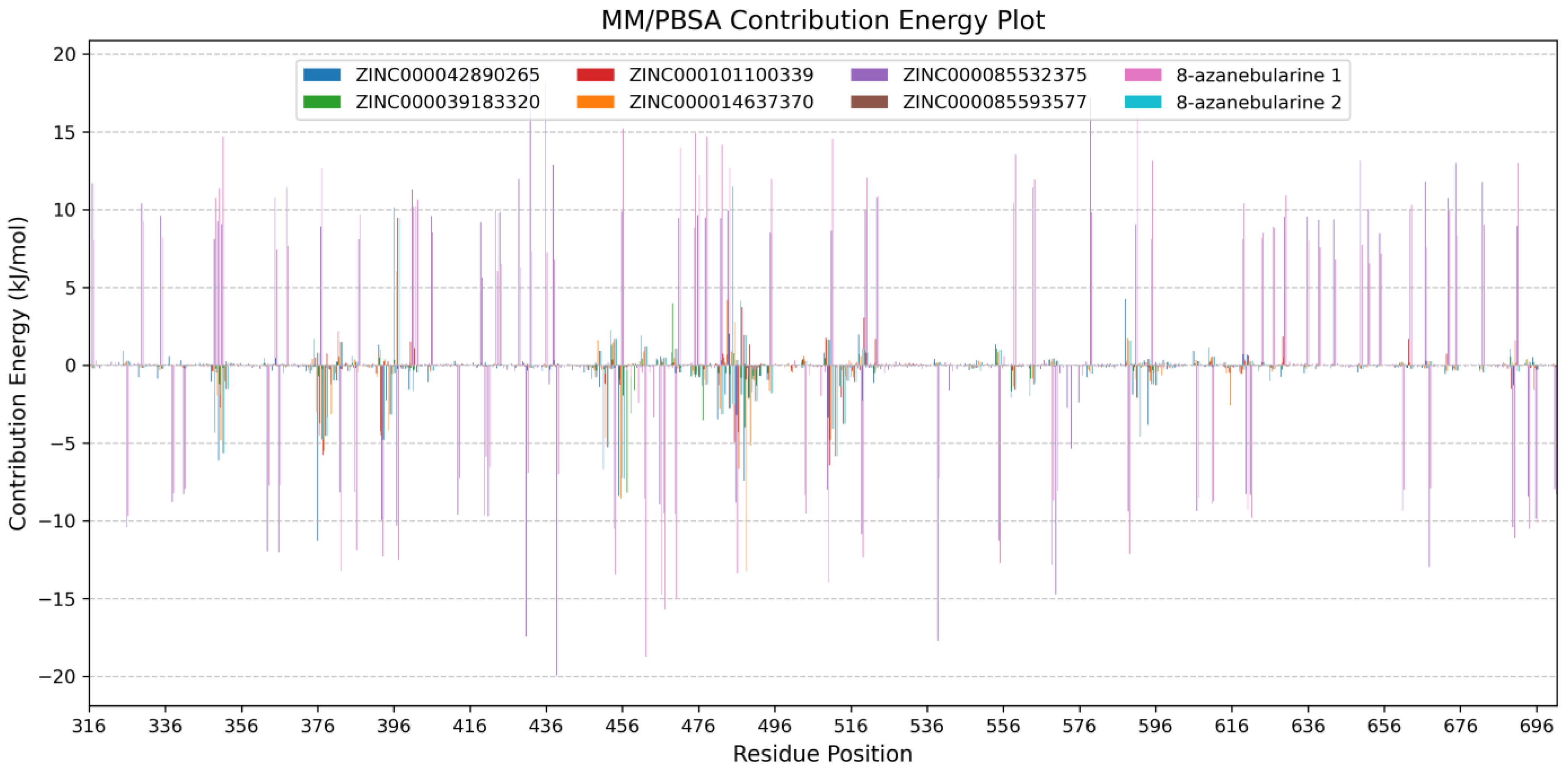
2.7.2. Analyzing Per-Residue Energy Contributions
2.8. Re-Docking of Top Compounds against the 5-HT2C Receptor
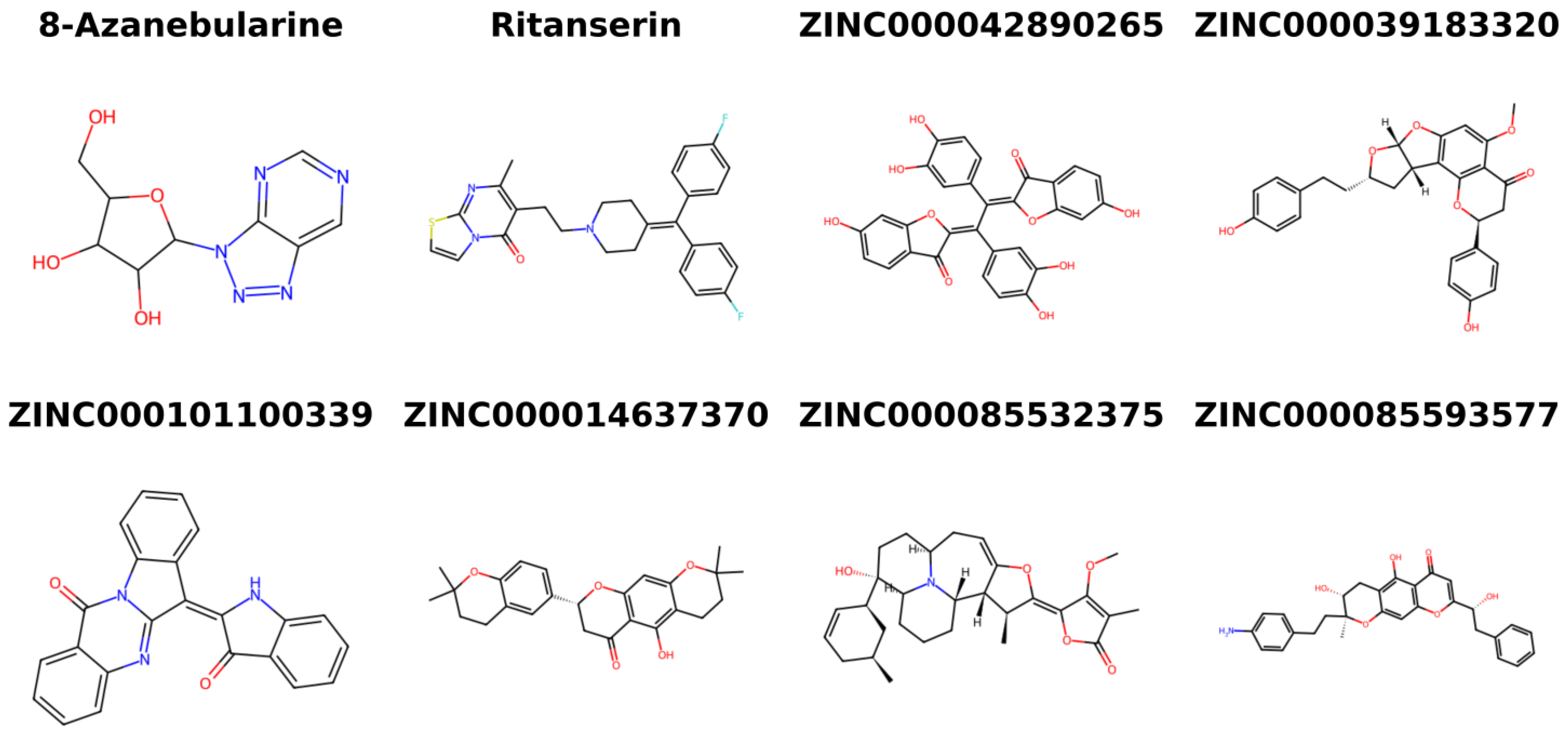
2.9. Provenance of Potential Lead Compounds
3. Materials and Methods
3.1. Protein and Ligands Preparation
3.2. Molecular Docking Studies
3.3. ADMET Profiling
3.4. Visualizing ADAR2–Ligand Interactions
3.5. Biological Activity Prediction of Shortlisted Compounds
3.6. Molecular Dynamics Simulations Study
3.7. Molecular Mechanics Poisson-Boltzmann Surface Area Calculation
3.8. Re-Docking Hit Compounds against the 5-HT2CR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bass, B.L.; Nishikura, K.; Keller, W.; Seeburg, P.H.; Emeson, R.B.; O’Connell, M.A.; Samuel, C.E.; Herbert, A. A standardized nomenclature for adenosine deaminases that act on RNA. RNA 1997, 3, 947–949. [Google Scholar] [PubMed]
- Polson, A.G.; Crain, P.F.; Pomerantz, S.C.; McCloskey, J.A.; Bass, B.L. The mechanism of adenosine to inosine conversion by the double-stranded RNA unwinding/modifying activity: A high-performance liquid chromatography-mass spectrometry analysis. Biochemistry 1991, 30, 11507–11514. [Google Scholar] [CrossRef] [PubMed]
- Cho, D.-S.C.; Yang, W.; Lee, J.T.; Shiekhattar, R.; Murray, J.M.; Nishikura, K. Requirement of Dimerization for RNA Editing Activity of Adenosine Deaminases Acting on RNA. J. Biol. Chem. 2003, 278, 17093–17102. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Maas, S.; Single, F.N.; Hartner, J.; Rozov, A.; Burnashev, N.; Feldmeyer, D.; Sprengel, R.; Seeburg, P.H. Point mutation in an AMPA receptor gene rescues lethality in mice deficient in the RNA-editing enzyme ADAR2. Nature 2000, 406, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Hideyama, T.; Yamashita, T.; Aizawa, H.; Tsuji, S.; Kakita, A.; Takahashi, H.; Kwak, S. Profound downregulation of the RNA editing enzyme ADAR2 in ALS spinal motor neurons. Neurobiol. Dis. 2012, 45, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Bazak, L.; Haviv, A.; Barak, M.; Jacob-Hirsch, J.; Deng, P.; Zhang, R.; Isaacs, F.J.; Rechavi, G.; Li, J.B.; Eisenberg, E.; et al. A-to-I RNA editing occurs at over a hundred million genomic sites, located in a majority of human genes. Genome Res. 2014, 24, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Athanasiadis, A.; Rich, A.; Maas, S. Widespread A-to-I RNA editing of Alu-containing mRNAs in the human transcriptome. PLoS Biol. 2004. [Google Scholar] [CrossRef] [Green Version]
- Piontkivska, H.; Wales-McGrath, B.; Miyamoto, M.; Wayne, M.L. ADAR Editing in Viruses: An Evolutionary Force to Reckon with. Genome Biol. Evol. 2021, 13. [Google Scholar] [CrossRef]
- Slotkin, W.; Nishikura, K. Adenosine-to-inosine RNA editing and human disease. Genome Med. 2013, 5, 105. [Google Scholar] [CrossRef]
- Rice, G.I.; Kasher, P.R.; Forte, G.M.A.; Mannion, N.M.; Greenwood, S.M.; Szynkiewicz, M.; Dickerson, J.E.; Bhaskar, S.S.; Zampini, M.; Briggs, T.A.; et al. Mutations in ADAR1 cause Aicardi-Goutières syndrome associated with a type I interferon signature. Nat. Genet. 2012, 44, 1243–1248. [Google Scholar] [CrossRef] [Green Version]
- Piekutowska-Abramczuk, D.; Mierzewska, H.; Bekiesińska-Figatowska, M.; Ciara, E.; Trubicka, J.; Pronicki, M.; Rokicki, D.; Rydzanicz, M.; Płoski, R.; Pronicka, E. Bilateral striatal necrosis caused by ADAR mutations in two siblings with dystonia and freckles-like skin changes that should be differentiated from Leigh syndrome. Folia Neuropathol. 2016, 4, 405–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatsiou, A.; Tual-Chalot, S.; Napoli, M.; Ortega-Gomez, A.; Regen, T.; Badolia, R.; Cesarini, V.; Garcia-Gonzalez, C.; Chevre, R.; Ciliberti, G.; et al. The RNA editor ADAR2 promotes immune cell trafficking by enhancing endothelial responses to interleukin-6 during sterile inflammation. Immunity 2023, 56, 979–997.e11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Qian, H.; Xu, J.; Gao, W. ADAR, the carcinogenesis mechanisms of ADAR and related clinical applications. Ann. Transl. Med. 2019, 7, 686. [Google Scholar] [CrossRef]
- Yang, Y.; Okada, S.; Sakurai, M. Adenosine-to-inosine RNA editing in neurological development and disease. RNA Biol. 2021, 18, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and Regulation of RNA Editing by ADAR Deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [Green Version]
- Nishikura, K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat. Rev. Mol. Cell Biol. 2016, 17, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Ma, C.-P.; Chen, Y.-T.; Schuyler, S.C.; Chang, K.-P.; Tan, B.C.-M. Functional Impact of RNA editing and ADARs on regulation of gene expression: Perspectives from deep sequencing studies. Cell Biosci. 2014, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Deng, P.; Khan, A.; Jacobson, D.; Sambrani, N.; McGurk, L.; Li, X.; Jayasree, A.; Hejatko, J.; Shohat-Ophir, G.; O’Connell, M.A.; et al. Adar RNA editing-dependent and -independent effects are required for brain and innate immune functions in Drosophila. Nat. Commun. 2020, 11, 1580. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Song, Y.; Chan, T.H.M.; Yang, H.; Lin, C.H.; Tay, D.J.T.; Hong, H.Q.; Tang, S.J.; Tan, K.T.; Huang, X.X.; et al. An RNA editing/dsRNA binding-independent gene regulatory mechanism of ADARs and its clinical implication in cancer. Nucleic Acids Res. 2017. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Grimberg, A.; Teegarden, S.; Mombereau, C.; Liu, S.; Bale, T.L.; Blendy, J.A.; Nishikura, K. Dysregulated Editing of Serotonin 2C Receptor mRNAs Results in Energy Dissipation and Loss of Fat Mass. J. Neurosci. 2008, 28, 12834–12844. [Google Scholar] [CrossRef] [Green Version]
- Mombereau, C.; Kawahara, Y.; Gundersen, B.B.; Nishikura, K.; Blendy, J.A. Functional relevance of serotonin 2C receptor mRNA editing in antidepressant- and anxiety-like behaviors. Neuropharmacology 2010, 59, 468–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eran, A.; Li, J.B.; Vatalaro, K.; McCarthy, J.; Rahimov, F.; Collins, C.; Markianos, K.; Margulies, D.M.; Brown, E.N.; Calvo, S.E.; et al. Comparative RNA editing in autistic and neurotypical cerebella. Mol. Psychiatry 2013, 18, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breen, M.S.; Dobbyn, A.; Li, Q.; Roussos, P.; Hoffman, G.E.; Stahl, E.; Chess, A.; Sklar, P.; Li, J.B.; Devlin, B.; et al. Global landscape and genetic regulation of RNA editing in cortical samples from individuals with schizophrenia. Nat. Neurosci. 2019, 22, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Barbon, A.; Magri, C. RNA Editing and Modifications in Mood Disorders. Genes 2020, 11, 872. [Google Scholar] [CrossRef]
- Tomaselli, S.; Galeano, F.; Alon, S.; Raho, S.; Galardi, S.; Polito, V.A.; Presutti, C.; Vincenti, S.; Eisenberg, E.; Locatelli, F.; et al. Modulation of microRNA editing, expression and processing by ADAR2 deaminase in glioblastoma. Genome Biol. 2015, 16, 5. [Google Scholar] [CrossRef]
