
Molecular Interplay between Non-Host Resistance, Pathogens and Basal Immunity as a Background for Fatal Yellowing in Oil Palm (Elaeis guineensis Jacq.) Plants

 , , and
, , and 
Abstract
:1. Introduction
2. Results
2.1. Soil Physicochemical and Leaf Chemical Analysis
2.2. Metabolomics Analysis
2.3. Transcriptomics Analysis
2.4. Multi-Omics Integration Analysis
3. Discussion
4. Materials and Methods
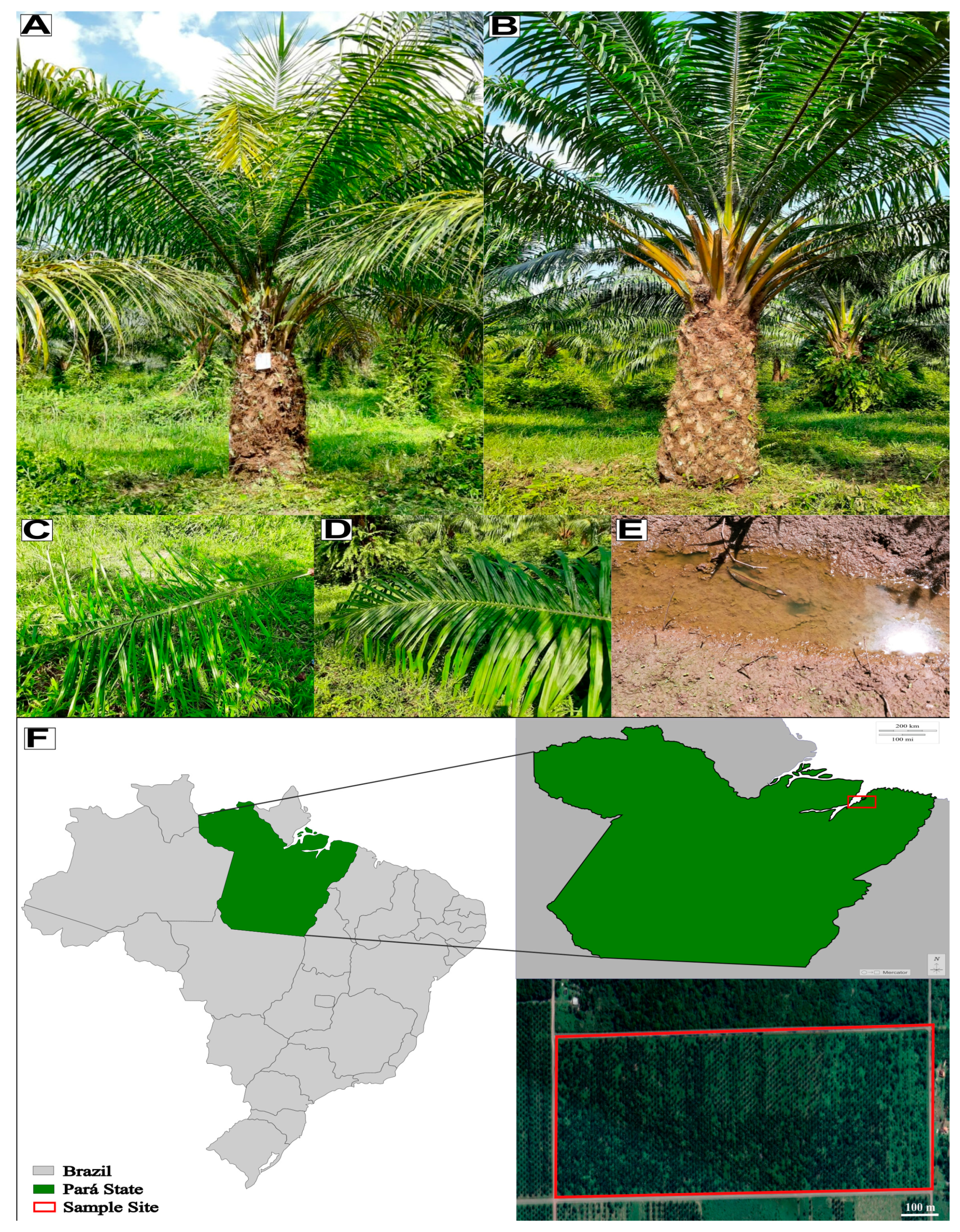
4.1. Soil and Leaf Samples—Collection and Chemical and Physicochemical Analysis
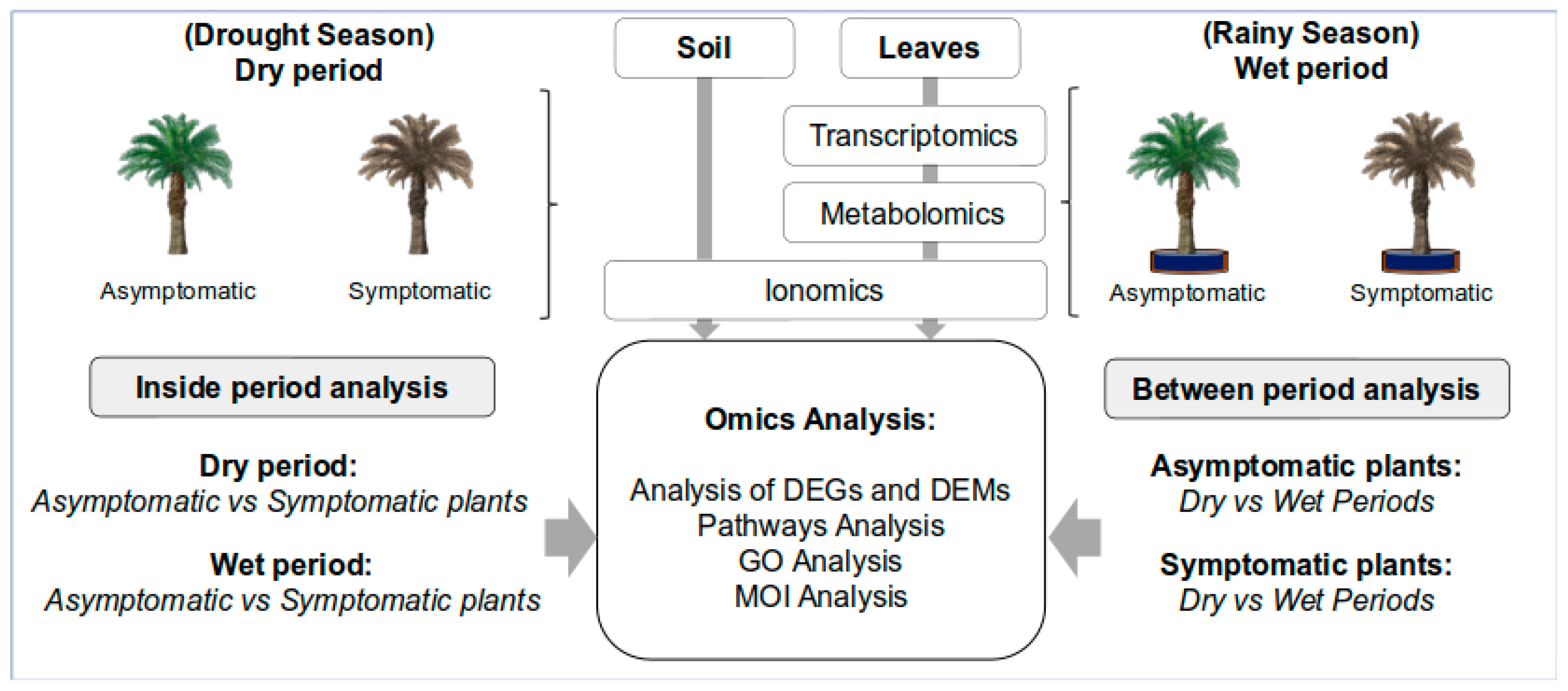
4.2. Experimental Design and Statistical Analysis
4.3. Transcriptomics Analysis
4.4. Metabolomics Analysis
4.5. Correlation and Integratomics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statista. 2023. Available online: www.statista.com (accessed on 30 June 2023).
- Murphy, D.J.; Goggin, K.; Paterson, R.R.M. Oil palm in the 2020s and beyond: Challenges and solutions. CABI Agric. Biosci. 2021, 2, 39. [Google Scholar] [CrossRef]
