
Deregulated miRNA Expression in Triple-Negative Breast Cancer of Ancestral Genomic-Characterized Latina Patients
 ,
,
Abstract
:1. Introduction
2. Results
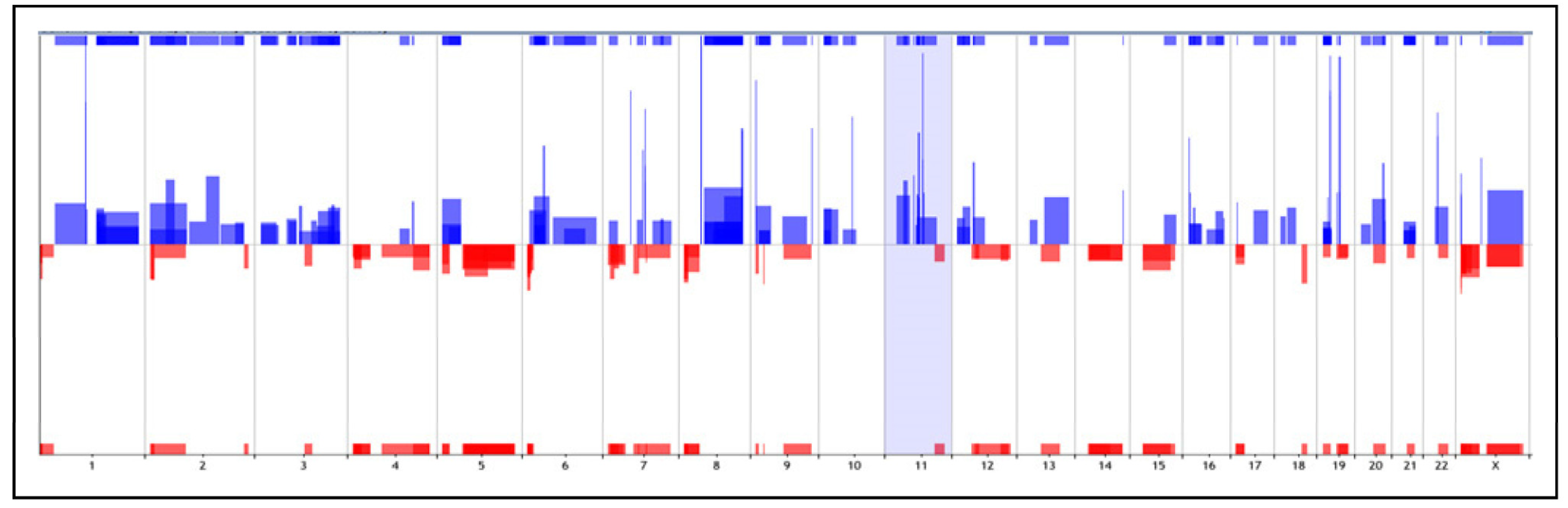
2.1. Copy Number Alterations (CNAs) Analysis
2.2. Global miRNA Expression profiling
2.3. Integration of miRNA Expression and Copy Number Alterations (CNAs) Analysis
2.4. The Cancer Genomic Atlas (TCGA) miRNA Analysis
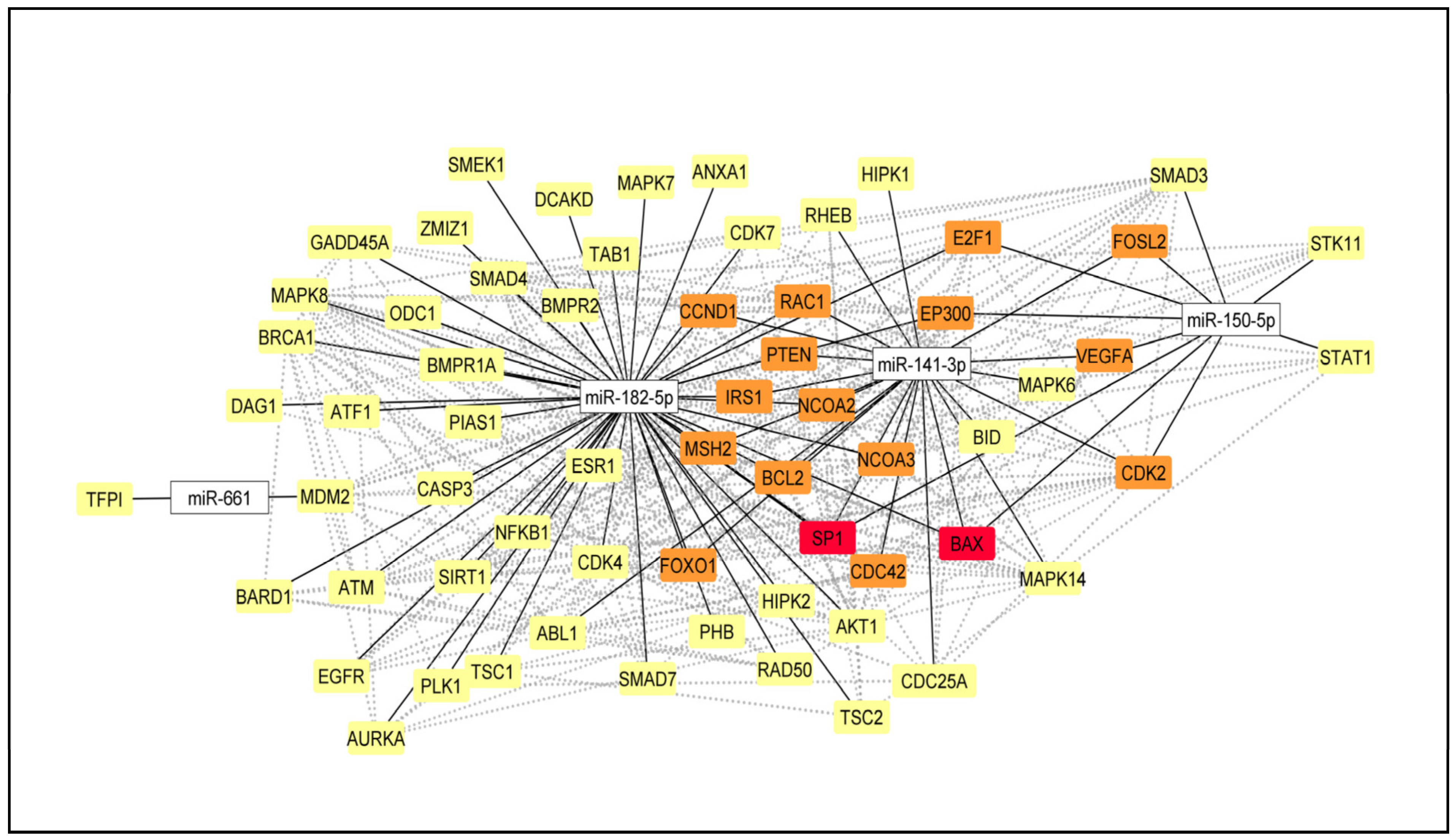
2.5. Selection of the DE miRNA and miRNA-mRNA Network
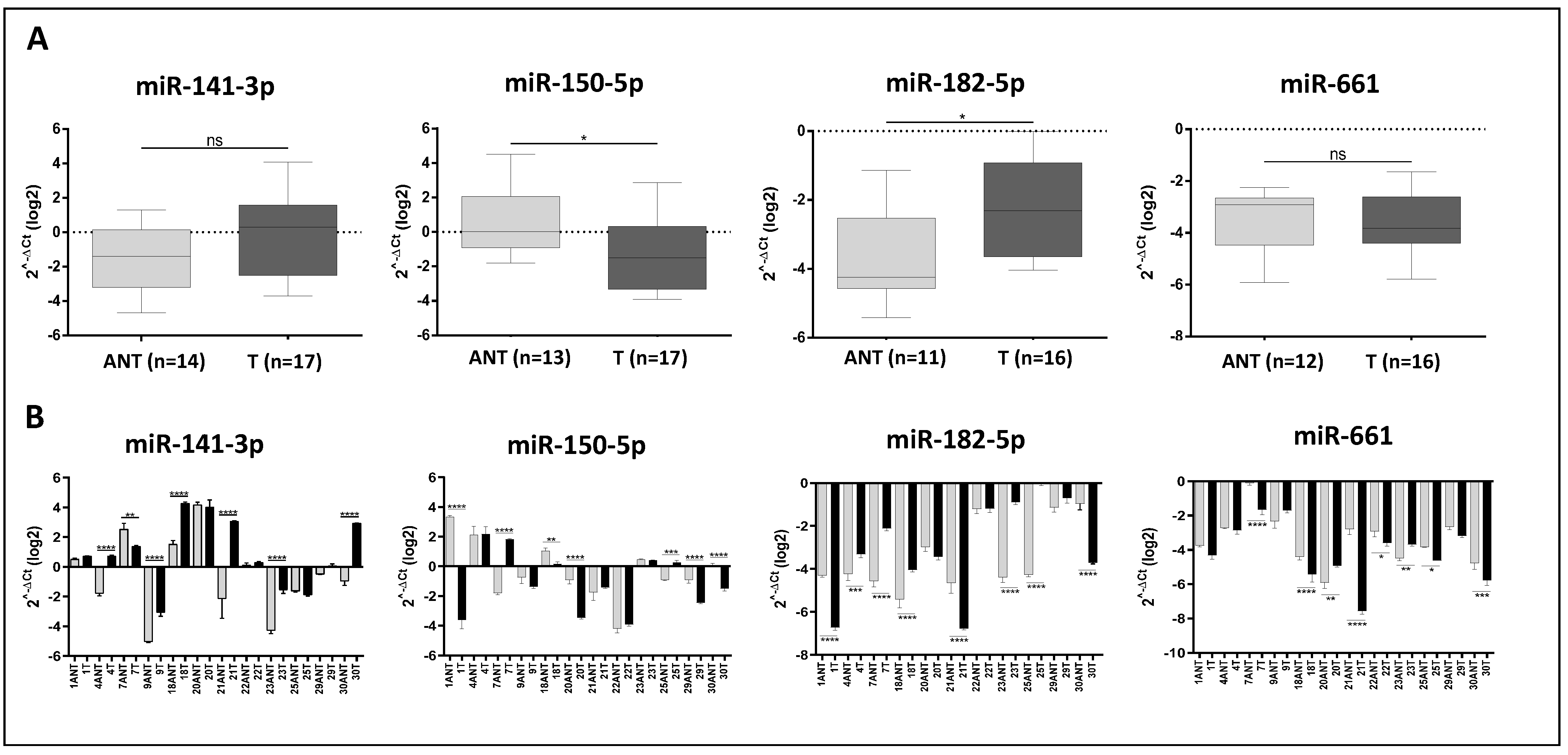
2.6. Validation of the Selected DE miRNA
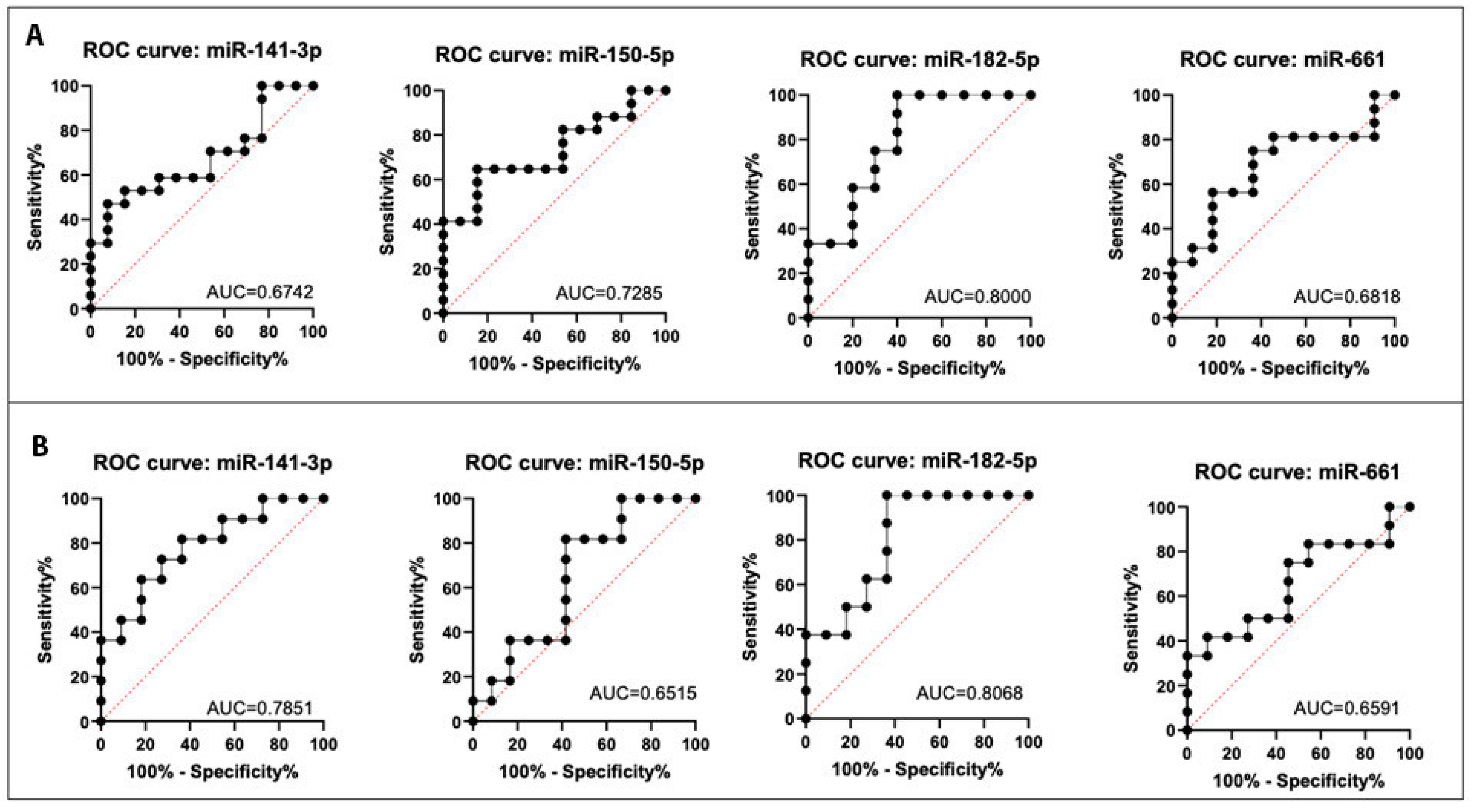
2.7. Discriminatory Power of the Selected DE miRNA
2.8. Association of miR-141-5p, miR-150-5p, miR-182-3p, and miR-661 Expression with the Clinical Parameters of the TNBC Latina Patients

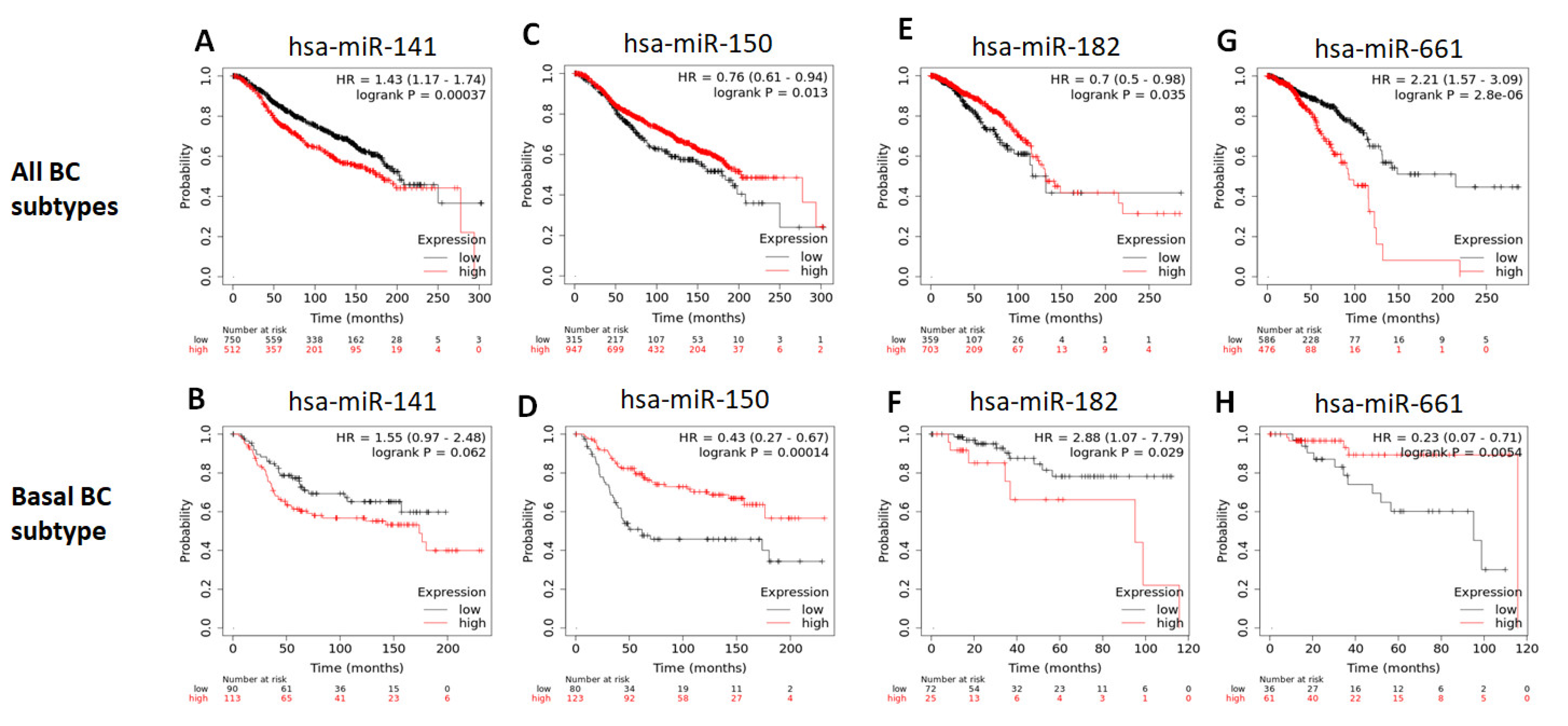
2.9. Survival Analysis
3. Discussion
4. Materials and methods
4.1. Patients’ Accrual and Samples Collection
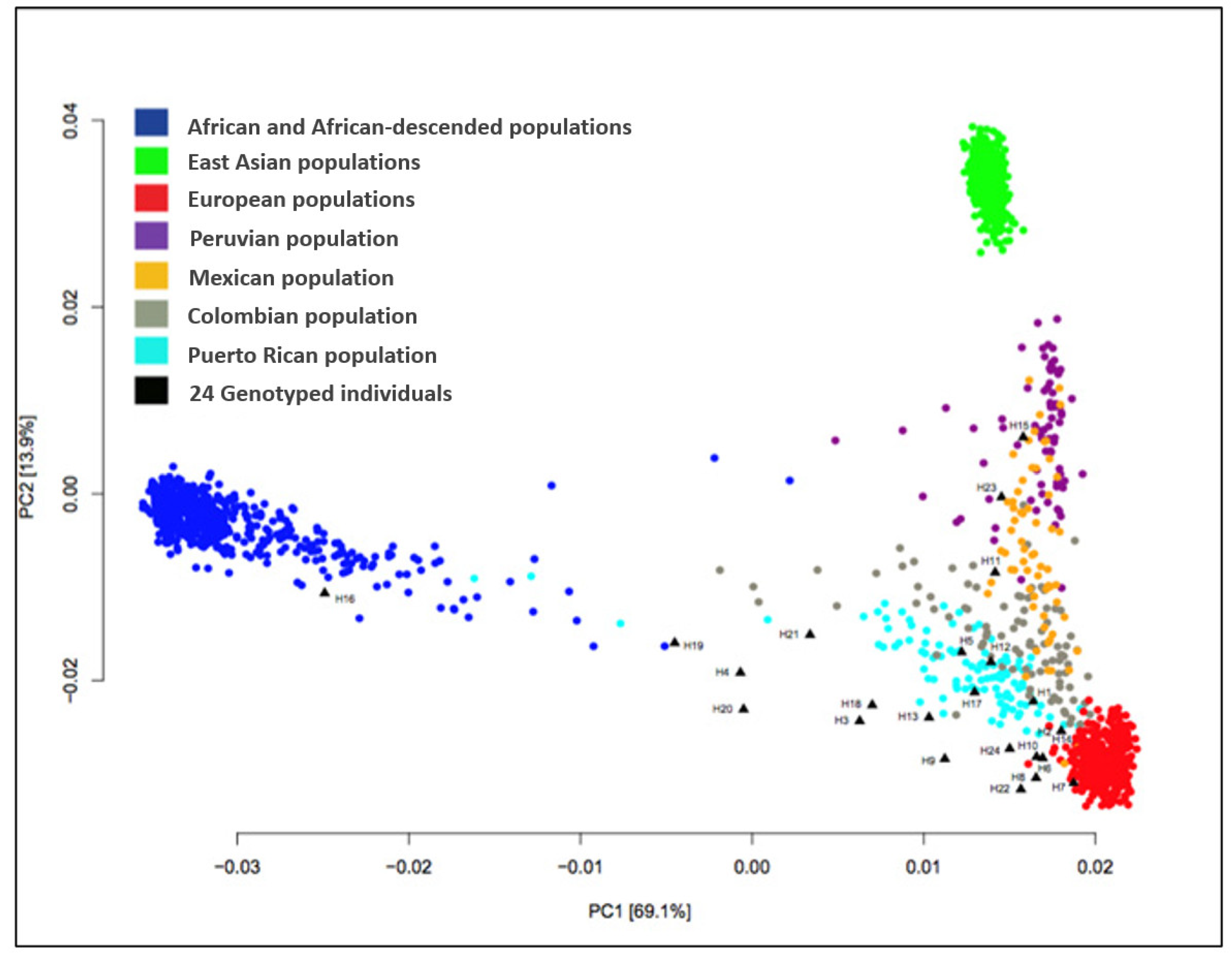
4.2. Ancestral Markers Analysis
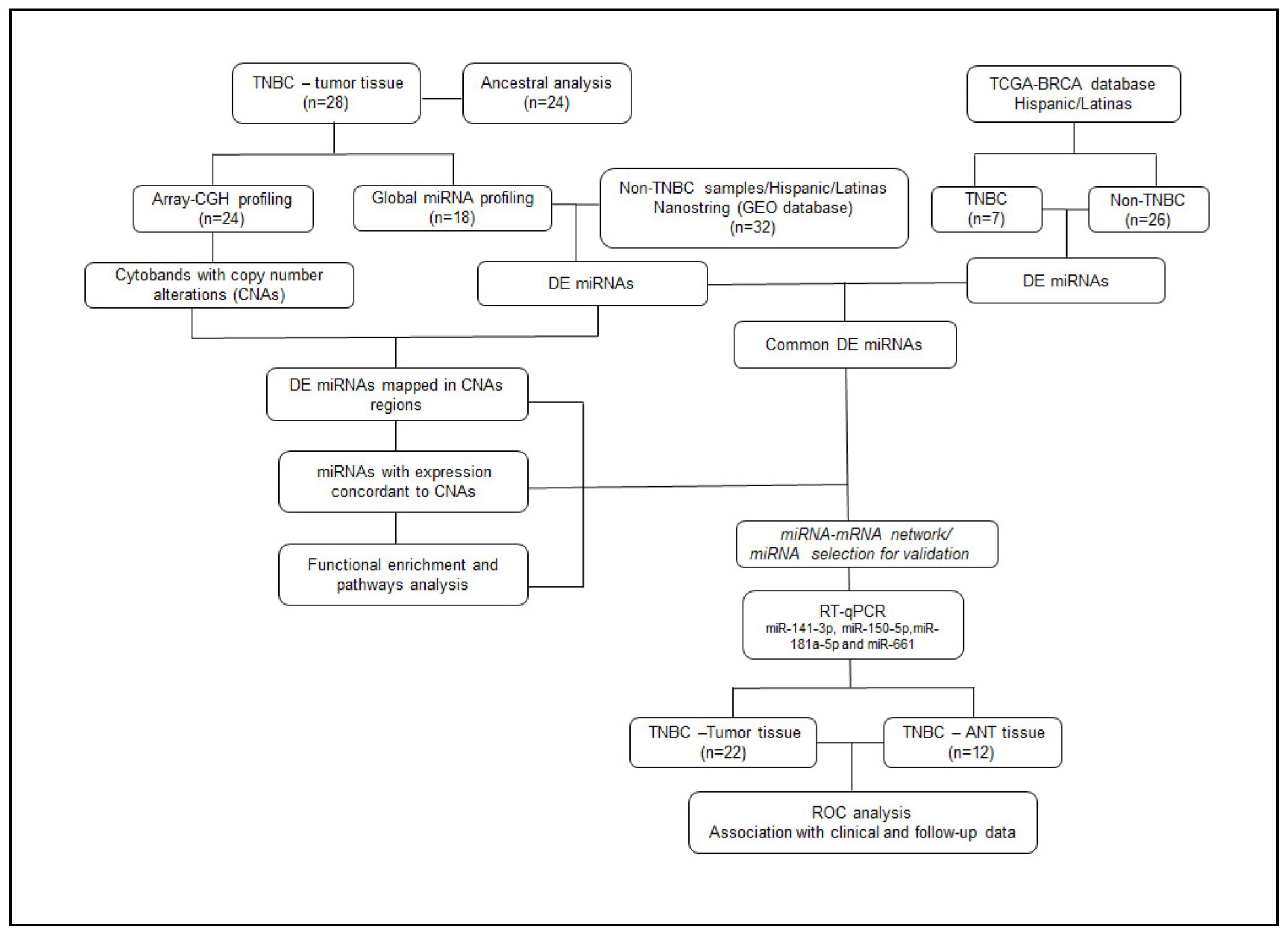
4.3. General Study Design
4.4. Tissue Microdissection and DNA and RNA Isolation
4.5. Array Comparative Genomic Hybridization (Array-CGH) and Analysis
4.6. Global miRNA Expression Analysis and Statistical Analyses
4.7. Integrated Analysis of Array-CGH and miRNA Data
4.8. Biological Function and Pathway Analysis
4.9. The Cancer Genome Atlas (TCGA) Data Processing and Analysis
4.10. Selection of miRNA for RT-qPCR Expression Analysis and miRNA–mRNA Network Construction
4.11. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR) Analysis
4.12. Receiver Operating Characteristic (ROC) Curve Analysis
4.13. Association of the RT-qPCR Results with the Patients’ Clinical–Pathological Data
4.14. Survival Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carey, L.A.; Perou, C.M.; Livasy, C.A.; Dressler, L.G.; Cowan, D.; Conway, K.; Karaca, G.; Troester, M.A.; Tse, C.K.; Edmiston, S.; et al. Race, breast cancer subtypes, and survival in the Carolina Breast Cancer Study. JAMA 2006, 295, 2492–2502. [Google Scholar] [CrossRef]
- Lara-Medina, F.; Pérez-Sánchez, V.; Saavedra-Pérez, D.; Blake-Cerda, M.; Arce, C.; Motola-Kuba, D.; Villarreal-Garza, C.; González-Angulo, A.M.; Bargalló, E.; Aguilar, J.L.; et al. Triple-negative breast cancer in Hispanic patients: High prevalence, poor prognosis, and association with menopausal status, body mass index, and parity. Cancer 2011, 117, 3658–3669. [Google Scholar] [CrossRef] [PubMed]
- Troester, M.A.; Sun, X.; Allott, E.H.; Geradts, J.; Cohen, S.M.; Tse, C.K.; Kirk, E.L.; Thorne, L.B.; Mathews, M.; Li, Y.; et al. Racial Differences in PAM50 Subtypes in the Carolina Breast Cancer Study. J. Natl. Cancer Inst. 2018, 110, 176–182. [Google Scholar] [CrossRef]
- Zhao, F.; Copley, B.; Niu, Q.; Liu, F.; Johnson, J.A.; Sutton, T.; Khramtsova, G.; Sveen, E.; Yoshimatsu, T.F.; Zheng, Y.; et al. Racial disparities in survival outcomes among breast cancer patients by molecular subtypes. Breast Cancer Res. Treat. 2021, 185, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, H., Jr.; Guerra, M.R.; Duarte Cintra, J.R.; Fayer, V.A.; Brum, I.V.; Bustamante Teixeira, M.T. Survival Study of Triple-Negative and Non-Triple-Negative Breast Cancer in a Brazilian Cohort. Clin. Med. Insights. Oncol. 2018, 12, 1179554918790563. [Google Scholar] [CrossRef] [PubMed]
- Rey-Vargas, L.; Sanabria-Salas, M.C.; Fejerman, L.; Serrano-Gómez, S.J. Risk Factors for Triple-Negative Breast Cancer among Latina Women. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1771. [Google Scholar] [CrossRef]
- Martínez, M.E.; Gomez, S.L.; Tao, L.; Cress, R.; Rodriguez, D.; Unkart, J.; Schwab, R.; Nodora, J.N.; Cook, L.; Komenaka, I.; et al. Contribution of clinical and socioeconomic factors to differences in breast cancer subtype and mortality between Hispanic and non-Hispanic white women. Breast Cancer Res. Treat. 2017, 166, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Sineshaw, H.M.; Gaudet, M.; Ward, E.M.; Flanders, W.D.; Desantis, C.; Lin, C.C.; Jemal, A. Association of race/ethnicity, socioeconomic status, and breast cancer subtypes in the National Cancer Data Base (2010–2011). Breast Cancer Res. Treat. 2014, 145, 753–763. [Google Scholar] [CrossRef]
- Edmonds, M.C.; Sutton, A.L.; He, J.; Perera, R.A.; Sheppard, V.B. Correlates of Adjuvant Therapy Attitudes in African American Breast Cancer Patients. J. Natl. Med. Assoc. 2020, 112, 167–175. [Google Scholar] [CrossRef]
