
Nano-Enabled Antivirals for Overcoming Antibody Escaped Mutations Based SARS-CoV-2 Waves

 , , and
, , and 
Abstract
:1. Introduction
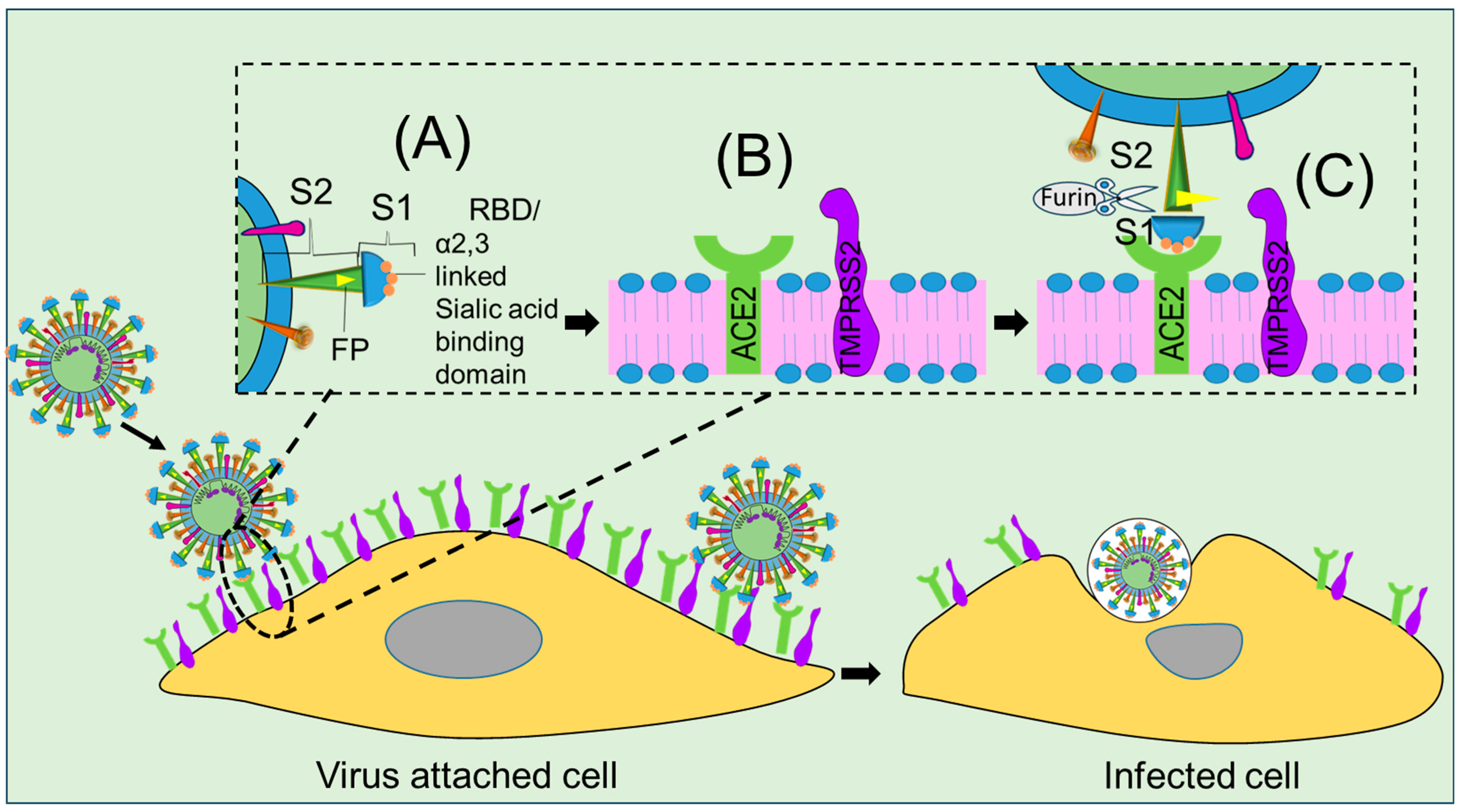
2. Viral Structures Associated with Cell Fusion
3. Escape Mutations and Clinical Waves of COVID-19
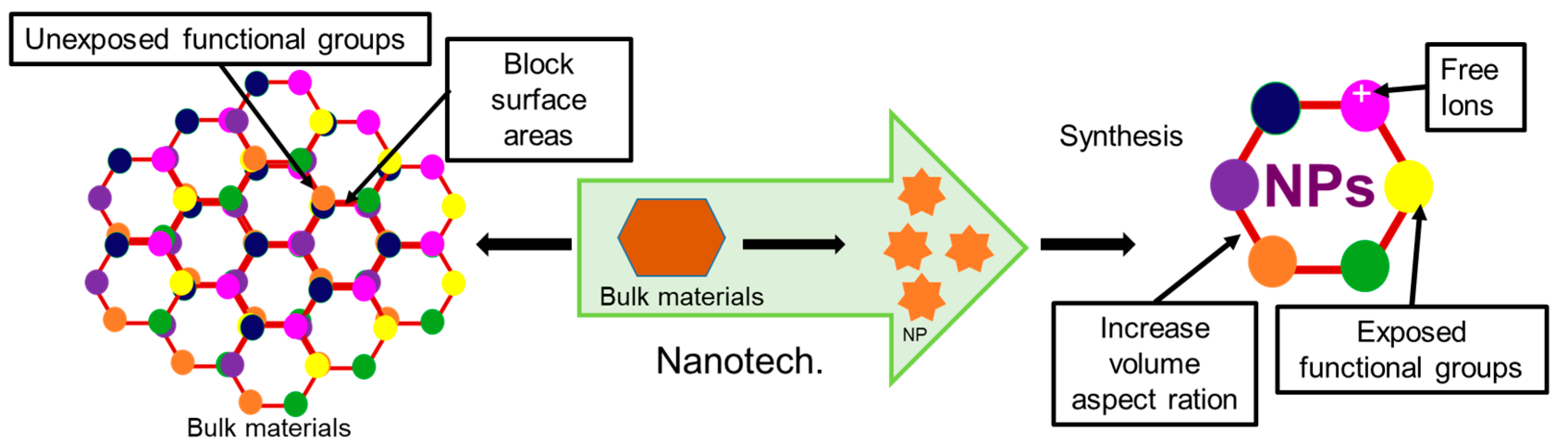
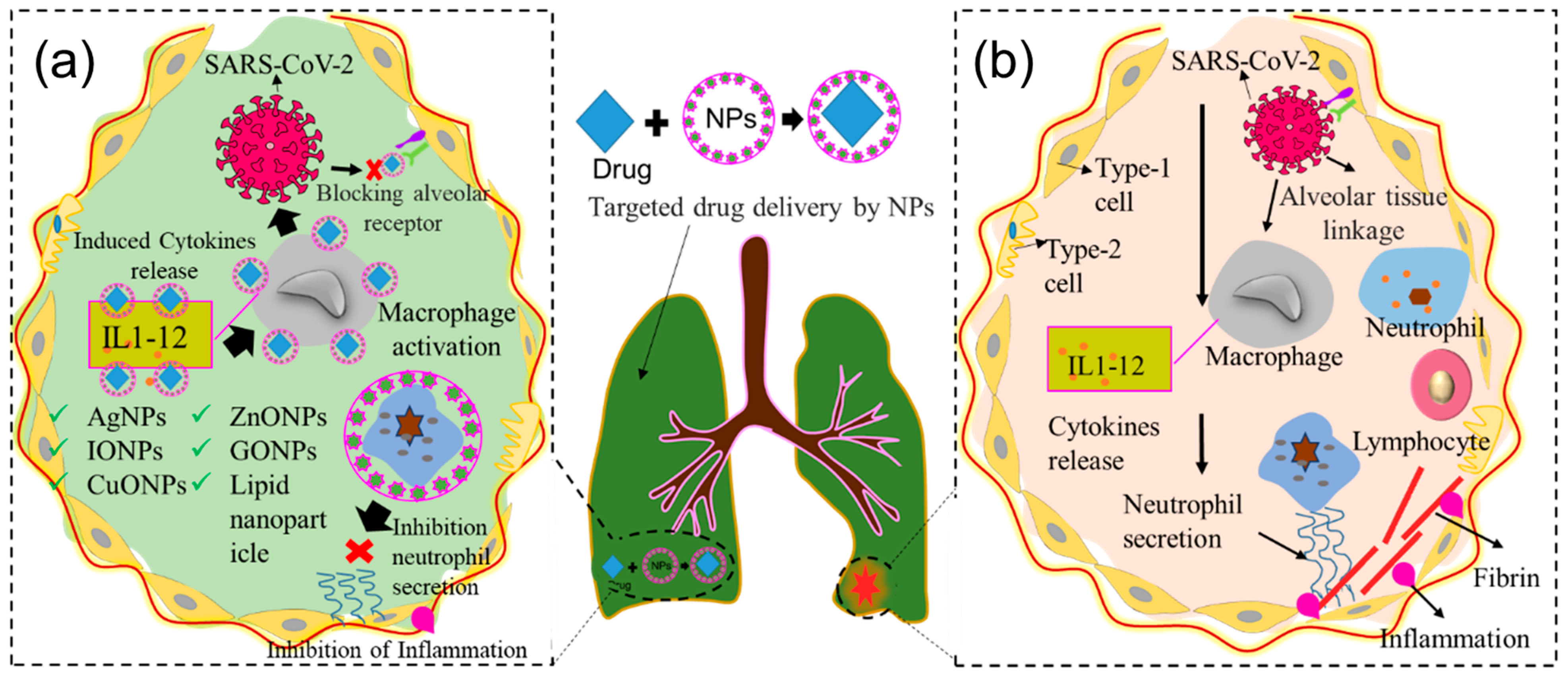
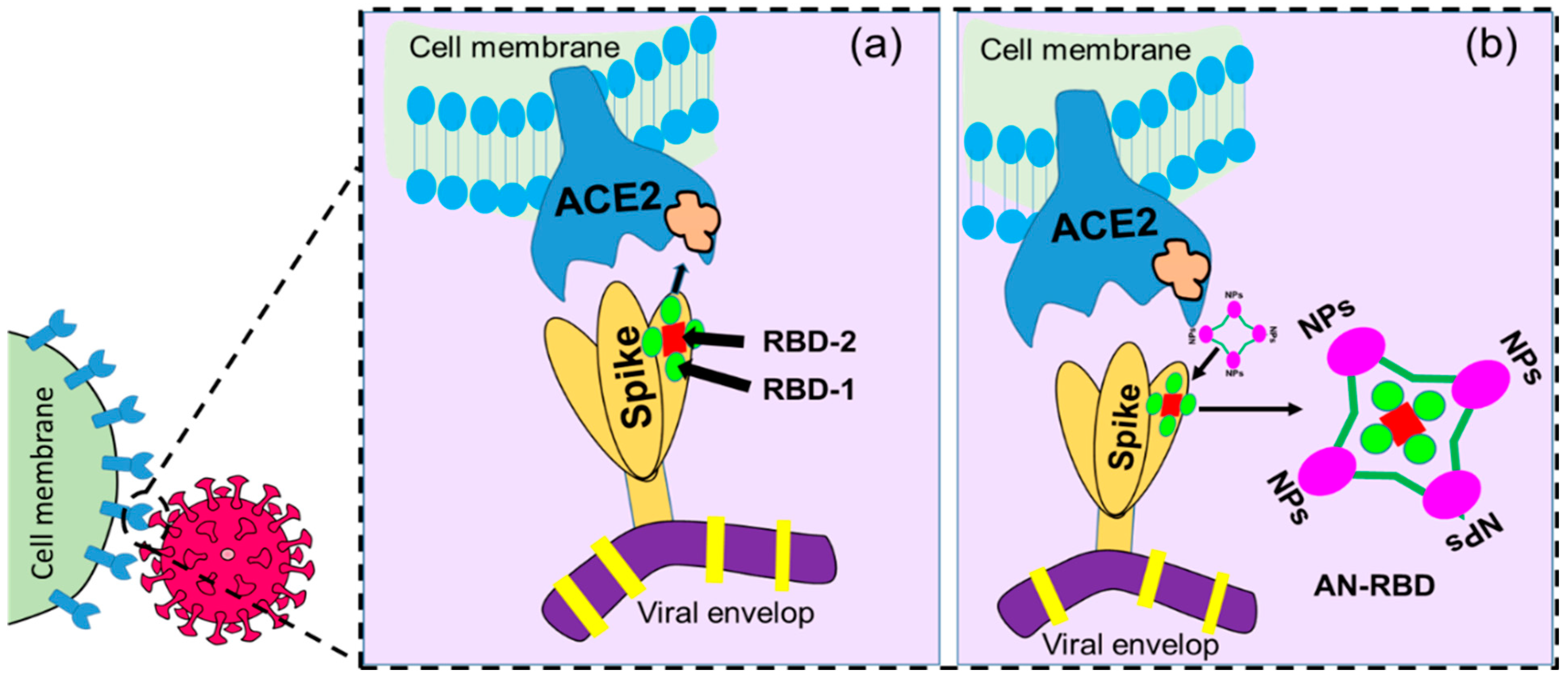
4. Importance of Nanotechnology
5. Antiviral Nanomaterials That Prevent Viral Infections
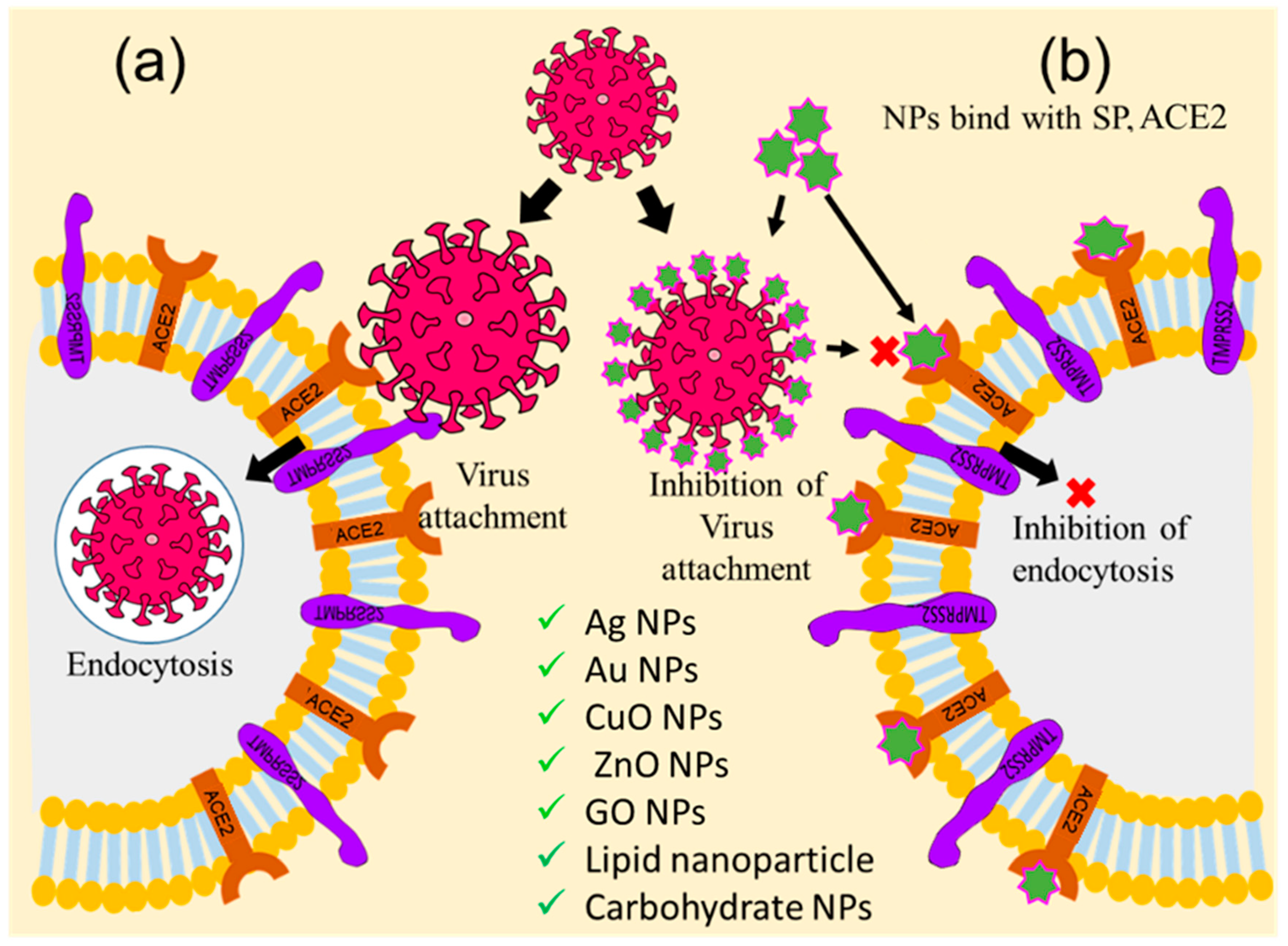
5.1. Inhibition of Receptor-Mediated Host–virus Attachments and Cell Fusion
5.2. Inhibition of Virus Uncoating
5.3. Inhibition of Viral Gene Expression
5.4. Inhibition of Protein Synthesis
5.5. Inhibition of the Viral Particle Assembly
5.6. Inhibition of Virion Release
6. Use of Nanomaterials in Targeted Drug Delivery
7. Use of Nanomaterials in Vaccine Preparations
8. Scope of Nanotechnology in Controlling Clinical Waves of COVID-19
8.1. Development of Nano-Biosensors
8.2. Development of SARS-CoV-2-Neutralizing Nanoparticles
8.3. Development of Nanoscale Antiviral Drugs and Vaccine Carriers
9. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, J.; Pandit, P.; McArthur, A.G.; Banerjee, A.; Mossman, K. Evolutionary trajectory of SARS-CoV-2 and emerging variants. Virol. J. 2021, 18, 166. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. ECDC SARS-CoV-2 Variant Classification Criteria and Recommended EU/EEA Member State Actions—29 June 2023 Recommended EU/EEA Member State Actions; European Centre for Disease Prevention and Control: Stockholm, Sweden, 2023.