- Ritchie, H.; Roser, M. Forests and Deforestation. Published Online at OurWorldInData.org. 2021. Available online: https://ourworldindata.org/forests-and-deforestation (accessed on 13 July 2023).
- MAPA—Ministério da Agricultura, Pecuária e Abastecimento. Diagnóstico da Produção Sustentável da Palma de Óleo. 2018. Available online: http://www.abrapalma.org/pt/wp-content/uploads/2018/06/DIAGNOSTICO_PALMA1.pdf (accessed on 30 June 2023).
- Bertone, M.V. A Importância do Programa de Produção Sustentável de Palma de Óleo: Produtividade e Sustentabilidade. 2011. Available online: https://issuu.com/embrapa/docs/revista_agroenergia_ed2 (accessed on 30 June 2023).
- Bittencourt, C.B.; Lins, P.d.C.; Boari, A.d.J.; Quirino, B.F.; Teixeira, W.G.; Souza Junior, M.T. Oil Palm Fatal Yellowing (FY), a Disease with an Elusive Causal Agent. In Elaeis Guineensis; Kamyab, H., Ed.; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Boari, A.d.J. Estudos Realizados Sobre o Amarelecimento Fatal do Dendezeiro (Elaeis Guineensis Jacq.) No Brasil. 2008. Available online: https://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/410160 (accessed on 30 June 2023).
