
Phylogenetic Relationships and Next-Generation Barcodes in the Genus Torreya Reveal a High Proportion of Misidentified Cultivated Plants

Abstract
:1. Introduction
2. Results
2.1. Plastomes, Mitochondrial Genes and nrDNA Cistrons of Torreya
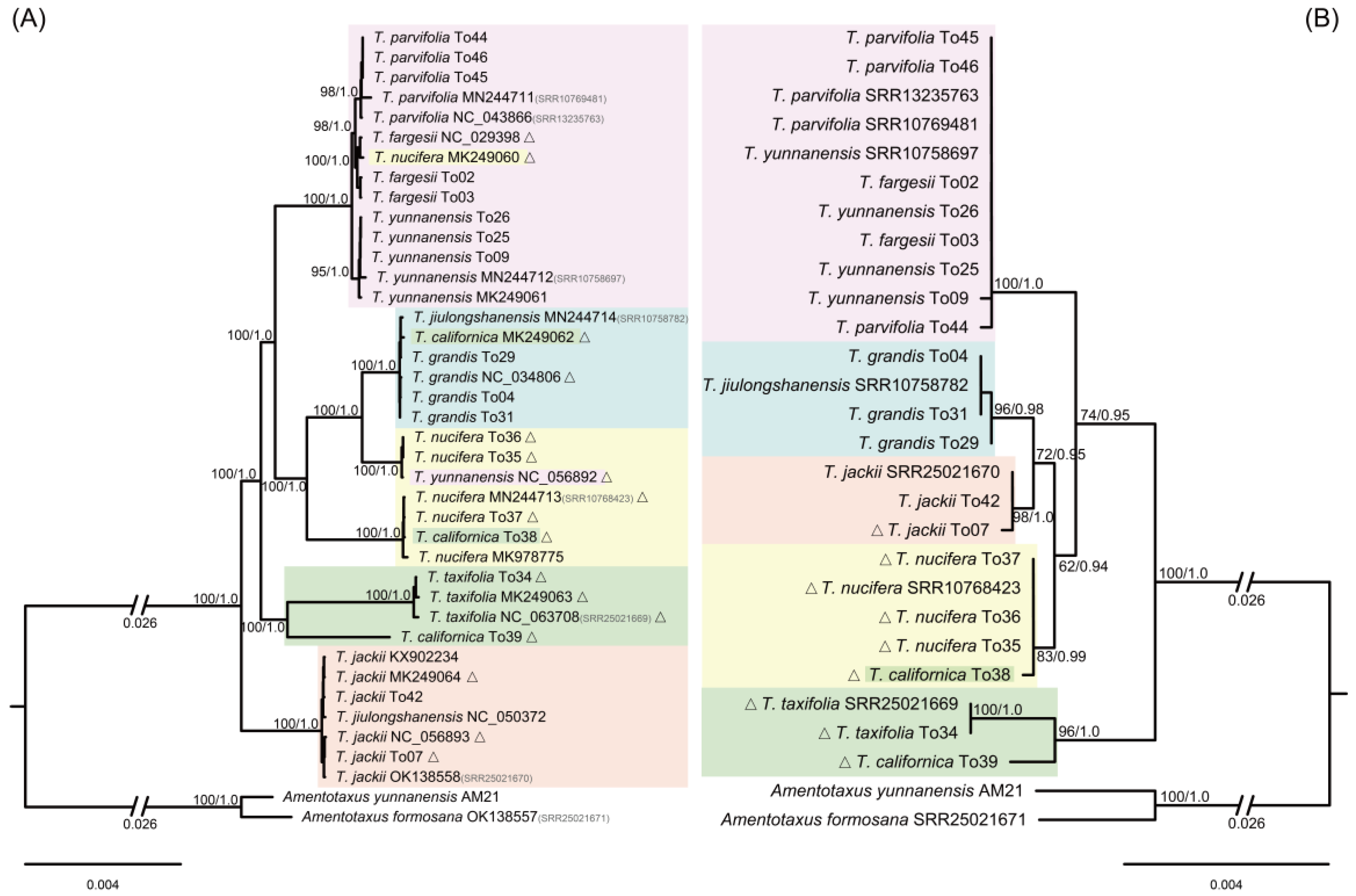
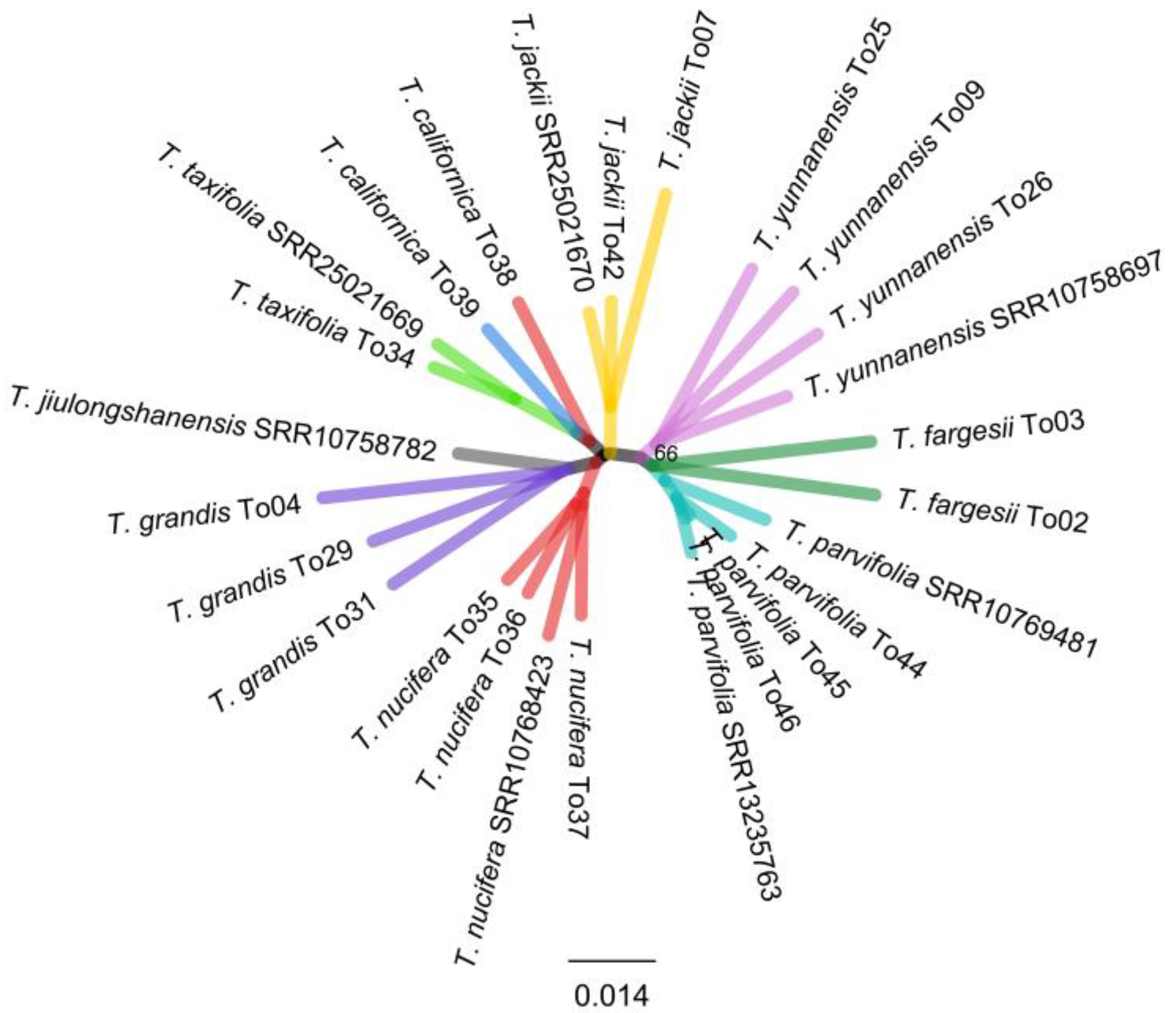
2.2. Phylogenetic Analyses—Species Delineation and Sample Identities
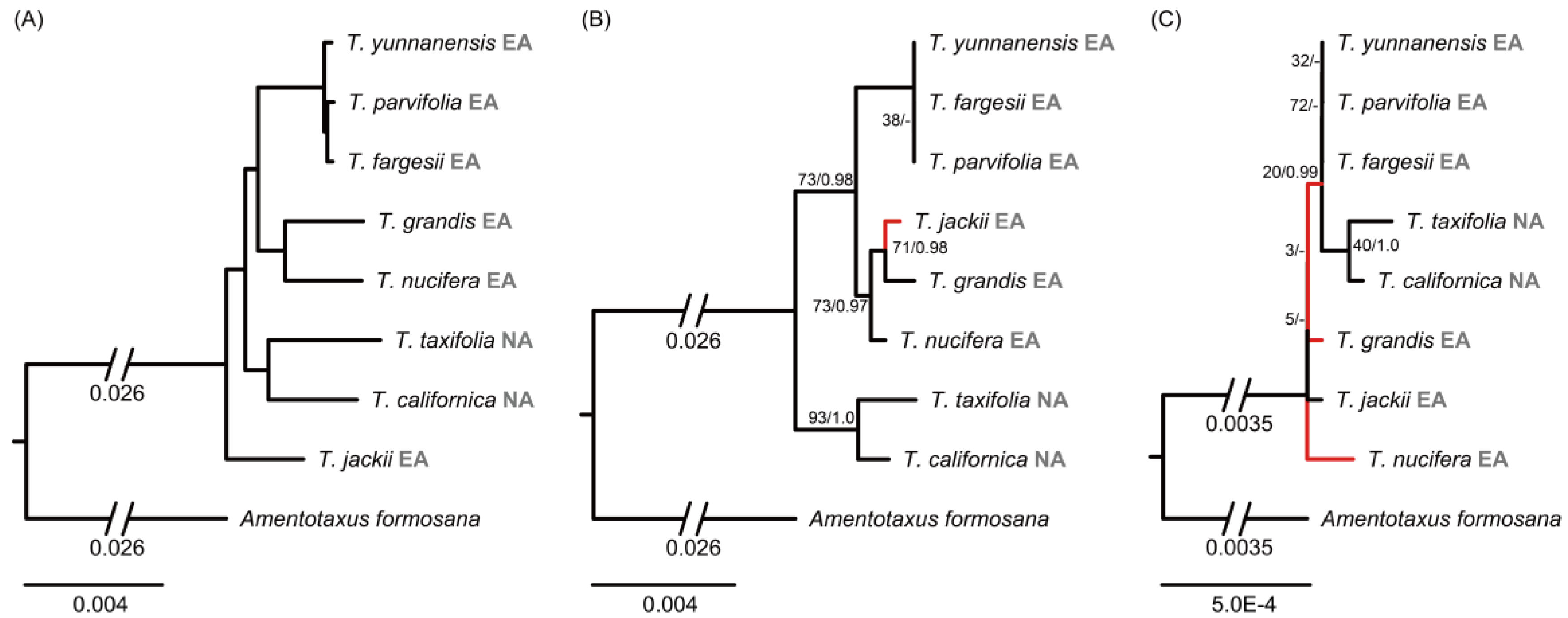
2.3. Discordance between Plastid, nrDNA Cistron, and Mitochondrial Phylogenies
3. Discussion
3.1. Species Delimitation and Correction of Misidentified Accessions
3.2. Hybrid Origin of T. jiulongshanensis
3.3. Phylogenetic Relationships and Conflicts among the Three Genomic Phylogenies
4. Materials and Methods
4.1. Sample Collection, DNA Extraction and Genome Skimming Sequencing
4.2. Assembly and Annotation
4.3. Phylogenetic Analyses
4.4. Skmer Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kane, N.; Sveinsson, S.; Dempewolf, H.; Yang, J.Y.; Zhang, D.; Engels, J.M.; Cronk, Q. Ultra-barcoding in cacao (Theobroma spp.; Malvaceae) using whole chloroplast genomes and nuclear ribosomal DNA. Am. J. Bot. 2012, 99, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Ruhsam, M.; Rai, H.S.; Mathews, S.; Ross, T.G.; Graham, S.W.; Raubeson, L.A.; Mei, W.; Thomas, P.I.; Gardner, M.F.; Ennos, R.A. Does complete plastid genome sequencing improve species discrimination and phylogenetic resolution in Araucaria? Mol. Ecol. Resour. 2015, 15, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.D.; Jin, J.J.; Chen, S.Y.; Chase, M.W.; Soltis, D.E.; Li, H.T.; Yang, J.B.; Li, D.Z.; Yi, T.S. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. New Phytol. 2017, 214, 1355–1367. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Niu, Z.; Xue, Q.; Wang, H.; Xie, X.; Ding, X. Accurate authentication of Dendrobium officinale and its closely related species by comparative analysis of complete plastomes. Acta Pharm. Sin. B 2018, 8, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.N.; Mo, Z.Q.; Yang, J.B.; Cai, J.; Ye, L.J.; Zou, J.Y.; Qin, H.T.; Zheng, W.; Hollingsworth, P.M.; Li, D.Z.; et al. Testing genome skimming for species discrimination in the large and taxonomically difficult genus Rhododendron. Mol. Ecol. Resour. 2022, 22, 404–414. [Google Scholar] [CrossRef]
- Li, H.T.; Luo, Y.; Gan, L.; Ma, P.F.; Gao, L.M.; Yang, J.B.; Cai, J.; Gitzendanner, M.A.; Fritsch, P.W.; Zhang, T.; et al. Plastid phylogenomic insights into relationships of all flowering plant families. BMC Biol. 2021, 19, 232. [Google Scholar] [CrossRef]
- Li, H.T.; Yi, T.S.; Gao, L.M.; Ma, P.F.; Zhang, T.; Yang, J.B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef]
- Yu, X.Q.; Yang, D.; Guo, C.; Gao, L.M. Plant phylogenomics based on genome-partitioning strategies: Progress and prospects. Plant Divers. 2018, 40, 158–164. [Google Scholar] [CrossRef]