- Sheppard, V.B.; Cavalli, L.R.; Dash, C.; Kanaan, Y.M.; Dilawari, A.A.; Horton, S.; Makambi, K.H. Correlates of Triple Negative Breast Cancer and Chemotherapy Patterns in Black and White Women With Breast Cancer. Clin. Breast Cancer 2017, 17, 232–238. [Google Scholar] [CrossRef]
- Burstein, M.D.; Tsimelzon, A.; Poage, G.M.; Covington, K.R.; Contreras, A.; Fuqua, S.A.; Savage, M.I.; Osborne, C.K.; Hilsenbeck, S.G.; Chang, J.C.; et al. Comprehensive genomic analysis identifies novel subtypes and targets of triple-negative breast cancer. Clin. Cancer Res. 2015, 21, 1688–1698. [Google Scholar] [CrossRef]
- Kudelova, E.; Smolar, M.; Holubekova, V.; Hornakova, A.; Dvorska, D.; Lucansky, V.; Koklesova, L.; Kudela, E.; Kubatka, P. Genetic Heterogeneity, Tumor Microenvironment and Immunotherapy in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2022, 23, 14937. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed]
- Mills, M.N.; Yang, G.Q.; Oliver, D.E.; Liveringhouse, C.L.; Ahmed, K.A.; Orman, A.G.; Laronga, C.; Hoover, S.J.; Khakpour, N.; Costa, R.L.B.; et al. Histologic heterogeneity of triple negative breast cancer: A National Cancer Centre Database analysis. Eur. J. Cancer 2018, 98, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Bareche, Y.; Venet, D.; Ignatiadis, M.; Aftimos, P.; Piccart, M.; Rothe, F.; Sotiriou, C. Unravelling triple-negative breast cancer molecular heterogeneity using an integrative multiomic analysis. Ann. Oncol. 2018, 29, 895–902. [Google Scholar] [CrossRef]
- Isaacs, J.; Anders, C.; McArthur, H.; Force, J. Biomarkers of Immune Checkpoint Blockade Response in Triple-Negative Breast Cancer. Curr. Treat. Options Oncol. 2021, 22, 38. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Colaprico, A.; Silva, T.C.; Chen, J.; An, H.; Ban, Y.; Huang, H.; Wang, L.; James, J.L.; Balko, J.M.; et al. Multi-omics analysis identifies therapeutic vulnerabilities in triple-negative breast cancer subtypes. Nat. Commun. 2021, 12, 6276. [Google Scholar] [CrossRef]
- Wang, D.-Y.; Jiang, Z.; Ben-David, Y.; Woodgett, J.R.; Zacksenhaus, E. Molecular stratification within triple-negative breast cancer subtypes. Sci. Rep. 2019, 9, 19107. [Google Scholar] [CrossRef] [PubMed]
- Philipovskiy, A.; Dwivedi, A.K.; Gamez, R.; McCallum, R.; Mukherjee, D.; Nahleh, Z.; Aguilera, R.J.; Gaur, S. Association between tumor mutation profile and clinical outcomes among Hispanic Latina women with triple-negative breast cancer. PLoS ONE 2020, 15, e0238262. [Google Scholar] [CrossRef]
- Sugita, B.M.; Pereira, S.R.; de Almeida, R.C.; Gill, M.; Mahajan, A.; Duttargi, A.; Kirolikar, S.; Fadda, P.; de Lima, R.S.; Urban, C.A.; et al. Integrated copy number and miRNA expression analysis in triple negative breast cancer of Latin American patients. Oncotarget 2019, 10, 6184–6203. [Google Scholar] [CrossRef] [PubMed]
- Arancibia, T.; Morales-Pison, S.; Maldonado, E.; Jara, L. Association between single-nucleotide polymorphisms in miRNA and breast cancer risk: An updated review. Biol. Res. 2021, 54, 26. [Google Scholar] [CrossRef] [PubMed]
- Machowska, M.; Galka-Marciniak, P.; Kozlowski, P. Consequences of genetic variants in miRNA genes. Comput. Struct. Biotechnol. J. 2022, 20, 6443–6457. [Google Scholar] [CrossRef] [PubMed]
- Rawlings-Goss, R.A.; Campbell, M.C.; Tishkoff, S.A. Global population-specific variation in miRNA associated with cancer risk and clinical biomarkers. BMC Med. Genom. 2014, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Kotsyfakis, M.; Patelarou, E. MicroRNAs as biomarkers of harmful environmental and occupational exposures: A systematic review. Biomarkers 2019, 24, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Mirón, P.; Gallego, A.; Bertranpetit, J.; Laayouni, H.; Espinosa-Parrilla, Y. Signatures of genetic variation in human microRNAs point to processes of positive selection and population-specific disease risks. Hum. Genet. 2022, 141, 1673–1693. [Google Scholar] [CrossRef]
- Gong, Z.; Chen, J.; Wang, J.; Liu, S.; Ambrosone, C.B.; Higgins, M.J. Differential methylation and expression patterns of microRNAs in relation to breast cancer subtypes among American women of African and European ancestry. PLoS ONE 2021, 16, e0249229. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target Ther. 2016, 1, 15004. [Google Scholar] [CrossRef]
- Van Roosbroeck, K.; Calin, G.A. Cancer Hallmarks and MicroRNAs: The Therapeutic Connection. Adv. Cancer Res. 2017, 135, 119–149. [Google Scholar] [CrossRef]