- WHO Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 9 August 2023).
- SARS-CoV-2 Variants of Concern as of 27 July 2023. Available online: https://www.ecdc.europa.eu/en/covid-19/variants-concern (accessed on 9 August 2023).
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Makatini, Z.; Moyo, S.; Amoako, D.G.; et al. Emergence of SARS-CoV-2 Omicron lineages BA.4 and BA.5 in South Africa. Nat. Med. 2022, 28, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Providers, H.C.; Variant, O.; Variant, O. SARS-CoV-2 Viral Mutations: Impact on COVID-19 Tests; Food and Drug Administration: Silver Spring, MD, USA, 2019; pp. 1–18.
- Starr, T.N.; Greaney, A.J.; Dingens, A.S.; Bloom, J.D. Complete map of SARS-CoV-2 RBD mutations that escape the monoclonal antibody LY-CoV555 and its cocktail with LY-CoV016. Cell Rep. Med. 2021, 2, 100255. [Google Scholar] [CrossRef] [PubMed]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Focosi, D.; Novazzi, F.; Genoni, A.; Dentali, F.; Gasperina, D.D.; Baj, A.; Maggi, F. Emergence of SARS-CoV-2 Spike Protein Escape Mutation Q493R after Treatment for COVID-19. Emerg. Infect. Dis. 2021, 27, 17–20. [Google Scholar] [CrossRef]
- Knoll, M.D.; Wonodi, C. Oxford-AstraZeneca COVID-19 vaccine efficacy. Lancet 2020, 397, 72–74. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Raman, R.; Patel, K.J.; Ranjan, K. COVID-19: Unmasking emerging SARS-CoV-2 variants, vaccines and therapeutic strategies. Biomolecules 2021, 11, 993. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef]
- Vijayan, V.; Mohapatra, A.; Uthaman, S.; Park, I.K. Recent advances in nanovaccines using biomimetic immunomodulatory materials. Pharmaceutics 2019, 11, 534. [Google Scholar] [CrossRef]
- Tejeda-Mansir, A.; García-Rendón, A.; Guerrero-Germán, P. Plasmid-DNA lipid and polymeric nanovaccines: A new strategic in vaccines development. Biotechnol. Genet. Eng. Rev. 2019, 35, 46–68. [Google Scholar] [CrossRef] [PubMed]
- McGill, J.L.; Kelly, S.M.; Kumar, P.; Speckhart, S.; Haughney, S.L.; Henningson, J.; Narasimhan, B.; Sacco, R.E. Efficacy of mucosal polyanhydride nanovaccine against respiratory syncytial virus infection in the neonatal calf. Sci. Rep. 2018, 8, 3021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Thao, T.T.N.; Hoffmann, D.; Taddeo, A.; Ebert, N.; Labroussaa, F.; Pohlmann, A.; King, J.; Steiner, S.; Kelly, J.N.; et al. SARS-CoV-2 spike D614G change enhances replication and transmission. Nature 2021, 592, 122–127. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Wang, J. Surface charge changes in spike RBD mutations of SARS-CoV-2 and its variant strains alter the virus evasiveness via HSPGs: A review and mechanistic hypothesis. Front. Public Health 2022, 10, 952916. [Google Scholar] [CrossRef] [PubMed]
- Fort, H. A very simple model to account for the rapid rise of the alpha variant of SARS-CoV-2 in several countries and the world. Virus Res. 2021, 304, 198531. [Google Scholar] [CrossRef]
- Khateeb, J.; Li, Y.; Zhang, H. Emerging SARS-CoV-2 variants of concern and potential intervention approaches. Crit. Care 2021, 25, 244. [Google Scholar] [CrossRef] [PubMed]
- Alizon, S.; Haim-Boukobza, S.; Foulongne, V.; Verdurme, L.; Trombert-Paolantoni, S.; Lecorche, E.; Roquebert, B.; Sofonea, M.T. Rapid spread of the SARS-CoV-2 Delta variant in some French regions, June 2021. Euro Surveill. 2021, 26, 2100573. [Google Scholar] [CrossRef]
- Zhand, S.; Jazi, M.S.; Mohammadi, S.; Rasekhi, R.T.; Rostamian, G.; Kalani, M.R.; Rostamian, A.; George, J.; Douglas, M.W. COVID-19: The immune responses and clinical therapy candidates. Int. J. Mol. Sci. 2020, 21, 5559. [Google Scholar] [CrossRef]
- Bilal, M.; Barani, M.; Sabir, F.; Rahdar, A.; Kyzas, G.Z. Nanomaterials for the treatment and diagnosis of Alzheimer’s disease: An overview. NanoImpact 2020, 20, 100251. [Google Scholar] [CrossRef]
- Nowak, M.A.; Lloyd, A.L.; Vasquez, G.M.; Wiltrout, T.A.; Wahl, L.M.; Bischofberger, N.; Williams, J.; Kinter, A.; Fauci, A.S.; Hirsch, V.M.; et al. Viral dynamics of primary viremia and antiretroviral therapy in simian immunodeficiency virus infection. J. Virol. 1997, 71, 7518–7525. [Google Scholar] [CrossRef]
- Hasöksüz, M.; Kiliç, S.; Saraç, F. Coronaviruses and SARS-CoV-2. Turk. J. Med. Sci. 2020, 50, 549–556. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.W.; Cheng, V.C.C.; Cai, J.P.; Chan, K.H.; Chen, L.L.; Wong, L.H.; Choi, C.Y.K.; Fong, C.H.Y.; Ng, A.C.K.; Lu, L.; et al. Seroprevalence of SARS-CoV-2 in Hong Kong and in residents evacuated from Hubei province, China: A multicohort study. Lancet Microbe 2020, 1, e111–e118. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Li, W.; Peng, G.; Li, F. Crystal structure of NL63 respiratory coronavirus receptor-binding domain complexed with its human receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 19970–19974. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, e00127-20. [Google Scholar] [CrossRef]
- Satarker, S.; Nampoothiri, M. Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef]
- Li, F. Evidence for a Common Evolutionary Origin of Coronavirus Spike Protein Receptor-Binding Subunits. J. Virol. 2012, 86, 2856–2858. [Google Scholar] [CrossRef]
- Mittal, A.; Manjunath, K.; Ranjan, R.K.; Kaushik, S.; Kumar, S.; Verma, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- DeDiego, M.L.; Nieto-Torres, J.L.; Jimenez-Guardeño, J.M.; Regla-Nava, J.A.; Castaño-Rodriguez, C.; Fernandez-Delgado, R.; Usera, F.; Enjuanes, L. Coronavirus virulence genes with main focus on SARS-CoV envelope gene. Virus Res. 2014, 194, 124–137. [Google Scholar] [CrossRef]
- Qu, X.-X.; Hao, P.; Song, X.-J.; Jiang, S.-M.; Liu, Y.-X.; Wang, P.-G.; Rao, X.; Song, H.-D.; Wang, S.-Y.; Zuo, Y.; et al. Identification of two critical amino acid residues of the severe acute respiratory syndrome coronavirus spike protein for its variation in zoonotic tropism transition via a double substitution strategy. J. Biol. Chem. 2005, 280, 29588–29595. [Google Scholar] [CrossRef]
- Wu, K.; Chen, L.; Peng, G.; Zhou, W.; Pennell, C.A.; Mansky, L.M.; Geraghty, R.J.; Li, F. A Virus-Binding Hot Spot on Human Angiotensin-Converting Enzyme 2 Is Critical for Binding of Two Different Coronaviruses. J. Virol. 2011, 85, 5331–5337. [Google Scholar] [CrossRef]
- Bakhshandeh, B.; Jahanafrooz, Z.; Abbasi, A.; Goli, M.B.; Sadeghi, M.; Mottaqi, M.S.; Zamani, M. Mutations in SARS-CoV-2; Consequences in structure, function, and pathogenicity of the virus. Microb. Pathog. 2021, 154, 104831. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, Á.; Pybus, O.G.; Abram, M.E.; Kelly, E.J.; Rambaut, A. Pango lineage designation and assignment using SARS-CoV-2 spike gene nucleotide sequences. BMC Genom. 2022, 23, 121. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.K.; Wegner, F.; Gsponer, S.; Helfenstein, F.; Roloff, T.; Tarnutzer, R.; Grosheintz, K.; Back, M.; Schaubhut, C.; Wagner, S.; et al. SARS-CoV-2 Vaccine Alpha and Delta Variant Breakthrough Infections Are Rare and Mild but Can Happen Relatively Early after Vaccination. Microorganisms 2022, 10, 857. [Google Scholar] [CrossRef] [PubMed]