- Alon, S.; Mor, E.; Vigneault, F.; Church, G.M.; Locatelli, F.; Galeano, F.; Gallo, A.; Shomron, N.; Eisenberg, E. Systematic identification of edited microRNAs in the human brain. Genome Res. 2012, 22, 1533–1540. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.-H.; Chen, C.-H.; Yeh, K.-H.; Li, C.-L.; Wu, Y.-J.; Chen, D.-S.; Chen, P.-J.; Yeh, S.-H. ADAR2-Mediated Editing of miR-214 and miR-122 Precursor and Antisense RNA Transcripts in Liver Cancers. PLoS ONE 2013, 8, e81922. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.H.M.; Lin, C.H.; Qi, L.; Fei, J.; Li, Y.; Yong, K.J.; Liu, M.; Song, Y.; Chow, R.K.K.; Ng, V.H.E.; et al. A disrupted RNA editing balance mediated by ADARs (Adenosine DeAminases that act on RNA) in human hepatocellular carcinoma. Gut 2014, 63, 832–843. [Google Scholar] [CrossRef]
- Samuel, C.E. Adenosine deaminases acting on RNA (ADARs) are both antiviral and proviral. Virology 2011, 411, 180–193. [Google Scholar] [CrossRef]
- Yanai, M.; Kojima, S.; Sakai, M.; Komorizono, R.; Tomonaga, K.; Makino, A. ADAR2 Is Involved in Self and Nonself Recognition of Borna Disease Virus Genomic RNA in the Nucleus. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [Green Version]
- Goodman, R.A.; Macbeth, M.R.; Beal, P.A. ADAR Proteins: Structure and Catalytic Mechanism. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–33. ISBN 9783642228001. [Google Scholar]
- Lehmann, K.A.; Bass, B.L. The importance of internal loops within RNA substrates of ADAR1. J. Mol. Biol. 1999, 291, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polson, A.G.; Bass, B.L. Preferential selection of adenosines for modification by double-stranded RNA adenosine deaminase. EMBO J. 1994, 13, 5701–5711. [Google Scholar] [CrossRef] [PubMed]
- Eggington, J.M.; Greene, T.; Bass, B.L. Predicting sites of ADAR editing in double-stranded RNA. Nat. Commun. 2011, 2, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, M.M.; Thomas, J.M.; Zheng, Y.; Tran, K.; Phelps, K.J.; Scott, A.I.; Havel, J.; Fisher, A.J.; Beal, P.A. Structures of human ADAR2 bound to dsRNA reveal base-flipping mechanism and basis for site selectivity. Nat. Struct. Mol. Biol. 2016, 23, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefl, R.; Oberstrass, F.C.; Hood, J.L.; Jourdan, M.; Zimmermann, M.; Skrisovska, L.; Maris, C.; Peng, L.; Hofr, C.; Emeson, R.B.; et al. The Solution Structure of the ADAR2 dsRBM-RNA Complex Reveals a Sequence-Specific Readout of the Minor Groove. Cell 2010, 143, 225–237. [Google Scholar] [CrossRef]
- Tian, B.; Bevilacqua, P.C.; Diegelman-Parente, A.; Mathews, M.B. The double-stranded-RNA-binding motif: Interference and much more. Nat. Rev. Mol. Cell Biol. 2004, 5, 1013–1023. [Google Scholar] [CrossRef]
- Wong, S.K.; Sato, S.; Lazinski, D.W. Substrate recognition by ADAR1 and ADAR2. RNA 2001, 7, S135583820101007X. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Beal, P.A. Probing RNA recognition by human ADAR2 using a high-throughput mutagenesis method. Nucleic Acids Res. 2016, 44, 9872–9880. [Google Scholar] [CrossRef]
- Tanaka, M.; Watanabe, Y. RNA Editing of Serotonin 2C Receptor and Alcohol Intake. Front. Neurosci. 2020, 13. [Google Scholar] [CrossRef] [Green Version]
- Zaidan, H.; Ramaswami, G.; Golumbic, Y.N.; Sher, N.; Malik, A.; Barak, M.; Galiani, D.; Dekel, N.; Li, J.B.; Gaisler-Salomon, I. A-to-I RNA editing in the rat brain is age-dependent, region-specific and sensitive to environmental stress across generations. BMC Genomics 2018, 19, 28. [Google Scholar] [CrossRef] [Green Version]
- Bhansali, P.; Dunning, J.; Singer, S.E.; David, L.; Schmauss, C. Early life stress alters adult serotonin 2C receptor pre-mRNA editing and expression of the α subunit of the heterotrimeric G-protein Gq. J. Neurosci. 2007. [Google Scholar] [CrossRef] [Green Version]
- Bombail, V.; Qing, W.; Chapman, K.E.; Holmes, M.C. Prevention of 5-hydroxytryptamine 2C receptor RNA editing and alternate splicing in C57BL/6 mice activates the hypothalamic-pituitary-adrenal axis and alters mood. Eur. J. Neurosci. 2014, 40, 3663–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sodhi, M.S.; Burnet, P.W.J.; Makoff, A.J.; Kerwin, R.W.; Harrison, P.J. RNA editing of the 5-HT2C receptor is reduced in schizophrenia. Mol. Psychiatry 2001, 6, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Narzo, A.F.; Kozlenkov, A.; Roussos, P.; Hao, K.; Hurd, Y.; Lewis, D.A.; Sibille, E.; Siever, L.J.; Koonin, E.; Dracheva, S. A unique gene expression signature associated with serotonin 2C receptor RNA editing in the prefrontal cortex and altered in suicide. Hum. Mol. Genet. 2014, 23, 4801–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissmann, D.; van der Laan, S.; Underwood, M.D.; Salvetat, N.; Cavarec, L.; Vincent, L.; Molina, F.; Mann, J.J.; Arango, V.; Pujol, J.F. Region-specific alterations of A-to-I RNA editing of serotonin 2c receptor in the cortex of suicides with major depression. Transl. Psychiatry 2016, 6, e878. [Google Scholar] [CrossRef] [Green Version]
- Parasuraman, S. Prediction of activity spectra for substances. J. Pharmacol. Pharmacother. 2011, 2, 52–53. [Google Scholar] [CrossRef] [Green Version]
- Lagunin, A.; Stepanchikova, A.; Filimonov, D.; Poroikov, V. PASS: Prediction of activity spectra for biologically active substances. Bioinformatics 2000, 16, 747–748. [Google Scholar] [CrossRef] [Green Version]
- Poroikov, V.V.; Filimonov, D.A.; Ihlenfeldt, W.-D.; Gloriozova, T.A.; Lagunin, A.A.; Borodina, Y.V.; Stepanchikova, A.V.; Nicklaus, M.C. PASS Biological Activity Spectrum Predictions in the Enhanced Open NCI Database Browser. J. Chem. Inf. Comput. Sci. 2003, 43, 228–236. [Google Scholar] [CrossRef]
- Broni, E.; Striegel, A.; Ashley, C.; Sakyi, P.O.; Peracha, S.; Velazquez, M.; Bebla, K.; Sodhi, M.; Kwofie, S.K.; Ademokunwa, A.; et al. Molecular Docking and Dynamics Simulation Studies Predict Potential Anti-ADAR2 Inhibitors: Implications for the Treatment of Cancer, Neurological, Immunological and Infectious Diseases. Int. J. Mol. Sci. 2023, 24, 6795. [Google Scholar] [CrossRef]
- Macbeth, M.R.; Schubert, H.L.; VanDemark, A.P.; Lingam, A.T.; Hill, C.P.; Bass, B.L. Inositol Hexakisphosphate Is Bound in the ADAR2 Core and Required for RNA Editing. Science 2005, 309, 1534–1539. [Google Scholar] [CrossRef] [Green Version]
- Véliz, E.A.; Easterwood, L.M.; Beal, P.A. Substrate Analogues for an RNA-Editing Adenosine Deaminase: Mechanistic Investigation and Inhibitor Design. J. Am. Chem. Soc. 2003, 125, 10867–10876. [Google Scholar] [CrossRef]
- Poli, G.; Tuccinardi, T. Consensus Docking in Drug Discovery. Curr. Bioact. Compd. 2020, 16, 182–190. [Google Scholar] [CrossRef]