- Moore, M.J.; Soltis, P.S.; Bell, C.D.; Burleigh, J.G.; Soltis, D.E. Phylogenetic analysis of 83 plastid genes further resolves the early diversification of eudicots. Proc. Natl. Acad. Sci. USA 2010, 107, 4623–4628. [Google Scholar] [CrossRef]
- Ji, Y.H.; Liu, C.K.; Yang, Z.Y.; Yang, L.F.; He, Z.S.; Wang, H.C.; Yang, J.B.; Yi, T.S. Testing and using complete plastomes and ribosomal DNA sequences as the next generation DNA barcodes in Panax (Araliaceae). Mol. Ecol. Resour. 2019, 19, 1333–1345. [Google Scholar] [CrossRef]
- Wang, J.; Fu, C.N.; Mo, Z.Q.; Möller, M.; Yang, J.B.; Zhang, Z.R.; Li, D.Z.; Gao, L.M. Testing the Complete Plastome for Species Discrimination, Cryptic Species Discovery and Phylogenetic Resolution in Cephalotaxus (Cephalotaxaceae). Front. Plant Sci. 2022, 13, 768810. [Google Scholar] [CrossRef]
- Fu, C.N.; Wu, C.S.; Ye, L.J.; Mo, Z.Q.; Liu, J.; Chang, Y.W.; Li, D.-Z.; Chaw, S.M.; Gao, L.M. Prevalence of isomeric plastomes and effectiveness of plastome super-barcodes in yews (Taxus) worldwide. Sci. Rep. 2019, 9, 2773. [Google Scholar] [CrossRef]
- Le, D.-T.; Zhang, Y.Q.; Xu, Y.; Guo, L.X.; Ruan, Z.P.; Burgess, K.S.; Ge, X.J. The utility of DNA barcodes to confirm the identification of palm collections in botanical gardens. PLoS ONE 2020, 15, e0235569. [Google Scholar] [CrossRef]
- Zhou, W.B.; Harris, A.J.; Xiang, Q.Y. Phylogenomics and biogeography of Torreya (Taxaceae)—Integrating data from three organelle genomes, morphology, and fossils and a practical method for reducing missing data from RAD-seq. J. Syst. Evol. 2022, 60, 1241–1262. [Google Scholar] [CrossRef]
- De Luca, A.; Sibilio, G.; De Luca, P.; Del Guacchio, E. DNA barcoding to confirm the morphological identification of the coral trees (Erythrina spp., Fabaceae) in the ancient gardens of Naples (Campania, Italy). Plants 2018, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Davis, C.C.; Donoghue, M.J.; Kelley, S.; Del Tredici, P. Phylogenetic relationships of Torreya (Taxaceae) inferred from sequences of nuclear ribosomal DNA ITS region. Harv. Pap. Bot. 2001, 6, 275–281. [Google Scholar]
- Wen, J. Evolution of eastern Asian and eastern North American disjunct distributions in flowering plants. Annu. Rev. Ecol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- Farjon, A. A Handbook of the World’s Conifers; Brill: Leiden, The Netherlands, 2010; Volume 2, pp. 1032–1041. [Google Scholar]
- Christenhusz, M.J.M.; Reveal, J.L.; Farjon, A.; Gardner, M.F.; Mill, R.R.; Chase, M.W. A new classification and linear sequence of extant gymnosperms. Phytotaxa 2011, 19, 55–70. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Z.H.; Xu, X.T. Taxonomy and Distribution of Global Gymnosperms; Shanghai Scientific and Technical Publishers: Shanghai, China, 2017; pp. 1073–1081. [Google Scholar]
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers: An Analysis of their Distribution, Biogeography, Diversity and Conservation Status; Brill: Leiden, The Netherlands, 2013. [Google Scholar]
