
Marine Bacterioplankton Community Dynamics and Potentially Pathogenic Bacteria in Seawater around Jeju Island, South Korea, via Metabarcoding


 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Seawater Environmental Characteristics
2.2. Metabarcoding Results
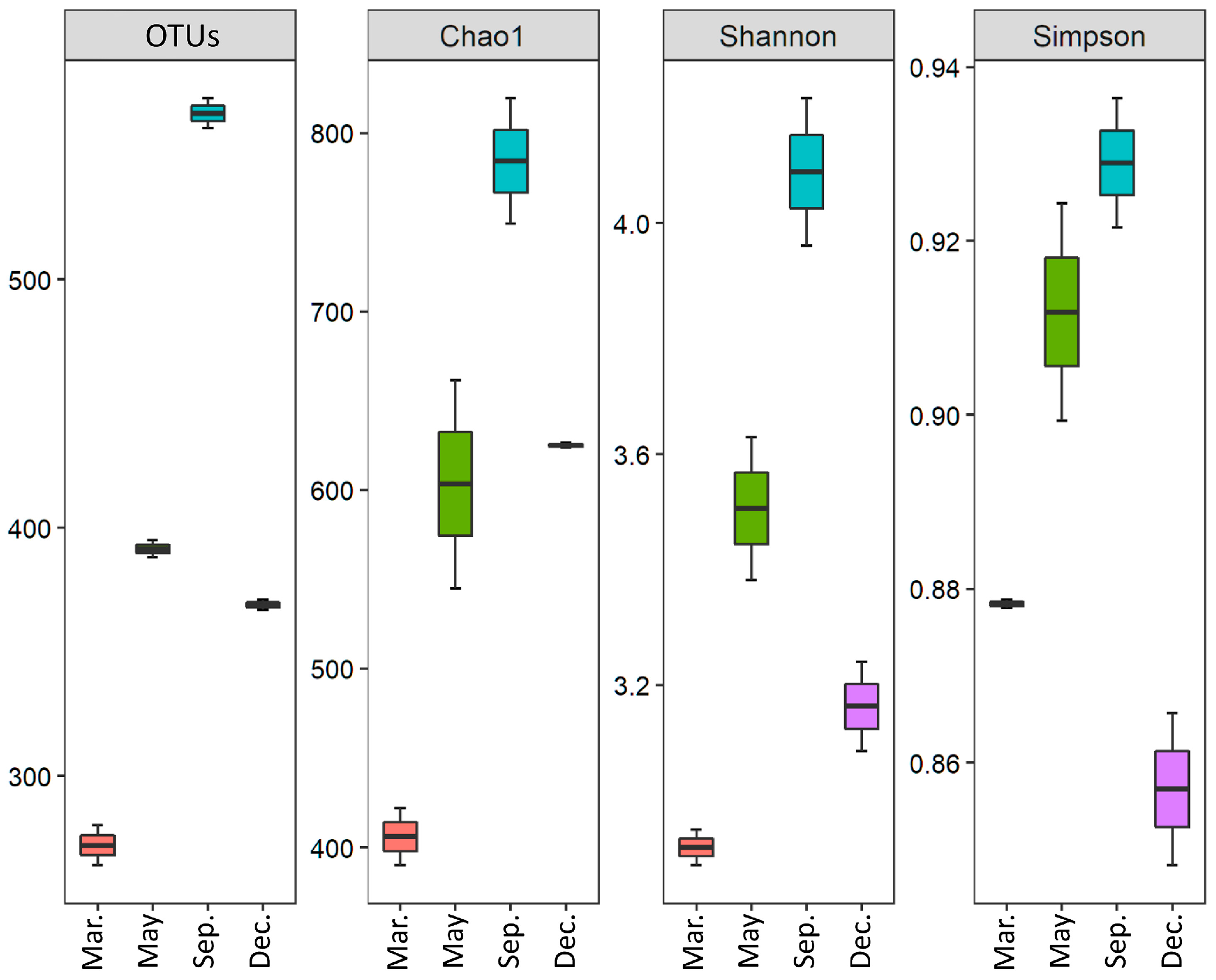
2.2.1. OTU Diversity
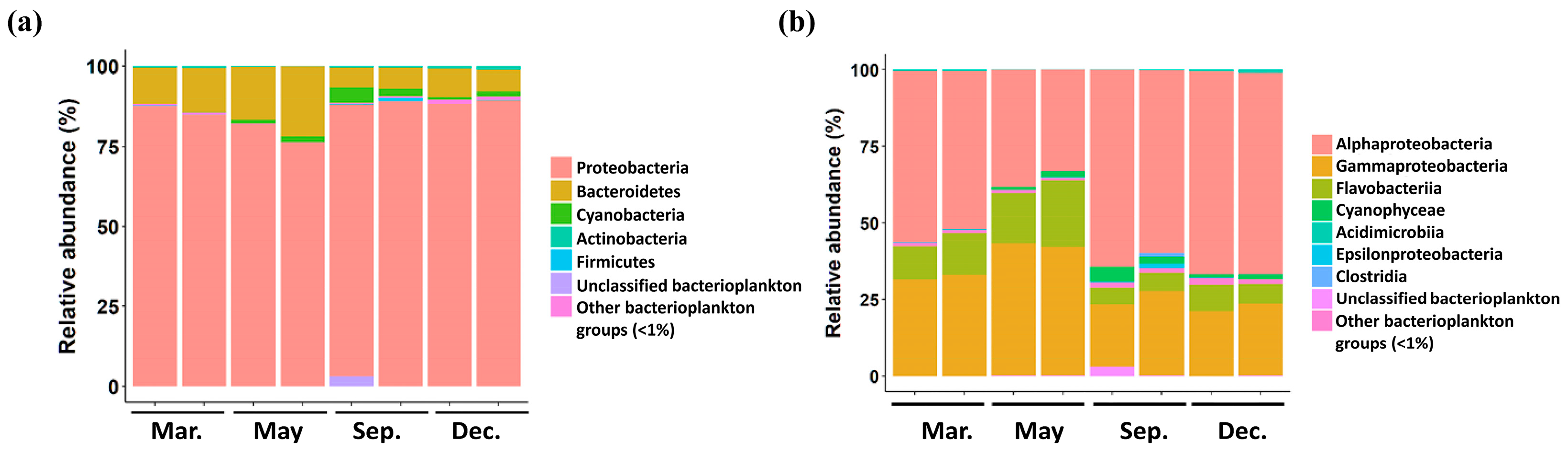
2.2.2. Diversity of Bacterioplankton
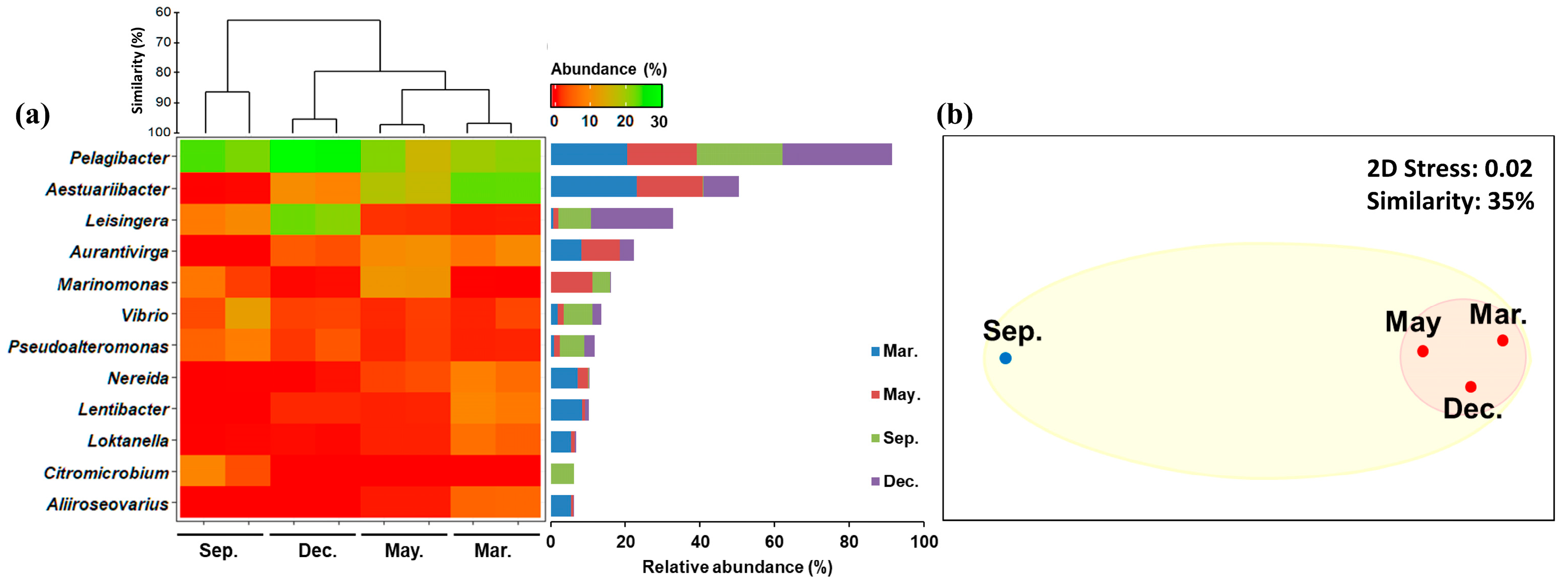
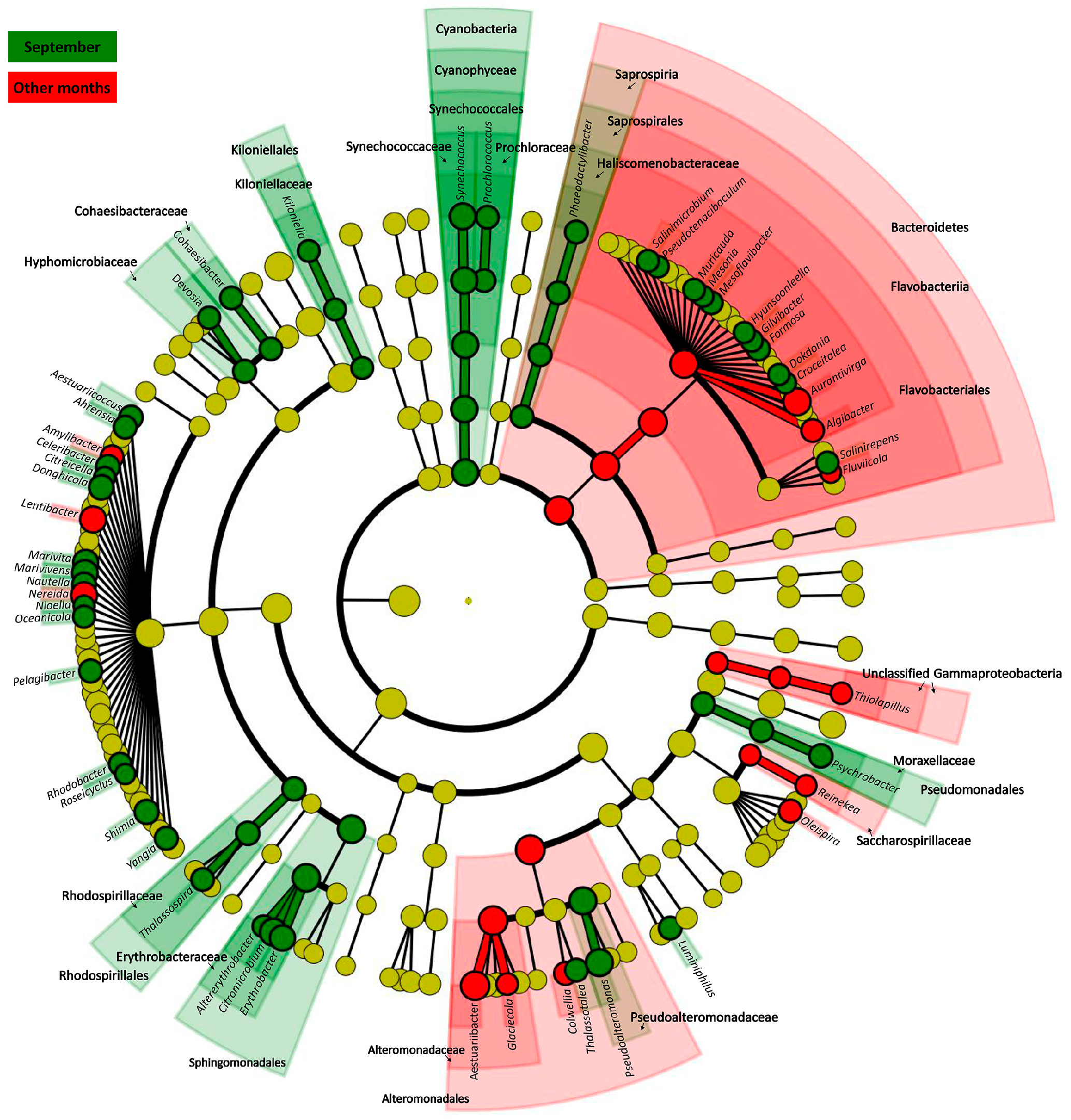
2.2.3. Changes in Common Bacterioplankton
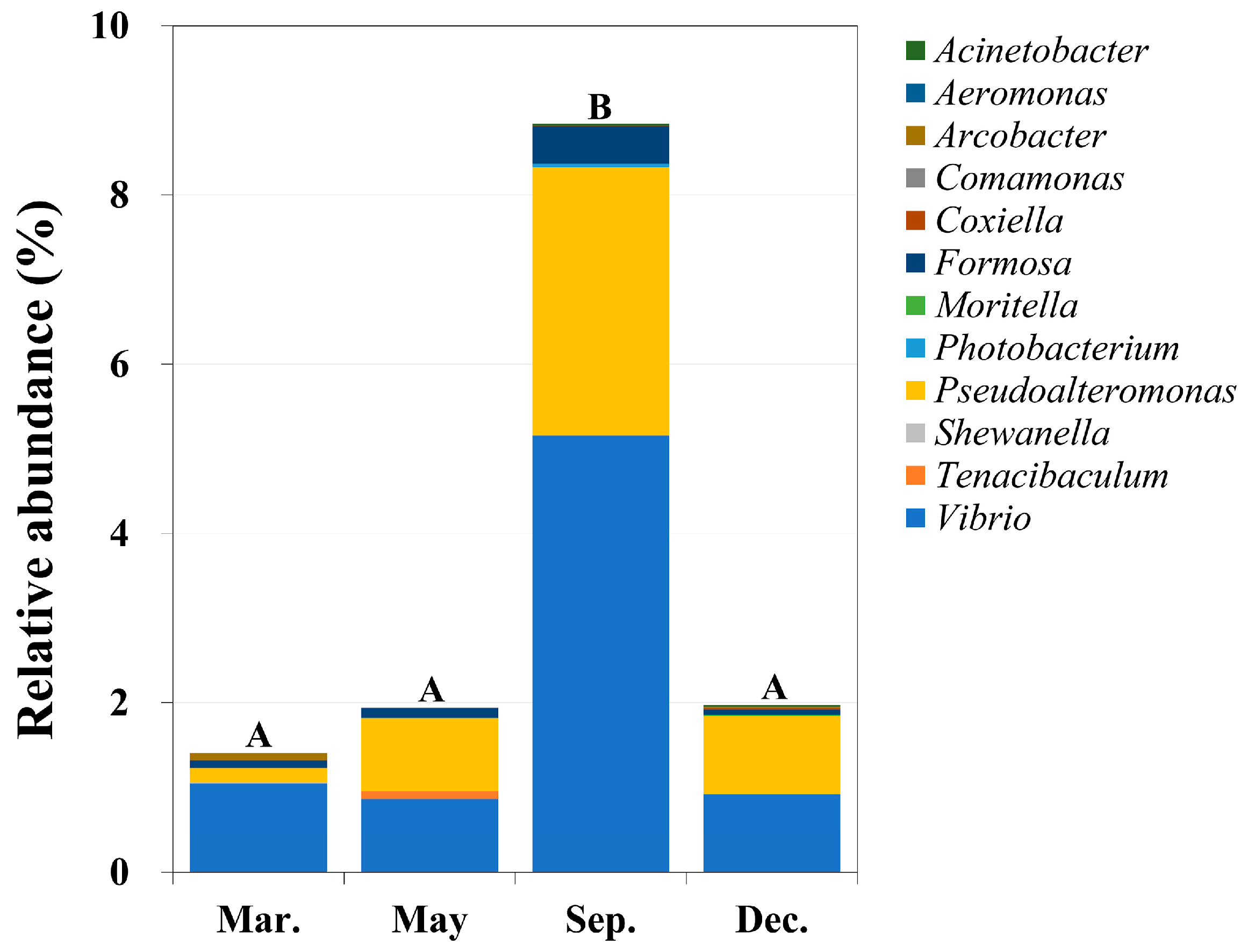
2.3. Identification of Potential Pathogens
3. Materials and Methods
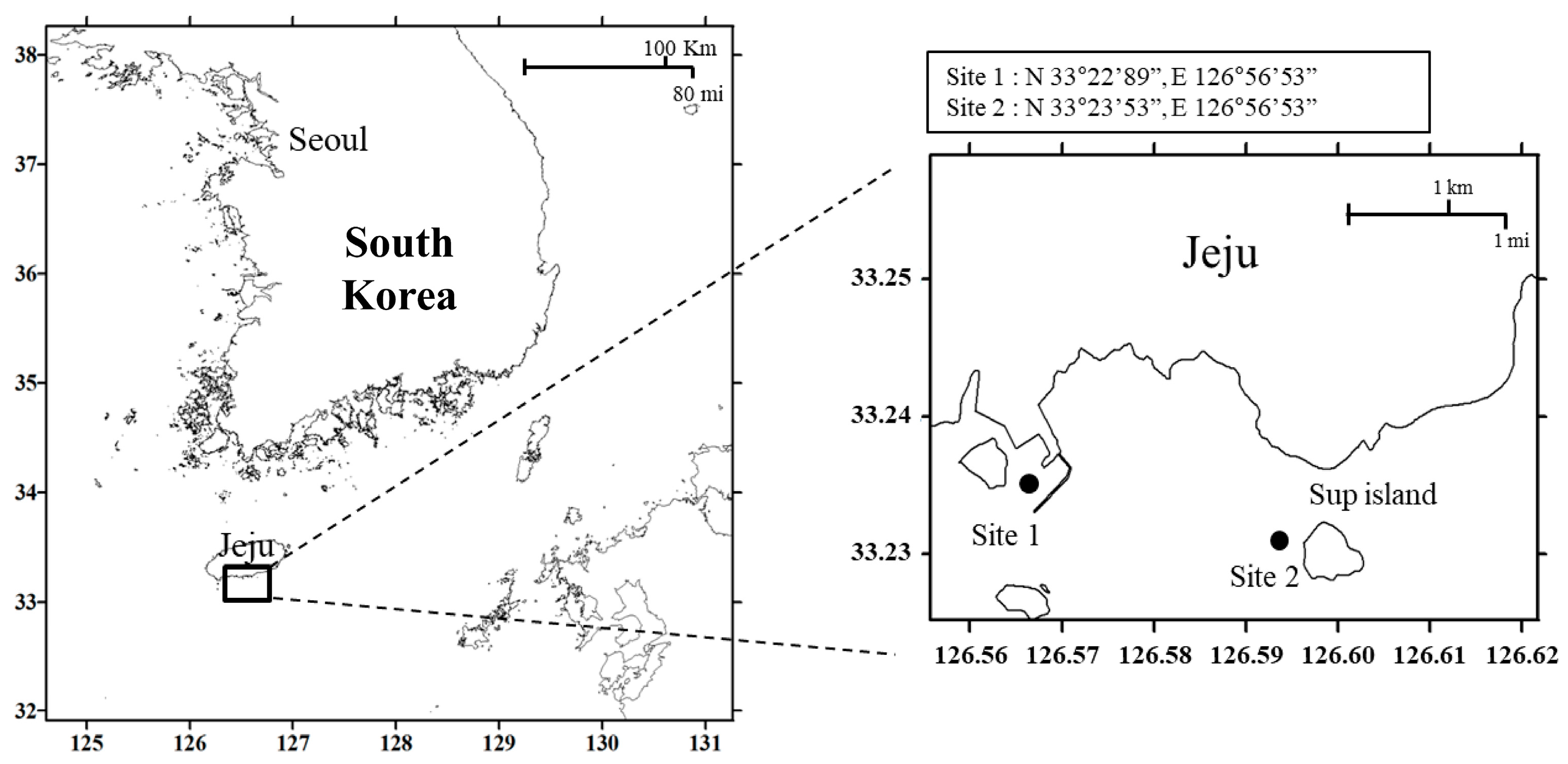
3.1. Sample Collection
3.2. Metabarcoding Analysis
3.3. Bioinformatics Analysis
3.4. Statistical Analysis and Selection of Potentially Pathogenic Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nemergut, D.R.; Costello, E.K.; Hamady, M.; Lozupone, C.; Jiang, L.; Schmidt, S.K.; Fierer, N.; Townsend, A.R.; Cleveland, C.C.; Stanish, L. Global patterns in the biogeography of bacterial taxa. Environ. Microbiol. 2011, 13, 135–144. [Google Scholar] [CrossRef]
- Baek, S.H.; Son, M.; Kim, D.; Choi, H.-W.; Kim, Y.-O. Assessing the ecosystem health status of Korea Gwangyang and Jinhae bays based on a planktonic index of biotic integrity (P-IBI). Ocean Sci. 2014, 49, 291–311. [Google Scholar] [CrossRef]
- Lee, O.O.; Wang, Y.; Yang, J.; Lafi, F.F.; Al-Suwailem, A.; Qian, P.-Y. Pyrosequencing reveals highly diverse and species-specific microbial communities in sponges from the Red Sea. ISME J. 2011, 5, 650–664. [Google Scholar] [CrossRef] [PubMed]
- Chow, C.-E.T.; Sachdeva, R.; Cram, J.A.; Steele, J.A.; Needham, D.M.; Patel, A.; Parada, A.E.; Fuhrman, J.A. Temporal variability and coherence of euphotic zone bacterial communities over a decade in the Southern California Bight. ISME J. 2013, 7, 2259–2273. [Google Scholar] [CrossRef]
- Cram, J.A.; Chow, C.-E.T.; Sachdeva, R.; Needham, D.M.; Parada, A.E.; Steele, J.A.; Fuhrman, J.A. Seasonal and interannual variability of the marine bacterioplankton community throughout the water column over ten years. ISME J. 2015, 9, 563–580. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Steele, J.A.; Caporaso, J.G.; Steinbrück, L.; Reeder, J.; Temperton, B.; Huse, S.; McHardy, A.C.; Knight, R.; Joint, I. Defining seasonal marine microbial community dynamics. ISME J. 2012, 6, 298–308. [Google Scholar] [CrossRef]