- Fan, J.; Fu, A.; Zhang, L. Progress in molecular docking. Quant. Biol. 2019, 7, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Sander, T.; Freyss, J.; Von Korff, M.; Rufener, C. DataWarrior: An open-source program for chemistry aware data visualization and analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- Han, J.; An, O.; Hong, H.Q.; Chan, T.H.M.; Song, Y.; Shen, H.; Tang, S.J.; Lin, J.S.; Ng, V.H.E.; Tay, D.J.T.; et al. Suppression of adenosine-to-inosine (A-to-I) RNA editome by death associated protein 3 (DAP3) promotes cancer progression. Sci. Adv. 2020. [Google Scholar] [CrossRef] [PubMed]
- Valles, I.; Pajares, M.J.; Segura, V.; Guruceaga, E.; Gomez-Roman, J.; Blanco, D.; Tamura, A.; Montuenga, L.M.; Pio, R. Identification of Novel Deregulated RNA Metabolism-Related Genes in Non-Small Cell Lung Cancer. PLoS ONE 2012, 7, e42086. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-B.; Liao, X.-Y.; Zhang, J.-B.; Wang, F.; Qin, H.-D.; Zhang, L.; Shugart, Y.Y.; Zeng, Y.-X.; Jia, W.-H. ADAR2 functions as a tumor suppressor via editing IGFBP7 in esophageal squamous cell carcinoma. Int. J. Oncol. 2017, 50, 622–630. [Google Scholar] [CrossRef]
- Sakata, K.-I.; Maeda, K.; Sakurai, N.; Liang, S.; Nakazawa, S.; Yanagihara, K.; Kubo, T.; Yoshiyama, H.; Kitagawa, Y.; Hamada, J.-I.; et al. ADAR2 Regulates Malignant Behaviour of Mesothelioma Cells Independent of RNA-editing Activity. Anticancer Res. 2020, 40, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Filimonov, D.A.; Lagunin, A.A.; Gloriozova, T.A.; Rudik, A.V.; Druzhilovskii, D.S.; Pogodin, P.V.; Poroikov, V.V. Prediction of the Biological Activity Spectra of Organic Compounds Using the Pass Online Web Resource. Chem. Heterocycl. Compd. 2014, 50, 444–457. [Google Scholar] [CrossRef]
- Fava, M.; Rosenbaum, J.F.; Hoog, S.L.; Tepner, R.G.; Kopp, J.B.; Nilsson, M.E. Fluoxetine versus sertraline and paroxetine in major depression: Tolerability and efficacy in anxious depression. J. Affect. Disord. 2000. [Google Scholar] [CrossRef]
- Cipriani, A.; La Ferla, T.; Furukawa, T.A.; Signoretti, A.; Nakagawa, A.; Churchill, R.; McGuire, H.; Barbui, C. Sertraline versus other antidepressive agents for depression. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, R.; Ravindran, A.V.; Chaput, Y.; Goldner, E.; Swinson, R.; van Ameringen, M.A.; Austin, C.; Hadrava, V. Sertraline and Fluoxetine Treatment of Obsessive-Compulsive Disorder: Results of a Double-Blind, 6-Month Treatment Study. J. Clin. Psychopharmacol. 2002, 22, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Górska, N.; Słupski, J.; Cubała, W.J.; Wiglusz, M.S.; Gałuszko-Węgielnik, M. Antidepressants in epilepsy. Neurol. Neurochir. Pol. 2018, 52, 657–661. [Google Scholar] [CrossRef]
- Abboud, R.; Noronha, C.; Diwadkar, V.A. Motor system dysfunction in the schizophrenia diathesis: Neural systems to neurotransmitters. Eur. Psychiatry 2017, 44, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Walther, S.; Strik, W. Motor Symptoms and Schizophrenia. Neuropsychobiology 2012, 66, 77–92. [Google Scholar] [CrossRef]
- Lefebvre, S.; Pavlidou, A.; Walther, S. What is the potential of neurostimulation in the treatment of motor symptoms in schizophrenia? Expert Rev. Neurother. 2020, 20, 697–706. [Google Scholar] [CrossRef]
- Varlet, M.; Marin, L.; Raffard, S.; Schmidt, R.C.; Capdevielle, D.; Boulenger, J.P.; Del-Monte, J.; Bardy, B.G. Impairments of social motor coordination in schizophrenia. PLoS ONE 2012, 7, e29772. [Google Scholar] [CrossRef] [Green Version]
- Posar, A.; Visconti, P. Early Motor Signs in Autism Spectrum Disorder. Children 2022, 9, 294. [Google Scholar] [CrossRef]
- Mohd Nordin, A.; Ismail, J.; Kamal Nor, N. Motor Development in Children with Autism Spectrum Disorder. Front. Pediatr. 2021, 9, 598276. [Google Scholar] [CrossRef]
- Behm, M.; Wahlstedt, H.; Widmark, A.; Eriksson, M.; Öhman, M. Accumulation of nuclear ADAR2 regulates A-to-I RNA editing during neuronal development. J. Cell Sci. 2017, 130, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Marcucci, R.; Brindle, J.; Paro, S.; Casadio, A.; Hempel, S.; Morrice, N.; Bisso, A.; Keegan, L.P.; Del Sal, G.; O’Connell, M.A. Pin1 and WWP2 regulate GluR2 Q/R site RNA editing by ADAR2 with opposing effects. EMBO J. 2011, 30, 4211–4222. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wu, Y.; Yang, H.; Li, X.; Jie, M.; Hu, C.; Wu, Y.; Yang, S.; Yang, Y. Prolyl isomerase Pin1: A promoter of cancer and a target for therapy. Cell Death Dis. 2018, 9, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yang, X.; Cui, Y.; Zhang, X. Suppression of RNA editing by miR-17 inhibits the stemness of melanoma stem cells. Mol. Ther. Nucleic Acids 2022, 27, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Tomonaga, K. Nucleocytoplasmic Shuttling of Viral Proteins in Borna Disease Virus Infection. Viruses 2013, 5, 1978–1990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubitt, B.; de la Torre, J.C. Borna disease virus (BDV), a nonsegmented RNA virus, replicates in the nuclei of infected cells where infectious BDV ribonucleoproteins are present. J. Virol. 1994, 68, 1371–1381. [Google Scholar] [CrossRef]
- Wang, Q.; Li, X.; Qi, R.; Billiar, T. RNA Editing, ADAR1, and the Innate Immune Response. Genes 2017, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, J.C. Bornavirus and the Brain. J. Infect. Dis. 2002, 186, S241–S247. [Google Scholar] [CrossRef]
- WENSMAN, J.J.; BERG, M.; BERG, A.-L. Experiences of Borna Disease Virus infection in Sweden. APMIS 2008, 116, 46–49. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, G.; Liu, J.; Song, J.; Zhang, Z.; Huang, Q.; Wei, F. Tanshinone I and Tanshinone IIA/B attenuate LPS-induced mastitis via regulating the NF-κB. Biomed. Pharmacother. 2021. [Google Scholar] [CrossRef]
- Wang, X.; Fan, J.; Ding, X.; Sun, Y.; Cui, Z.; Liu, W. Tanshinone i inhibits il-1β-induced apoptosis, inflammation and extracellular matrix degradation in chondrocytes CHON-001 cells and attenuates murine osteoarthritis. Drug Des. Devel. Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Jing, H.; Yang, H.; Liu, Z.; Guo, H.; Chai, L.; Hu, L. Tanshinone i selectively suppresses pro-inflammatory genes expression in activated microglia and prevents nigrostriatal dopaminergic neurodegeneration in a mouse model of Parkinson’s disease. J. Ethnopharmacol. 2015, 164, 247–255. [Google Scholar] [CrossRef]
- Nizamutdinova Tanshinone I effectively induces apoptosis in estrogen receptor-positive (MCF-7) and estrogen receptor-negative (MDA-MB-231) breast cancer cells. Int. J. Oncol. 2008, 33, 485–491. [CrossRef]
- Zheng, L.; Zhang, Y.; Liu, G.; Cheng, S.; Zhang, G.; An, C.; Sun, S.; Wang, J.; Pang, B.; Li, S. Tanshinone i regulates autophagic signaling via the activation of AMP-activated protein kinase in cancer cells. Anticancer. Drugs 2020, 31, 601–608. [Google Scholar] [CrossRef]
- Lu, M.; Wang, C.; Wang, J. Tanshinone I induces human colorectal cancer cell apoptosis: The potential roles of Aurora A-p53 and survivin-mediated signaling pathways. Int. J. Oncol. 2016, 49, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng Su Growth inhibition and apoptosis induction by tanshinone I in human colon cancer Colo 205 cells. Int. J. Mol. Med. 2008, 22, 613–618. [CrossRef]