- Blenkiron, C.; Goldstein, L.D.; Thorne, N.P.; Spiteri, I.; Chin, S.F.; Dunning, M.J.; Barbosa-Morais, N.L.; Teschendorff, A.E.; Green, A.R.; Ellis, I.O.; et al. MicroRNA expression profiling of human breast cancer identifies new markers of tumor subtype. Genome Biol. 2007, 8, R214. [Google Scholar] [CrossRef]
- Dvinge, H.; Git, A.; Gräf, S.; Salmon-Divon, M.; Curtis, C.; Sottoriva, A.; Zhao, Y.; Hirst, M.; Armisen, J.; Miska, E.A.; et al. The shaping and functional consequences of the microRNA landscape in breast cancer. Nature 2013, 497, 378–382. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Søkilde, R.; Persson, H.; Ehinger, A.; Pirona, A.C.; Fernö, M.; Hegardt, C.; Larsson, C.; Loman, N.; Malmberg, M.; Rydén, L.; et al. Refinement of breast cancer molecular classification by miRNA expression profiles. BMC Genom. 2019, 20, 503. [Google Scholar] [CrossRef]
- Arun, R.P.; Cahill, H.F.; Marcato, P. Breast Cancer Subtype-Specific miRNAs: Networks, Impacts, and the Potential for Intervention. Biomedicines 2022, 10, 651. [Google Scholar] [CrossRef] [PubMed]
- Avery-Kiejda, K.A.; Mathe, A.; Scott, R.J. Genome-wide miRNA, gene and methylation analysis of triple negative breast cancer to identify changes associated with lymph node metastases. Genom. Data 2017, 14, 1–4. [Google Scholar] [CrossRef]
- Gasparini, P.; Cascione, L.; Fassan, M.; Lovat, F.; Guler, G.; Balci, S.; Irkkan, C.; Morrison, C.; Croce, C.M.; Shapiro, C.L.; et al. microRNA expression profiling identifies a four microRNA signature as a novel diagnostic and prognostic biomarker in triple negative breast cancers. Oncotarget 2014, 5, 1174–1184. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cai, Q.; Bao, P.P.; Su, Y.; Cai, H.; Wu, J.; Ye, F.; Guo, X.; Zheng, W.; Zheng, Y.; et al. Tumor tissue microRNA expression in association with triple-negative breast cancer outcomes. Breast Cancer Res. Treat. 2015, 152, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Lü, L.; Mao, X.; Shi, P.; He, B.; Xu, K.; Zhang, S.; Wang, J. MicroRNAs in the prognosis of triple-negative breast cancer: A systematic review and meta-analysis. Medicine 2017, 96, e7085. [Google Scholar] [CrossRef]
- Piasecka, D.; Braun, M.; Kordek, R.; Sadej, R.; Romanska, H. MicroRNAs in regulation of triple-negative breast cancer progression. J. Cancer Res. Clin. Oncol. 2018, 144, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Turkistani, S.; Sugita, B.M.; Fadda, P.; Marchi, R.; Afsari, A.; Naab, T.; Apprey, V.; Copeland, R.L.; Campbell, M.C.; Cavalli, L.R.; et al. A panel of miRNAs as prognostic markers for African-American patients with triple negative breast cancer. BMC Cancer 2021, 21, 861. [Google Scholar] [CrossRef]
- Gong, Z.; Wang, J.; Wang, D.; Buas, M.F.; Ren, X.; Freudenheim, J.L.; Belinsky, S.A.; Liu, S.; Ambrosone, C.B.; Higgins, M.J. Differences in microRNA expression in breast cancer between women of African and European ancestry. Carcinogenesis 2019, 40, 61–69. [Google Scholar] [CrossRef]
- Gupta, I.; Sareyeldin, R.M.; Al-Hashimi, I.; Al-Thawadi, H.A.; Al Farsi, H.; Vranic, S.; Al Moustafa, A.E. Triple Negative Breast Cancer Profile, from Gene to microRNA, in Relation to Ethnicity. Cancers 2019, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Nassar, F.J.; Talhouk, R.; Zgheib, N.K.; Tfayli, A.; El Sabban, M.; El Saghir, N.S.; Boulos, F.; Jabbour, M.N.; Chalala, C.; Boustany, R.M.; et al. microRNA Expression in Ethnic Specific Early Stage Breast Cancer: An Integration and Comparative Analysis. Sci. Rep. 2017, 7, 16829. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.; Burns, P.A.; Hughes, T.A.; Ho-Yen, C.; Jones, J.L.; Mukherjee, G.; Omoniyi-Esan, G.O.; Titloye, N.A.; Speirs, V.; Shaaban, A.M. Differential Expression of MicroRNAs in Breast Cancers from Four Different Ethnicities. Pathobiology 2018, 85, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Sugita, B.; Gill, M.; Mahajan, A.; Duttargi, A.; Kirolikar, S.; Almeida, R.; Regis, K.; Oluwasanmi, O.L.; Marchi, F.; Marian, C.; et al. Differentially expressed miRNAs in triple negative breast cancer between African-American and non-Hispanic white women. Oncotarget 2016, 7, 79274–79291. [Google Scholar] [CrossRef] [PubMed]
- Angajala, A.; Raymond, H.; Muhammad, A.; Uddin Ahmed, M.S.; Haleema, S.; Haque, M.; Wang, H.; Campbell, M.; Martini, R.; Karanam, B.; et al. MicroRNAs within the Basal-like signature of Quadruple Negative Breast Cancer impact overall survival in African Americans. Sci. Rep. 2022, 12, 22178. [Google Scholar] [CrossRef]
- Jinna, N.; Jovanovic-Talisman, T.; LaBarge, M.; Natarajan, R.; Kittles, R.; Sistrunk, C.; Rida, P.; Seewaldt, V.L. Racial Disparity in Quadruple Negative Breast Cancer: Aggressive Biology and Potential Therapeutic Targeting and Prevention. Cancers 2022, 14, 4484. [Google Scholar] [CrossRef]