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar] [CrossRef]
- Laranjeira, F.F.; Amorin, L.; Bergamin Filho, A.; Berger, R.D.; Hau, B. Análise espacial do Amarelecimento Fatal do dendezeiro como ferramenta para elucidar sua etiologia. Fitopatol. Bras. 1998, 23, 397–403. [Google Scholar]
- Silveira, R.I.; Veiga, A.S.; Ramos, E.J.A.; Parente, J.R. Evolução da Sintomatologia do Amarelecimento Fatal a Adubações com Omissão de Macro e Micronutrientes; Denpasa: Belém, PA, Brazil, 2000; p. 35. [Google Scholar]
- Viégas, I.d.J.M.; Frazão, D.A.C.; Júnior, J.F.; Trindade, D.R.; Thomazl, M.A.A. Teores de Micronutrientes em Folhas de Dendezeiros Sadios e com Sintomas de Amarelecimento Fatal. XXV Reunião Brasileira de Fertilidade Do Solo e Nutrição de Plantas. Available online: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/100995/1/4469.pdf (accessed on 30 June 2023).
- Muniz, R.S. Alterações do Fluxo Hídrico e Seus Efeitos na Dinâmica do Ferro e na Estrutura de um Latossolo Amarelo na Amazônia; Universidade Federal do Rio de Janeiro: Rio de Janeiro, Brazil, 2017; Available online: https://pantheon.ufrj.br/handle/11422/9680 (accessed on 30 June 2023).
- Costa, O.Y.d.A.; Tupinambá, D.D.; Bergmann, J.C.; Barreto, C.C.; Quirino, B.F. Fungal diversity in oil palm leaves showing symptoms of Fatal Yellowing disease. PLoS ONE 2018, 13, e0191884. [Google Scholar] [CrossRef]
- Rodrigues-Neto, J.C.; Correia, M.V.; Souto, A.L.; Ribeiro, J.A.A.; Vieira, L.R.; Souza, M.T., Jr.; Rodrigues, C.M.; Abdelnur, P.V. Metabolic fingerprinting analysis of oil palm reveals a set of differentially expressed metabolites in fatal yellowing symptomatic and non-symptomatic plants. Metabolomics Off. J. Metabolomic Soc. 2018, 14, 142. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, S.V.D.; Magalhães, M.M.; Cunha, R.L.; Costa, P.H.O.; Alves, R.C.O.; Oliveira, G.C.; Valadares, R.B.D.S. Differential accumulation of proteins in oil palms affected by fatal yellowing disease. PLoS ONE 2018, 13, e0195538. [Google Scholar] [CrossRef] [PubMed]
- Torres, G.A.; Sarria, G.A.; Varon, F.; Coffey, M.D.; Elliott, M.L.; Martinez, G. First Report of Bud Rot Caused by Phytophthora palmivora on African Oil Palm in Colombia. Plant Dis. 2010, 94, 1163. [Google Scholar] [CrossRef]
- Torres, G.A.; Sarria, G.A.; Martinez, G.; Varon, F.; Drenth, A.; Guest, D.I. Bud Rot Caused by Phytophthora palmivora: A Destructive Emerging Disease of Oil Palm. Phytopathology 2016, 106, 320–329. [Google Scholar] [CrossRef]
- Reddy, K.R.; Patrick, W.H. Effect of alternate aerobic and anaerobic conditions on redox potential, organic matter decomposition and nitrogen loss in a flooded soil. Soil Biol. Biochem. 1975, 7, 87–94. [Google Scholar] [CrossRef]
- Favre, F.; Tessier, D.; Abdelmoula, M.; Génin, J.M.; Gates, W.P.; Boivin, P. Iron reduction and changes in cation exchange capacity in intermittently waterlogged soil. Eur. J. Soil Sci. 2002, 53, 175–183. [Google Scholar] [CrossRef]
- Bittencourt, C.B.; Carvalho da Silva, T.L.; Rodrigues Neto, J.C.; Vieira, L.R.; Leão, A.P.; de Aquino Ribeiro, J.A.; Abdelnur, P.V.; de Sousa, C.A.F.; Souza, M.T., Jr. Insights from a Multi-Omics Integration (MOI) Study in Oil Palm (Elaeis guineensis Jacq.) Response to Abiotic Stresses: Part One-Salinity. Plants 2022, 11, 1755. [Google Scholar] [CrossRef]
- Gertsman, I.; Barshop, B.A. Promises and pitfalls of untargeted metabolomics. J. Inherit. Metab. Dis. 2018, 41, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Sudheer, S.; Usmani, Z.; Rani, R.; Gupta, P. Deciphering the Omics of Plant-Microbe Interaction: Perspectives and New Insights. Curr. Genom. 2020, 21, 343–362. [Google Scholar] [CrossRef] [PubMed]