- Jing, X.; Xu, Y.; Cheng, W.; Guo, S.; Zou, Y.; He, L. Tanshinone I induces apoptosis and pro-survival autophagy in gastric cancers. Cancer Chemother. Pharmacol. 2016, 77, 1171–1181. [Google Scholar] [CrossRef]
- Cui, S.; Chen, T.; Wang, M.; Chen, Y.; Zheng, Q.; Feng, X.; Li, S.; Wang, J. Tanshinone I inhibits metastasis of cervical cancer cells by inducing BNIP3/NIX-mediated mitophagy and reprogramming mitochondrial metabolism. Phytomedicine 2022, 98, 153958. [Google Scholar] [CrossRef]
- Zhou, J.; Jiang, Y.; Chen, H.; Wu, Y.-C.; Zhang, L. Tanshinone I attenuates the malignant biological properties of ovarian cancer by inducing apoptosis and autophagy via the inactivation of PI3K/AKT/mTOR pathway. Cell Prolif. 2020, 53, e12739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, S.; Chen, L.; Minxue, L.; Hongmin, C.; Ronghua, T.; Xiaoxuan, F.; Binbin, Z.; Shiwen, G. Tanshinone I induces apoptosis and protective autophagy in human glioblastoma cells via a reactive oxygen species-dependent pathway. Int. J. Mol. Med. 2020, 45, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Liu, Y.; Dong, Z. Tanshinone I alleviates motor and cognitive impairments via suppressing oxidative stress in the neonatal rats after hypoxic-ischemic brain damage. Mol. Brain 2017, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Jeon, S.J.; Jung, J.W.; Lee, S.; Yoon, B.H.; Shin, B.Y.; Son, K.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; et al. Tanshinone congeners improve memory impairments induced by scopolamine on passive avoidance tasks in mice. Eur. J. Pharmacol. 2007. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, S.; Jeon, S.J.; Son, K.H.; Lee, S.; Yoon, B.H.; Cheong, J.H.; Ko, K.H.; Ryu, J.H. Tanshinone I enhances learning and memory, and ameliorates memory impairment in mice via the extracellular signal-regulated kinase signalling pathway. Br. J. Pharmacol. 2009, 158, 1131–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patlolla, J.M.R.; Rao, C.V. Anti-inflammatory and Anti-cancer Properties of β-Escin, a Triterpene Saponin. Curr. Pharmacol. Rep. 2015, 1, 170–178. [Google Scholar] [CrossRef]
- Kenny, H.A.; Hart, P.C.; Kordylewicz, K.; Lal, M.; Shen, M.; Kara, B.; Chen, Y.-J.; Grassl, N.; Alharbi, Y.; Pattnaik, B.R.; et al. The Natural Product β-Escin Targets Cancer and Stromal Cells of the Tumor Microenvironment to Inhibit Ovarian Cancer Metastasis. Cancers 2021, 13, 3931. [Google Scholar] [CrossRef] [PubMed]
- Mojžišová, G.; Kello, M.; Pilátová, M.; Tomečková, V.; Vašková, J.; Vaško, L.; Bernátová, S.; Mirossay, L.; Mojžiš, J. Antiproliferative effect of β-escin—An in vitro study. Acta Biochim. Pol. 2016, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Fu, F.; Zhang, X.; Zhu, M.; Wang, T.; Fan, H. Escin attenuates cognitive deficits and hippocampal injury after transient global cerebral ischemia in mice via regulating certain inflammatory genes. Neurochem. Int. 2010, 57, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, G.P.; Janakiraman, U.; Essa, M.M.; Justin Thenmozhi, A.; Manivasagam, T. Escin attenuates behavioral impairments, oxidative stress and inflammation in a chronic MPTP/probenecid mouse model of Parkinson’s disease. Brain Res. 2014, 1585, 23–36. [Google Scholar] [CrossRef]
- Barter, J.W.; Li, S.; Lu, D.; Bartholomew, R.A.; Rossi, M.A.; Shoemaker, C.T.; Salas-Meza, D.; Gaidis, E.; Yin, H.H. Beyond reward prediction errors: The role of dopamine in movement kinematics. Front. Integr. Neurosci. 2015, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Faynveitz, A.; Lavian, H.; Jacob, A.; Korngreen, A. Proliferation of Inhibitory Input to the Substantia Nigra in Experimental Parkinsonism. Front. Cell. Neurosci. 2019, 13, 417. [Google Scholar] [CrossRef]
- Ayano, G. Dopamine: Receptors, Functions, Synthesis, Pathways, Locations and Mental Disorders: Review of Literatures. J. Ment. Disord. Treat. 2016, 2, 2. [Google Scholar] [CrossRef]
- Yoshida, Y.; Fujigaki, H.; Kato, K.; Yamazaki, K.; Fujigaki, S.; Kunisawa, K.; Yamamoto, Y.; Mouri, A.; Oda, A.; Nabeshima, T.; et al. Selective and competitive inhibition of kynurenine aminotransferase 2 by glycyrrhizic acid and its analogues. Sci. Rep. 2019, 9, 10243. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Nakagome, I.; Hirono, S.; Itoh, T.; Fujiwara, R. Inhibition of adenosine deaminase (ADA)-mediated metabolism of cordycepin by natural substances. Pharmacol. Res. Perspect. 2015, 3, e00121. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-J.; Lee, J.-C.; Hsieh, J.-H.; Lin, M.-Y.; Shih, I.-A.; You, H.-L.; Wang, K. Cordycepin inhibits the proliferation of malignant peripheral nerve sheath tumor cells through the p53/Sp1/tubulin pathway. Am. J. Cancer Res. 2021, 11, 1247–1266. [Google Scholar] [PubMed]
- Rawat, R.S.; Kumar, S. Understanding the mode of inhibition and molecular interaction of taxifolin with human adenosine deaminase. J. Biomol. Struct. Dyn. 2023, 41, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Ishola, I.O.; Ben-Azu, B.; Adebayo, O.A.; Ajayi, A.M.; Omorodion, I.L.; Edje, K.E.; Adeyemi, O.O. Prevention and reversal of ketamine-induced experimental psychosis in mice by the neuroactive flavonoid, hesperidin: The role of oxidative and cholinergic mechanisms. Brain Res. Bull. 2021, 177, 239–251. [Google Scholar] [CrossRef]
- Thayumanavan, G.; Jeyabalan, S.; Fuloria, S.; Sekar, M.; Ravi, M.; Selvaraj, L.K.; Bala, L.; Chidambaram, K.; Gan, S.H.; Rani, N.N.I.M.; et al. Silibinin and Naringenin against Bisphenol A-Induced Neurotoxicity in Zebrafish Model—Potential Flavonoid Molecules for New Drug Design, Development, and Therapy for Neurological Disorders. Molecules 2022, 27, 2572. [Google Scholar] [CrossRef]
- Carry, E.; Kshatriya, D.; Silva, J.; Davies, D.L.; Yuan, B.; Wu, Q.; Patel, H.; Park, E.R.; Gilleran, J.; Hao, L.; et al. Identification of Dihydromyricetin and Metabolites in Serum and Brain Associated with Acute Anti-Ethanol Intoxicating Effects in Mice. Int. J. Mol. Sci. 2021, 22, 7460. [Google Scholar] [CrossRef]
- Al Omran, A.J.; Shao, A.S.; Watanabe, S.; Zhang, Z.; Zhang, J.; Xue, C.; Watanabe, J.; Davies, D.L.; Shao, X.M.; Liang, J. Social isolation induces neuroinflammation and microglia overactivation, while dihydromyricetin prevents and improves them. J. Neuroinflamm. 2022, 19, 2. [Google Scholar] [CrossRef]
- Durrant, J.D.; McCammon, J.A. Molecular dynamics simulations and drug discovery. BMC Biol. 2011, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- De Vivo, M.; Masetti, M.; Bottegoni, G.; Cavalli, A. Role of Molecular Dynamics and Related Methods in Drug Discovery. J. Med. Chem. 2016, 59, 4035–4061. [Google Scholar] [CrossRef]