- Schauer, M.; Balagué, V.; Pedrós-Alió, C.; Massana, R. Seasonal changes in the taxonomic composition of bacterioplankton in a coastal oligotrophic system. Aquat. Microb. Ecol. 2003, 31, 163–174. [Google Scholar] [CrossRef]
- Pinhassi, J.; Hagström, Å. Seasonal succession in marine bacterioplankton. Aquat. Microb. Ecol. 2000, 21, 245–256. [Google Scholar] [CrossRef]
- Acinas, S.G.; Klepac-Ceraj, V.; Hunt, D.E.; Pharino, C.; Ceraj, I.; Distel, D.L.; Polz, M.F. Fine-scale phylogenetic architecture of a complex bacterial community. Nature 2004, 430, 551–554. [Google Scholar] [CrossRef]
- Kirchman, D.L.; Cottrell, M.T.; Lovejoy, C. The structure of bacterial communities in the western Arctic Ocean as revealed by pyrosequencing of 16S rRNA genes. Environ. Microbiol. 2010, 12, 1132–1143. [Google Scholar] [CrossRef] [PubMed]
- Ladau, J.; Sharpton, T.J.; Finucane, M.M.; Jospin, G.; Kembel, S.W.; O’dwyer, J.; Koeppel, A.F.; Green, J.L.; Pollard, K.S. Global marine bacterial diversity peaks at high latitudes in winter. ISME J. 2013, 7, 1669–1677. [Google Scholar] [CrossRef] [PubMed]
- Aravindraja, C.; Viszwapriya, D.; Karutha Pandian, S. Ultradeep 16S rRNA sequencing analysis of geographically similar but diverse unexplored marine samples reveal varied bacterial community composition. PLoS ONE 2013, 8, e76724. [Google Scholar] [CrossRef]
- Jeong, G.; Kim, H.-J.; Kim, K.E.; Kim, Y.J.; Lee, T.-K.; Shim, W.J.; Jung, S.W. Selective attachment of prokaryotes and emergence of potentially pathogenic prokaryotes on four plastic surfaces: Adhesion study in a natural marine environment. Mar. Pollut. Bull. 2023, 193, 115149. [Google Scholar] [CrossRef]
- Kim, H.-J.; Jeoung, G.; Kim, K.E.; Park, J.S.; Kang, D.; Baek, S.H.; Lee, C.Y.; Kim, H.; Cho, S.; Lee, T.-K. Co-variance between free-living bacteria and Cochlodinium polykrikoides (Dinophyta) harmful algal blooms, South Korea. Harmful Algae 2023, 122, 102371. [Google Scholar] [CrossRef]
