
The Role of Gender Differences and Menopause in Obesity-Related Renal Disease, Renal Inflammation and Lipotoxicity
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Inflammation in Renal Tissue
2.1.1. Inflammation in Male Mice
2.1.2. Inflammation in Female Mice
2.2. Lipidomic Analysis in Renal Tissue
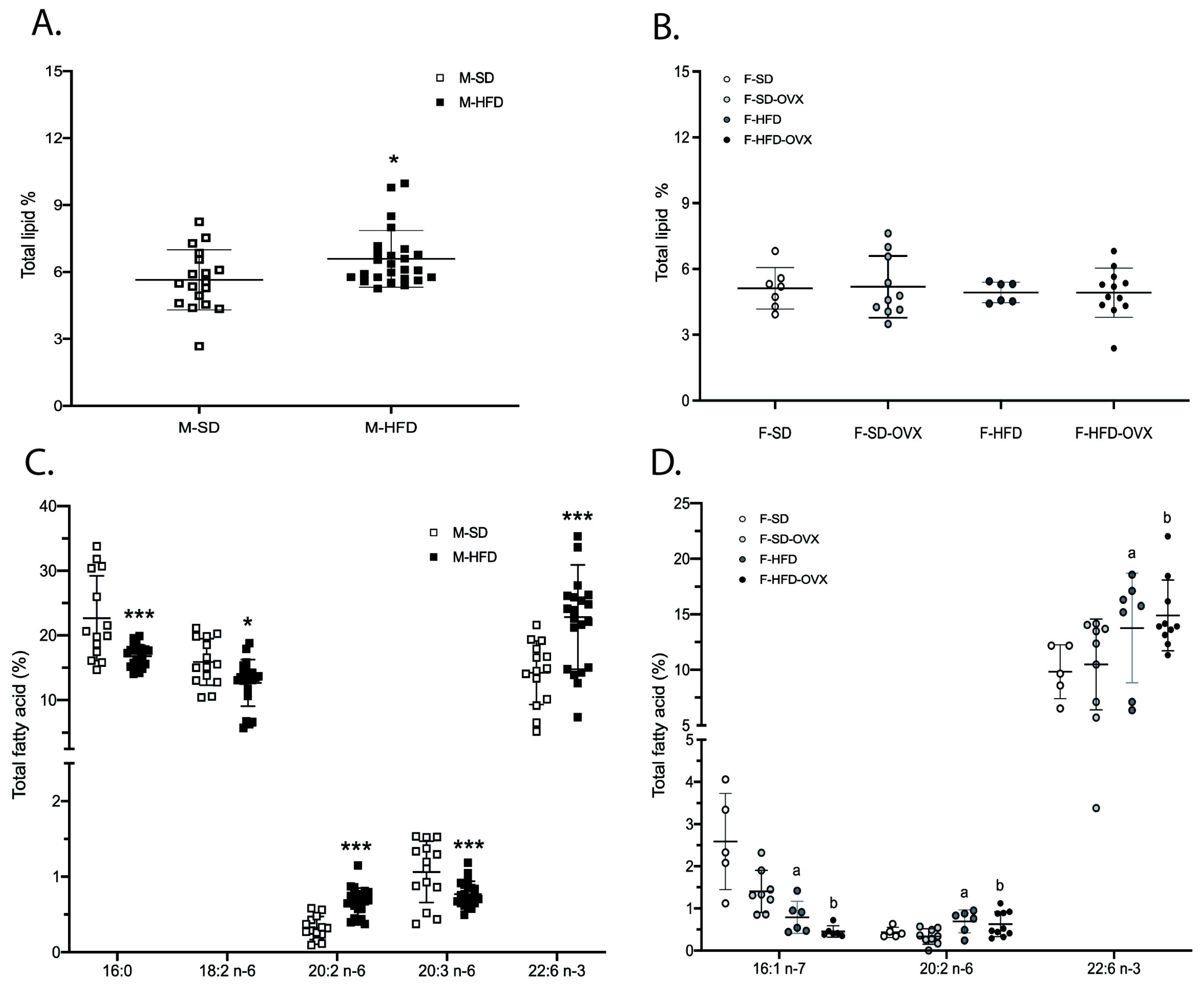
2.2.1. Total Lipid Content and Lipid Classes Profile
2.2.2. Fatty Acids Profile in Total Lipids
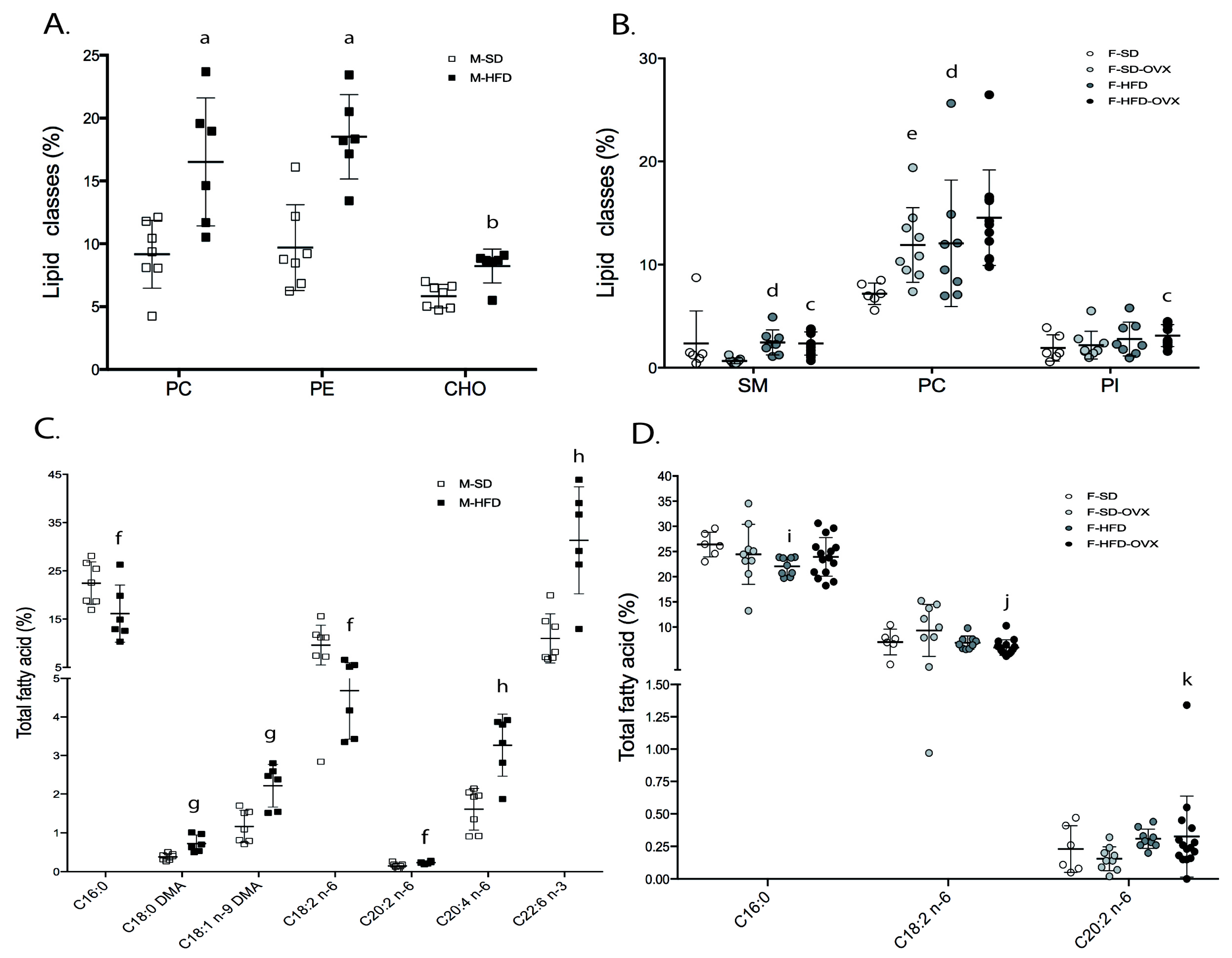
2.2.3. Fatty Acids Profile in Individual Lipid Classes
- Male animals: in phosphatidylcholine (PC), obese mice had lower 16:1 n-7, 18:1 n-7, 18:2 n-6, 20:1 n-9, 20:3 n-6, and 22:5 n-6, and higher 22:6 n-3 than lean animals (Figure 4A-left, Table S4). Phosphatidylethanolamine (PE) of obese mice contained lower 18:1 n-7, 18:2 n-6, 20:1 n-9, 20:3 n-6, and 22:5 n-6, and higher 22:6 n-3 (Figure 4B-left,Table S5). In phosphatidylinositol (PI), 20:3 n-6 was lower and 20:2 n-6 higher in obese animals (Figure 4C-left, Table S6). In triglycerides (TAG), except 18:1 n-9, which was higher, major monounsaturated fatty acids (MUFAs) and saturated fatty acids (SFAs) were lower in obese animals compared with leans (Figure 4D-left, Table S7).