- Carabelli, A.M.; Peacock, T.P.; Thorne, L.G.; Harvey, W.T.; Hughes, J.; de Silva, T.I.; Peacock, S.J.; Barclay, W.S.; de Silva, T.I.; Towers, G.J.; et al. SARS-CoV-2 variant biology: Immune escape, transmission and fitness. Nat. Rev. Microbiol. 2023, 21, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Zhang, X.E.; Sun, X.; Zhang, X.; Yao, Y.; Liu, S.; Chen, Z.; Li, W.; Zhang, Z.; Chen, J.; et al. Intranasal Nanovaccine Confers Homo- and Hetero-Subtypic Influenza Protection. Small 2018, 14, e1703207. [Google Scholar] [CrossRef] [PubMed]
- Vicente, S.; Diaz-Freitas, B.; Peleteiro, M.; Sanchez, A.; Pascual, D.W.; Gonzalez-Fernandez, A.; Alonso, M.J. A Polymer/Oil Based Nanovaccine as a Single-Dose Immunization Approach. PLoS ONE 2013, 8, e62500. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Zhang, L.; Li, Q.; Liang, Z.; Li, T.; Liu, S.; Cui, Q.; Nie, J.; Wu, Q.; Qu, X.; Huang, W.; et al. The significant immune escape of pseudotyped SARS-CoV-2 variant Omicron. Emerg. Microbes Infect. 2022, 11, 1–5. [Google Scholar] [CrossRef]
- Malik, S.; Muhammad, K.; Waheed, Y. Nanotechnology: A Revolution in Modern Industry. Molecules 2023, 28, 661. [Google Scholar] [CrossRef]
- Rahman, A.; Rasid, H.; Ali, I.; Yeachin, N.; Alam, S.; Hossain, K.S.; Kaf, A. Facile Synthesis and Application of Ag-NPs for Controlling Antibiotic-Resistant Pseudomonas spp. and Bacillus spp. in a Poultry Farm Environment. J. Nanotechnol. 2023, 2023, 6260066. [Google Scholar] [CrossRef]
- Khan, I.; Khan, M.; Umar, M.N.; Oh, D.H. Nanobiotechnology and its applications in drug delivery system: A review. IET Nanobiotechnol. 2015, 9, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Kafi, M.A.; Kim, T.H.; Yagati, A.K.; Kim, H.; Choi, J.W. Nanoscale fabrication of a peptide layer in cell chip to detect effects of environmental toxins on HEK293 cells. Biotechnol. Lett. 2010, 32, 1797–1802. [Google Scholar] [CrossRef] [PubMed]
- Kafi, M.A.; Cho, H.Y.; Choi, J.W. Engineered peptide-based nanobiomaterials for electrochemical cell chip. Nano Converg. 2016, 3, 17. [Google Scholar] [CrossRef]
- Kafi, M.A.; Kim, T.H.; Yea, C.H.; Kim, H.; Choi, J.W. Effects of nanopatterned RGD peptide layer on electrochemical detection of neural cell chip. Biosens. Bioelectron. 2010, 26, 1359–1365. [Google Scholar] [CrossRef]
- Kafi, M.A.; Cho, H.Y.; Choi, J.W. Neural cell chip based electrochemical detection of nanotoxicity. Nanomaterials 2015, 5, 1181–1199. [Google Scholar] [CrossRef]
- Rani Sarkar, M.; Rashid, M.H.; Rahman, A.; Kafi, M.A.; Hosen, M.I.; Rahman, M.S.; Khan, M.N. Recent advances in nanomaterials based sustainable agriculture: An overview. Environ. Nanotechnol. Monit. Manag. 2022, 18, 100687. [Google Scholar] [CrossRef]
- Wong, K.V. Nanoscience and Nanotechnology. In Nanotechnology and Energy; Jenny Stanford Publishing: New York, NY, USA, 2018; ISBN 9781259007323. [Google Scholar]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; Rehman, H.U.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef]
- Weiss, C.; Carriere, M.; Fusco, L.; Fusco, L.; Capua, I.; Regla-Nava, J.A.; Pasquali, M.; Pasquali, M.; Pasquali, M.; Scott, J.A.; et al. Toward Nanotechnology-Enabled Approaches against the COVID-19 Pandemic. ACS Nano 2020, 14, 6383–6406. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef]
- Vahedifard, F.; Chakravarthy, K. Nanomedicine for COVID-19: The role of nanotechnology in the treatment and diagnosis of COVID-19. Emergent Mater. 2021, 4, 75–99. [Google Scholar] [CrossRef] [PubMed]
- Caputo, F.; Metcalfe, S.; Tosi, G.; Spring, K.; Åslund, A.K.O.; Pottier, A.; Schi, R.; Ceccaldi, A.; Schmid, R. Since January 2020 Elsevier has created a COVID-19 resource centre with free information in English and Mandarin on the novel coronavirus COVID-19. The COVID-19 resource centre is hosted on Elsevier Connect, the company ’s public news and information. J. Control. Release 2020, 326, 164–171. [Google Scholar]
- Umair, M.; Javed, I.; Rehman, M.; Madni, A.; Javeed, A.; Ghafoor, A.; Ashraf, M. Nanotoxicity of inert materials: The case of gold, silver and iron. J. Pharm. Pharm. Sci. 2016, 19, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Trigilio, J.; Antoine, T.E.; Paulowicz, I.; Mishra, Y.K.; Adelung, R.; Shukla, D. Tin Oxide Nanowires Suppress Herpes Simplex Virus-1 Entry and Cell-to-Cell Membrane Fusion. PLoS ONE 2012, 7, e48147. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, N.; Kaur, G.; Kumar, N.; Tiwari, A. Application of silver nanoparticles in viral inhibition: A new hope for antivirals. Dig. J. Nanomater. Biostruct. 2014, 9, 175–186. [Google Scholar]
- Lin, Z.; Li, Y.; Xu, T.; Guo, M.; Wang, C.; Zhao, M.; Chen, H.; Kuang, J.; Li, W.; Zhang, Y.; et al. Inhibition of Enterovirus 71 by Selenium Nanoparticles Loaded with siRNA through Bax Signaling Pathways. ACS Omega 2020, 5, 12495–12500. [Google Scholar] [CrossRef]
- Mainardes, R.M.; Diedrich, C. The potential role of nanomedicine on COVID-19 therapeutics. Ther. Deliv. 2020, 11, 411–414. [Google Scholar] [CrossRef]
- Tavakoli, A.; Hashemzadeh, M.S. Inhibition of herpes simplex virus type 1 by copper oxide nanoparticles. J. Virol. Methods 2020, 275, 113688. [Google Scholar] [CrossRef]
- Ren, S.; Fraser, K.; Kuo, L.; Chauhan, N.; Adrian, A.T.; Zhang, F.; Linhardt, R.J.; Kwon, P.S.; Wang, X. Designer DNA nanostructures for viral inhibition. Nat. Protoc. 2022, 17, 282–326. [Google Scholar] [CrossRef]
- Wang, P.; Wu, S.; Tian, C.; Yu, G.; Jiang, W.; Wang, G.; Mao, C. Retrosynthetic Analysis-Guided Breaking Tile Symmetry for the Assembly of Complex DNA Nanostructures. J. Am. Chem. Soc. 2016, 138, 13579–13585. [Google Scholar] [CrossRef]
- Mehanna, M.M.; Mohyeldin, S.M.; Elgindy, N.A. Respirable nanocarriers as a promising strategy for antitubercular drug delivery. J. Control. Release 2014, 187, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.; Zheng, Y.; Duan, W.; Li, X.; Yin, J.; Shigdar, S.; O’Connor, M.L.; Marappan, M.; Zhao, X.; Miao, Y.; et al. Inhibition of A/Human/Hubei/3/2005 (H3N2) influenza virus infection by silver nanoparticles in vitro and in vivo. Int. J. Nanomed. 2013, 8, 4103–4114. [Google Scholar] [CrossRef] [PubMed]
- Benarba, B.; Pandiella, A. Medicinal Plants as Sources of Active Molecules Against COVID-19. Front. Pharmacol. 2020, 11, 1189. [Google Scholar] [CrossRef] [PubMed]
- Aderibigbe, B.A. Metal-based nanoparticles for the treatment of infectious diseases. Molecules 2017, 22, 1370. [Google Scholar] [CrossRef] [PubMed]
- Szunerits, S.; Barras, A.; Khanal, M.; Pagneux, Q.; Boukherroub, R. Nanostructures for the inhibition of viral infections. Molecules 2015, 20, 14051–14081. [Google Scholar] [CrossRef]
- Mohammed Fayaz, A.; Ao, Z.; Girilal, M.; Chen, L.; Xiao, X.; Kalaichelvan, P.T.; Yao, X. Inactivation of microbial infectiousness by silver nanoparticles-coated condom: A new approach to inhibit HIV- and HSV-transmitted infection. Int. J. Nanomed. 2012, 7, 5007–5018. [Google Scholar]
- Baram-Pinto, D.; Shukla, S.; Perkas, N.; Gedanken, A.; Sarid, R. Inhibition of herpes simplex virus type 1 infection by silver nanoparticles capped with mercaptoethane sulfonate. Bioconjug. Chem. 2009, 20, 1497–1502. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Ganesan, S. Gold Nanoparticles as an HIV Entry Inhibitor. Curr. HIV Res. 2014, 10, 643–646. [Google Scholar] [CrossRef]
- Kesarkar, R. L-Cysteine Functionalized Gold Nanocargos Potentiates Anti-HIV Activity of Azidothymydine against HIV-1Ba-L Virus. Juniper Online J. Immuno Virol. 2015, 1, 555552. [Google Scholar] [CrossRef]