- Kim, H.-J.; Park, J.S.; Lee, T.-K.; Kang, D.; Kang, J.-H.; Shin, K.; Jung, S.W. Dynamics of marine bacterial biofouling communities after initial Alteromonas genovensis biofilm attachment to anti-fouling paint substrates. Mar. Pollut. Bull. 2021, 172, 112895. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.W.; Kang, J.; Park, J.S.; Joo, H.M.; Suh, S.-S.; Kang, D.; Lee, T.-K.; Kim, H.-J. Dynamic bacterial community response to Akashiwo sanguinea (Dinophyceae) bloom in indoor marine microcosms. Sci. Rep. 2021, 11, 6983. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Park, J.S.; Jung, S.W.; Kim, H.J.; Joo, H.M.; Kang, D.; Seo, H.; Kim, S.; Jang, M.C.; Lee, K.W. Zooming on dynamics of marine microbial communities in the phycosphere of Akashiwo sanguinea (Dinophyta) blooms. Mol. Ecol. 2021, 30, 207–221. [Google Scholar] [CrossRef]
- Kim, H.J.; Jung, S.W.; Lim, D.-I.; Jang, M.-C.; Lee, T.-K.; Shin, K.; Ki, J.-S. Effects of temperature and nutrients on changes in genetic diversity of bacterioplankton communities in a semi-closed bay, South Korea. Mar. Pollut. Bull. 2016, 106, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-S.; Park, M.; Hwang, J.; Kil, E.-J.; Jung, S.W.; Lee, S.; Lee, T.-K. Seasonal dynamics of marine microbial community in the South Sea of Korea. PLoS ONE 2015, 10, e0131633. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.H.; Jeong, H.J.; Du Yoo, Y. Gambierdiscus jejuensis sp. nov., an epiphytic dinoflagellate from the waters of Jeju Island, Korea, effect of temperature on the growth, and its global distribution. Harmful Algae 2018, 80, 149–157. [Google Scholar] [CrossRef]
- Kim, S.H.; Choi, B.K.; Kim, E. Study on the behavior of the water temperature inversion layer in the northern East China Sea. J. Mar. Sci. Eng. 2020, 8, 157. [Google Scholar] [CrossRef]
- Qian, W.; Kang, H.-S.; Lee, D.-K. Distribution of seasonal rainfall in the East Asian monsoon region. Theor. Appl. Climatol. 2002, 73, 151–168. [Google Scholar] [CrossRef]
- Yu, B.; Xie, G.; Shen, Z.; Shao, K.; Tang, X. Spatiotemporal variations, assembly processes, and co-occurrence patterns of particle-attached and free-living bacteria in a large drinking water reservoir in China. Front. Microbiol. 2023, 13, 1056147. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C.; Zhao, D.; Zeng, J.; Guo, L.; Yu, Z. Disentangling the seasonal co-occurrence patterns and ecological stochasticity of planktonic and benthic bacterial communities within multiple lakes. Sci. Total Environ. 2020, 740, 140010. [Google Scholar] [CrossRef] [PubMed]