- Female animals: in PC, animals on HFD had lower 16:1 n-7, 18:1 n-7, and 20:3 n-6, and higher 22:5 n-3 than those on SD, irrespective of ovariectomy. Obese not ovariectomized had lower 18:2 n-6 and higher levels of 22:6 n-3 than animals on SD. Finally, obese ovariectomized mice had a higher 18:0 than obese not-castrated or castrated animals on SD (Figure 4A-right, Table S4). In PE, obese animals had lower 20:3 n-6 and 22:4 n-6 and higher levels of 18:0 DMA than those on SD, irrespective of ovariectomy. Obese mice not ovariectomized contained lower 20:0, 20:1 n-9, and 22:5 n-6, and higher 20:5 n-3 and 22:6 n-3 compared to animals on SD (Figure 4B-right, Table S5). In PI, HFD females showed lower 20:3 n-6 than SD irrespective of ovariectomy (Figure 4C-right, Table S6). TAG from obese mice showed lower 16:0, 16:1 n-7, and higher 17:0, 18:1 n-9, and 20:2 n-6 than lean animals, irrespective of ovariectomy. Obese castrated animals had high 18:0 and 20:1 n-9 compared with the other groups but lower 15:0 than mice ovariectomized on SD. Obese not ovariectomized showed higher 18:3 n-3 compared with animals obese, castrated, or on SD. Finally, the SD group had a higher 14:0 than ovariectomized animals on the same diet (Figure 4D-right, Table S7).
2.3. Lipidomic Analysis in Urine
2.3.1. Total Lipid Content and Lipid Classes Profile
2.3.2. Fatty Acids Profile
3. Discussion
4. Materials and Methods
4.1. The Animal Model
4.2. Diets
4.3. Inflammatory Markers in Renal Tissue
4.4. Lipidomic Analysis in Renal Tissues and Urine
4.5. Total Lipid Extraction
4.6. Lipid Classes Analysis
4.7. Fatty Acid Methyl Esters from Total Lipids
4.8. FAMEs from Phospholipids and Triglycerides
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 16 October 2022).
- Bikbov, B.; Purcell, C.A.; Levey, A.S.; Smith, M.; Abdoli, A.; Abebe, M.; Adebayo, O.M.; Afarideh, M.; Agarwal, S.K.; Agudelo-Botero, M.; et al. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Kurella, M.; Lo, J.C.; Chertow, G.M. Metabolic Syndrome and the Risk for Chronic Kidney Disease among Nondiabetic Adults. J. Am. Soc. Nephrol. 2005, 16, 2134–2140. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Tanaka, M.; Okada, H.; Senmaru, T.; Hamaguchi, M.; Asano, M.; Yamazaki, M.; Oda, Y.; Hasegawa, G.; Toda, H.; et al. Metabolically Healthy Obesity and Risk of Incident CKD. Clin. J. Am. Soc. Nephrol. 2015, 10, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, V.T.; Schei, J.; Solbu, M.D.; Jenssen, T.G.; Melsom, T.; Eriksen, B.O. Metabolic syndrome but not obesity measures are risk factors for accelerated age-related glomerular filtration rate decline in the general population. Kidney Int. 2018, 93, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ryu, D.-R.; Kim, S.-J.; Choi, K.-B.; Kang, D.-H. Clinical implication of metabolic syndrome on chronic kidney disease depends on gender and menopausal status: Results from the Korean National Health and Nutrition Examination Survey. Nephrol. Dial. Transplant. 2009, 25, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Jafar, T.H.; Schmid, C.H.; Stark, P.C.; Toto, R.; Remuzzi, G.; Ruggenenti, P.; Marcantoni, C.; Becker, G.; Shahinfar, S.; de Jong, P.E.; et al. The rate of progression of renal disease may not be slower in women compared with men: A patient-level meta-analysis. Nephrol. Dial. Transplant. 2003, 18, 2047–2053. [Google Scholar] [CrossRef]