- Dev Sareshma, S.; Subha, B. Assessing Host-Pathogen Interaction Networks via RNA-Seq Profiling: A Systems Biology Approach; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Zheng, S.; Terauchi, R. Plant–Pathogen Interaction: New Era of Plant–Pathogen Interaction Studies: “Omics” Perspectives. In Plant Omics: Advances in Big Data Biology; CABI Biotechnology Series; CABI International: Wallingford, UK, 2022. [Google Scholar] [CrossRef]
- Liu, Z.; Pan, Y.; Li, Y.; Ouellet, T.; Foroud, N.A. RNA-Seq Data Processing in Plant-Pathogen Interaction System: A Case Study. Methods Mol. Biol. 2023, 2659, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Gregory, B.; Rahman, N.; Bommakanti, A.; Shamsuzzaman, M.; Thapa, M.; Lescure, A.; Zengel, J.M.; Lindahl, L. The small and large ribosomal subunits depend on each other for stability and accumulation. Life Sci. Alliance 2019, 2, e201800150. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Senthil-Kumar, M.; Ramu, V.S.; Wang, K.; Mysore, K.S. Plant Ribosomal Proteins, RPL12 and RPL19, Play a Role in Nonhost Disease Resistance against Bacterial Pathogens. Front. Plant Sci. 2016, 6, 1192. [Google Scholar] [CrossRef]
- Ramu, V.S.; Dawane, A.; Lee, S.; Oh, S.; Lee, H.K.; Sun, L.; Senthil-Kumar, M.; Mysore, K.S. Ribosomal protein QM/RPL10 positively regulates defence and protein translation mechanisms during nonhost disease resistance. Mol. Plant Pathol. 2020, 21, 1481–1494. [Google Scholar] [CrossRef]
- Ayliffe, M.; Sørensen, C.K. Plant nonhost resistance: Paradigms and new environments. Curr. Opin. Plant Biol. 2019, 50, 104–113. [Google Scholar] [CrossRef]
- Panstruga, R.; Moscou, M.J. What is the Molecular Basis of Nonhost Resistance? Mol. Plant Microbe Interact. 2020, 33, 1253–1264. [Google Scholar] [CrossRef]
- Wu, Y.; Sexton, W.; Yang, B.; Xiao, S. Genetic approaches to dissect plant nonhost resistance mechanisms. Mol. Plant Pathol. 2023, 24, 272–283. [Google Scholar] [CrossRef]
- Semrad, K.; Green, R.; Schroeder, R. RNA chaperone activity of large ribosomal subunit proteins from Escherichia coli. RNA 2004, 10, 1855–1860. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, D.; Rakacs, M.; Agoston, B.; Lenkey, K.; Semrad, K.; Schroeder, R.; Tompa, P. Janus chaperones: Assistance of both RNA- and protein-folding by ribosomal proteins. FEBS Lett. 2009, 583, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Gorelova, V.; De Lepeleire, J.; Van Daele, J.; Pluim, D.; Meï, C.; Cuypers, A.; Leroux, O.; Rébeillé, F.; Schellens, J.H.M.; Blancquaert, D.; et al. Dihydrofolate Reductase/Thymidylate Synthase Fine-Tunes the Folate Status and Controls Redox Homeostasis in Plants. Plant Cell 2017, 29, 2831–2853. [Google Scholar] [CrossRef]
- Singh, R.; Ong-Abdullah, M.; Low, E.T.; Manaf, M.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.; Ooi, S.E.; Chan, K.L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in Old and New worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Carvalho da Silva, T.L.; The Brazilian Agricultural Research Corporation, Embrapa Agroenergy, Brasília, DF, Brazil; Grynberg, P.; The Brazilian Agricultural Research Corporation, Embrapa Agroenergy, Brasília, DF, Brazil; Togawa, R.C.; The Brazilian Agricultural Research Corporation, Embrapa Agroenergy, Brasília, DF, Brazil; Souza, M.T., Jr.; The Brazilian Agricultural Research Corporation, Embrapa Agroenergy, Brasília, DF, Brazil. Unpublished work, 2023.
- McCoy, R.M.; Julian, R.; Kumar, S.R.V.; Ranjan, R.; Varala, K.; Li, Y. A Systems Biology Approach to Identify Essential Epigenetic Regulators for Specific Biological Processes in Plants. Plants 2021, 10, 364. [Google Scholar] [CrossRef]
- Souza, M.T., Jr.; The Brazilian Agricultural Research Corporation, Embrapa Agroenergy, Brasília, DF, Brazil. Unpublished work, 2023.