- Rooney-Varga, J.N.; Giewat, M.W.; Savin, M.C.; Sood, S.; LeGresley, M.; Martin, J. Links between phytoplankton and bacterial community dynamics in a coastal marine environment. Microb. Ecol. 2005, 49, 163–175. [Google Scholar] [CrossRef]
- Riemann, L.; Winding, A. Community dynamics of free-living and particle-associated bacterial assemblages during a freshwater phytoplankton bloom. Microb. Ecol. 2001, 42, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, W.; Yang, D.; Jiang, J. The current water environment of Tianmuhu Lake and countermeasures for the sustainable development of the ecological tourism. Ecol. Environ. 2003, 12, 405–408. [Google Scholar]
- Rappé, M.S.; Connon, S.A.; Vergin, K.L.; Giovannoni, S.J. Cultivation of the ubiquitous SAR11 marine bacterioplankton clade. Nature 2002, 418, 630–633. [Google Scholar] [CrossRef]
- Lankiewicz, T.S.; Cottrell, M.T.; Kirchman, D.L. Growth rates and rRNA content of four marine bacteria in pure cultures and in the Delaware estuary. ISME J. 2016, 10, 823–832. [Google Scholar] [CrossRef]
- Herlemann, D.P.; Woelk, J.; Labrenz, M.; Jürgens, K. Diversity and abundance of “Pelagibacterales”(SAR11) in the Baltic Sea salinity gradient. Syst. Appl. Microbiol. 2014, 37, 601–604. [Google Scholar] [CrossRef]
- Yi, H.; Bae, K.S.; Chun, J. Aestuariibacter salexigens gen. nov., sp. nov. and Aestuariibacter halophilus sp. nov., isolated from tidal flat sediment, and emended description of Alteromonas macleodii. Int. J. Syst. Evol. Microbiol. 2004, 54, 571–576. [Google Scholar] [CrossRef]
- Yurkov, V.V.; Krieger, S.; Stackebrandt, E.; Beatty, J.T. Citromicrobium bathyomarinum, a novel aerobic bacterium isolated from deep-sea hydrothermal vent plume waters that contains photosynthetic pigment-protein complexes. J. Bacteriol. 1999, 181, 4517–4525. [Google Scholar] [CrossRef]