- Levental, K.R.; Malmberg, E.; Symons, J.L.; Fan, Y.-Y.; Chapkin, R.S.; Ernst, R.; Levental, I. Lipidomic and biophysical homeostasis of mammalian membranes counteracts dietary lipid perturbations to maintain cellular fitness. Nat. Commun. 2020, 11, 1339. [Google Scholar] [CrossRef]
- de Vries, A.P.J.; Ruggenenti, P.; Ruan, X.Z.; Praga, M.; Cruzado, J.M.; Bajema, I.M.; D’Agati, V.D.; Lamb, H.J.; Barlovic, D.P.; Hojs, R.; et al. Fatty kidney: Emerging role of ectopic lipid in obesity-related renal disease. Lancet Diabetes Endocrinol. 2014, 2, 417–426. [Google Scholar] [CrossRef]
- D’Agati, V.D.; Chagnac, A.; de Vries, A.P.; Levi, M.; Porrini, E.; Herman-Edelstein, M.; Praga, M. Obesity-related glomerulopathy: Clinical and pathologic characteristics and pathogenesis. Nat. Rev. Nephrol. 2016, 12, 453–471. [Google Scholar] [CrossRef]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose Tissue in Obesity-Related Inflammation and Insulin Resistance: Cells, Cytokines, and Chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef]
- Cotter, T.G.; Rinella, M. Nonalcoholic Fatty Liver Disease 2020: The State of the Disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Dasilva, G.; Medina, I. Lipidomic methodologies for biomarkers of chronic inflammation in nutritional research: ω-3 and ω-6 lipid mediators. Free Radic. Biol. Med. 2019, 144, 90–109. [Google Scholar] [CrossRef]
- Schebb, N.H.; Kühn, H.; Kahnt, A.S.; Rund, K.M.; O’Donnell, V.B.; Flamand, N.; Peters-Golden, M.; Jakobsson, P.-J.; Weylandt, K.H.; Rohwer, N.; et al. Formation, Signaling and Occurrence of Specialized Pro-Resolving Lipid Mediators—What is the Evidence so far? Front. Pharmacol. 2022, 13, 838782. [Google Scholar] [CrossRef]
- Ma, H.-Y.; Chen, S.; Du, Y. Estrogen and estrogen receptors in kidney diseases. Ren. Fail. 2021, 43, 619–642. [Google Scholar] [CrossRef] [PubMed]
- Al-Safi, Z.A.; Polotsky, A.J. Obesity and Menopause. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Christakis, M.K.; Hasan, H.B.; De Souza, L.R.; Shirreff, L.M. The effect of menopause on metabolic syndrome: Cross-sectional results from the Canadian Longitudinal Study on Aging. Menopause 2020, 27, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, A.E.; Donate-Correa, J.; Luis-Lima, S.; Díaz-Martín, L.L.; Rodríguez-González, C.; Pérez-Pérez, J.A.; Acosta-González, N.G.; Fumero, C.L.; Navarro-Díaz, M.; López-Álvarez, D.; et al. Obesity and metabolic syndrome induce hyperfiltration, glomerulomegaly, and albuminuria in obese ovariectomized female mice and obese male mice. Menopause 2021, 28, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Hewage, S.M.; Prashar, S.; Debnath, S.C.; Karmin, O.; Siow, Y.L. Inhibition of Inflammatory Cytokine Expression Prevents High-Fat Diet-Induced Kidney Injury: Role of Lingonberry Supplementation. Front. Med. 2020, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.; Gai, Z.; Kullak-Ublick, G.A.; Liu, Z. TNF-α Deficiency Prevents Renal Inflammation and Oxidative Stress in Obese Mice. Kidney Blood Press. Res. 2017, 42, 416–427. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, S.; Ma, Y.; Bai, X.-Y.; Chen, X. Role of Toll-like receptors in diabetic renal lesions in a miniature pig model. Sci. Adv. 2015, 1, e1400183. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, Z.; Xiang, Z.; Zeng, C.; Chen, Z.; Ma, X.; Li, L. Obesity-Related Glomerulopathy: Insights from Gene Expression Profiles of the Glomeruli Derived from Renal Biopsy Samples. Endocrinology 2006, 147, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lozada, L.G.; Tapia, E.; Johnson, R.J.; Rodríguez-Iturbe, B.; Herrera-Acosta, J. Glomerular hemodynamic changes associated with arteriolar lesions and tubulointerstitial inflammation. Kidney Int. 2003, 64, S9–S14. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.D.; Ryan, M.J. Immune and Inflammatory Role in Renal Disease. Compr. Physiol. 2013, 3, 957–976. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Chun, S.Y.; Lee, E.; Kim, B.; Yoon, B.; Gil, H.; Han, M.-H.; Ha, Y.-S.; Lee, J.N.; Kwon, T.G.; et al. IL-10 Deficiency Aggravates Renal Inflammation, Fibrosis and Functional Failure in High-Fat Dieted Obese Mice. Tissue Eng. Regen. Med. 2021, 18, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Trauner, M.; Arrese, M.; Wagner, M. Fatty liver and lipotoxicity. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2010, 1801, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Hansell, P.; Welch, W.J.; Blantz, R.C.; Palm, F. Determinants of kidney oxygen consumption and their relationship to tissue oxygen tension in diabetes and hypertension. Clin. Exp. Pharmacol. Physiol. 2012, 40, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Console, L.; Scalise, M.; Giangregorio, N.; Tonazzi, A.; Barile, M.; Indiveri, C. The Link Between the Mitochondrial Fatty Acid Oxidation Derangement and Kidney Injury. Front. Physiol. 2020, 11, 794. [Google Scholar] [CrossRef]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.C. COX-2 and the Kidney. J. Cardiovasc. Pharmacol. 2006, 47 (Suppl. 1), S37–S42. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-H. Renal Effects of Prostaglandins and Cyclooxygenase-2 Inhibitors. Electrolytes Blood Press. 2008, 6, 35–41. [Google Scholar] [CrossRef]
- Li, Y.; Xia, W.; Zhao, F.; Wen, Z.; Zhang, A.; Huang, S.; Jia, Z.; Zhang, Y. Prostaglandins in the pathogenesis of kidney diseases. Oncotarget 2018, 9, 26586–26602. [Google Scholar] [CrossRef]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [PubMed]
- Alfaidi, M.A.; Chamberlain, J.; Rothman, A.; Crossman, D.; Villa-Uriol, M.; Hadoke, P.; Wu, J.; Schenkel, T.; Evans, P.C.; Francis, S.E. Dietary Docosahexaenoic Acid Reduces Oscillatory Wall Shear Stress, Atherosclerosis, and Hypertension, Most Likely Mediated via an IL-1–Mediated Mechanism. J. Am. Heart Assoc. 2018, 7, e008757. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, R.; Picon-Ruiz, M.; Aurrekoetxea-Rodriguez, I.; de Paiva, V.N.; D’amico, M.; Yoon, H.; Radhakrishnan, R.; Morata-Tarifa, C.; Ince, T.; Lippman, M.E.; et al. The Major Pre- and Postmenopausal Estrogens Play Opposing Roles in Obesity-Driven Mammary Inflammation and Breast Cancer Development. Cell Metab. 2020, 31, 1154–1172.e9. [Google Scholar] [CrossRef] [PubMed]
- Lizcano, F.; Guzmán, G. Estrogen Deficiency and the Origin of Obesity during Menopause. BioMed Res. Int. 2014, 2014, 757461. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.B. Menopause and Chronic Kidney Disease. Semin. Nephrol. 2017, 37, 404–411. [Google Scholar] [CrossRef]
- Riant, E.; Waget, A.; Cogo, H.; Arnal, J.-F.; Burcelin, R.; Gourdy, P. Estrogens protect against high-fat diet-induced insulin resistance and glucose intolerance in mice. Endocrinology 2009, 150, 2109–2117. [Google Scholar] [CrossRef]