- Mishra, Y.K.; Adelung, R.; Röhl, C.; Shukla, D.; Spors, F.; Tiwari, V. Virostatic potential of micro-nano filopodia-like ZnO structures against herpes simplex virus-1. Antivir. Res. 2011, 92, 305–312. [Google Scholar] [CrossRef]
- Shoji, M.; Takahashi, E.; Hatakeyama, D.; Iwai, Y.; Morita, Y.; Shirayama, R.; Echigo, N.; Kido, H.; Nakamura, S.; Mashino, T.; et al. Anti-Influenza Activity of C60 Fullerene Derivatives. PLoS ONE 2013, 8, e66337. [Google Scholar] [CrossRef]
- Ziem, B.; Rahn, J.; Donskyi, I.; Silberreis, K.; Cuellar, L.; Dernedde, J.; Keil, G.; Mettenleiter, T.C.; Haag, R. Polyvalent 2D Entry Inhibitors for Pseudorabies and African Swine Fever Virus. Macromol. Biosci. 2017, 17, 1600499. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Liang, J.; Dong, N.; Liu, L.; Fang, L.; Xiao, S.; Han, H. Carbon dots as inhibitors of virus by activation of type I interferon response. Carbon N. Y. 2016, 110, 278–285. [Google Scholar] [CrossRef]
- Łoczechin, A.; Séron, K.; Barras, A.; Giovanelli, E.; Belouzard, S.; Chen, Y.T.; Metzler-Nolte, N.; Boukherroub, R.; Dubuisson, J.; Szunerits, S. Functional Carbon Quantum Dots as Medical Countermeasures to Human Coronavirus. ACS Appl. Mater. Interfaces 2019, 11, 42964–42974. [Google Scholar] [CrossRef]
- Tahara, K.; Kobayashi, M.; Yoshida, S.; Onodera, R.; Inoue, N.; Takeuchi, H. Effects of cationic liposomes with stearylamine against virus infection. Int. J. Pharm. 2018, 543, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Liu, J.; Valyi-Nagy, T.; Shukla, D. Anti-heparan sulfate peptides that block herpes simplex virus infection in vivo. J. Biol. Chem. 2011, 286, 25406–25415. [Google Scholar] [CrossRef]
- Figueroa, S.M.; Veser, A.; Abstiens, K.; Fleischmann, D.; Beck, S.; Goepferich, A. Influenza A virus mimetic nanoparticles trigger selective cell uptake. Proc. Natl. Acad. Sci. USA 2019, 116, 9831–9836. [Google Scholar] [CrossRef]
- Cavalli, R.; Donalisio, M.; Civra, A.; Ferruti, P.; Ranucci, E.; Trotta, F.; Lembo, D. Enhanced antiviral activity of Acyclovir loaded into β-cyclodextrin-poly(4-acryloylmorpholine) conjugate nanoparticles. J. Control. Release 2009, 137, 116–122. [Google Scholar] [CrossRef]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef]
- Chakravarty, M.; Vora, A. Nanotechnology-based antiviral therapeutics. Drug Deliv. Transl. Res. 2020, 11, 748–787. [Google Scholar] [CrossRef]
- Lategan, K.; Alghadi, H.; Bayati, M.; de Cortalezzi, M.F.; Pool, E. Effects of graphene oxide nanoparticles on the immune system biomarkers produced by RAW 264.7 and human whole blood cell cultures. Nanomaterials 2018, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Speshock, J.L.; Murdock, R.C.; Braydich-Stolle, L.K.; Schrand, A.M.; Hussain, S.M. Interaction of silver nanoparticles with Tacaribe virus. J. Nanobiotechnol. 2010, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Soiza, R.L.; Donaldson, A.I.C.; Myint, P.K. Vaccine against arteriosclerosis: An update. Ther. Adv. Vaccines 2018, 9, 259–261. [Google Scholar]
- Chen, L.; Liang, J. An overview of functional nanoparticles as novel emerging antiviral therapeutic agents. Mater. Sci. Eng. C 2020, 112, 110924. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.T. Interfering viral-like particles inhibit SARS-CoV-2 replication. Nat. Rev. Drug Discov. 2022, 21, 19. [Google Scholar] [CrossRef] [PubMed]
- Ishida, N. Laboratory diagnosis of virus diseases. Boei. Eisei. 1962, 9, 330–333. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Báez-Santos, Y.M.; John, S.E.S.; Mesecar, A.D. The SARS-coronavirus papain-like protease: Structure, function and inhibition by designed antiviral compounds. Antivir. Res. 2015, 115, 21–38. [Google Scholar] [CrossRef]
- Ishida, T. Review on The Role of Zn2+ Ions in Viral Pathogenesis and the Effect of Zn2+ Ions for Host Cell-Virus Growth Inhibition. Am. J. Biomed. Sci. Res. 2019, 2, 28–37. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef]
- Sagripanti, J.L.; Routson, L.B.; Lytle, C.D. Virus inactivation by copper or iron ions alone and in the presence of peroxide. Appl. Environ. Microbiol. 1993, 59, 4374–4376. [Google Scholar] [CrossRef] [PubMed]
- Noyce, J.O.; Michels, H.; Keevil, C.W. Inactivation of influenza A virus on copper versus stainless steel surfaces. Appl. Environ. Microbiol. 2007, 73, 2748–2750. [Google Scholar] [CrossRef] [PubMed]
- te Velthuis, A.J.W.; van den Worml, S.H.E.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn2+ inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, H.; Tavakoli, A.; Moradi, A.; Tabarraei, A.; Bokharaei-Salim, F.; Zahmatkeshan, M.; Farahmand, M.; Javanmard, D.; Kiani, S.J.; Esghaei, M.; et al. Inhibition of H1N1 influenza virus infection by zinc oxide nanoparticles: Another emerging application of nanomedicine. J. Biomed. Sci. 2019, 26, 70. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.; Dearnley, M.; Hinton, T.M. Polymers in the Delivery of siRNA for the Treatment of Virus Infections; Springer International Publishing: New York, NY, USA, 2017; Volume 375, ISBN 4106101701276. [Google Scholar]
- Ting, D.; Dong, N.; Fang, L.; Lu, J.; Bi, J.; Xiao, S.; Han, H. Multisite inhibitors for enteric coronavirus: Antiviral cationic carbon dots based on curcumin. ACS Appl. Nano Mater. 2018, 1, 5451–5459. [Google Scholar] [CrossRef]
- Hu, C.M.J.; Chen, Y.T.; Fang, Z.S.; Chang, W.S.; Chen, H.W. Antiviral efficacy of nanoparticulate vacuolar ATPase Inhibitors Against Influenza virus infection. Int. J. Nanomed. 2018, 13, 8579–8593. [Google Scholar] [CrossRef]
- Croci, R.; Bottaro, E.; Chan, K.W.K.; Watanabe, S.; Pezzullo, M.; Mastrangelo, E.; Nastruzzi, C. Liposomal Systems as Nanocarriers for the Antiviral Agent Ivermectin. Int. J. Biomater. 2016, 2016, 8043983. [Google Scholar] [CrossRef]
- Xing-Guo, Z.; Jing, M.; Min-Wei, L.; Sai-Ping, J.; Fu-Qiang, H.; Yong-Zhong, D. Solid lipid nanoparticles loading adefovir dipivoxil for antiviral therapy. J. Zhejiang Univ. Sci. B 2008, 9, 506–510. [Google Scholar] [CrossRef]
- Rogers, J.V.; Parkinson, C.V.; Choi, Y.W.; Speshock, J.L.; Hussain, S.M. A preliminary assessment of silver nanoparticle inhibition of monkeypox virus plaque formation. Nanoscale Res. Lett. 2008, 3, 129–133. [Google Scholar] [CrossRef]
- López, N.; Jácamo, R.; Franze-Fernández, M.T. Transcription and RNA Replication of Tacaribe Virus Genome and Antigenome Analogs Require N and L Proteins: Z Protein Is an Inhibitor of These Processes. J. Virol. 2001, 75, 12241–12251. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Nayak, M.; Sahoo, G.C.; Pandey, K.; Sarkar, M.C.; Ansari, Y.; Das, V.N.R.; Topno, R.K.; Bhawna; Madhukar, M.; et al. Iron oxide nanoparticles based antiviral activity of H1N1 influenza A virus. J. Infect. Chemother. 2019, 25, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Ramphul, K.; Mejias, S.G. Coronavirus Disease: A Review of a New Threat to Public Health. Cureus 2020, 20, 2019–2020. [Google Scholar] [CrossRef] [PubMed]
- Krishnaraj, R.; Chandran, S.; Pal, P.; Berchmans, S. Investigations on the Antiretroviral Activity of Carbon Nanotubes Using Computational Molecular Approach. Comb. Chem. High Throughput Screen. 2014, 17, 531–535. [Google Scholar] [CrossRef]
- Martinez, Z.S.; Castro, E.; Seong, C.S.; Cerón, M.R.; Echegoyen, L.; Llano, M. Fullerene derivatives strongly inhibit HIV-1 replication by affecting virus maturation without impairing protease activity. Antimicrob. Agents Chemother. 2016, 60, 5731–5741. [Google Scholar] [CrossRef]