- Sanchez-Porro, C.; Mellado, E.; Bertoldo, C.; Antranikian, G.; Ventosa, A. Screening and characterization of the protease CP1 produced by the moderately halophilic bacterium Pseudoalteromonas sp. strain CP76. Extremophiles 2003, 7, 221–228. [Google Scholar] [CrossRef]
- Brinkmeyer, R.; Knittel, K.; Jurgens, J.; Weyland, H.; Amann, R.; Helmke, E. Diversity and structure of bacterial communities in Arctic versus Antarctic pack ice. Appl. Environ. Microbiol. 2003, 69, 6610–6619. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-I.; Paillard, C.; Le Chevalier, P.; Choi, K.-S. Report on the occurrence of brown ring disease (BRD) in Manila clam, Ruditapes philippinarum, on the west coast of Korea. Aquaculture 2006, 255, 610–613. [Google Scholar] [CrossRef]
- Osaka, K.; Komatsuzaki, M.; Takahashi, H.; Sakano, S.; Okabe, N. Vibrio vulnificus septicaemia in Japan: An estimated number of infections and physicians’ knowledge of the syndrome. Epidemiol. Infect. 2004, 132, 993–996. [Google Scholar] [CrossRef]
- Aeby, G.; Ross, M.; Williams, G.; Lewis, T.; Work, T. Disease dynamics of Montipora white syndrome within Kaneohe Bay, Oahu, Hawaii: Distribution, seasonality, virulence, and transmissibility. Dis. Aquat. Org. 2010, 91, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kunin, V.; Engelbrektson, A.; Ochman, H.; Hugenholtz, P. Wrinkles in the rare biosphere: Pyrosequencing errors can lead to artificial inflation of diversity estimates. Environ. Microbiol. 2010, 12, 118–123. [Google Scholar] [CrossRef]
- Gontcharova, V.; Youn, E.; Wolcott, R.D.; Hollister, E.B.; Gentry, T.J.; Dowd, S.E. Black box chimera check (B2C2): A windows-based software for batch depletion of chimeras from bacterial 16S rRNA gene datasets. Open Microbiol. J. 2010, 4, 47–52. [Google Scholar] [CrossRef]
- Huse, S.M.; Huber, J.A.; Morrison, H.G.; Sogin, M.L.; Welch, D.M. Accuracy and quality of massively parallel DNA pyrosequencing. Genome Biol. 2007, 8, R143. [Google Scholar] [CrossRef]
- Li, W.; Fu, L.; Niu, B.; Wu, S.; Wooley, J. Ultrafast clustering algorithms for metagenomic sequence analysis. Brief Bioinform. 2012, 13, 656–668. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; et al. “Package ‘Vegan’,” in Community Ecology Package. 2022. Available online: https://github.com/vegandevs/vegan (accessed on 17 June 2023).