- MacCuaig, W.M.; Thomas, A.; Claros-Sorto, J.C.; Gomez-Gutierrez, J.G.; Alexander, A.C.; Wellberg, E.A.; Grizzle, W.E.; McNally, L.R. Differential expression of microRNA between triple negative breast cancer patients of African American and European American descent. Biotech. Histochem. 2022, 97, 1–10. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.C.; Korkut, A.; Kanchi, R.S.; Hegde, A.M.; Lenoir, W.; Liu, W.; Liu, Y.; Fan, H.; Shen, H.; Ravikumar, V.; et al. A Comprehensive Pan-Cancer Molecular Study of Gynecologic and Breast Cancers. Cancer Cell 2018, 33, 690–705.e699. [Google Scholar] [CrossRef] [PubMed]
- Ciriello, G.; Gatza, M.L.; Beck, A.H.; Wilkerson, M.D.; Rhie, S.K.; Pastore, A.; Zhang, H.; McLellan, M.; Yau, C.; Kandoth, C.; et al. Comprehensive Molecular Portraits of Invasive Lobular Breast Cancer. Cell 2015, 163, 506–519. [Google Scholar] [CrossRef]
- Puppe, J.; Seifert, T.; Eichler, C.; Pilch, H.; Mallmann, P.; Malter, W. Genomic Signatures in Luminal Breast Cancer. Breast Care 2020, 15, 355–365. [Google Scholar] [CrossRef]
- Spratt, D.E.; Chan, T.; Waldron, L.; Speers, C.; Feng, F.Y.; Ogunwobi, O.O.; Osborne, J.R. Racial/Ethnic Disparities in Genomic Sequencing. JAMA Oncol. 2016, 2, 1070–1074. [Google Scholar] [CrossRef]
- Yuan, J.; Hu, Z.; Mahal, B.A.; Zhao, S.D.; Kensler, K.H.; Pi, J.; Hu, X.; Zhang, Y.; Wang, Y.; Jiang, J.; et al. Integrated Analysis of Genetic Ancestry and Genomic Alterations across Cancers. Cancer Cell 2018, 34, 549–560.e549. [Google Scholar] [CrossRef] [PubMed]
- Carrot-Zhang, J.; Chambwe, N.; Damrauer, J.S.; Knijnenburg, T.A.; Robertson, A.G.; Yau, C.; Zhou, W.; Berger, A.C.; Huang, K.L.; Newberg, J.Y.; et al. Comprehensive Analysis of Genetic Ancestry and Its Molecular Correlates in Cancer. Cancer Cell 2020, 37, 639–654.e636. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Garza, C.; Mohar, A.; Bargallo-Rocha, J.E.; Lasa-Gonsebatt, F.; Reynoso-Noverón, N.; Matus-Santos, J.; Cabrera, P.; Arce-Salinas, C.; Lara-Medina, F.; Alvarado-Miranda, A.; et al. Molecular Subtypes and Prognosis in Young Mexican Women With Breast Cancer. Clin. Breast Cancer 2017, 17, e95–e102. [Google Scholar] [CrossRef]
- Ding, Y.C.; Steele, L.; Warden, C.; Wilczynski, S.; Mortimer, J.; Yuan, Y.; Neuhausen, S.L. Molecular subtypes of triple-negative breast cancer in women of different race and ethnicity. Oncotarget 2019, 10, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Zevallos, A.; Bravo, L.; Bretel, D.; Paez, K.; Infante, U.; Cárdenas, N.; Alvarado, H.; Posada, A.M.; Pinto, J.A. The hispanic landscape of triple negative breast cancer. Crit. Rev. Oncol. Hematol. 2020, 155, 103094. [Google Scholar] [CrossRef]
- Romero-Cordoba, S.L.; Salido-Guadarrama, I.; Rebollar-Vega, R.; Bautista-Piña, V.; Dominguez-Reyes, C.; Tenorio-Torres, A.; Villegas-Carlos, F.; Fernández-López, J.C.; Uribe-Figueroa, L.; Alfaro-Ruiz, L.; et al. Comprehensive omic characterization of breast cancer in Mexican-Hispanic women. Nat. Commun. 2021, 12, 2245. [Google Scholar] [CrossRef]
- Ortiz Valdez, E.; Rangel-Escareño, C.; Matus Santos, J.A.; Vázquez Romo, R.; Guijosa, A.; Villarreal-Garza, C.; Arrieta, O.; Rodríguez-Bautista, R.; Caro-Sánchez, C.H.; Ortega Gómez, A. Characterization of triple negative breast cancer gene expression profiles in Mexican patients. Mol. Clin. Oncol. 2023, 18, 5. [Google Scholar] [CrossRef]
- Hastings, P.J.; Lupski, J.R.; Rosenberg, S.M.; Ira, G. Mechanisms of change in gene copy number. Nat. Rev. Genet. 2009, 10, 551–564. [Google Scholar] [CrossRef]
- Shlien, A.; Malkin, D. Copy number variations and cancer. Genome Med. 2009, 1, 62. [Google Scholar] [CrossRef]
- Lupicki, K.; Elifio-Esposito, S.; Fonseca, A.S.; Weber, S.H.; Sugita, B.; Langa, B.C.; Pereira, S.R.F.; Govender, D.; Panieri, E.; Hiss, D.C.; et al. Patterns of copy number alterations in primary breast tumors of South African patients and their impact on functional cellular pathways. Int. J. Oncol. 2018, 53, 2745–2757. [Google Scholar] [CrossRef]
- Calin, G.A.; Sevignani, C.; Dumitru, C.D.; Hyslop, T.; Noch, E.; Yendamuri, S.; Shimizu, M.; Rattan, S.; Bullrich, F.; Negrini, M.; et al. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc. Natl. Acad. Sci. USA 2004, 101, 2999–3004. [Google Scholar] [CrossRef]