- Cheng, X.; Ivanov, I. Molecular Dynamics. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: Totowa, NJ, USA, 2012; Volume 929, pp. 243–285. ISBN 9781627030496. [Google Scholar]
- Childers, M.C.; Daggett, V. Validating Molecular Dynamics Simulations against Experimental Observables in Light of Underlying Conformational Ensembles. J. Phys. Chem. B 2018, 122, 6673–6689. [Google Scholar] [CrossRef]
- Wadhwa, R.; Yadav, N.S.; Katiyar, S.P.; Yaguchi, T.; Lee, C.; Ahn, H.; Yun, C.-O.; Kaul, S.C.; Sundar, D. Molecular dynamics simulations and experimental studies reveal differential permeability of withaferin-A and withanone across the model cell membrane. Sci. Rep. 2021, 11, 2352. [Google Scholar] [CrossRef]
- Adelusi, T.I.; Oyedele, A.-Q.K.; Boyenle, I.D.; Ogunlana, A.T.; Adeyemi, R.O.; Ukachi, C.D.; Idris, M.O.; Olaoba, O.T.; Adedotun, I.O.; Kolawole, O.E.; et al. Molecular modeling in drug discovery. Inform. Med. Unlocked 2022, 29, 100880. [Google Scholar] [CrossRef]
- Singh, W.; Karabencheva-Christova, T.G.; Black, G.W.; Ainsley, J.; Dover, L.; Christov, C.Z. Conformational Dynamics, Ligand Binding and Effects of Mutations in NirE an S-Adenosyl-L-Methionine Dependent Methyltransferase. Sci. Rep. 2016, 6, 20107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Mackerell, A.D.; Brooks, C.L. CHARMM fluctuating charge force field for proteins: II Protein/solvent properties from molecular dynamics simulations using a nonadditive electrostatic model. J. Comput. Chem. 2004, 25, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Lobanov, M.Y.; Bogatyreva, N.S.; Galzitskaya, O.V. Radius of gyration as an indicator of protein structure compactness. Mol. Biol. 2008, 42, 623–628. [Google Scholar] [CrossRef]
- De Vita, S.; Chini, M.G.; Bifulco, G.; Lauro, G. Insights into the Ligand Binding to Bromodomain-Containing Protein 9 (BRD9): A Guide to the Selection of Potential Binders by Computational Methods. Molecules 2021, 26, 7192. [Google Scholar] [CrossRef]
- Wen, C.C.; Kuo, Y.H.; Jan, J.T.; Liang, P.H.; Wang, S.Y.; Liu, H.G.; Lee, C.K.; Chang, S.T.; Kuo, C.J.; Lee, S.S.; et al. Specific plant terpenoids and lignoids possess potent antiviral activities against severe acute respiratory syndrome coronavirus. J. Med. Chem. 2007, 50, 4087–4095. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Oezguen, N.; Urvil, P.; Ferguson, C.; Dann, S.M.; Savidge, T.C. Regulation of protein-ligand binding affinity by hydrogen bond pairing. Sci. Adv. 2016, 2, e1501240. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Kumar, R.; Lynn, A. g_mmpbsa—A GROMACS Tool for High-Throughput MM-PBSA Calculations. J. Chem. Inf. Model. 2014, 54, 1951–1962. [Google Scholar] [CrossRef]
- Wang, C.; Nguyen, P.H.; Pham, K.; Huynh, D.; Le, T.B.N.; Wang, H.; Ren, P.; Luo, R. Calculating protein-ligand binding affinities with MMPBSA: Method and error analysis. J. Comput. Chem. 2016, 37, 2436–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Greene, D.; Xiao, L.; Qi, R.; Luo, R. Recent Developments and Applications of the MMPBSA Method. Front. Mol. Biosci. 2018, 4, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- José dos Santos Nascimento, I.; Mendonça de Aquino, T.; da Silva Júnior, E.F.; Olimpio de Moura, R. Insights on Microsomal Prostaglandin E2 synthase 1 (mPGES-1) Inhibitors Using Molecular Dynamics and MM/PBSA calculations. Lett. Drug Des. Discov. 2023, 22, 2435–2462. [Google Scholar] [CrossRef]
- Silva, L.R.; Guimarães, A.S.; do Nascimento, J.; do Santos Nascimento, I.J.; da Silva, E.B.; McKerrow, J.H.; Cardoso, S.H.; da Silva-Júnior, E.F. Computer-aided design of 1,4-naphthoquinone-based inhibitors targeting cruzain and rhodesain cysteine proteases. Bioorganic Med. Chem. 2021, 41, 116213. [Google Scholar] [CrossRef] [PubMed]
- Obiol-Pardo, C.; Rubio-Martinez, J. Comparative Evaluation of MMPBSA and XSCORE to Compute Binding Free Energy in XIAP−Peptide Complexes. J. Chem. Inf. Model. 2007, 47, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Breznik, M.; Ge, Y.; Bluck, J.P.; Briem, H.; Hahn, D.F.; Christ, C.D.; Mortier, J.; Mobley, D.L.; Meier, K. Prioritizing Small Sets of Molecules for Synthesis through in-silico Tools: A Comparison of Common Ranking Methods. ChemMedChem 2023, 18, e202200425. [Google Scholar] [CrossRef]
- Warren, G.L.; Andrews, C.W.; Capelli, A.-M.; Clarke, B.; LaLonde, J.; Lambert, M.H.; Lindvall, M.; Nevins, N.; Semus, S.F.; Senger, S.; et al. A Critical Assessment of Docking Programs and Scoring Functions. J. Med. Chem. 2006, 49, 5912–5931. [Google Scholar] [CrossRef] [PubMed]
- Broni, E.; Ashley, C.; Adams, J.; Manu, H.; Aikins, E.; Okom, M.; Miller, W.A.; Wilson, M.D.; Kwofie, S.K. Cheminformatics-Based Study Identifies Potential Ebola VP40 Inhibitors. Int. J. Mol. Sci. 2023, 24, 6298. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.J.; Beal, P.A. Effects of Aicardi-Goutières syndrome mutations predicted from ADAR-RNA structures. RNA Biol. 2017, 14, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Rosani, U.; Bai, C.-M.; Maso, L.; Shapiro, M.; Abbadi, M.; Domeneghetti, S.; Wang, C.-M.; Cendron, L.; MacCarthy, T.; Venier, P. A-to-I editing of Malacoherpesviridae RNAs supports the antiviral role of ADAR1 in mollusks. BMC Evol. Biol. 2019, 19, 149. [Google Scholar] [CrossRef] [Green Version]
- Sterling, T.; Irwin, J.J. ZINC 15-Ligand Discovery for Everyone. J. Chem. Inf. Model. 2015, 55, 2324–2337. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Mendez, D.; Gaulton, A.; Bento, A.P.; Chambers, J.; De Veij, M.; Félix, E.; Magariños, M.P.; Mosquera, J.F.; Mutowo, P.; Nowotka, M.; et al. ChEMBL: Towards direct deposition of bioassay data. Nucleic Acids Res. 2019, 47, D930–D940. [Google Scholar] [CrossRef] [PubMed]
- Gaulton, A.; Bellis, L.J.; Bento, A.P.; Chambers, J.; Davies, M.; Hersey, A.; Light, Y.; McGlinchey, S.; Michalovich, D.; Al-Lazikani, B.; et al. ChEMBL: A large-scale bioactivity database for drug discovery. Nucleic Acids Res. 2012, 40, D1100–D1107. [Google Scholar] [CrossRef] [Green Version]
- Bento, A.P.; Gaulton, A.; Hersey, A.; Bellis, L.J.; Chambers, J.; Davies, M.; Krüger, F.A.; Light, Y.; Mak, L.; McGlinchey, S.; et al. The ChEMBL bioactivity database: An update. Nucleic Acids Res. 2014, 42, D1083–D1090. [Google Scholar] [CrossRef] [Green Version]
- Rutz, A.; Sorokina, M.; Galgonek, J.; Mietchen, D.; Willighagen, E.; Gaudry, A.; Graham, J.G.; Stephan, R.; Page, R.; Vondrášek, J.; et al. The LOTUS initiative for knowledge sharing in Natural Products research. Planta Med. 2021, 87, 1248–1249. [Google Scholar]
- Westenburg, H.E.; Lee, K.-J.; Lee, S.K.; Fong, H.H.S.; van Breemen, R.B.; Pezzuto, J.M.; Kinghorn, A.D. Activity-Guided Isolation of Antioxidative Constituents of Cotinus coggygria. J. Nat. Prod. 2000, 63, 1696–1698. [Google Scholar] [CrossRef]
- Sukhikh, S.; Noskova, S.; Pungin, A.; Ivanova, S.; Skrypnik, L.; Chupakhin, E.; Babich, O. Study of the Biologically Active Properties of Medicinal Plant Cotinus coggygria. Plants 2021, 10, 1224. [Google Scholar] [CrossRef]