- Schwarzer, G. Meta-Analysis in R. In Systematic Reviews in Health Research: Meta-Analysis in Context, 3rd ed.; Matthias, E., Julian, P.T.H., George, D.S., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2022; pp. 510–534. [Google Scholar]
- Wilcoxon, F. Individual comparisons by ranking methods. In Breakthroughs in Statistics: Methodology and Distribution; Kotz, S., Johnson, N.L., Eds.; Springer: New York, NY, USA, 1992; pp. 196–202. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Audic, S.; Claverie, J.-M. The significance of digital gene expression profiles. Genome Res. 1997, 7, 986–995. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Q.; Chen, J.; Wu, H. The occurrence of potential pathogenic bacteria on international ships’ ballast water at Yangshan Port, Shanghai, China. Mar. Pollut. Bull. 2022, 184, 114190. [Google Scholar] [CrossRef]
- Houf, K.; Stephan, R. Isolation and characterization of the emerging foodborn pathogen Arcobacter from human stool. J. Microbiol. Methods 2007, 68, 408–413. [Google Scholar] [CrossRef]
- Johnson, L.G.; Murano, E.A. Arcobacter Isolates from Various Sources. J. Food Prot. 2002, 65, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Levican, A.; Alkeskas, A.; Günter, C.; Forsythe, S.J.; Figueras, M.J. Adherence to and Invasion of Human Intestinal Cells by Arcobacter Species and Their Virulence Genotypes. Appl. Environ. Microbiol. 2013, 79, 4951–4957. [Google Scholar] [CrossRef] [PubMed]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Phylogenetic analysis and antibiotic activity of bacteria isolated from the surface of two co-occurring macroalgae from the Baltic Sea. Eur. J. Phycol. 2013, 48, 47–60. [Google Scholar] [CrossRef]
- Piñeiro-Vidal, M.; Riaza, A.; Santos, Y. Tenacibaculum discolor sp. nov. and Tenacibaculum gallaicum sp. nov., isolated from sole (Solea senegalensis) and turbot (Psetta maxima) culture systems. Int. J. Syst. Evol. Microbiol. 2008, 58, 21–25. [Google Scholar] [CrossRef]
- Fernández-Álvarez, C.; Santos, Y. Phenotypic and Molecular Characterization of Lacinutrix venerupis Isolated from Atlantic Horse Mackerel Trachurus trachurus. J. Aquat. Anim. Health 2019, 31, 320–327. [Google Scholar] [CrossRef]
- Kozińska, A.; Paździor, E.; Pękala, A.; Niemczuk, W. Acinetobacter johnsonii and Acinetobacter lwoffii—The emerging fish pathogens. J. Vet. Res. 2014, 58, 193–199. [Google Scholar] [CrossRef]
- Seifert, H.; Strate, A.; Schulze, A.; Pulverer, G. Vascular Catheter—Related Bloodstream Infection Due to Acinetobacter johnsonii (Formerly Acinetobacter calcoaceticus var. lwoffii): Report of 13 Cases. Clin. Infect. Dis. 1993, 17, 632–636. [Google Scholar] [CrossRef]
- Manan, H.; Rosland, N.A.; Deris, Z.M.; Hashim, N.F.C.; Kasan, N.A.; Ikhwanuddin, M.; Suloma, A.; Fauzan, F. 16S rRNA sequences of Exiguobacterium spp. bacteria dominant in a biofloc pond cultured with whiteleg shrimp, Penaeus vannamei. Aquac. Res. 2022, 53, 2029–2041. [Google Scholar] [CrossRef]
- Jalali, S.; Kohli, S.; Latka, C.; Bhatia, S.; Vellarikal, S.K.; Sivasubbu, S.; Scaria, V.; Ramachandran, S. Screening Currency Notes for Microbial Pathogens and Antibiotic Resistance Genes Using a Shotgun Metagenomic Approach. PLoS ONE 2015, 10, e0128711. [Google Scholar] [CrossRef]
- Kazar, J. Coxiella burnetii infection. Ann. N. Y. Acad. Sci. 2005, 1063, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Løvoll, M.; Wiik-Nielsen, C.; Tunsjø, H.; Colquhoun, D.; Lunder, T.; Sørum, H.; Grove, S. Atlantic salmon bath challenged with Moritella viscosa—Pathogen invasion and host response. Fish Shellfish. Immunol. 2009, 26, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.; Cleary, D.F.R.; Pires, A.C.C.; Rodrigues, A.M.; Quintino, V.; Calado, R.; Gomes, N.C.M. Molecular Analysis of Bacterial Communities and Detection of Potential Pathogens in a Recirculating Aquaculture System for Scophthalmus maximus and Solea senegalensis. PLoS ONE 2013, 8, e80847. [Google Scholar] [CrossRef] [PubMed]
- Urbanczyk, H.; Ogura, Y.; Hendry, T.A.; Gould, A.L.; Kiwaki, N.; Atkinson, J.T.; Hayashi, T.; Dunlap, P.V. Genome Sequence of Photobacterium mandapamensis Strain svers. 1.1, the Bioluminescent Symbiont of the Cardinal Fish Siphamia versicolor. J. Bacteriol. 2011, 193, 3144–3145. [Google Scholar] [CrossRef]
- Fichi, G.; Cardeti, G.; Perrucci, S.; Vanni, A.; Cersini, A.; Lenzi, C.; De Wolf, T.; Fronte, B.; Guarducci, M.; Susini, F. Skin lesion-associated pathogens from Octopus vulgaris: First detection of Photobacterium swingsii, Lactococcus garvieae and betanodavirus. Dis. Aquat. Org. 2015, 115, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Weinberger, F.; Saha, M.; Majzoub, M.E.; Egan, S. Cross-Host Protection of Marine Bacteria Against Macroalgal Disease. Microb. Ecol. 2021, 84, 1288–1293. [Google Scholar] [CrossRef]
- Weiland-Bräuer, N.; Pinnow, N.; Langfeldt, D.; Roik, A.; Güllert, S.; Chibani, C.M.; Reusch, T.B.H.; Schmitz, R.A. The Native Microbiome is Crucial for Offspring Generation and Fitness of Aurelia aurita. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mou, Z.; Yang, H.; Mao, Y.; Yan, Y.; Mo, Z. Isolation and identification the pathogen of Pyropia yezoensis green spot disease. Prog. Fish. Sci. 2019, 40, 140–146. [Google Scholar]