- Rosen, Y.; Gurman, P. Carbon Nanotubes for Drug Delivery Applications. In Nanotechnology and Drug Delivery; CRC Press: Boca Raton, FL, USA, 2014; pp. 233–248. [Google Scholar] [CrossRef]
- Steinbach, J.M.; Weller, C.E.; Booth, C.J.; Saltzman, W.M. Polymer nanoparticles encapsulating siRNA for treatment of HSV-2 genital infection. J. Control. Release 2012, 162, 102–110. [Google Scholar] [CrossRef]
- Ye, S.; Shao, K.; Li, Z.; Guo, N.; Zuo, Y.; Li, Q.; Lu, Z.; Chen, L.; He, Q.; Han, H. Antiviral Activity of Graphene Oxide: How Sharp Edged Structure and Charge Matter. ACS Appl. Mater. Interfaces 2015, 7, 21578–21579. [Google Scholar] [CrossRef]
- Cagno, V.; Andreozzi, P.; D’alicarnasso, M.; Silva, P.J.; Mueller, M.; Galloux, M.; Goffic, R.L.; Jones, S.T.; Vallino, M.; Hodek, J.; et al. Broad-spectrum non-toxic antiviral nanoparticles with a virucidal inhibition mechanism. Nat. Mater. 2018, 17, 195–203. [Google Scholar] [CrossRef]
- Bromberg, L.; Bromberg, D.J.; Hatton, T.A.; Bandín, I.; Concheiro, A.; Alvarez-Lorenzo, C. Antiviral properties of polymeric aziridine- and biguanide-modified core-shell magnetic nanoparticles. Langmuir 2012, 28, 4548–4558. [Google Scholar] [CrossRef]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef]
- Ghaffari, E.; Rezatofighi, S.E.; Ardakani, M.R.; Rastegarzadeh, S. Delivery of antisense peptide nucleic acid by gold nanoparticles for the inhibition of virus replication. Nanomedicine 2019, 14, 1827–1840. [Google Scholar] [CrossRef] [PubMed]
- Baram-Pinto, D.; Shukla, S.; Gedanken, A.; Sarid, R. Inhibition of HSV-1 Attachment, Entry, and Cell-to-Cell Spread by Functionalized Multivalent Gold Nanoparticles. Small 2010, 6, 1044–1050. [Google Scholar] [CrossRef]
- Szabó, G.T.; Mahiny, A.J.; Vlatkovic, I. COVID-19 mRNA vaccines: Platforms and current developments. Mol. Ther. 2022, 30, 1850–1868. [Google Scholar] [CrossRef] [PubMed]
- Maduray, K.; Parboosing, R. Metal Nanoparticles: A Promising Treatment for Viral and Arboviral Infections. Biol. Trace Elem. Res. 2020, 199, 3159–3176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Guo, H.; Guo, J.T.; Cuconati, A.; Mehta, A.; Block, T.M. Hepatitis B virus e antigen production is dependent upon covalently closed circular (ccc) DNA in HepAD38 cell cultures and may serve as a cccDNA surrogate in antiviral screening assays. Antivir. Res. 2006, 72, 116–124. [Google Scholar] [CrossRef]
- Sametband, M.; Kalt, I.; Gedanken, A.; Sarid, R. Herpes simplex virus type-1 attachment inhibition by functionalized graphene oxide. ACS Appl. Mater. Interfaces 2014, 6, 1228–1235. [Google Scholar] [CrossRef]
- Martínez-Gualda, B.; Sun, L.; Martí-Marí, O.; Mirabelli, C.; Delang, L.; Neyts, J.; Schols, D.; Camarasa, M.J.; San-Félix, A. Modifications in the branched arms of a class of dual inhibitors of HIV and EV71 replication expand their antiviral spectrum. Antivir. Res. 2019, 168, 210–214. [Google Scholar] [CrossRef]
- Jamali, A.; Mottaghitalab, F.; Abdoli, A.; Dinarvand, M.; Esmailie, A.; Kheiri, M.T.; Atyabi, F. Inhibiting influenza virus replication and inducing protection against lethal influenza virus challenge through chitosan nanoparticles loaded by siRNA. Drug Deliv. Transl. Res. 2018, 8, 12–20. [Google Scholar] [CrossRef]
- Yuan, M.; Das, R.; Ghannam, R.; Wang, Y.; Reboud, J.; Fromme, R.; Moradi, F.; Heidari, H. Electronic Contact Lens: A Platform for Wireless Health Monitoring Applications. Adv. Intell. Syst. 2020, 2, 1900190. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, S. Mechanics of interfacial delamination in epidermal electronics systems. J. Appl. Mech. Trans. ASME 2014, 81, 1–3. [Google Scholar] [CrossRef]
- Baldassi, D.; Ambike, S.; Feuerherd, M.; Cheng, C.C.; Peeler, D.J.; Feldmann, D.P.; Porras-Gonzalez, D.L.; Wei, X.; Keller, L.A.; Kneidinger, N.; et al. Inhibition of SARS-CoV-2 replication in the lung with siRNA/VIPER polyplexes. J. Control. Release 2022, 345, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Simion, V.; Stan, D.; Constantinescu, C.A.; Deleanu, M.; Dragan, E.; Tucureanu, M.M.; Gan, A.M.; Butoi, E.; Constantin, A.; Manduteanu, I.; et al. Conjugation of curcumin-loaded lipid nanoemulsions with cell-penetrating peptides increases their cellular uptake and enhances the anti-inflammatory effects in endothelial cells. J. Pharm. Pharmacol. 2016, 68, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Garg, T.; Aman, A.; Panchal, K.; Sharma, R.; Kumar, S.; Markandeywar, T. Nanogel—An advanced drug delivery tool: Current and future. Artif. Cells Nanomed. Biotechnol. 2016, 44, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Gigliobianco, M.R.; Censi, R.; Di Martino, P. Polymeric nanocapsules as nanotechnological alternative for drug delivery system: Current status, challenges and opportunities. Nanomaterials 2020, 10, 847. [Google Scholar] [CrossRef] [PubMed]
- Argenziano, M.; Foglietta, F.; Canaparo, R.; Spagnolo, R.; Pepa, C.D.; Caldera, F.; Trotta, F.; Serpe, L.; Cavalli, R. Biological effect evaluation of glutathione-responsive cyclodextrin-based nanosponges: 2D and 3D studies. Molecules 2020, 25, 2775. [Google Scholar] [CrossRef]
- Silva, M.M.; Calado, R.; Marto, J.; Bettencourt, A.; Almeida, A.J.; Gonçalves, L.M.D. Chitosan nanoparticles as a mucoadhesive drug delivery system for ocular administration. Mar. Drugs 2017, 15, 370. [Google Scholar] [CrossRef]
- Patil, N.H.; Devarajan, P.V. Insulin-loaded alginic acid nanoparticles for sublingual delivery. Drug Deliv. 2016, 23, 429–436. [Google Scholar] [CrossRef]
- Menzel, C.; Jelkmann, M.; Laffleur, F.; Bernkop-Schnürch, A. Nasal drug delivery: Design of a novel mucoadhesive and in situ gelling polymer. Int. J. Pharm. 2017, 517, 196–202. [Google Scholar] [CrossRef]
- Agarwal, T.; Narayana, S.N.G.H.; Pal, K.; Pramanik, K.; Giri, S.; Banerjee, I. Calcium alginate-carboxymethyl cellulose beads for colon-targeted drug delivery. Int. J. Biol. Macromol. 2015, 75, 409–417. [Google Scholar] [CrossRef]
- Kotla, N.G.; Chandrasekar, B.; Rooney, P.; Sivaraman, G.; Larrañaga, A.; Krishna, K.V.; Pandit, A.; Rochev, Y. Biomimetic Lipid-Based Nanosystems for Enhanced Dermal Delivery of Drugs and Bioactive Agents. ACS Biomater. Sci. Eng. 2017, 3, 1262–1272. [Google Scholar] [CrossRef]
- Li, Q.; Lai, K.L.; Chan, P.S.; Leung, S.C.; Li, H.Y.; Fang, Y.; To, K.K.W.; Choi, C.H.J.; Gao, Q.Y.; Lee, T.W.Y. Micellar delivery of dasatinib for the inhibition of pathologic cellular processes of the retinal pigment epithelium. Colloids Surf. B Biointerfaces 2016, 140, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Jain, K.; Mehra, N.K.; Jain, N.K. Development and characterization of surface engineered PPI dendrimers for targeted drug delivery. Artif. Cells Nanomed. Biotechnol. 2017, 45, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Prusty, K.; Swain, S.K. Nano silver decorated polyacrylamide/dextran nanohydrogels hybrid composites for drug delivery applications. Mater. Sci. Eng. C 2018, 85, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Ni, R.; Zhao, J.; Liu, Q.; Liang, Z.; Muenster, U.; Mao, S. Nanocrystals embedded in chitosan-based respirable swellable microparticles as dry powder for sustained pulmonary drug delivery. Eur. J. Pharm. Sci. 2017, 99, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Pramanik, A.; Tchounwou, C.; Pedraza, F.; Crouch, R.A.; Chavva, S.R.; Vangara, A.; Sinha, S.S.; Jones, S.; Sardar, D.; et al. Multifunctional Biocompatible Graphene Oxide Quantum Dots Decorated Magnetic Nanoplatform for Efficient Capture and Two-Photon Imaging of Rare Tumor Cells. ACS Appl. Mater. Interfaces 2015, 7, 10935–10943. [Google Scholar] [CrossRef]