- Qi, H.; Zhu, X.; Guo, F.; Lv, L.; Zhang, Z. The Wall-Associated Receptor-Like Kinase TaWAK7D Is Required for Defense Responses to Rhizoctonia cerealis in Wheat. Int. J. Mol. Sci. 2021, 22, 5629. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, J.; Liu, H.; Ou, M.; Ye, H.; Zhao, P. Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica. Genes 2022, 13, 134. [Google Scholar] [CrossRef]
- Boutrot, F.; Zipfel, C. Function, Discovery, and Exploitation of Plant Pattern Recognition Receptors for Broad-Spectrum Disease Resistance. Annu. Rev. Phytopathol. 2017, 55, 257–286. [Google Scholar] [CrossRef]
- Shingaki-Wells, R.; Millar, A.H.; Whelan, J.; Narsai, R. What happens to plant mitochondria under low oxygen? An omics review of the responses to low oxygen and reoxygenation. Plant Cell Environ. 2014, 37, 2260–2277. [Google Scholar] [CrossRef]
- León, J.; Castillo, M.C.; Gayubas, B. The hypoxia-reoxygenation stress in plants. J. Exp. Bot. 2021, 72, 5841–5856. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.A.F.; Sodek, L. The metabolic response of plants to oxygen deficiency. Braz. J. Plant Physiol. 2002, 14, 83–94. [Google Scholar] [CrossRef]
- Fukao, T.; Bailey-Serres, J. Plant responses to hypoxia—Is survival a balancing act? Trends Plant Sci. 2004, 9, 449–456. [Google Scholar] [CrossRef]
- Souza, R.; Veiga, A.; Ramos, E. Amarelecimento Fatal do Dendezeiro: Identificação Prática; Denpasa: Belém, PA, Brazil, 2000; p. 27. [Google Scholar]
- OmicsBox—Bioinformatics Made Easy, BioBam Bioinformatics, 3 March 2019. Available online: https://www.biobam.com/omicsbox (accessed on 18 July 2023).
- Andrews, S. FastQC: A Quality Control Tool for High throughput Sequence Data. 2010. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 18 July 2023).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Vargas, L.H.G.; Neto, J.C.R.; de Aquino Ribeiro, J.A.; Ricci-Silva, M.E.; Souza, M.T., Jr.; Rodrigues, C.M.; de Oliveira, A.E.; Abdelnur, P.V. Metabolomics analysis of oil palm (Elaeis guineensis) leaf: Evaluation of sample preparation steps using, UHPLC–MS/MS. Metabolomics 2016, 12, 153. [Google Scholar] [CrossRef]
- Neto, J.C.R.; Vieira, L.R.; de Aquino Ribeiro, J.A.; de Sousa, C.A.F.; Júnior, M.T.S.; Abdelnur, P.V. Metabolic effect of drought stress on the leaves of young oil palm (Elaeis guineensis) plants using UHPLC-MS and multivariate analysis. Sci. Rep. 2021, 11, 18271. [Google Scholar] [CrossRef]
- Gowda, H.; Ivanisevic, J.; Johnson, C.H.; Kurczy, M.E.; Benton, H.P.; Rinehart, D.; Nguyen, T.; Ray, J.; Kuehl, J.; Arevalo, B.; et al. Interactive XCMS Online: Simplifying Advanced Metabolomic Data Processing and Subsequent Statistical Analyses. Anal. Chem. 2014, 86, 6931–6939. [Google Scholar] [CrossRef]
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A web-based platform to process untargeted metabolomic data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Wold, S. Introduction to Multi-and Megavariate Data Analysis Using Projection Methods (PCA & PLS); Umetrics: Umeå, Sweden, 1999. [Google Scholar]