- Antal, D.S.; Ardelean, F.; Jijie, R.; Pinzaru, I.; Soica, C.; Dehelean, C. Integrating Ethnobotany, Phytochemistry, and Pharmacology of Cotinus coggygria and Toxicodendron vernicifluum: What Predictions can be Made for the European Smoketree? Front. Pharmacol. 2021, 12, 662852. [Google Scholar] [CrossRef]
- Thapa, P.; Prakash, O.; Rawat, A.; Kumar, R.; Srivastava, R.M.; Rawat, D.S.; Pant, A.K. Essential Oil Composition, Antioxidant, Anti-inflammatory, Insect Antifeedant and Sprout Suppressant Activity in Essential Oil from Aerial Parts of Cotinus coggygria Scop. J. Essent. Oil Bear. Plants 2020, 23, 65–76. [Google Scholar] [CrossRef]
- Marčetić, M.; Božić, D.; Milenković, M.; Malešević, N.; Radulović, S.; Kovačević, N. Antimicrobial, antioxidant and anti-inflammatory activity of young shoots of the smoke tree, Cotinus coggygria Scop. Phyther. Res. 2013, 27, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Banskota, A.H.; Tezuka, Y.; Saiki, I.; Kadota, S. Antiproliferative Activity of Diarylheptanoids from the Seeds of Alpinia blepharocalyx. Biol. Pharm. Bull. 2001, 24, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, Y.; Gewali, M.B.; Ali, M.S.; Banskota, A.H.; Kadota, S. Eleven Novel Diarylheptanoids and Two Unusual Diarylheptanoid Derivatives from the Seeds of Alpinia blepharocalyx. J. Nat. Prod. 2001, 64, 208–213. [Google Scholar] [CrossRef]
- Kadota, S.; Tezuka, Y.; Prasain, J.; Shawkat Ali, M.; Banskota, A. Novel Diarylheptanoids of Alpinia blepharocalyx. Curr. Top. Med. Chem. 2003, 3, 203–225. [Google Scholar] [CrossRef]
- Bergman, J. The identity of candidine and qingdainone. Phytochemistry 1989, 28, 3547. [Google Scholar] [CrossRef]
- Li, Z.; Li, L.; Chen, T.; Li, C.; Wang, D.; Yang, Z.; Zhong, N. Efficacy and safety of Ban-Lan-Gen granules in the treatment of seasonal influenza: Study protocol for a randomized controlled trial. Trials 2015, 16, 126. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Fan, F.; Hou, X.; Wu, H.; Wang, J.; Xu, H.; Sun, Y. 4(3H)-Quinazolone regulates innate immune signaling upon respiratory syncytial virus infection by moderately inhibiting the RIG-1 pathway in RAW264.7 cell. Int. Immunopharmacol. 2017, 52, 245–252. [Google Scholar] [CrossRef]
- Zhang, D.; Shi, Y.; Li, J.; Ruan, D.; Jia, Q.; Zhu, W.; Chen, K.; Li, Y.; Wang, R. Alkaloids with Nitric Oxide Inhibitory Activities from the Roots of Isatis tinctoria. Molecules 2019, 24, 4033. [Google Scholar] [CrossRef] [Green Version]
- Lotts, T.; Kabrodt, K.; Hummel, J.; Binder, D.; Schellenberg, I.; Ständer, S.; Agelopoulos, K. Isatis tinctoria L.-derived Petroleum Ether Extract Mediates Anti-inflammatory Effects via Inhibition of Interleukin-6, Interleukin-33 and Mast Cell Degranulation. Acta Derm. Venereol. 2020, 100, adv00131-9. [Google Scholar] [CrossRef]
- Nicosia, N.; Kwiecień, I.; Mazurek, J.; Mika, K.; Bednarski, M.; Miceli, N.; Ragusa, S.; Ekiert, H.; Maes, M.; Kotańska, M. Hydroalcoholic Leaf Extract of Isatis tinctoria L. via Antioxidative and Anti-Inflammatory Effects Reduces Stress-Induced Behavioral and Cellular Disorders in Mice. Oxid. Med. Cell. Longev. 2022, 2022, 3567879. [Google Scholar] [CrossRef]
- Rose, P.W.; Prlić, A.; Altunkaya, A.; Bi, C.; Bradley, A.R.; Christie, C.H.; Di Costanzo, L.; Duarte, J.M.; Dutta, S.; Feng, Z.; et al. The RCSB protein data bank: Integrative view of protein, gene and 3D structural information. Nucleic Acids Res. 2017, 45, D271–D281. [Google Scholar] [CrossRef]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef]
- Lu, C.; Wu, C.; Ghoreishi, D.; Chen, W.; Wang, L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; Von Bargen, C.D.; et al. OPLS4: Improving Force Field Accuracy on Challenging Regimes of Chemical Space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef]
- Chen, C.Y.-C. TCM Database@Taiwan: The world’s largest traditional Chinese medicine database for drug screening in silico. PLoS ONE 2011, 6, e15939. [Google Scholar] [CrossRef] [Green Version]
- Kwofie, S.; Broni, E.; Yunus, F.; Nsoh, J.; Adoboe, D.; Miller, W.; Wilson, M. Molecular Docking Simulation Studies Identifies Potential Natural Product Derived-Antiwolbachial Compounds as Filaricides against Onchocerciasis. Biomedicines 2021, 9, 1682. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veber, D.F.; Johnson, S.R.; Cheng, H.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular Properties That Influence the Oral Bioavailability of Drug Candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings1PII of original article: S0169-409X(96)00423-1. The article was originally published in Advanced Drug Delivery Reviews 23 (1997) 3. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]
- Barret, R. Lipinski’s Rule of Five. In Therapeutical Chemistry; Elsevier: Amsterdam, The Netherlands, 2018; Volume 1, pp. 7–100. [Google Scholar]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand–Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Dodda, L.S.; Cabeza de Vaca, I.; Tirado-Rives, J.; Jorgensen, W.L. LigParGen web server: An automatic OPLS-AA parameter generator for organic ligands. Nucleic Acids Res. 2017, 45, W331–W336. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.; Viet, M.H.; Li, M.S. Effects of water models on binding affinity: Evidence from all-atom simulation of binding of tamiflu to A/H5N1 neuraminidase. Sci. World J. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Z.; Liu, L.; Zhao, L.; Wang, J. Effects of different force fields and temperatures on the structural character of abeta (12–28) peptide in aqueous solution. Int. J. Mol. Sci. 2011, 12, 8259. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; McCorvy, J.D.; Harpsøe, K.; Lansu, K.; Yuan, S.; Popov, P.; Qu, L.; Pu, M.; Che, T.; Nikolajsen, L.F.; et al. 5-HT2C Receptor Structures Reveal the Structural Basis of GPCR Polypharmacology. Cell 2018, 172, 719–730.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Binding Energy (kcal/mol) | Interacting Residues | |
|---|---|---|---|
| Hydrogen Bonds (Å) | Hydrophobic Contacts | ||
| 8-Azanebularine | −6.8 | Thr375 (2.94, 3.14), Ile484 (2.82, 3.2), and Gly487 (3.21) | Lys376, Cys377, Ile378, His394, Lys483, and Glu485 |
| ZINC000095913861 ((2Z)-2,11,28-trimethyl-19-methylidene-13,30- dioxaheptacyclo[21.10.1.06,18.07,15.010,14.024,32.027,31]tetratriaconta- 1(33),2,6(18),7(15),10(14),11,16,23(34),24(32),27(31),28- undecaene-8,9,25,26-tetrone) | −12.0 | Asn379 (2.92) and Gly489 (2.99) | Thr375, Lys376, Cys377, Ile378, His394, Arg455, Ile456, Lys483, Ile484, Glu485, Thr490, and Leu511 |
| ZINC000085996580 (Lespedezol B2 or 8-[[2-(2,4-dihydroxyphenyl)-6-hydroxy-1-benzofuran-3-yl]methyl]-6H-[1]benzofuro[3,2-c]chromene-3,9-diol) | −11.0 | Ile378 (3.01), Arg455 (2.99), Lys483 (3.17), Ile484 (2.55, 2.88), Gly487 (3.05), Leu511 (2.85), Leu512 (3.11), Thr513 (2.97), and Arg590 (2.8) | Val351, Thr375, Lys376, Cys377, His394, Thr448, Cys451, Glu485, and Leu512 |