- Sillman, B.; Bade, A.N.; Dash, P.K.; Bhargavan, B.; Kocher, T.; Mathews, S.; Su, H.; Kanmogne, G.D.; Poluektova, L.Y.; Gorantla, S.; et al. Creation of a long-acting nanoformulated dolutegravir. Nat. Commun. 2018, 9, 443. [Google Scholar] [CrossRef]
- Dhoke, D.M.; Basaiyye, S.S.; Khedekar, P.B. Development and characterization of L-HSA conjugated PLGA nanoparticle for hepatocyte targeted delivery of antiviral drug. J. Drug Deliv. Sci. Technol. 2018, 47, 77–94. [Google Scholar] [CrossRef]
- Makwana, V.; Jain, R.; Patel, K.; Nivsarkar, M.; Joshi, A. Solid lipid nanoparticles (SLN) of Efavirenz as lymph targeting drug delivery system: Elucidation of mechanism of uptake using chylomicron flow blocking approach. Int. J. Pharm. 2015, 495, 439–446. [Google Scholar] [CrossRef]
- Renu, S.; Feliciano-Ruiz, N.; Ghimire, S.; Han, Y.; Schrock, J.; Dhakal, S.; Patil, V.; Krakowka, S.; Renukaradhya, G.J. Poly(I:C) augments inactivated influenza virus-chitosan nanovaccine induced cell mediated immune response in pigs vaccinated intranasally. Vet. Microbiol. 2020, 242, 108611. [Google Scholar] [CrossRef]
- Mori, Y.; Ono, T.; Miyahira, Y.; Nguyen, V.Q.; Matsui, T.; Ishihara, M. Antiviral activity of silver nanoparticle/chitosan composites against H1N1 influenza A virus. Nanoscale Res. Lett. 2013, 8, 93. [Google Scholar] [CrossRef]
- Coleman, C.M.; Venkataraman, T.; Liu, Y.V.; Glenn, G.M.; Smith, G.E.; Flyer, D.C.; Frieman, M.B. MERS-CoV spike nanoparticles protect mice from MERS-CoV infection. Vaccine 2017, 35, 1586–1589. [Google Scholar] [CrossRef]
- Roux, X.; Dubuquoy, C.; Durand, G.; Tran-Tolla, T.L.; Castagné, N.; Bernard, J.; Petit-Camurdan, A.; Eléouët, J.F.; Riffault, S. Sub-nucloecapsid nanoparticles: A nasal vaccine against respiratory syncytial virus. PLoS ONE 2008, 3, e1766. [Google Scholar] [CrossRef]
- Favaro, M.T.P.; Rodrigues-Jesus, M.J.; Venceslau-Carvalho, A.A.; Alves, R.P.D.S.; Pereira, L.R.; Pereira, S.S.; Andreata-Santos, R.; de Souza Ferreira, L.C. Nanovaccine based on self-assembling nonstructural protein 1 boosts antibody responses to Zika virus. Nanomed. Nanotechnol. Biol. Med. 2021, 32, 102334. [Google Scholar] [CrossRef]
- Hozáková, L.; Vokatá, B.; Ruml, T.; Ulbrich, P. Targeting the Virus Capsid as a Tool to Fight RNA Viruses. Viruses 2022, 14, 174. [Google Scholar] [CrossRef]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M.; Bachmann, M.F. Interaction of viral capsid-derived virus-like particles (VLPs) with the innate immune system. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, A.; Rassek, O.; Vrohlings, M.; Marrero-Nodarse, A.; Moehle, K.; Robinson, J.A.; Ghasparian, A. An epitope-specific chemically defined nanoparticle vaccine for respiratory syncytial virus. Npj Vaccines 2021, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Roland, V., Jr.; Lori, M.; Millner, M.W.L. Myocardium extract from suckling rat HHS Public Access. Physiol. Behav. 2019, 176, 139–148. [Google Scholar]
- Yu, F.; Ao, M.; Zheng, X.; Li, N.; Xia, J.; Li, Y.; Li, D.; Hou, Z.; Qi, Z.; Chen, X.D. PEG-lipid-PLGA hybrid nanoparticles loaded with berberine-phospholipid complex to facilitate the oral delivery efficiency. Drug Deliv. 2017, 24, 825–833. [Google Scholar] [CrossRef]
- Zhang, N.; Chittasupho, C.; Duangrat, C.; Siahaan, T.J.; Berkland, C. PLGA Nanoparticle−Peptide Conjugate Effectively Targets Intercellular Cell-Adhesion Molecule-1. Bioconjug Chem. 2008, 19, 145–152. [Google Scholar] [CrossRef]
- Emami, J.; Maghzi, P.; Hasanzadeh, F.; Sadeghi, H.; Mirian, M.; Rostami, M. PLGA-PEG-RA-based polymeric micelles for tumor targeted delivery of irinotecan. Pharm. Dev. Technol. 2018, 23, 41–54. [Google Scholar] [CrossRef]
- Shand, H.; Dutta, S.; Rajakumar, S.; James Paulraj, S.; Mandal, A.K.; KT, R.D.; Ghorai, S. New Age Detection of Viruses: The Nano-Biosensors. Front. Nanotechnol. 2022, 3, 814550. [Google Scholar] [CrossRef]
- Günl, F.; Mecate-zambrano, A.; Rehländer, S.; Hinse, S.; Ludwig, S.; Brunotte, L. Shooting at a Moving Target—Effectiveness and Emerging Challenges for SARS-CoV-2 Vaccine Development. Vaccines 2021, 9, 1052. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Mohammadifar, M.; Choi, S. A single-use, self-powered, paper-based sensor patch for detection of exercise-induced hypoglycemia. Micromachines 2017, 8, 265. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, S.R.; Kil, H.J.; Kim, Y.C.; Park, J.W. Highly Conformable, Transparent Electrodes for Epidermal Electronics. Nano Lett. 2018, 18, 4531–4540. [Google Scholar] [CrossRef]
- Zhang, Z.; Michniak-Kohn, B.B. Tissue engineered human skin equivalents. Pharmaceutics 2012, 4, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Mutashar, S.; Hannan, M.A.; Samad, S.A.; Hussain, A. Analysis and optimization of spiral circular inductive coupling link for bio-implanted applications on air and within human tissue. Sensors 2014, 14, 11522–11541. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Lee, S.Y.; Permana, H.; Ghorbani, K.; Cosic, I. Developing a wireless implantable body sensor network in MICS band. IEEE Trans. Inf. Technol. Biomed. 2011, 15, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Roy, K.J.; Rahman, K.M.A.; Aktar, M.K.; Kafi, M.A.; Islam, M.S.; Rahman, M.B.; Islam, M.R.; Hossain, K.S.; Rahman, M.M.; et al. Adhesion and proliferation of living cell on surface functionalized with glycine nanostructures. Nano Sel. 2021, 2, 188–200. [Google Scholar] [CrossRef]
- Rajan, V.; Sivaraman, G.K.; Vijayan, A.; Elangovan, R.; Prendiville, A.; Bachmann, T.T. Genotypes and phenotypes of methicillin-resistant staphylococci isolated from shrimp aquaculture farms. Environ. Microbiol. Rep. 2022, 14, 391–399. [Google Scholar] [CrossRef]
- Kafi, M.A.; Aktar, M.K.; Heidari, H. Mammalian Cell-Based Electrochemical Sensor for Label-Free Monitoring of Analytes. Smart Sens. Environ. Med. Appl. 2020, 9, 43–60. [Google Scholar] [CrossRef]
- Roy, K.J.; Rahman, A.; Hossain, K.; Rahman, B. Antibacterial Investigation of Silver Nanoparticle against Staphylococcus, E. coli and Salmonella Isolated from Selected Live Bird Markets. Appl. Microbiol. Open Access 2020, 6, 173. [Google Scholar]
- Soliman, G.M. Nanoparticles as safe and effective delivery systems of antifungal agents: Achievements and challenges. Int. J. Pharm. 2017, 523, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, S.; Falanga, A.; Vitiello, M.; Cantisani, M.; Marra, V.; Galdiero, M. Silver nanoparticles as potential antiviral agents. Molecules 2011, 16, 8894–8918. [Google Scholar] [CrossRef]
- Lysenko, V.; Lozovski, V.; Lokshyn, M.; Gomeniuk, Y.V.; Dorovskih, A.; Rusinchuk, N.; Pankivska, Y.; Povnitsa, O.; Zagorodnya, S.; Tertykh, V.; et al. Nanoparticles as antiviral agents against adenoviruses. Adv. Nat. Sci. Nanosci. Nanotechnol. 2018, 9, 025021. [Google Scholar] [CrossRef]
- Milovanovic, M.; Arsenijevic, A.; Milovanovic, J.; Kanjevac, T.; Arsenijevic, N. Nanoparticles in Antiviral Therapy; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780323527347. [Google Scholar]