- Li, S.; Park, Y.; Duraisingham, S.; Strobel, F.H.; Khan, N.; Soltow, Q.A.; Jones, D.P.; Pulendran, B. Predicting network activity from high throughput metabolomics. PLoS Comput. Biol. 2013, 9, e1003123. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef]
- Brink, B.G.; Seidel, A.; Kleinbölting, N.; Nattkemper, T.W.; Albaum, S.P. Omics Fusion—A Platform for Integrative Analysis of Omics Data. J. Integr. Bioinform. 2016, 13, 296. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Query Mass | Matched Compound | Matched Form | Mass Diff | Compound Name | Log2(FC) | FDR | Profile |
|---|---|---|---|---|---|---|---|
| 792.12440 | C00024 | M-H2O+H[1+] | 1.95 × 10−3 | Acetyl-CoA | −5.37 | 0.0004 | Down |
| 742.22112 | C03541 | M+K[1+] | 2.18 × 10−3 | THF-polyglutamate | −5.17 | 0.0012 | Down |
| 293.21349 | C06427 | M-H+O[-] | 1.28 × 10−3 | alpha-Linolenic acid | −1.11 | 0.0026 | Down |
| 312.16523 | C16448 | M-C3H4O2+H[1+] | 1.43 × 10−3 | Dihydrozeatin-O-glucoside | 1.93 | 0.0111 | Up |
| 836.28348 | C05275 | M-HCOOK+H[1+] | 2.18 × 10−3 | trans-Dec-2-enoyl-CoA | −2.53 | 0.0220 | Down |
| 409.38261 | C01054 | M-H2O+H[1+] | 2.33 × 10−4 | (S)-2,3-Epoxysqualene | −4.13 | 0.0305 | Down |
| 425.37851 | C22116 | M-HCOOH+H[1+] | 6.34 × 10−4 | 3beta-Hydroxy-4beta | −2.62 | 0.0305 | Down |
| 309.20812 | C04785 | M-H[-] | 9.86 × 10−4 | (9Z,11E,15Z)-(13S)-Hydroperoxyoctadeca-9,11,15-trienoate | −1.33 | 0.0346 | Down |
| 361.20077 | C18016 | M+HCOO[-] | 7.15 × 10−4 | 3beta-Hydroxy-9beta-pimara-7,15-diene-19,6beta-olide | −1.20 | 0.0346 | Down |
| 426.38263 | C22121 | M(C13)+H[1+] | 1.45 × 10−3 | Cycloeucalenone | −2.26 | 0.0359 | Down |
| 407.36819 | C03313 | M-HCOOH+H[1+] | 8.83 × 10−4 | Phylloquinol | −1.90 | 0.0372 | Down |
| 87.00852 | C00258 | M-H2O-H[-] | 2.12 × 10−4 | D-Glycerate | 0.75 | 0.0384 | Up |
| 446.16191 | C00101 | M(Cl37)-H[-] | 1.00 × 10−3 | Tetrahydrofolate | −0.64 | 0.0398 | Down |
| 129.01926 | C06032 | M-H2O-H[-] | 3.35 × 10−5 | D-erythro-3-Methylmalate | 1.00 | 0.0401 | Up |
| 173.00911 | C00311 | M-H2O-H[-] | 1.61 × 10−5 | Isocitrate | 1.00 | 0.0447 | Up |
| Pathway | Pathway ID | Common (Symptomatic and Asymptomatic) | Only in Symptomatic | ||||
|---|---|---|---|---|---|---|---|
| Enzymes & Metabolites | Enzymes | Metabolites | Enzymes & Metabolites | Enzymes | Metabolites | ||
| Purine metabolism | 230 | 32 | 15 | 17 | 9 | 4 | 5 |
| Porphyrin and chlorophyll metabolism | 860 | 29 | 10 | 19 | 4 | 3 | 1 |
| Phenylpropanoid biosynthesis | 940 | 20 | 4 | 16 | 4 | 2 | 2 |
| Starch and sucrose metabolism | 500 | 19 | 17 | 2 | 5 | 4 | 1 |
| Glycolysis/Gluconeogenesis | 10 | 17 | 14 | 3 | 12 | 9 | 3 |
| Carbon fixation pathways in prokaryotes | 720 | 17 | 5 | 12 | 8 | 2 | 6 |
| Cysteine and methionine metabolism | 270 | 16 | 8 | 8 | 10 | 7 | 3 |
| Ubiquinone and other terpenoid-quinone biosynthesis | 130 | 16 | 2 | 14 | 5 | 1 | 4 |
| Pentose phosphate pathway | 30 | 15 | 9 | 6 | 8 | 5 | 3 |
| Aminoacyl-tRNA biosynthesis | 970 | 14 | 12 | 2 | 3 | 3 | 0 |
| Methane metabolism | 680 | 14 | 8 | 6 | 10 | 7 | 3 |
| Glyoxylate and dicarboxylate metabolism | 630 | 14 | 7 | 7 | 5 | 3 | 2 |
| Pyruvate metabolism | 620 | 13 | 8 | 5 | 7 | 5 | 2 |
| Glycerophospholipid metabolism | 564 | 12 | 10 | 2 | 5 | 4 | 1 |
| Glutathione metabolism | 480 | 12 | 7 | 5 | 7 | 5 | 2 |