- Spector, T.; Determann, R.; Gardner, M. Torreya taxifolia. The IUCN Red List of Threatened Species 2011: E.T30968A9585489. Available online: https://doi.org/10.2305/IUCN.UK.2011-2.RLTS.T30968A9585489.en (accessed on 17 August 2023). [CrossRef]
- Farjon, A. Torreya californica. The IUCN Red List of Threatened Species 2013: E.T34026A2840781. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T34026A2840781.en (accessed on 17 August 2023). [CrossRef]
- Fu, L.K.; Li, N.; Robert, R.M. Taxaceae. In Flora of China; Hong, D.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 1999; pp. 89–96. [Google Scholar]
- Yang, Y.; Luscombe, D. Torreya fargesii. The IUCN Red List of Threatened Species 2013: E.T191877A2011625. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T191877A2011625.en (accessed on 17 August 2023). [CrossRef]
- Yang, Y.; Luscombe, D. Torreya jackii. The IUCN Red List of Threatened Species 2013: E.T32379A2816957. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T32379A2816957.en (accessed on 17 August 2023). [CrossRef]
- Yang, Y.; Luscombe, D. Torreya fargesii var. yunnanensis. The IUCN Red List of Threatened Species 2013: E.T44050A2992243. Available online: https://doi.org/10.2305/IUCN.UK.2013-1.RLTS.T44050A2992243.en (accessed on 17 August 2023). [CrossRef]
- Teng, T.Y.; Pan, C.C.; Liu, J.L.; Chen, Z.L.; Xie, W.Y.; Jin, X.F. Leaf variation of Torreya plants in Zhejiang and its taxonomic implications. J. Hangzhou Norm. Univ. Nat. Sci. Ed. 2017, 16, 9–12, 18. [Google Scholar]
- Kou, Y.X.; Xiao, K.; Lai, X.R.; Wang, Y.J.; Zhang, Z.Y. Natural hybridization between Torreya jackii and T. grandis (Taxaceae) in southeast China. J. Syst. Evol. 2017, 55, 25–33. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, H.J.; Landis, J.B.; Deng, T.; Meng, A.P.; Sun, H.; Peng, Y.S.; Wang, H.C.; Sun, Y.X. Plastome phylogenomic analysis of Torreya (Taxaceae). J. Syst. Evol. 2019, 57, 607–615. [Google Scholar] [CrossRef]
- Ji, Y.H.; Liu, C.K.; Landis, J.B.; Deng, M.; Chen, J.H. Plastome phylogenomics of Cephalotaxus (Cephalotaxaceae) and allied genera. Ann. Bot. 2021, 127, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Xiao, K.; Kou, Y.X. Torreya jackii (Taxaceae): A special species that is genetically admixed, morphologically distinct, and geographically sympatric with parent species. Forests 2019, 10, 174. [Google Scholar] [CrossRef]
- Hao, D.C.; Xiao, P.G.; Huang, B.; Ge, G.B.; Yang, L. Interspecific relationships and origins of Taxaceae and Cephalotaxaceae revealed by partitioned Bayesian analyses of chloroplast and nuclear DNA sequences. Plant Syst. Evol. 2008, 276, 89–104. [Google Scholar] [CrossRef]
- Zhang, L.S.; Mao, X.X.; Qian, X.Y.; Wang, Z.F. The complete chloroplast genome of Torreya parvifolia, a species with extremely small population in China. Mitochondrial DNA Part B Resour. 2021, 6, 387–388. [Google Scholar] [CrossRef]