- Wang, Y. Etiology of skin ulcer syndrome in cultured juveniles of Apostichopus japonicus and analysis of reservoir of the pathogens. J. Fish. Sci. China 2006, 13. [Google Scholar] [CrossRef]
- Simidu, U.; Kita-Tsukamoto, K.; Yasumoto, T.; Yotsu, M. Taxonomy of four marine bacterial strains that produce tetrodotoxin. Int. J. Syst. Evol. Microbiol. 1990, 40, 331–336. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Romanenko, L.A.; Matte, M.H.; Matte, G.R.; Lysenko, A.M.; Simidu, U.; Kita-Tsukamoto, K.; Sawabe, T.; Vysotskii, M.V.; Frolova, G.M. Retrieval of the species Alteromonas tetraodonis Simidu et al. 1990 as Pseudoalteromonas tetraodonis comb. nov. and emendation of description. Int. J. Syst. Evol. Microbiol. 2001, 51, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zheng, F.; Sun, X.; Hong, X.; Dong, S.; Wang, B.; Tang, X.; Wang, Y. Identification of the pathogens associated with skin ulceration and peristome tumescence in cultured sea cucumbers Apostichopus japonicus (Selenka). J. Invertebr. Pathol. 2010, 105, 236–242. [Google Scholar] [CrossRef]
- Popović, N.T.; Kazazić, S.; Bilić, B.; Babić, S.; Bojanić, K.; Bujak, M.; Bujak, I.T.; Jadan, M.; Strunjak-Perović, I.; Kepec, S.; et al. Shewanella spp. from wastewater treatment plant-affected environment: Isolation and characterization. Environ. Sci. Pollut. Res. 2022, 29, 82986–83003. [Google Scholar] [CrossRef]
- Labreuche, Y.; Lambert, C.; Soudant, P.; Boulo, V.; Huvet, A.; Nicolas, J.-L. Cellular and molecular hemocyte responses of the Pacific oyster, Crassostrea gigas, following bacterial infection with Vibrio aestuarianus strain 01/32. Microbes Infect. 2006, 8, 2715–2724. [Google Scholar] [CrossRef]
- Costa, J.C.C.P.; Floriano, B.; Villegas, I.M.B.; Rodríguez-Ruiz, J.P.; Posada-Izquierdo, G.D.; Zurera, G.; Pérez-Rodríguez, F. Study of the microbiological quality, prevalence of foodborne pathogens and product shelf-life of Gilthead sea bream (Sparus aurata) and Sea bass (Dicentrarchus labrax) from aquaculture in estuarine ecosystems of Andalusia (Spain). Food Microbiol. 2020, 90, 103498. [Google Scholar] [CrossRef]
- Li, S.-Y.; Huang, Y.-E.; Chen, J.-Y.; Lai, C.-H.; Mao, Y.-C.; Huang, Y.-T.; Liu, P.-Y. Genomics of Ochrobactrum pseudogrignonense (newly named Brucella pseudogrignonensis) reveals a new bla OXA subgroup. Microb. Genom. 2021, 7, 000626. [Google Scholar] [CrossRef]
- Li, S.; Wang, S.; Xie, L.; Liu, Y.; Chen, H.; Feng, J.; Ouyang, L. Identification and Optimization of the Algicidal Activity of a Novel Marine Bacterium Against Akashiwo sanguinea. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Huang, H.; Huang, Z.; Chen, H.; Shao, Z. Isolation and identification of Vibrio campbellii as a bacterial pathogen for luminous vibriosis of Litopenaeus vannamei. Aquac. Res. 2015, 46, 395–404. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.; Sutherland, R.; Thompson, F.; Swings, J. Pathogenicity of vibrios to rainbow trout (Oncorhynchus mykiss, Walbaum) and Artemia nauplii. Environ. Microbiol. 2005, 7, 1488–1495. [Google Scholar] [CrossRef]
- Pérez-Cataluña, A.; Lucena, T.; Tarazona, E.; Arahal, D.R.; Macián, M.C.; Pujalte, M.J. An MLSA approach for the taxonomic update of the Splendidus clade, a lineage containing several fish and shellfish pathogenic Vibrio spp. Syst. Appl. Microbiol. 2016, 39, 361–369. [Google Scholar] [CrossRef]
- Bruto, M.; James, A.; Petton, B.; Labreuche, Y.; Chenivesse, S.; Alunno-Bruscia, M.; Polz, M.F.; Le Roux, F. Vibrio crassostreae, a benign oyster colonizer turned into a pathogen after plasmid acquisition. ISME J. 2016, 11, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, X.; Wu, C.; Zhang, Y.; Fan, K.; Zhang, X.; Wei, Y. Isolation, identification and pathogenesis study of Vibrio diabolicus. Aquaculture 2020, 533, 736043. [Google Scholar] [CrossRef]
- Ramamurthy, T.; Chowdhury, G.; Pazhani, G.P.; Shinoda, S. Vibrio fluvialis: An emerging human pathogen. Front. Microbiol. 2014, 5, 91. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Qin, G.; Luo, W.; Lin, Q. A novel pathogenic bacteria (Vibrio fortis) causing enteritis in cultured seahorses, Hippocampus erectus Perry, 1810. J. Fish Dis. 2016, 39, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Roux, F.L.; Goubet, A.; Thompson, F.; Faury, N.; Gay, M.; Swings, J.; Saulnier, D. Vibrio gigantis sp. nov., isolated from the haemolymph of cultured oysters (Crassostrea gigas). Int. J. Syst. Evol. Microbiol. 2005, 55, 2251–2255. [Google Scholar] [CrossRef] [PubMed]
- Sicuro, B.; Luzzana, U. The state of Seriola spp. other than yellowtail (S. quinqueradiata) farming in the world. Rev. Fish. Sci. Aquac. 2016, 24, 314–325. [Google Scholar] [CrossRef]
- Harrison, J.; Nelson, K.; Morcrette, H.; Morcrette, C.; Preston, J.; Helmer, L.; Titball, R.W.; Butler, C.S.; Wagley, S. The increased prevalence of Vibrio species and the first reporting of Vibrio jasicida and Vibrio rotiferianus at UK shellfish sites. Water Res. 2022, 211, 117942. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, X.; Wang, C.; Bai, C.; Li, C.; Li, C.; Xin, L. Isolation and characterization of Vibrio kanaloae as a major pathogen associated with mass mortalities of Ark Clam, Scapharca broughtonii, in cold season. Microorganisms 2021, 9, 2161. [Google Scholar] [CrossRef] [PubMed]