- Selcuklu, S.D.; Yakicier, M.C.; Erson, A.E. An investigation of microRNAs mapping to breast cancer related genomic gain and loss regions. Cancer Genet. Cytogenet. 2009, 189, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Aure, M.R.; Jernström, S.; Krohn, M.; Vollan, H.K.M.; Due, E.U.; Rødland, E.; Kåresen, R.; Ram, P.; Lu, Y.; Mills, G.B.; et al. Integrated analysis reveals microRNA networks coordinately expressed with key proteins in breast cancer. Genome Med. 2015, 7, 21. [Google Scholar] [CrossRef]
- Ferrari, N.; Mohammed, Z.M.A.; Nixon, C.; Mason, S.M.; Mallon, E.; McMillan, D.C.; Morris, J.S.; Cameron, E.R.; Edwards, J.; Blyth, K. Expression of RUNX1 correlates with poor patient prognosis in triple negative breast cancer. PLoS ONE 2014, 9, e100759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Jia, S.; Wang, Y.; Kang, Y.; Zhang, W. Down-regulation of lncRNA-ATB inhibits epithelial–mesenchymal transition of breast cancer cells by increasing miR-141-3p expression. Biochem. Cell Biol. 2019, 97, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-M.; Kim, H.; Lee, K.-H.; Cho, J.-Y. Analysis of opposing histone modifications H3K4me3 and H3K27me3 reveals candidate diagnostic biomarkers for TNBC and gene set prediction combination. BMB Rep. 2020, 53, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Du, Z.; Chen, Y.; Liu, N.; Zhong, Z.; Shen, Y.; Tang, L. Identification of Metastasis-Associated Genes in Triple-Negative Breast Cancer Using Weighted Gene Co-expression Network Analysis. Evol. Bioinform. 2020, 16, 1176934320954868. [Google Scholar] [CrossRef]
- Chang, C.C.; Chiu, C.C.; Liu, P.F.; Wu, C.H.; Tseng, Y.C.; Lee, C.H.; Shu, C.W. Kinome-Wide siRNA Screening Identifies DYRK1B as a Potential Therapeutic Target for Triple-Negative Breast Cancer Cells. Cancers 2021, 13, 5779. [Google Scholar] [CrossRef]
- Dookeran, K.A.; Zhang, W.; Stayner, L.; Argos, M. Associations of two-pore domain potassium channels and triple negative breast cancer subtype in The Cancer Genome Atlas: Systematic evaluation of gene expression and methylation. BMC Res. Notes 2017, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mu, X.; Huang, O.; Wang, Z.; Chen, J.; Chen, D.; Wang, G. ZNF703 promotes triple-negative breast cancer cells through cell-cycle signaling and associated with poor prognosis. BMC Cancer 2022, 22, 226. [Google Scholar] [CrossRef]
- Võsa, U.; Esko, T.; Kasela, S.; Annilo, T. Altered Gene Expression Associated with microRNA Binding Site Polymorphisms. PLoS ONE 2015, 10, e0141351. [Google Scholar] [CrossRef]
- Sugita, B.M.; Rodriguez, Y.; Fonseca, A.S.; Nunes Souza, E.; Kallakury, B.; Cavalli, I.J.; Ribeiro, E.; Aneja, R.; Cavalli, L.R. MiR-150-5p Overexpression in Triple-Negative Breast Cancer Contributes to the In Vitro Aggressiveness of This Breast Cancer Subtype. Cancers 2022, 14, 2156. [Google Scholar] [CrossRef] [PubMed]
- Mendes, D.C.C.; Filho, C.; Garcia, N.; Ricci, M.D.; Soares, J.M.J.; Carvalho, K.C.; Baracat, E.C. Could be FOXO3a, miR-96-5p and miR-182-5p useful for Brazilian women with luminal A and triple negative breast cancers prognosis and target therapy? Clinics 2023, 78, 100155. [Google Scholar] [CrossRef]
- Darbeheshti, F.; Kadkhoda, S.; Keshavarz-Fathi, M.; Razi, S.; Bahramy, A.; Mansoori, Y.; Rezaei, N. Investigation of BRCAness associated miRNA-gene axes in breast cancer: Cell-free miR-182-5p as a potential expression signature of BRCAness. BMC Cancer 2022, 22, 668. [Google Scholar] [CrossRef]
- Lu, C.; Zhao, Y.; Wang, J.; Shi, W.; Dong, F.; Xin, Y.; Zhao, X.; Liu, C. Breast cancer cell-derived extracellular vesicles transfer miR-182-5p and promote breast carcinogenesis via the CMTM7/EGFR/AKT axis. Mol. Med. 2021, 27, 78. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-T.; Tan, C.-C.; Wu, X.-R.; He, R.; Zhang, X.; Wang, Q.-S.; Li, X.-Q.; Zhang, R.; Feng, Y.-M. FOXF2 deficiency accelerates the visceral metastasis of basal-like breast cancer by unrestrictedly increasing TGF-β and miR-182-5p. Cell Death Differ. 2020, 27, 2973–2987. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Chen, B.; Song, X.; Li, Y.; Liang, Y.; Han, D.; Zhang, N.; Zhang, H.; Liu, Y.; Chen, T.; et al. circRNA_0025202 Regulates Tamoxifen Sensitivity and Tumor Progression via Regulating the miR-182-5p/FOXO3a Axis in Breast Cancer. Mol. Ther. 2019, 27, 1638–1652. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Chen, H.; Wu, M.; Peng, S.; Zhang, L. Downregulation of miR-182-5p inhibits the proliferation and invasion of triple-negative breast cancer cells through regulating TLR4/NF-κB pathway activity by targeting FBXW7. Ann. Transl. Med. 2020, 8, 995. [Google Scholar] [CrossRef]