- Miao, Z.P.; Niu, X.N.; Wang, R.B.; Huang, L.; Ma, B.B.; Li, J.H.; Hong, X. Study of the genus Torreya (Taxaceae) based on chloroplast genomes. Front. Biosci. 2022, 27, 9. [Google Scholar] [CrossRef]
- Guo, C.; Luo, Y.; Gao, L.M.; Yi, T.S.; Li, H.T.; Yang, J.B.; Li, D.Z. Phylogenomics and the flowering plant tree of life. J. Integr. Plant Biol. 2023, 65, 299–323. [Google Scholar] [CrossRef]
- Mogensen, H.L. The hows and whys of cytoplasmic inheritance in seed plants. Am. J. Bot. 1996, 83, 383–404. [Google Scholar] [CrossRef]
- Gianordoli, M. A cytological investigation on gametes and fecundation among Cephalotaxus drupacea. In Fertilization in Higher Plants; Linskens, H.F., Ed.; American Elsevier: New York, NY, USA, 1974; pp. 221–232. [Google Scholar]
- Jaramillo-Correa, J.P.; Bousquet, J. Mitochondrial genome recombination in the zone of contact between two hybridizing conifers. Genetics 2005, 171, 1951–1962. [Google Scholar] [CrossRef]
- Pennell, R.I.; Bell, P.R. Insemination of the archegonium and fertilization in Taxus baccata L. J. Cell Sci. 1988, 89, 551–559. [Google Scholar] [CrossRef]
- Buchholz, J.T. The embryogeny of Torreya, with a note on Austrotaxus. Bull. Torrey Bot. Club 1940, 67, 731–754. [Google Scholar] [CrossRef]
- Lu, Y.F.; Chen, Z.L.; He, A.G.; Liu, J.L.; Wang, P.; Chen, W.J.; Jin, X.F. Torreya dapanshanica (Taxaceae), a new species of gymnosperm from Zhejiang, East China. Phytokeys 2022, 192, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Sarmashghi, S.; Bohmann, K.; Gilbert, M.T.P.; Bafna, V.; Mirarab, S. Skmer: Assembly-free and alignment-free sample identification using genome skims. Genome Biol. 2019, 20, 34. [Google Scholar] [CrossRef] [PubMed]
- Bohmann, K.; Mirarab, S.; Bafna, V.; Gilbert, M.T.P. Beyond DNA barcoding: The unrealized potential of genome skim data in sample identification. Mol. Ecol. 2020, 29, 2521–2534. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Gao, L.; Li, J.; Chen, S.; Su, Y.; Wang, T. The complete chloroplast genome of Torreya fargesii (Taxaceae). Mitochondrial DNA Part A 2016, 27, 3512–3513. [Google Scholar] [CrossRef] [PubMed]
- Miu, Z.P.; Zhang, J.M.; Li, J.H.; Hong, X.; Pan, T. The complete chloroplast genome sequence of an conifer plant Torreya grandis (Pinales, Taxaceae). Mitochondrial DNA Part B Resour. 2018, 3, 1152–1153. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Cheng, H.X.; Liu, L.L.; Wang, X.Y.; Pan, T.; Hong, X. The complete chloroplast genome sequence of an endangered plant Torreya jackii (Pinales, Taxaceae). Conserv. Genet. Resour. 2018, 10, 487–489. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, J.; Zhang, H. Characterization of the complete plastome sequence of Torreya grandis var. jiulongshanensis (Taxaceae), a rare and endangered plant species endemic to Zhejiang province, China. Mitochondrial DNA Part B Resour. 2020, 5, 834–836. [Google Scholar] [CrossRef]