| ZINC000070454467 ((1S,2S,4R,6S,11R,12S,15S,18S,19S,20R,21S,23R,26S)-15-hydroxy-11,18,21-trimethyl-5,17,24,28,29-pentaoxanonacyclo[17.9.1.11,20.02,12.04,6.06,11.015,19.018,23.021,26]triacont-8-ene-10,16,25,30-tetrone) | −10.9 | His394 (3.15) and Gly487 (2.91) | Thr375, Lys376, Cys377, Lys483, Ile484, Ser486, Gln488, Gly489, Arg590, and Ala595 |
| ZINC000042890265 (Disulfuretin) | −10.6 | Cys377 (3.08), Cys451 (3.15), Gly452 (3.19), Ser449 (2.7), and Arg590 (3.05) | Lys350, Val351, Thr375, Lys376, His394, Glu396, Thr448, Pro450, Arg455, Ile456, Lys483, Ile484, Gly487, Gly489, and Thr490 |
| ZINC000039183320 (Neocalyxin A) | −10.5 | Cys377 (3.01), Ile378 (3.19), Asn379 (3.11), Glu396 (2.77), and Ser449 (3.14) | Val351, Thr375, Lys376, His394, Thr448, Pro450, Cys451, Arg455, Lys483, Ile484, Glu485, Gln488, Gly489, Arg590, and Ala595 |
| ZINC000085593577 ((2S,3R)-2-[2-(4-aminophenyl)ethyl]-3,5-dihydroxy-8-[(1R)-1-hydroxy-2-phenylethyl]-2-methyl-3,4-dihydropyrano[3,2-g]chromen-6-one) | −10.5 | Lys483 (2.97), Ile484 (2.97), Gly487 (2.78), and Leu511 (2.89) | Val351, Thr375, Lys376, Cys377, Ile378, His394, Glu396, Thr448, Ser449, Cys451, Arg455, Glu485, Ser486, Gly489, Thr490, and Thr513 |
| ZINC000070454124 ((3S,10S,11S,12S)-10,11-dihydroxy-7,18-bis(2-phenylethyl)-2,8,13,17-tetraoxapentacyclo[12.8.0.03,12.04,9.016,21]docosa-1(14),4(9),6,15,18,21-hexaene-5,20-dione) | −10.2 | Lys376 (3.08), Cys377 (3.13), Asn379 (3.07), Ile484 (2.94), Ser486 (2.85), and Gly487 (3.27) | Val351, Thr375, His394, Glu396, Thr448, Ser449, Cys451, Arg455, Glu485, Ala595, Asn597, and Thr615 |
| ZINC000103585067 ((1R,2S,5S,8S,9R,17R,18S,21S,24R,26S,27S)-5-hydroxy-2,9,26-trimethyl-3,19,23,28-tetraoxaoctacyclo[16.9.1.118,27.01,5.02,24.08,17.09,14.021,26]nonacosa-11,14-diene-4,10,22,29-tetrone) | −10.2 | Thr375 (2.9), Lys376 (3.29), Cys377 (3.0), Asn379 (3.33), His394 (3.13), Ile484 (3.05), and Gly487 (2.99) | Lys483, Ser486, Gln488, and Thr615 |
| ZINC000014637370 ((8R)-8-(2,2-dimethyl-3,4-dihydrochromen-6-yl)-5-hydroxy-2,2-dimethyl-3,4,7,8-tetrahydropyrano[3,2-g]chromen-6-one) | −10.2 | Cys377 (3.34), Asn379 (2.97), Ser486 (3.31), and Gly489 (3.19) | Thr375, Lys376, His394, Cys451, Lys483, Ile484, Glu485, Gly487, and Thr490 |
| ZINC000013384051 (Cassigarol E) | −10.1 | Asn379 (3.03), Glu396 (3.13), Ser449 (2.79), and Cys451 (2.97) | Val351, Thr375, Cys377, Ile378, His394, Pro450, Arg455, Lys483, Ile484, Glu485, Gln488, Gly489, and Leu511 |
| ZINC000059586224 ((5S)-9-methoxy-14-methyl-5,19-diphenyl-4,12,18-trioxapentacyclo[11.7.1.02,11.03,8.017,21]henicosa-1(21),2,6,8,10,13,16,19-octaen-15-one) | −10.1 | Gly489 (3.07) | Thr375, Lys376, Cys377, Ile378, Asn379, His394, Arg455, Lys483, Ile484, Glu485, Gly487, Gln488, Thr490, Leu511, Asn597, and Thr615 |
| ZINC000070454074 ((1S,2R,7R,10R,13R,14S,16R,19R,20R)-19-[(2S)-2-hydroxy-5-oxo-2H-furan-3-yl]-9,9,13,20-tetramethyl-4,15,18-trioxahexacyclo[11.9.0.02,7.02,10.014,16.014,20]docosane-5,12,17-trione) | −10.1 | His394 (2.97), Arg455 (2.97, 2.96, 3.2) and Arg590 (2.81) | Val351, Thr375, Lys376, Cys377, Cys451, Ile484, Gly487, and Gln488 |
| ZINC000085530502 ((1S,2R,4S,7S,8S,11R,12R,17S,19R,20S,24S)-19-cyclohexyl-7-(furan-3-yl)-24-hydroxy-8,19-dimethyl-3,6,14,18-tetraoxaheptacyclo[18.3.2.01,11.02,4.02,8.012,17.012,20]pentacos-21-ene-5,15,25-trione) | −10.1 | Cys377 (3.22) and Arg455 (3.31) | Thr375, Lys376, His394, Cys451, Lys483, Ile484, Ser486, Gly487, Gln488, Gly489, and Thr490 |
| ZINC000085532258 ((5E)-5-[(1S,2R,3S,11S,13S)-13-benzyl-11-[(S)-hydroxy-[(1S,5R)-5-methylcyclohex-2-en-1-yl]methyl]-3-methyl-5-oxa-10-azatricyclo[8.4.0.02,6]tetradec-6-en-4-ylidene]-3-(hydroxymethyl)-4-methoxyfuran-2-one) | −10.1 | Asn379 (2.92) and Arg455 (3.32) | Val351, Thr375, Cys377, His394, Glu396, Thr448, Ser449, Cys451, Pro459, Lys483, Ile484, Glu485, Ser486, Gly487, Gln488, and Gly489 |
| ZINC000085532442 (5-[(1S,2R,3S,4E,11S,13S)-13-benzyl-11-[(1S)-2- cyclopentyl-1-hydroxyethyl]-3-methyl-5-oxa-10- azatricyclo[8.4.0.02,6]tetradec-6-en-4-ylidene]-3- (hydroxymethyl)-4-methoxy-2,5-dihydrofuran-2-one) | −10.1 | Cys377 (3.04), Ile378 (2.91), and Gly489 (3.05) | Val351, Thr375, Lys376, Asn379, His394, Glu396, Ser449, Cys451, Arg455, Ile456 Lys483, Ile484, Glu485, Ser486, Gln488, and Thr490 |
| ZINC000095911347 ((1R,2S,4R,6S,11R,12S,15R,18S,19R,20S,21S,23R,26S)-15-hydroxy-11,18,21-trimethyl-5,17,24,28,29-pentaoxanonacyclo[17.9.1.11,20.02,12.04,6.06,11.015,19.018,23.021,26]triacont-8-ene-10,16,25,30-tetrone) | −10.1 | Lys376 (3.19), Cys377 (3.28), and Arg455 (2.99) | Thr375, Hs394, Ile484, Ser486, Gly487, Gln488, Gly489, and Ala595 |
| ZINC000095914813 (5-[(Z)-2-[(2S,3S)-3-(3,5-dihydroxyphenyl)-2-(4-hydroxyphenyl)-2,3-dihydro-1-benzofuran-5-yl]ethenyl]benzene-1,3-diol) | −10.1 | Gly374 (3.25), Lys376 (2.9), Cys377 (3.07), Glu396 (2.96), Ser449 (2.54), Cys451 (3.16), and Arg455 (2.8) | Lys350, Val351, Thr375, His394, Thr448, Pro450, Gly487, Gln488, Gly489, Arg590, and Ala595 |
| ZINC000085530478 ((1S,2R,4S,7S,8S,10S,13S,17R,18S,21S,25S,27R)-7-(furan-3-yl)-25-hydroxy-8,20,20-trimethyl-3,6,15,19-tetraoxaoctacyclo[19.3.2.11,10.02,4.02,8.013,18.017,21.017,27]heptacos-22-ene-5,14,26-trione) | −10.0 | Arg455 (2.81) and Gly487 (3.06) | Thr375, Lys376, Cys377, His394, Cys451, Lys483, Ile484, Ser486, and Gln488 |
| ZINC000085530490 ((1R,2R,3’R,7S,9S,10S,12S,13S,14R,16S,19S,20S)-19-(furan-3-yl)-12-hydroxy-13,20-dimethyl-3’-propan-2-ylspiro[4,8,15,18-tetraoxahexacyclo[11.9.0.02,7.02,10.014,16.014,20]docosane-9,1’-cyclohexane]-5,11,17-trione) | −10.0 | Thr375 (3.12) and Lys376 (2.89) | Cys377, His394, Cys451, Arg455, Lys483, Ile484, Ser486, Gly487, Gln488, and Gly489 |
| ZINC000085543539 (3-[[(1R,3R)-3-[(1S,5S)-1,5-dimethylcyclohex-2-en-1-yl]cyclohexyl]methyl]-5-[(1R,4S)-4-(ethylamino)-1,2,3,4-tetrahydronaphthalen-1-yl]phenol) | −10.0 | Ile484 (2.86, 3.2) and Gly487 (2.86) | Val351, Gly374, Thr375, Lys376, Cys377, His394, Ala395, Glu396, Ser449, Cys451, Arg455, Lys483, Gly489, Arg590, and Thr615 |
| ZINC000085592995 ((1R,2R)-2-[(3S,4S)-4-hydroxy-8-[(3-hydroxyphenyl)methyl]-6-methoxy-3,4-dihydro-2H-chromen-3-yl]-1,2,3,8,9,10-hexahydropyrano[3,2-f]chromen-1-ol) | −10.0 | Ser449 (2.55), Ser486 (3.19), and Gly489 (2.88) | Val351, Thr375, Cys377, His394, Glu396, Thr448, Pro450, Cys451, Arg455, Lys483, Ile484, Glu485, Gly487, Gln488, and Arg590 |
| ZINC000085633079 (9-[[(2S,4S)-5,5-dimethyl-4’-(3-methylbut-2-enoxy)spiro[1,3-dioxolane-2,7’-furo[3,2-g]chromene]-4-yl]methoxy]furo[3,2-g]chromen-7-one) | −10.0 | Lys376 (2.98), Cys377 (3.31), and Thr490 (3.07) | Ile378, His394, Arg455, Ile456, Lys483, Ile484, Glu485, Gln488, Gly489, Leu511, and Thr513 |
| ZINC000101100339 (Qingdainone) | −9.7 | Asn379 (3.01) | Lys376, Cys377, Ile378, Lys483, Ile484, Glu485, Gly487, Gly489, Thr490, Leu511, and Thr513 |