- Zhao, Y.S.; Yang, W.C.; Xin, H.W.; Han, J.X.; Ma, S.G. MiR-182-5p Knockdown Targeting PTEN Inhibits Cell Proliferation and Invasion of Breast Cancer Cells. Yonsei Med. J. 2019, 60, 148–157. [Google Scholar] [CrossRef]
- Gorski, J.J.; James, C.R.; Quinn, J.E.; Stewart, G.E.; Staunton, K.C.; Buckley, N.E.; McDyer, F.A.; Kennedy, R.D.; Wilson, R.H.; Mullan, P.B.; et al. BRCA1 transcriptionally regulates genes associated with the basal-like phenotype in breast cancer. Breast Cancer Res. Treat. 2010, 122, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Stefansson, O.A.; Jonasson, J.G.; Johannsson, O.T.; Olafsdottir, K.; Steinarsdottir, M.; Valgeirsdottir, S.; Eyfjord, J.E. Genomic profiling of breast tumours in relation to BRCA abnormalities and phenotypes. Breast Cancer Res. 2009, 11, R47. [Google Scholar] [CrossRef] [PubMed]
- Churpek, J.E.; Walsh, T.; Zheng, Y.; Moton, Z.; Thornton, A.M.; Lee, M.K.; Casadei, S.; Watts, A.; Neistadt, B.; Churpek, M.M.; et al. Inherited predisposition to breast cancer among African American women. Breast Cancer Res. Treat. 2015, 149, 31–39. [Google Scholar] [CrossRef]
- Han, G.; Qiu, N.; Luo, K.; Liang, H.; Li, H. Downregulation of miroRNA-141 mediates acquired resistance to trastuzumab and is associated with poor outcome in breast cancer by upregulating the expression of ERBB4. J. Cell Biochem. 2019, 120, 11390–11400. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Jeong, J.Y.; Park, J.Y.; Kim, S.W.; Heo, J.H.; Kang, H.; Kim, G.; An, H.J. miR-150 enhances apoptotic and anti-tumor effects of paclitaxel in paclitaxel-resistant ovarian cancer cells by targeting Notch3. Oncotarget 2017, 8, 72788–72800. [Google Scholar] [CrossRef]
- Singh, B.; Sarli, V.N.; Lucci, A. Inhibition of resistant triple-negative breast cancer cells with low-dose 6-mercaptopurine and 5-azacitidine. Oncotarget 2021, 12, 626–637. [Google Scholar] [CrossRef]
- Song, W.; Wu, S.; Wu, Q.; Zhou, L.; Yu, L.; Zhu, B.; Gong, X. The microRNA-141-3p/CDK8 pathway regulates the chemosensitivity of breast cancer cells to trastuzumab. J. Cell Biochem. 2019, 120, 14095–14106. [Google Scholar] [CrossRef]
- Yao, Y.S.; Qiu, W.S.; Yao, R.Y.; Zhang, Q.; Zhuang, L.K.; Zhou, F.; Sun, L.B.; Yue, L. miR-141 confers docetaxel chemoresistance of breast cancer cells via regulation of EIF4E expression. Oncol. Rep. 2015, 33, 2504–2512. [Google Scholar] [CrossRef]
- Hammond, M.E.; Hayes, D.F.; Dowsett, M.; Allred, D.C.; Hagerty, K.L.; Badve, S.; Fitzgibbons, P.L.; Francis, G.; Goldstein, N.S.; Hayes, M.; et al. American Society of Clinical Oncology/College Of American Pathologists guideline recommendations for immunohistochemical testing of estrogen and progesterone receptors in breast cancer. J. Clin. Oncol. 2010, 28, 2784–2795. [Google Scholar] [CrossRef]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. Arch. Pathol. Lab. Med. 2014, 138, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef]
- Torresan, C.; Oliveira, M.M.; Pereira, S.R.; Ribeiro, E.M.; Marian, C.; Gusev, Y.; Lima, R.S.; Urban, C.A.; Berg, P.E.; Haddad, B.R.; et al. Increased copy number of the DLX4 homeobox gene in breast axillary lymph node metastasis. Cancer Genet. 2014, 207, 177–187. [Google Scholar] [CrossRef]
- Kunc, M.; Popęda, M.; Niemira, M.; Szałkowska, A.; Bieńkowski, M.; Pęksa, R.; Łacko, A.; Radecka, B.S.; Braun, M.; Pikiel, J.; et al. microRNA Expression Profile in Single Hormone Receptor-Positive Breast Cancers is Mainly Dependent on HER2 Status-A Pilot Study. Diagnostics 2020, 10, 617. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.; Sugita, B.M.; Bortoletto, S.M.; Fonseca, A.S.; Cavalli, L.R.; Aneja, R. QNBC Is Associated with High Genomic Instability Characterized by Copy Number Alterations and miRNA Deregulation. Int. J. Mol. Sci. 2021, 22, 11548. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Qu, H.; Wang, S.; Wei, J.; Zhang, L.; Ma, R.; Lu, J.; Zhu, J.; Zhong, W.D.; Jia, Z. GDCRNATools: An R/Bioconductor package for integrative analysis of lncRNA, miRNA and mRNA data in GDC. Bioinformatics 2018, 34, 2515–2517. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Chowbina, S.R.; Wu, X.; Zhang, F.; Li, P.M.; Pandey, R.; Kasamsetty, H.N.; Chen, J.Y. HPD: An online integrated human pathway database enabling systems biology studies. BMC Bioinform. 2009, 10, S5. [Google Scholar] [CrossRef]
- Huang, H.Y.; Lin, Y.C.; Li, J.; Huang, K.Y.; Shrestha, S.; Hong, H.C.; Tang, Y.; Chen, Y.G.; Jin, C.N.; Yu, Y.; et al. miRTarBase 2020: Updates to the experimentally validated microRNA-target interaction database. Nucleic Acids Res. 2020, 48, D148–D154. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Cytoband | Start | Stop | Size (kb) | Cases (%) | Gains/Losses | of Probes |