- Shin, S.; Kim, S.-C.; Hong, K.N.; Kang, H.; Lee, J.-W. The complete chloroplast genome of Torreya nucifera (Taxaceae) and phylogenetic analysis. Mitochondrial DNA Part B Resour. 2019, 4, 2537–2538. [Google Scholar] [CrossRef]
- Xu, T.; Kong, L.; Li, Q. Testing efficacy of assembly-free and alignment-free methods for species identification using genome skims, with Patellogastropoda as a test case. Genes 2022, 13, 1192. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, Y.W.; Huang, J.L.; Ya, J.D.; Zhe, M.Q.; Zeng, C.X.; Zhang, Z.R.; Zhang, S.B.; Li, D.Z.; Li, H.T.; et al. DNA barcoding of Cymbidium by genome skimming: Call for next-generation nuclear barcodes. Mol. Ecol. Resour. 2023, 23, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.; Tang, Z.X. Studies on the taxonomy of the genus Torreya Bull. Bot. Res. 1995, 15, 349–362. [Google Scholar]
- Yi, T.P.; Yang, L.; Long, T.L. Torreya parvifolia, a new species of the Taxaceae from Sichuan, China. Bull. Bot. Res. 2006, 26, 513–515. [Google Scholar]
- Liu, H.; Wei, J.; Yang, T.; Mu, W.; Song, B.; Yang, T.; Fu, Y.; Wang, X.; Hu, G.; Li, W.; et al. Molecular digitization of a botanical garden: High-depth whole-genome sequencing of 689 vascular plant species from the Ruili Botanical Garden. GigaScience 2019, 8, giz007. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.B.; Dong, J.; Carlson, M.R.; Yanchuk, A.D. Paternal leakage of mitochondrial DNA in Pinus. Theor. Appl. Genet. 1991, 82, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; dePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Wick, R.R. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Borowiec, M.L. AMAS: A fast tool for alignment manipulation and computing of summary statistics. PeerJ 2016, 4, e1660. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Kainer, D.; Mayer, C.; Stamatakis, A. Selecting optimal partitioning schemes for phylogenomic datasets. BMC Evol. Biol. 2014, 14, 82. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Taxon | Correct Scientific Name | Voucher | Sample ID | GenBank (SRA) Accession Number of Plastome | GenBank (SRA) Accession Number of nrDNA Cistron | Locality Information | Origin | No. of Bases | No. of Reads | Plastome Size (bp) | No. of Reads Mapped to Plastome | Mean Coverage of Plastome (×) | Platome GC Content (%) | Source |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amentotaxus formosana * | - | AM10 | OK138557 (SRR25021671) | (SRR25021671) | China: Taiwan, Pingtung | 3,662,110,500 | 24,414,070 | 136,361 | 450,086 | 500 | 35.8 | Wang et al. [11] | ||
| Amentotaxus yunnanensis | GBOWS079 | AM21 | OR197346 | OR195109 | China: Yunnan, Malipo, Xiajinchang | 1,927,772,700 | 12,851,818 | 137,609 | 134,071 | 146.2 | 35.8 | this study | ||