| Citrate cycle (TCA cycle) | 20 | 12 | 7 | 5 | 4 | 3 | 1 |
| Glycine, serine and threonine metabolism | 260 | 12 | 7 | 5 | 4 | 3 | 1 |
| Galactose metabolism | 52 | 11 | 6 | 5 | 5 | 4 | 1 |
| Pyrimidine metabolism | 240 | 11 | 4 | 7 | 4 | 1 | 3 |
| Carotenoid biosynthesis | 906 | 11 | 0 | 11 | 6 | 0 | 6 |
| Flavonoid biosynthesis | 941 | 11 | 0 | 11 | 4 | 0 | 4 |
| Amino sugar and nucleotide sugar metabolism | 520 | 10 | 9 | 1 | 5 | 4 | 1 |
| Carbon fixation in photosynthetic organisms | 710 | 10 | 7 | 3 | 7 | 5 | 2 |
| Sulfur metabolism | 920 | 10 | 6 | 4 | 4 | 1 | 3 |
| Terpenoid backbone biosynthesis | 900 | 10 | 5 | 5 | 4 | 2 | 2 |
| Steroid biosynthesis | 100 | 10 | 1 | 9 | 8 | 1 | 7 |
| Biosynthesis of various secondary metabolites—part 2 | 998 | 10 | 0 | 10 | 2 | 0 | 2 |
| Protein ID | UniProt Accession | EC Number | FC Symptomatic | Profile Symptomatic | FC Asymptomatic | Profile Asymptomatic |
|---|---|---|---|---|---|---|
| XP_010912022.1 | A0A6I9QPT3 | 1.17.4.1 | −4.0 | DOWN | −2.3 | DOWN |
| XP_010938967.1 | A0A6I9S8I9 | 2.7.1.25 | −2.5 | DOWN | −3.0 | DOWN |
| XP_010911123.2 | A0A6I9QKC5 | 2.7.1.40 | −5.9 | DOWN | −3.6 | DOWN |
| XP_010930617.1 | A0A6I9RQ67 | 2.7.1.40 | −1.8 | DOWN | −2.8 | DOWN |
| XP_010919863.2 | A0A6I9R3I3 | 2.7.1.40 | −2.9 | DOWN | −2.2 | DOWN |
| XP_010924524.1 | A0A6I9RE71 | 2.7.4.6 | −2.5 | DOWN | −2.7 | DOWN |
| XP_010937073.1 | A0A6I9S4K9 | 2.7.4.8 | −4.2 | DOWN | −3.3 | DOWN |
| XP_010910297.1 | A0A6I9QJ47 | 2.7.6.5 | −4.0 | DOWN | −3.6 | DOWN |
| XP_010933384.1 | A0A6I9RX86 | 2.7.6.5 | −2.6 | DOWN | −2.9 | DOWN |
| XP_010932410.1 | A0A6I9RU27 | 2.7.6.5 | −2.8 | DOWN | −2.7 | DOWN |
| XP_010921622.1 | A0A6I9R798 | 2.7.6.5 | −4.9 | DOWN | −5.4 | DOWN |
| XP_029119510.1 | A0A8N4F2W4 | 2.7.7.4 | −3.6 | DOWN | −4.7 | DOWN |
| XP_010932834.1 | A0A6I9RV40 | 2.7.7.4 | −2.2 | DOWN | −2.4 | DOWN |
| XP_010920819.1 | A0A6I9R728 | 3.5.4.6 | −3.3 | DOWN | −3.7 | DOWN |
| XP_010937877.2 | A0A6I9S697 | 3.5.4.6 | −4.6 | DOWN | −2.2 | DOWN |
| XP_029116569.1 | A0A8N4EWM4 | 5.4.2.2 | −2.0 | DOWN | −3.0 | DOWN |
| XP_010934074.1 | A0A6I9RXY5 | 5.4.2.2 | −1.9 | DOWN | −2.1 | DOWN |
| XP_010911922.1 | A0A6I9QMW6 | 2.4.2.7 | 1.8 | UP | −1.3 | NDE |
| XP_010920467.1 | A0A6I9R4T4 | 2.7.1.20 | 1.9 | UP | 1.2 | NDE |
| XP_029117373.1 | A0A8N4ID85 | 2.7.4.6 | 1.5 | UP | −1.2 | NDE |
| XP_010933513.1 | A0A6I9RXJ3 | 2.7.1.40 | −6.4 | DOWN | No | No |
| XP_010905734.1 | A0A6I9QAR4 | 1.7.3.3 | 2.1 | UP | 1.6 | UP |
| XP_010907802.1 | A0A6I9QFG8 | 1.7.3.3 | 3.2 | UP | 2.8 | UP |
| XP_010941354.1 | A0A6I9SCA5 | 2.7.4.3 | 1.7 | UP | 1.7 | UP |
| XP_010908713.1 | A0A6I9QHG6 | 2.7.4.3 | 1.8 | UP | 1.9 | UP |
| XP_010935173.1 | A0A6I9RZH1 | 2.7.4.3 | 4.6 | UP | 3.9 | UP |
| XP_010919758.1 | A0A6I9R9M7 | 2.7.4.6 | 2.4 | UP | 1.5 | UP |
| XP_010933580.1 | A0A6I9RXP4 | 2.7.4.6 | 3.6 | UP | 2.6 | UP |
| XP_010943858.1 | A0A6I9SHJ3 | 2.7.6.5 | 3.9 | UP | 2.8 | UP |
| XP_010914531.2 | A0A6I9QT08 | 6.3.3.1 | 1.7 | UP | 1.8 | UP |
| XP_010910143.1 | A0A6I9QKR6 | 6.3.4.13 | 2.2 | UP | 1.9 | UP |
| KEGG ID | Compound | Matched Form Symptomatic | Fold Change Asymptomatic | Profile Asymptomatic | Fold Change Symptomatic | Profile Symptomatic |