- Wu, C.-C.; Chang, C.-Y.; Chang, S.-T.; Chen, S.-H. 17β-Estradiol Accelerated Renal Tubule Regeneration in Male Rats After Ischemia/Reperfusion-Induced Acute Kidney Injury. Shock 2016, 46, 158–163. [Google Scholar] [CrossRef]
- Inada, A.; Inada, O.; Fujii, N.L.; Nagafuchi, S.; Katsuta, H.; Yasunami, Y.; Matsubara, T.; Arai, H.; Fukatsu, A.; Nabeshima, Y.-I. Adjusting the 17β–Estradiol-to-Androgen Ratio Ameliorates Diabetic Nephropathy. J. Am. Soc. Nephrol. 2016, 27, 3035–3050. [Google Scholar] [CrossRef]
- Bonanome, A.; Bennett, M.; Grundy, S.M. Metabolic effects of dietary stearic acid in mice: Changes in the fatty acid composition of triglycerides and phospholipids in various tissues. Atherosclerosis 1992, 94, 119–127. [Google Scholar] [CrossRef]
- Rodríguez, R.R.; González-Bulnes, A.; Garcia-Contreras, C.; Rodriguez-Rodriguez, A.E.; Astiz, S.; Vazquez-Gomez, M.; Pesantez, J.L.; Isabel, B.; Salido-Ruiz, E.; González, J.; et al. The Iberian pig fed with high-fat diet: A model of renal disease in obesity and metabolic syndrome. Int. J. Obes. 2019, 44, 457–465. [Google Scholar] [CrossRef] [PubMed]
- The Jackson Laboratory. Phenotype Information for Diet-Induced Obese C57BL/6J (380050). Available online: https://www.jax.org/jax-mice-and-services/strain-data-sheet-pages/phenotype-information-380050 (accessed on 12 November 2022).
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.E.; Henderson, R.J. The rapid analysis of neutral and polar marine lipids using double- development HPTLC and scanning densitometry. J. Exp. Mar. Biol. Ecol. 1989, 129, 189–197. [Google Scholar] [CrossRef]
- Christie, W.W. Lipid Analysis, 3rd ed.; The Oily Press: Bridgewater, UK, 2003; pp. 205–224. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afonso-Alí, A.; Porrini, E.; Teixido-Trujillo, S.; Pérez-Pérez, J.A.; Luis-Lima, S.; Acosta-González, N.G.; Sosa-Paz, I.; Díaz-Martín, L.; Rodríguez-González, C.; Rodríguez-Rodríguez, A.E. The Role of Gender Differences and Menopause in Obesity-Related Renal Disease, Renal Inflammation and Lipotoxicity. Int. J. Mol. Sci. 2023, 24, 12984. https://doi.org/10.3390/ijms241612984
Afonso-Alí A, Porrini E, Teixido-Trujillo S, Pérez-Pérez JA, Luis-Lima S, Acosta-González NG, Sosa-Paz I, Díaz-Martín L, Rodríguez-González C, Rodríguez-Rodríguez AE. The Role of Gender Differences and Menopause in Obesity-Related Renal Disease, Renal Inflammation and Lipotoxicity. International Journal of Molecular Sciences. 2023; 24(16):12984. https://doi.org/10.3390/ijms241612984
Chicago/Turabian StyleAfonso-Alí, Aaron, Esteban Porrini, Silvia Teixido-Trujillo, José Antonio Pérez-Pérez, Sergio Luis-Lima, Nieves Guadalupe Acosta-González, Irene Sosa-Paz, Laura Díaz-Martín, Covadonga Rodríguez-González, and Ana Elena Rodríguez-Rodríguez. 2023. "The Role of Gender Differences and Menopause in Obesity-Related Renal Disease, Renal Inflammation and Lipotoxicity" International Journal of Molecular Sciences 24, no. 16: 12984. https://doi.org/10.3390/ijms241612984
APA StyleAfonso-Alí, A., Porrini, E., Teixido-Trujillo, S., Pérez-Pérez, J. A., Luis-Lima, S., Acosta-González, N. G., Sosa-Paz, I., Díaz-Martín, L., Rodríguez-González, C., & Rodríguez-Rodríguez, A. E. (2023). The Role of Gender Differences and Menopause in Obesity-Related Renal Disease, Renal Inflammation and Lipotoxicity. International Journal of Molecular Sciences, 24(16), 12984. https://doi.org/10.3390/ijms241612984