| Torreya californica | Torreya nucifera | Xianglab209 | To38 | OR197360 | OR195123 | USA: North Carolina JC Arboretum | cultivated | 1,980,844,200 | 13,205,628 | 137,215 | 593,920 | 645.8 | 35.4 | this study |
| Torreya californica | Torreya grandis | - | - | MK249062 | - | UK: Edingburgh, Royal Botanic Garden Edinburgh | cultivated | 136,693 | 35.4 | Zhang et al. [30] | ||||
| Torreya californica * | Xianglab276 | To39 | OR197361 | OR195124 | USA: Washington Arboretum | cultivated | 3,243,790,200 | 21,625,268 | 136,957 | 429,684 | 468.5 | 35.4 | this study | |
| Torreya fargesii | Zhdq-195 | To03 | OR197348 | OR195111 | China: Gansu, Wenxian, Bikou | wild | 4,098,586,800 | 27,323,912 | 137,055 | 261,206 | 279.2 | 35.5 | this study | |
| Torreya fargesii | - | - | NC_029398 | - | China: Hubei, Wuhang, Wuhan Botanic Garden | cultivated | 137,075 | 35.5 | Tao et al. [45] | |||||
| Torreya fargesii * | Zhdq-081 | To02 | OR197347 | OR195110 | China: Sichuan, Maoxian, Fengyi | wild | 2,744,998,800 | 18,299,992 | 137,047 | 206,326 | 223.3 | 35.5 | this study | |
| Torreya grandis | LJ-10759 | To31 | OR197355 | OR195118 | China: Zhejiang, Linan, Tianmushan Nature Reserve | wild | 3,982,402,800 | 26,549,352 | 136,991 | 487,543 | 556.7 | 35.4 | this study | |
| Torreya grandis | 061013-8 | To04 | OR197349 | OR195112 | China: Jiangxi, Qianshan, Wuyishan Natural Reserve | wild | 2,342,818,500 | 15,618,790 | 136,962 | 219,046 | 236.1 | 35.4 | this study | |
| Torreya grandis | - | - | NC_034806 | - | China: Zhejiang, Hangzhou, Hangzhou Botanical Garden | cultivated | 136,949 | 35.4 | Miu et al. [46] | |||||
| Torreya grandis * | ZLN-2011120 | To29 | OR197354 | OR195117 | China: Fujian, Nanping, Wuyishan Natural Reserve | wild | 4,690,482,300 | 31,269,882 | 136,948 | 281,539 | 302.7 | 35.4 | this study | |
| Torreya jackii | GLM-07317 | To07 | OR197350 | OR195113 | China: Yunnan, Kunming, Kunming Botanical Garden | cultivated | 2,766,930,600 | 18,446,204 | 136,798 | 398,596 | 433.6 | 35.5 | this study | |
| Torreya jackii | - | To43 | OK138558 (SRR25021670) | (SRR25021670) | China: Jiangxi, Zixi, Matuoshan | wild | 3,865,107,900 | 25,767,386 | 136,884 | 451,825 | 495 | 35.5 | Wang et al. [11] | |
| Torreya jackii | - | - | NC_056893 | - | China: Yunnan, Kunming, Kunming Botanical Garden | cultivated | 136,924 | 35.5 | Ji et al. [31] | |||||
| Torreya jackii | - | - | MK249064 | - | China: Zhejiang, Hangzhou, Hangzhou Botanical Garden | cultivated | 136,728 | 35.5 | Zhang et al. [30] | |||||
| Torreya jackii | - | - | KX902234 | - | China: Zhejiang, Tonglu, Baiyunyuan Forest Park | wild | 136,720 | 35.5 | Li et al. [47] | |||||
| Torreya jackii * | PVHJX03232 | To42 | OR197362 | OR195125 | China: Jiangxi, Zixi, Matuoshan | wild | 3,073,291,500 | 20,488,610 | 136,751 | 343,308 | 374.3 | 35.5 | this study | |
| Torreya jiulongshanensis | - | - | NC_050372 | - | China: Zhejiang, Jingning, Xikengxia Village | wild | 136,705 | 35.5 | Jiang et al. [48] | |||||