- Hua, S.; Wu, S.Y. The use of lipid-based nanocarriers for targeted pain therapies. Front. Pharmacol. 2013, 4, 143. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F.; Hong, V.; Spoerke, E.D.; Lu, P.; Breitenkamp, K.; Finn, M.G.; Manchester, M. Buckyballs meet viral nanoparticles: Candidates for biomedicine. J. Am. Chem. Soc. 2009, 131, 17093–17095. [Google Scholar] [CrossRef]
- Johnsen, K.B.; Gudbergsson, J.M.; Skov, M.N.; Pilgaard, L.; Moos, T.; Duroux, M. A comprehensive overview of exosomes as drug delivery vehicles—Endogenous nanocarriers for targeted cancer therapy. Biochim. Biophys. Acta—Rev. Cancer 2014, 1846, 75–87. [Google Scholar] [CrossRef]
- Prompetchara, E.; Ketloy, C.; Palaga, T. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic. Asian Pac. J. Allergy Immunol. 2020, 38, 1–9. [Google Scholar] [CrossRef]
- Duan, K.; Liu, B.; Li, C.; Zhang, H.; Yu, T.; Qu, J.; Zhou, M.; Chen, L.; Meng, S.; Hu, Y.; et al. Effectiveness of convalescent plasma therapy in severe COVID-19 patients. Proc. Natl. Acad. Sci. USA 2020, 117, 9490–9496. [Google Scholar] [CrossRef]
- Brogden, K.A. The sweet spot: Defining virus–sialic acid interactions Jennifer. Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Pescatori, M.; Bedognetti, D.; Venturelli, E.; Ménard-Moyon, C.; Bernardini, C.; Muresu, E.; Piana, A.; Maida, G.; Manetti, R.; Sgarrella, F.; et al. Functionalized carbon nanotubes as immunomodulator systems. Biomaterials 2013, 34, 4395–4403. [Google Scholar] [CrossRef] [PubMed]
- Loutfy, S.A.; Elberry, M.H.; Farroh, K.Y.; Taha Mohamed, H.; Mohamed, A.A.; Mohamed, E.B.; Hassan, A.; Faraag, I.; Mousa, S.A. Antiviral Activity of Chitosan Nanoparticles Encapsulating Curcumin Against Hepatitis C Virus Genotype 4a in Human Hepatoma Cell Lines. Int. J. Nanomed. 2020, 2020, 15–2699. [Google Scholar] [CrossRef] [PubMed]
- De Medeiros, A.M.Z.; Khan, L.U.; Gabriela, H.; Ospina, C.A.; Alves, O.L.; Lúcia, V.; Castro, D.; Martinez, D.S.T. Ecotoxicology and Environmental Safety Graphene oxide-silver nanoparticle hybrid material: An integrated nanosafety study in zebrafish embryos. Ecotoxicol. Environ. Saf. 2021, 209, 111776. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regional Variant | Scientific Name | WHO Name | RBD Mutation | Spreading Nature | Reduced Neutralizing Ability | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mutation Residues | |||||||||||
| K 417 | L 452 | K 477 | T 478 | E 484 | Q 498 | N 501 | |||||
| United Kingdom, September 2020 | B.1.1.7 | Alpha | K | L | K | T | E | Q | Y | + | 3-fold (Pfizer/BioNTech) and 6-fold (AstraZeneca) |
| South Africa, October 2020 | B.1.351 | Beta | N | L | K | T | K | Q | Y | ++ | ≤86-fold (AstraZeneca) ≤6.5, ≤8.6, and ≤1.6-fold (Moderna, Pfizer-BioNTech, and Sinopharm) |
| Brazil and Japan, December 2020 | P.1 | Gamma | T | L | K | T | K | Q | Y | ++ | 6.7- and 4.5-fold (Pfizer-BioNTech and Moderna) |
| India, December 2020 | B.1.617.2 | Delta | N | R | K | K | Q | Q | N | + | 2.5-fold (Pfizer/BioNTech, Moderna, and Janssen vaccine) |
| South Africa, November 2021 | B.1.1.529 | Omicron | N | L | N | K | A | R | Y | +++ | 41-fold (Pfizer-BioNTech) |
| Nanoparticles | Targeted Site | Action | Reference |
|---|---|---|---|
| Nanoemulsion | Monocytes, lung cells, cancer cells | Prevention from hydrolysis and oxidation allows for durable action Minimize vascular inflammation | [128] |
| Nanogel | Blood cell, THP-1, and HaCaT cell lines | Stability in blood circulation Enhanced anti-inflammatory action through inhibition of LOX and COX activities in cells | [129] |
| Nano-capsule (resveratrol-charged lipid-core-nano-capsule) | Cancer cell, HT29 cell lines | Controlled drug release Destruction of colon cancer cells Enhanced anticancer activity in HT29 cancer cells | [130] |
| Nanosponges (cyclodextrin drug-coated nanosponges) | Tumor cells | Increased anti-tumor activities | [131] |
| Chitosan | Buccal, intestinal, nasal, ocular, and pulmonary cells | Interaction with the ocular mucosa and prolonged release of the antibiotic Enhanced half-life of the drug in the eyes | [132] |
| Alginate | Sublingual cells | Dip in serum glucose levels and increase in serum insulin levels in diabetic rats | [133] |
| Xanthan gum | Buccal cells | Increased adhesion to buccal cells and release of tannin in buccal mucosa to treat diarrhea | [134] |
| Cellulose | Colon, nasal mucosa | Sustained release of cellulose nanocrystals Calcium alginate beads with carboxymethylcellulose (CMC)-loaded 5-fuoroacyl (5-FU); 90% release of 5-FU encapsulated in the beads Increased permeation of acyclovir into the nasal mucosa | [135] |
| Liposomes | cell membranes | Increased opsonization and immunogenicity of RES (reticuloendothelial system) Boost in drug delivery efficiency of the liposomes | [136] |
| Polymeric micelles | ARPE-19 cells, Eye tissues | Enhancement of cell proliferation, attachment, and relocation Inhibition of rear eye tissue damage | [137] |
| Dendrimers | Cancer cells and MCF-7 cells | Folate-attached poly-l-lysine dendrimers control cancer cell Increased concentration of doxorubicin in the tumor Increased cell uptake and low cytotoxicity in MCF-7 cell lines | [138] |
| Inorganic (silver, gold, iron oxide, and silica) nanoparticles | Bacteria and virus cell membranes | Control of release through biological stimuli or light activation | [139] |
| Nanocrystals | Pulmonary tissues | Enhanced dissolution velocity and increased glueyness to surface/cell membranes Continuous release of nanoparticles helps with swelling and shows muco-adhesive potential Enhanced inhalation efficacy under disease conditions | [140] |
| Quantum dots | Bone marrow cells, liver, cancer, and tumor cells | Diffusion into the entire bone marrow and labeling of rare populations of cells, such as hematopoietic and progenitor cells Attachment of an anti-GPC3-antibody to the nanoplatform results in selective separation of HepG2 hepatocellular carcinoma cells from infected blood samples | [141] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.; Roy, K.J.; Deb, G.K.; Ha, T.; Rahman, S.; Aktar, M.K.; Ali, M.I.; Kafi, M.A.; Choi, J.-W. Nano-Enabled Antivirals for Overcoming Antibody Escaped Mutations Based SARS-CoV-2 Waves. Int. J. Mol. Sci. 2023, 24, 13130. https://doi.org/10.3390/ijms241713130
Rahman A, Roy KJ, Deb GK, Ha T, Rahman S, Aktar MK, Ali MI, Kafi MA, Choi J-W. Nano-Enabled Antivirals for Overcoming Antibody Escaped Mutations Based SARS-CoV-2 Waves. International Journal of Molecular Sciences. 2023; 24(17):13130. https://doi.org/10.3390/ijms241713130
Chicago/Turabian StyleRahman, Aminur, Kumar Jyotirmoy Roy, Gautam Kumar Deb, Taehyeong Ha, Saifur Rahman, Mst. Khudishta Aktar, Md. Isahak Ali, Md. Abdul Kafi, and Jeong-Woo Choi. 2023. "Nano-Enabled Antivirals for Overcoming Antibody Escaped Mutations Based SARS-CoV-2 Waves" International Journal of Molecular Sciences 24, no. 17: 13130. https://doi.org/10.3390/ijms241713130
APA StyleRahman, A., Roy, K. J., Deb, G. K., Ha, T., Rahman, S., Aktar, M. K., Ali, M. I., Kafi, M. A., & Choi, J. -W. (2023). Nano-Enabled Antivirals for Overcoming Antibody Escaped Mutations Based SARS-CoV-2 Waves. International Journal of Molecular Sciences, 24(17), 13130. https://doi.org/10.3390/ijms241713130