| ZINC000085532375 ((5E)-5-[(1S,2R,3S,9S,12S,13S)-12-hydroxy-3-methyl-12-[(1S,5S)-5-methylcyclohex-2-en-1-yl]-5-oxa-17-azatetracyclo[7.7.1.02,6.013,17]heptadec-6-en-4-ylidene]-4-methoxy-3-methylfuran-2-one) | −9.6 | Arg455 (3.11) and Gly487 (3.05) | Thr375, Lys376, Cys377, Asn379, His394, Ile456, Lys483, Glu485, Ser486, Gly489, and Thr490 |
| Compound | MW (g/mol) | Consensus logP o/w | TPSA (Å2) | BBB Permeant | GI Absorption | ESOL Solubility Class | No. of Lipinski’s Rule Violations | No. of Veber’s Rule Violations |
|---|---|---|---|---|---|---|---|---|
| ZINC000095913861 | 556.6 | 5.97 | 94.56 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000085996580 | 508.48 | 4.47 | 136.66 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000070454467 | 526.53 | 4.47 | 137.96 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000042890265 | 538.46 | 5.27 | 173.98 | No | High | Poorly soluble | 1 | 0 |
| ZINC000039183320 | 474.6 | 3.93 | 94.45 | No | High | Moderately soluble | 1 | 0 |
| ZINC000085593577 | 487.54 | 3.55 | 126.15 | No | High | Moderately soluble | 0 | 0 |
| ZINC000070454124 | 564.58 | 3.38 | 119.34 | No | High | Moderately soluble | 1 | 0 |
| ZINC000103585067 | 510.53 | 1.27 | 125.43 | No | High | Soluble | 1 | 0 |
| ZINC000014637370 | 408.49 | 4.58 | 64.99 | Yes | High | Moderately soluble | 0 | 0 |
| ZINC000013384051 | 486.47 | 3.44 | 139.84 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000059586224 | 486.51 | 5.7 | 61.81 | No | Low | Poorly soluble | 0 | 0 |
| ZINC000070454074 | 500.54 | 2.17 | 128.73 | No | High | Soluble | 1 | 0 |
| ZINC000085530502 | 578.65 | 3.31 | 124.8 | No | High | Moderately soluble | 1 | 0 |
| ZINC000085532258 | 561.71 | 4.27 | 88.46 | No | High | Poorly soluble | 1 | 0 |
| ZINC000085532442 | 548.7 | 4.36 | 88.46 | No | High | Poorly soluble | 1 | 0 |
| ZINC000095911347 | 526.53 | 1.37 | 137.96 | No | High | Poorly soluble | 1 | 0 |
| ZINC000095914813 | 454.47 | 4.01 | 110.38 | No | High | Poorly soluble | 0 | 0 |
| ZINC000085530478 | 536.57 | 2.32 | 124.8 | No | High | Moderately soluble | 1 | 0 |
| ZINC000085530490 | 568.65 | 3.35 | 124.8 | No | High | Moderately soluble | 1 | 0 |
| ZINC000085543539 | 471.72 | 7.08 | 32.26 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000085592995 | 490.54 | 3.47 | 97.61 | No | High | Moderately soluble | 0 | 0 |
| ZINC000085633079 | 556.56 | 5.46 | 102.64 | No | Low | Poorly soluble | 1 | 0 |
| ZINC000101100339 | 363.37 | 3.25 | 63.47 | Yes | High | Moderately soluble | 0 | 0 |
| ZINC000085532375 | 481.62 | 3.91 | 68.23 | Yes | High | Moderately soluble | 0 | 0 |
| Compound | vdW | Electrostatic | Polar Solvation | SASA | Binding |
|---|---|---|---|---|---|
| ZINC000042890265 | −185.615 ± 1.989 | −81.409 ± 2.341 | 174.426 ± 3.098 | −24.866 ± 0.150 | −117.236 ± 4.040 |
| ZINC000039183320 | −110.150 ± 2.956 | −23.952 ± 1.891 | 82.789 ± 2.665 | −15.755 ± 0.366 | −67.023 ± 3.022 |
| ZINC000101100339 | −156.644 ± 1.535 | 12.671 ± 1.635 | 95.394 ± 2.438 | −16.419 ± 0.171 | −64.913 ± 2.029 |
| ZINC000014637370 | −218.832 ± 1.310 | −42.038 ± 1.376 | 108.673 ± 1.974 | −22.854 ± 0.099 | −174.911 ± 2.104 |
| ZINC000085532375 | −69.224 ± 5.194 | 289.884 ± 7.174 | 18.341 ± 2.572 | −9.868 ± 0.750 | 228.669 ± 3.288 |
| ZINC000085593577 | −224.512 ± 2.058 | −47.130 ± 1.820 | 158.815 ± 2.933 | −24.585 ± 0.180 | −137.369 ± 2.365 |
| 8-Azanebularine | −49.600 ± 2.917 | 227.826 ± 11.031 | 75.515 ± 7.041 | −7.374 ± 0.412 | 246.374 ± 5.841 |
| 8-Azanebularine rerun | −109.745 ± 1.390 | 310.967 ± 6.418 | 123.630 ± 5.771 | −13.942 ± 0.096 | 310.779 ± 2.145 |
| Compound | Binding Energy | Interacting Residues | |
|---|---|---|---|
| Hydrogen Bonds (Å) | Hydrophobic Bonds | ||
| Ritanserin | −12.7 | Asp134 (2.82) and Tyr358 (2.93) | Ser110, Tyr118, Val135, Ser138, Thr139, Ile142, Val208, Ser219, Ala222, Phe223, Trp324, Phe327, Phe328, Asn351, and Val354 |
| ZINC000095913861 | −12.9 | - | Tyr118, Asp134, Ser138, Val208, Leu209, Val215, Ser219, Ala222, Phe223, Phe327, Phe328, Asn331, Asn351, Val354, and Tyr358 |
| ZINC000101100339 | −12.5 | - | Asp134, Ser138, Leu209, Phe214, Val215, Gly218, Ser219, Ala222, Phe223, Trp324, Phe327, Phe328, and Val354 |
| ZINC000085996580 | −12.1 | Ser110 (2.82), Leu209 (2.81), Ala222 (2.85), Asn331 (3.00), and Asn351 (2.78) | Ile114, Tyr118, Ile131, Asp134, Val135, Thr139, Ile142, Val208, Phe223, Trp324, Phe327, Phe328, Leu350, and Val354, |
| ZINC000085593577 | −11.4 | Asp134 (3.00, 3.06, and 3.07) and Asn331 (3.21) | Ser110, Tyr118, Trp130, Val135, Ser138, Leu209, Phe214, Val215, Ser219, Trp324, Phe327, Phe328, Val354, and Tyr358 |
| ZINC000085532375 | −11.2 | Asn331 (3.06) | Asp134, Val135, Ser138, Val208, Leu209, Val215, Gly218, Ser219, Ala222, Phe223, Phe327, Phe328, Leu350, Asn351, and Val354 |
| ZINC000014637370 | −10.9 | - | Asp134, Val135, Val208, Leu209, Val215, Trp324, Phe327, Phe328, Asn331, Leu350, Asn351, and Val354 |
| ZINC000039183320 | −10.3 | Thr139 (2.83), Leu209 (2.92), and Asn331 (3.30) | Asp134, Val135, Ser138, Val208, Val215, Phe327, Phe328, Glu347, Leu350, Asn351, and Val354 |
| ZINC000070454467 | −8.4 | Leu209 (3.01) | Asp134, Val135, Val208, Trp324, Phe327, Phe328, Asn331, Leu350, and Val354 |
| ZINC000042890265 | −7.8 | Val215 (2.75) and Ser219 (2.52) | Trp130, Asp134, Val135, Ser138, Leu209, Phe214, Phe223, Trp324, Phe327, Phe328, Asn331, Leu350, and Val354 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broni, E.; Ashley, C.; Velazquez, M.; Khan, S.; Striegel, A.; Sakyi, P.O.; Peracha, S.; Bebla, K.; Sodhi, M.; Kwofie, S.K.; et al. In Silico Discovery of Potential Inhibitors Targeting the RNA Binding Loop of ADAR2 and 5-HT2CR from Traditional Chinese Natural Compounds. Int. J. Mol. Sci. 2023, 24, 12612. https://doi.org/10.3390/ijms241612612
Broni E, Ashley C, Velazquez M, Khan S, Striegel A, Sakyi PO, Peracha S, Bebla K, Sodhi M, Kwofie SK, et al. In Silico Discovery of Potential Inhibitors Targeting the RNA Binding Loop of ADAR2 and 5-HT2CR from Traditional Chinese Natural Compounds. International Journal of Molecular Sciences. 2023; 24(16):12612. https://doi.org/10.3390/ijms241612612
Chicago/Turabian StyleBroni, Emmanuel, Carolyn Ashley, Miriam Velazquez, Sufia Khan, Andrew Striegel, Patrick O. Sakyi, Saqib Peracha, Kristeen Bebla, Monsheel Sodhi, Samuel K. Kwofie, and et al. 2023. "In Silico Discovery of Potential Inhibitors Targeting the RNA Binding Loop of ADAR2 and 5-HT2CR from Traditional Chinese Natural Compounds" International Journal of Molecular Sciences 24, no. 16: 12612. https://doi.org/10.3390/ijms241612612
APA StyleBroni, E., Ashley, C., Velazquez, M., Khan, S., Striegel, A., Sakyi, P. O., Peracha, S., Bebla, K., Sodhi, M., Kwofie, S. K., Ademokunwa, A., & Miller, W. A., III. (2023). In Silico Discovery of Potential Inhibitors Targeting the RNA Binding Loop of ADAR2 and 5-HT2CR from Traditional Chinese Natural Compounds. International Journal of Molecular Sciences, 24(16), 12612. https://doi.org/10.3390/ijms241612612