|---|---|---|---|---|---|---|---|
| chr1 | q21.1–q24.2 | 144,374,546 | 1.68 × 108 | 23,435,182 | 6 (28.5%) | Gain | 533 |
| chr3 | q26.1–q27.2 | 166,346,288 | 1.85 × 108 | 18,991,985 | 5 (23.8%) | Gain | 132 |
| chr4 | p16.3–p15.31 | 1,914,109 | 20,323,997 | 18,409,889 | 4 (19.01%) | Loss | 243 |
| chr5 | q21.1–q35.3 | 99,381,621 | 1.72 × 108 | 72,831,986 | 4 (19.01%) | Loss | 1048 |
| chr6 | p25.3–p24.2 | 248,239 | 10,815,671 | 10,567,433 | 8 (33.33%) | Loss | 16 |
| chr6 | p22.3–p21.32 | 17,745,590 | 32,262,768 | 14,517,179 | 4 (19.01%) | Gain | 321 |
| chr8 | q13–q24.3 | 69,999,338 | 1.46 × 108 | 75,139,299 | 7 (33.33%) | Gain | 1038 |
| chr8 | q24.3 | 141,355,101 | 1.45 × 108 | 3,898,583 | 7 (33.33%) | Gain | 587 |
| chr11 | q13.2–q13.3 | 68,249,411 | 70,012,823 | 1,763,413 | 4 (19.01%) | Gain | 31 |
| chr19 | p13.3–p13.11 | 781,586 | 17,833,369 | 17,051,784 | 7 (33.33%) | Gain | 560 |
| chr21 | q21.3–q22.3 | 28,834,275 | 45,382,723 | 16,549,449 | 4 (19.01%) | Gain | 344 |
| chrX | p22.33 | 1,179,089 | 2,353,577 | 1,174,489 | 7 (33.33%) | Loss | 42 |
| chrX | p22.33 | 218,292 | 2,622,294 | 2,404,303 | 5 (23.8%) | Gain | 75 |
| chrX | p22.33–p22.2 | 2,662,039 | 13,621,701 | 10,959,663 | 10 (47.6%) | Loss | 149 |
| Up-Regulated | Down-Regulated | ||||
|---|---|---|---|---|---|
| miRNA | FC (log2) | p-Value | MiRNA | FC (log2) | p-Value |
| hsa-miR-661 | 4.12434 | 1.75 × 10−13 | hsa-miR-141-3p | −4.11176 | 1.00 × 10−6 |
| hsa-miR-1270 | 4.04835 | <0.0001 | hsa-miR-125a-5p | −5.17837 | 2.40 × 10−8 |
| hsa-miR-548 | 3.92366 | <0.0001 | hsa-miR-222-3p | −5.24417 | 1.10 × 10−8 |
| hsa-miR-548h-3p | 3.92366 | <0.0001 | hsa-let-7b-5p | −5.27658 | 1.00 × 10−7 |
| hsa-miR-517a-3p | 3.72959 | 1.11 × 10−15 | hsa-miR-29b-3p | −5.32329 | 5.97 × 10−7 |
| hsa-miR-548al | 3.66887 | 1.33 × 10−8 | hsa-miR-15a-5p | −5.33016 | 1.86 × 10−7 |
| hsa-miR-765 | 3.65976 | 4.88 × 10−15 | hsa-miR-200c-3p | −5.35346 | 1.52× 10−6 |
| hsa-miR-761 | 3.58448 | 5.15 × 10−11 | hsa-miR-4286 | −5.44997 | 1.26× 10−6 |
| hsa-miR-219b-3p | 3.38808 | 7.44 × 10−9 | hsa-miR-93-5p | −5.46019 | 2.19 × 10−7 |
| hsa-miR-605-5p | 3.14118 | 4.53 × 10−8 | hsa-miR-126-3p | −5.70673 | 3.22 × 10−7 |
| hsa-miR-608 | 3.11135 | 4.31 × 10−10 | hsa-miR-181a-5p | −5.71558 | 4.93 × 10−7 |
| hsa-miR-212-3p | 3.04930 | 1.88 × 10−13 | hsa-miR-21-5p | −5.98850 | 1.52× 10−6 |
| hsa-miR-508-3p | 3.00980 | 7.44 × 10−10 | hsa-miR-191-5p | −6.27011 | 4.31 × 10−8 |
| hsa-miR-219a-5p | 3.00575 | 3.08 × 10−8 | hsa-miR-150-5p | −6.33220 | 7.32 × 10−8 |
| hsa-miR-325 | 2.95349 | 9.22 × 10−10 | hsa-let-7a-5p | −8.81173 | 5.50 × 10−7 |
| miRNA | FC (log2) | p-Value | FDR | Cytoband | CNA |
|---|---|---|---|---|---|
| hsa-miR-661 | 4.12434 | 1.75 × 10−13 | 1.75 × 10−11 | 8q24.3 | gain |
| hsa-miR-765 | 3.65976 | 4.88 × 10−15 | 9.75 × 10−13 | 1q23.1 | gain |
| hsa-miR-3151-5p | 2.63406 | 7.04 × 10−7 | 6.46 × 10−6 | 8q22.3 | gain |
| hsa-miR-2053 | 2.94650 | 1.75 × 10−8 | 3.49 × 10−7 | 8q23.3 | gain |
| hsa-miR-548d-5p | 1.98169 | 6.27 × 10−6 | 3.58 × 10−5 | 8q24.13 | gain |
| hsa-miR-6721-5p | 1.58740 | 4.62 × 10−4 | 0.00129 | 6p21.32 | gain |
| hsa-miR-548d-3p | 1.50149 | 2.90 × 10−5 | 1.32 × 10−4 | 8q24.13 | gain |
| hsa-miR-638 | 1.17150 | 1.68 × 10−4 | 5.82 × 10−4 | 19p13.2 | gain |
| hsa-miR-1224-5p | 0.99921 | 3.44 × 10−4 | 0.00103 | 3q27.1 | gain |
| hsa-miR-3150b-3p | 0.49202 | 2.65 × 10−4 | 8.55 × 10−4 | 8q22.1 | gain |
| hsa-miR-1204 | 0.40531 | 6.10 × 10−4 | 0.00150 | 8q24.21 | gain |
| hsa-miR-4448 | 0.39551 | 7.85 × 10−4 | 0.00179 | 3q27.1 | gain |
| hsa-miR-218-5p | −2.71112 | 5.71 × 10−8 | 9.12 × 10−7 | 5q34 | loss |
| hsa-miR-146a-5p | −2.81117 | 1.85 × 10−6 | 1.35 × 10−5 | 5q33.3 | loss |
| hsa-miR-145-5p | −4.69634 | 8.57 × 10−8 | 1.24 × 10−6 | 5q32 | loss |
| TNBC vs. Non-TNBC (This Study) | TNBC vs. Non-TNBC TCGA | |||||
|---|---|---|---|---|---|---|
| FC (log2) | p-Value | FDR | FC (log2) | p-Value | FDR | |
| hsa-let-7a-5p | −8.81174 | 5.50 × 10−7 | 5.29 × 10−6 | −1.03695 | 0.004 | 0.06594 |
| hsa-let-7b-5p | −5.27658 | 1.00 × 10−7 | 1.40 × 10−6 | −1.15425 | 0.010 | 0.10343 |
| hsa-let-7f-5p | −4.62780 | 2.86 × 10−7 | 3.26 × 10−6 | −1.05391 | 0.006 | 0.07720 |
| hsa-let-7g-5p | −4.45426 | 2.27 × 10−6 | 1.59 × 10−5 | −0.57203 | 0.032 | 0.17183 |
| hsa-miR-10a-5p | −2.32238 | 5.63 × 10−9 | 1.40 × 10−7 | −1.60710 | 0.005 | 0.07131 |
| hsa-miR-10b-5p | −1.86443 | 4.05 × 10−6 | 2.55 × 10−5 | −0.93191 | 0.022 | 0.16072 |