| Torreya jiulongshanensis | - | - | MN244714 (SRR10758782) | (SRR10758782) | China: Zhejiang, Suichang, Jiulong Mountain Nature Reserve | wild | 3,842,990,166 | 25,450,266 | 137,320 | 574,469 | 755.8 | 35.4 | Miao et al. [35] | |
| Torreya nucifera | 19940566 | To35 | OR197357 | OR195120 | USA: Atlanda, Atlanta Botanical Garden | cultivated | 6,329,296,224 | 42,242,036 | 136,944 | 630,323 | 685.3 | 35.5 | this study | |
| Torreya nucifera | 19940647 | To36 | OR197358 | OR195121 | USA: Atlanda, Atlanta Botanical Garden | cultivated | 5,910,584,366 | 39,445,718 | 136,944 | 1,051,344 | 1144.5 | 35.5 | this study | |
| Torreya nucifera | - | - | MK978775 | - | South Korea: Pyungdae-ri, Jeju-do Island | wild | 136,985 | 35.4 | Shin et al. [49] | |||||
| Torreya nucifera | Torreya fargesii | - | - | MK249060 | - | China: Jiangxi, Jiujiang, Lushan Botanical Garden | cultivated | 136,970 | 35.5 | Zhang et al. [30] | ||||
| Torreya nucifera | - | - | MN244713 (SRR10768423) | (SRR10768423) | China: Jiangshu, Nanjing, Nanjing University | cultivated | 3,495,500,208 | 23,149,008 | 136,955 | 238,714 | 261.5 | 35.5 | Miao et al. [35] | |
| Torreya nucifera * | 19980776 | To37 | OR197359 | OR195122 | USA: Atlanda, Atlanta Botanical Garden | cultivated | 5,231,130,270 | 34,919,542 | 137,276 | 247,792 | 269.2 | 35.4 | this study | |
| Torreya parvifolia | 5025 | To44 | OR197363 | OR195126 | China: Sichuan, Tuowu, Wuyi Town | wild | 7,353,802,222 | 49,770,264 | 137,160 | 694,509 | 779.7 | 35.5 | this study | |
| Torreya parvifolia | W24 | To46 | OR197365 | OR195128 | China: Sichuan, Tuowu, Wuyi Town | wild | 7,520,255,400 | 50,135,036 | 137,183 | 587,446 | 639 | 35.5 | this study | |
| Torreya parvifolia | - | - | NC_043866 (SRR13235763) | (SRR13235763) | China: Sichuan, Liangshan | wild | 12,258,497,819 | 81,276,138 | 137,106 | 512,642 | 561.7 | 35.5 | Zhang et al. [34] | |
| Torreya parvifolia | - | - | MN244711 (SRR10769481) | (SRR10769481) | China: Sichuan, Butuo, Wuyi, Wandun Mountain | wild | 4,853,642,830 | 32,143,330 | 136,781 | 264,197 | 290.5 | 35.5 | Miao et al. [35] | |
| Torreya parvifolia * | W21 | To45 | OR197364 | OR195127 | China: Sichuan, Tuowu, Wuyi Town | wild | 7,492,564,200 | 49,950,428 | 137,198 | 342,748 | 374.4 | 35.5 | this study | |
| Torreya taxifolia | - | To33 | NC_063708 (SRR25021669) | (SRR25021669) | USA: Atlanda, Atlanta Botanical Garden | cultivated | 5,860,851,364 | 39,120,638 | 137,117 | 452,291 | 498.8 | 35.4 | Wang et al. [11] | |
| Torreya taxifolia | - | - | MK249063 | - | USA: Atlanda, Atlanta Botanical Garden | cultivated | 137,285 | 35.4 | Zhang et al. [30] | |||||
| Torreya taxifolia * | 20121421 | To34 | OR197356 | OR195119 | USA: Atlanda, Atlanta Botanical Garden | cultivated | 5,707,656,428 | 38,089,352 | 136,972 | 444,007 | 485.5 | 35.4 | this study | |