|---|---|---|---|---|---|---|
| C00104 | IDP | M-HCOOK+H[1+] | 0.07 | DOWN | 87.59 | UP |
| C06197 | P1,P3-Bis(5′-adenosyl) triphosphate | M+NaCl[1+] | 0.16 | DOWN | 0.05 | DOWN |
| C00212 | Adenosine | M+Cl[-] | 0.30 | DOWN | 4.97 | UP |
| C00387 | Guanosine | M+Na[1+] | 0.52 | DOWN | 2.54 | UP |
| C04640 | 2-(Formamido)-N1-(5′-phosphoribosyl) acetamidine | M+HCOONa[1+] | 0.27 | DOWN | 0.20 | DOWN |
| C12248 | 5-Hydroxy-2-oxo-4-ureido-2,5-dihydro 1H-imidazole-5-carboxylate | M[1+] | 0.42 | DOWN | 2.88 | UP |
| C00242 | Guanine | M+Na[1+] | 0.12 | DOWN | 9.74 | UP |
| C00224 | Adenylyl sulfate | M-NH3+H[1+] | 0.19 | DOWN | 2.74 | UP |
| C00655 | Xanthosine 5′-phosphate | M+NaCl[1+] | 0.37 | DOWN | 2.98 | UP |
| C04823 | 1-(5′-Phosphoribosyl)-5-amino-4 (N-succinocarboxamide)-imidazole | M-HCOOH+H[1+] | 0.20 | DOWN | 7.89 | UP |
| C00301 | ADP-ribose | M-H2O-H[-] | No | No | 0.22 | DOWN |
| C00385 | Xanthine | M+Na-2H[-] | No | No | 7.90 | UP |
| C00206 | dADP | M+3H[3+] | No | No | 14.49 | UP |
| C02091 | (S)-Ureidoglycine | M[1+] | No | No | 2.51 | UP |
| C00059 | Sulfate | M(S34)-H[-] | No | No | 2.27 | UP |
| C04677 | 1-(5′-Phosphoribosyl)-5-amino-4 imidazolecarboxamide | M-H[-] | 10.95 | UP | 66.60 | UP |
| C00130 | IMP | M-H4O2+H[1+] | 2.93 | UP | 112.58 | UP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bittencourt, C.B.; Carvalho da Silva, T.L.; Rodrigues Neto, J.C.; Leão, A.P.; de Aquino Ribeiro, J.A.; Maia, A.d.H.N.; de Sousa, C.A.F.; Quirino, B.F.; Souza Júnior, M.T. Molecular Interplay between Non-Host Resistance, Pathogens and Basal Immunity as a Background for Fatal Yellowing in Oil Palm (Elaeis guineensis Jacq.) Plants. Int. J. Mol. Sci. 2023, 24, 12918. https://doi.org/10.3390/ijms241612918
Bittencourt CB, Carvalho da Silva TL, Rodrigues Neto JC, Leão AP, de Aquino Ribeiro JA, Maia AdHN, de Sousa CAF, Quirino BF, Souza Júnior MT. Molecular Interplay between Non-Host Resistance, Pathogens and Basal Immunity as a Background for Fatal Yellowing in Oil Palm (Elaeis guineensis Jacq.) Plants. International Journal of Molecular Sciences. 2023; 24(16):12918. https://doi.org/10.3390/ijms241612918
Chicago/Turabian StyleBittencourt, Cleiton Barroso, Thalliton Luiz Carvalho da Silva, Jorge Cândido Rodrigues Neto, André Pereira Leão, José Antônio de Aquino Ribeiro, Aline de Holanda Nunes Maia, Carlos Antônio Ferreira de Sousa, Betania Ferraz Quirino, and Manoel Teixeira Souza Júnior. 2023. "Molecular Interplay between Non-Host Resistance, Pathogens and Basal Immunity as a Background for Fatal Yellowing in Oil Palm (Elaeis guineensis Jacq.) Plants" International Journal of Molecular Sciences 24, no. 16: 12918. https://doi.org/10.3390/ijms241612918
APA StyleBittencourt, C. B., Carvalho da Silva, T. L., Rodrigues Neto, J. C., Leão, A. P., de Aquino Ribeiro, J. A., Maia, A. d. H. N., de Sousa, C. A. F., Quirino, B. F., & Souza Júnior, M. T. (2023). Molecular Interplay between Non-Host Resistance, Pathogens and Basal Immunity as a Background for Fatal Yellowing in Oil Palm (Elaeis guineensis Jacq.) Plants. International Journal of Molecular Sciences, 24(16), 12918. https://doi.org/10.3390/ijms241612918