| hsa-miR-146b-3p | 0.40531 | 6.10 × 10−4 | 0.001503 | 1.46182 | 0.002 | 0.03960 |
| hsa-miR-181c-5p | −1.87328 | 8.15 × 10−5 | 3.24 × 10−4 | −0.82176 | 0.045 | 0.21220 |
| hsa-miR-191-5p | −6.27011 | 4.31 × 10−8 | 7.16 × 10−7 | −1.33551 | 0.000 | 0.01904 |
| hsa-miR-195-5p | −2.90058 | 4.43 × 10−7 | 4.53 × 10−6 | −1.10876 | 0.044 | 0.20910 |
| hsa-miR-200a-3p | −2.42572 | 1.90 × 10−4 | 6.41 × 10−4 | −1.31242 | 0.011 | 0.11347 |
| hsa-miR-200b-3p | −5.03990 | 2.95 × 10−6 | 1.93 × 10−5 | −1.07680 | 0.016 | 0.14109 |
| hsa-miR-26a-5p | −4.78689 | 2.76 × 10−7 | 3.24 × 10−6 | −0.70270 | 0.042 | 0.20410 |
| hsa-miR-26b-5p | −4.82779 | 1.57 × 10−7 | 2.03 × 10−6 | −0.95776 | 0.017 | 0.14177 |
| hsa-miR-29b-3p | −5.32329 | 5.97 × 10−7 | 5.68 × 10−6 | −0.95562 | 0.031 | 0.16993 |
| hsa-miR-29c-3p | −4.26978 | 2.44 × 10−8 | 4.64 × 10−7 | −1.67903 | 0.000 | 0.01904 |
| hsa-miR-30a-5p | −3.51938 | 1.90 × 10−5 | 9.15 × 10−5 | −1.90155 | 5.71 × 10−5 | 0.00646 |
| hsa-miR-30b-5p | −4.62253 | 8.25 × 10−8 | 1.24 × 10−6 | −0.89401 | 0.003 | 0.05813 |
| hsa-miR-342-3p | −4.73665 | 8.75 × 10−9 | 2.05 × 10−7 | −1.49716 | 0.005 | 0.06655 |
| hsa-miR-34a-5p | −2.44624 | 3.34 × 10−7 | 3.65 × 10−6 | −0.72461 | 0.025 | 0.16814 |
| hsa-miR-423-5p | −2.72085 | 7.42 × 10−6 | 4.14 × 10−5 | −0.84124 | 0.021 | 0.16072 |
| hsa-miR-664a-3p | −1.71081 | 1.43 × 10−4 | 5.20 × 10−4 | −0.87318 | 0.021 | 0.16072 |
| hsa-miR-766-3p | 2.89894 | 1.71 × 10−9 | 5.93 × 10−8 | 1.20479 | 0.000 | 0.02093 |
| Clinical Variable | miR-141-3p | miR-150-5p | miR-182-5p | miR-661 |
|---|---|---|---|---|
| Age at diagnosis | n = 17, p = 0.753 | n = 17, p = 0.410 | n = 16, p = 0.646 | n = 17, p = 0.111 |
| >55.5, ≤55.5 | ||||
| Tumor size (cm) | n = 17, p = 0.218 | n = 16, p = 0.01 | n = 16, p = 0.537 | n = 17, p = 0.013 |
| >1.7, ≤1.7 | ||||
| Tumor grade | n = 15, p = 0.434 | n = 15, p = 0.803 | n = 15, p = 0.273 | n = 15, p = 0.198 |
| 2, 3 | ||||
| Tumor stage | n = 17, p = 0.968 | n = 17, p = 0.569 | n = 16, p = 0.597 | n = 17, p = 0.704 |
| T1/T2, T3/T4 | ||||
| Ki-67 | n = 16, p = 0.272 | n = 16, p = 0.630 | n = 15, p = 0.236 | n = 16, p = 0.969 |
| >10%, ≤10% | ||||
| p53 | n = 16, p = 0.138 | n = 16, p = 0.001 | n = 15, p = 0.297 | n = 16, p = 0.406 |
| >10%, ≤10% | ||||
| BC recurrence | n = 16, p = 0.007 | n = 16, p = 0.009 | n = 16, p = 0.489 | n = 17, p = 0.510 |
| Yes/No | ||||
| Dist Mets | n = 17, p = 0.437 | n = 16, p = 0.011 | n = 16, p = 0.321 | n = 17, p = 0.889 |
| Yes/No | ||||
| Survival status | n = 17, p = 0.846 | n = 17, p = 0.021 | n = 16, p = 0.459 | n = 17, p = 0.654 |
| Alive/Deceased | ||||
| BMI values | n = 17, p = 0.429 | n = 17, p = 0.132 | n = 16, p = 0.627 | n = 17, p = 0.157 |
| >28.3, ≤28.3 | ||||
| Co-morbidities | n = 17, p = 0.155 | n = 17, p = 0.766 | n = 16, p = 0.982 | n = 17, p = 0.779 |
| Yes/No | ||||
| HTN | n = 17, p = 0.173 | n = 17, p = 0.479 | n = 16, p = 0.482 | n = 17, p = 0.160 |
| Yes/No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almohaywi, M.; Sugita, B.M.; Centa, A.; Fonseca, A.S.; Antunes, V.C.; Fadda, P.; Mannion, C.M.; Abijo, T.; Goldberg, S.L.; Campbell, M.C.; et al. Deregulated miRNA Expression in Triple-Negative Breast Cancer of Ancestral Genomic-Characterized Latina Patients. Int. J. Mol. Sci. 2023, 24, 13046. https://doi.org/10.3390/ijms241713046
Almohaywi M, Sugita BM, Centa A, Fonseca AS, Antunes VC, Fadda P, Mannion CM, Abijo T, Goldberg SL, Campbell MC, et al. Deregulated miRNA Expression in Triple-Negative Breast Cancer of Ancestral Genomic-Characterized Latina Patients. International Journal of Molecular Sciences. 2023; 24(17):13046. https://doi.org/10.3390/ijms241713046
Chicago/Turabian StyleAlmohaywi, Maram, Bruna M. Sugita, Ariana Centa, Aline S. Fonseca, Valquiria C. Antunes, Paolo Fadda, Ciaran M. Mannion, Tomilowo Abijo, Stuart L. Goldberg, Michael C. Campbell, and et al. 2023. "Deregulated miRNA Expression in Triple-Negative Breast Cancer of Ancestral Genomic-Characterized Latina Patients" International Journal of Molecular Sciences 24, no. 17: 13046. https://doi.org/10.3390/ijms241713046
APA StyleAlmohaywi, M., Sugita, B. M., Centa, A., Fonseca, A. S., Antunes, V. C., Fadda, P., Mannion, C. M., Abijo, T., Goldberg, S. L., Campbell, M. C., Copeland, R. L., Kanaan, Y., & Cavalli, L. R. (2023). Deregulated miRNA Expression in Triple-Negative Breast Cancer of Ancestral Genomic-Characterized Latina Patients. International Journal of Molecular Sciences, 24(17), 13046. https://doi.org/10.3390/ijms241713046