| Torreya yunnanensis | GLM-07342 | To09 | OR197351 | OR195114 | China: Yunnan, Yulong, Xinzhu | wild | 3,523,821,900 | 23,492,146 | 137,074 | 506,145 | 549.1 | 35.5 | this study | |
| Torreya yunnanensis | GLM-092567-1 | To26 | OR197353 | OR195116 | China: Yunnan, Weixi, Weideng | wild | 4,241,168,700 | 28,274,458 | 137,122 | 399,311 | 430.7 | 35.5 | this study | |
| Torreya yunnanensis | Torreya nucifera | - | - | NC_056892 | - | China: Yunnan, Kunming, Kunming Botanical Garden | cultivated | 137,412 | 35.4 | Ji et al. [31] | ||||
| Torreya yunnanensis | GLM-07342-1 | - | MK249061 | - | China: Yunnan, Yulong | wild | 136,844 | 35.5 | Zhang et al. [30] | |||||
| Torreya yunnanensis | - | - | MN244712 (SRR10758697) | (SRR10758697) | China: Yunnan, Weixi | wild | 4,151,493,132 | 27,493,332 | 136,807 | 233,613 | 256 | 35.5 | Miao et al. [35] | |
| Torreya yunnanensis * | GLM-07469 | To25 | OR197352 | OR195115 | China: Yunnan, Gongshan, Bingzhongluo | wild | 4,142,196,600 | 27,614,644 | 137,029 | 280,765 | 301.4 | 35.5 | this study |
| Dataset | No. of Samples | Length (bp) | Length (bp) * | Variable Sites (%) * | PI Sites (%) * | Identical Sites (%) * |
|---|---|---|---|---|---|---|
| CP | 40 | 145,775 | 140,787 | 2561 (1.82%) | 2063 (1.47%) | 138,226 (98.18%) |
| CP-gb | 40 | 135,852 | 135,852 | 2258 (1.66%) | 1860 (1.37%) | 133,594 (98.34%) |
| MT | 27 | 21,493 | 21,385 | 14 (0.07%) | 7 (0.03%) | 21,371 (99.93%) |
| MT-gb | 27 | 21,303 | 21,303 | 13 (0.06%) | 6 (0.03%) | 21,290 (99.94%) |
| nrDNA | 28 | 6137 | 6115 | 52 (0.85%) | 44 (0.72%) | 6063 (99.15%) |
| nrDNA-gb | 28 | 6100 | 6100 | 52 (0.85%) | 44 (0.72%) | 6048 (99.15%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Z.-Q.; Wang, J.; Möller, M.; Yang, J.-B.; Gao, L.-M. Phylogenetic Relationships and Next-Generation Barcodes in the Genus Torreya Reveal a High Proportion of Misidentified Cultivated Plants. Int. J. Mol. Sci. 2023, 24, 13216. https://doi.org/10.3390/ijms241713216
Mo Z-Q, Wang J, Möller M, Yang J-B, Gao L-M. Phylogenetic Relationships and Next-Generation Barcodes in the Genus Torreya Reveal a High Proportion of Misidentified Cultivated Plants. International Journal of Molecular Sciences. 2023; 24(17):13216. https://doi.org/10.3390/ijms241713216
Chicago/Turabian StyleMo, Zhi-Qiong, Jie Wang, Michael Möller, Jun-Bo Yang, and Lian-Ming Gao. 2023. "Phylogenetic Relationships and Next-Generation Barcodes in the Genus Torreya Reveal a High Proportion of Misidentified Cultivated Plants" International Journal of Molecular Sciences 24, no. 17: 13216. https://doi.org/10.3390/ijms241713216
APA StyleMo, Z. -Q., Wang, J., Möller, M., Yang, J. -B., & Gao, L. -M. (2023). Phylogenetic Relationships and Next-Generation Barcodes in the Genus Torreya Reveal a High Proportion of Misidentified Cultivated Plants. International Journal of Molecular Sciences, 24(17), 13216. https://doi.org/10.3390/ijms241713216