- Schaeck, M.; Duchateau, L.; Broeck, W.V.D.; Van Trappen, S.; De Vos, P.; Coulombet, C.; Boon, N.; Haesebrouck, F.; Decostere, A. Vibrio lentus protects gnotobiotic sea bass (Dicentrarchus labrax L.) larvae against challenge with Vibrio harveyi. Vet. Microbiol. 2016, 185, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Farto, R.; Armada, S.; Montes, M.; Guisande, J.; Pérez, M.; Nieto, T. Vibrio lentus associated with diseased wild octopus (Octopus vulgaris). J. Invertebr. Pathol. 2003, 83, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Andree, K.B.; Carrasco, N.; Carella, F.; Furones, D.; Prado, P. Vibrio mediterranei, a potential emerging pathogen of marine fauna: Investigation of pathogenicity using a bacterial challenge in Pinna nobilis and development of a species-specific PCR. J. Appl. Microbiol. 2021, 130, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Reina, J.C.; Fuentes-Monteverde, J.C.; Fernández, G.; Rodríguez, J.; Jiménez, C.; Llamas, I. AHL-lactonase expression in three marine emerging pathogenic Vibrio spp. reduces virulence and mortality in brine shrimp (Artemia salina) and Manila clam (Venerupis philippinarum). PLoS ONE 2018, 13, e0195176. [Google Scholar] [CrossRef]
- Mondal, S.K.; Lijon, B.; Reza, R.; Ishika, T. Isolation and identification of Vibrio nereis and Vibrio harveyi in farm raised Penaeus monodon marine shrimp. Int. J. Biosci. 2016, 8, 55–61. [Google Scholar]
- Ruiz, P.; Balado, M.; Fuentes-Monteverde, J.C.; Toranzo, A.E.; Rodríguez, J.; Jiménez, C.; Avendaño-Herrera, R.; Lemos, M.L. The Fish Pathogen Vibrio ordalii Under Iron Deprivation Produces the Siderophore Piscibactin. Microorganisms 2019, 7, 313. [Google Scholar] [CrossRef]
- Amin, A.M.R.; Feng, G.; Al-Saari, N.; Meirelles, P.M.; Yamazaki, Y.; Mino, S.; Thompson, F.L.; Sawabe, T.; Sawabe, T. The First Temporal and Spatial Assessment of Vibrio Diversity of the Surrounding Seawater of Coral Reefs in Ishigaki, Japan. Front. Microbiol. 2016, 7, 1185. [Google Scholar] [CrossRef] [PubMed]
- Villamil, L.; Figueras, A.; Aranguren, R.; Novoa, B. Non-specific immune response of turbot, Scophthalmus maximus (L.), experimentally infected with a pathogenic Vibrio pelagius. J. Fish Dis. 2003, 26, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Guisande, J.A.; Lago, E.P.; Prado, S.; Nieto, T.P.; Seguín, R.F. Genotypic Diversity of Culturable Vibrio Species Associated with the Culture of Oysters and Clams in Galicia and Screening of Their Pathogenic Potential. J. Shellfish. Res. 2008, 27, 801–809. [Google Scholar] [CrossRef]
- Verschuere, L.; Heang, H.; Criel, G.; Sorgeloos, P.; Verstraete, W. Selected bacterial strains protect Artemia spp. from the pathogenic effects of Vibrio proteolyticus CW8T2. Appl. Environ. Microbiol. 2000, 66, 1139–1146. [Google Scholar] [CrossRef]
- Ray, A.; Kinch, L.N.; Santos, M.d.S.; Grishin, N.V.; Orth, K.; Salomon, D. Proteomics Analysis Reveals Previously Uncharacterized Virulence Factors in Vibrio proteolyticus. mBio 2016, 7. [Google Scholar] [CrossRef]
- Gökbulak, F. Effect of American bison (Bison bison L.) on the recovery and germinability of seeds of range forage species. Grass Forage Sci. 2002, 57, 395–400. [Google Scholar] [CrossRef]
- Qiao, G.; Jang, I.-K.; Won, K.M.; Woo, S.H.; Xu, D.-H.; Park, S.I. Pathogenicity comparison of high- and low-virulence strains of Vibrio scophthalmi in olive flounder Paralichthys olivaceus. Fish. Sci. 2012, 79, 99–109. [Google Scholar] [CrossRef]
- Duperthuy, M.; Schmitt, P.; Garzón, E.; Caro, A.; Rosa, R.D.; Le Roux, F.; Lautrédou-Audouy, N.; Got, P.; Romestand, B.; de Lorgeril, J.; et al. Use of OmpU porins for attachment and invasion of Crassostrea gigas immune cells by the oyster pathogen Vibrio splendidus. Proc. Natl. Acad. Sci. USA 2011, 108, 2993–2998. [Google Scholar] [CrossRef]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.; Gibson, L. Two pathogens of Greenshell™ mussel larvae, Perna canaliculus: Vibrio splendidus and a V. coralliilyticus/neptunius-like isolate. J. Fish Dis. 2009, 32, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Robino, E.; Poirier, A.C.; Amraoui, H.; Le Bissonnais, S.; Perret, A.; Lopez-Joven, C.; Auguet, J.C.; Rubio, T.P.; Cazevieille, C.; Rolland, J.L. Resistance of the oyster pathogen Vibrio tasmaniensis LGP32 against grazing by Vannella sp. marine amoeba involves Vsm and CopA virulence factors. Environ. Microbiol. 2020, 22, 4183–4197. [Google Scholar] [CrossRef]
- Ho, H.T.; Lipman, L.J.; Gaastra, W. Arcobacter, what is known and unknown about a potential foodborne zoonotic agent! Vet. Microbiol. 2006, 115, 1–13. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Month | Trimmed Reads | OTUs | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|
| March | 6673 | 272 | 406.1 | 2.92 | 0.88 |
| May | 9782 | 393 | 603.4 | 3.51 | 0.91 |
| September | 9813 | 567 | 784.2 | 4.09 | 0.93 |
| December | 5745 | 369 | 625.1 | 3.16 | 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Kim, K.E.; Kim, Y.J.; Kang, H.; Shin, J.W.; Kim, S.; Lee, S.H.; Jung, S.W.; Lee, T.-K. Marine Bacterioplankton Community Dynamics and Potentially Pathogenic Bacteria in Seawater around Jeju Island, South Korea, via Metabarcoding. Int. J. Mol. Sci. 2023, 24, 13561. https://doi.org/10.3390/ijms241713561
Kim H-J, Kim KE, Kim YJ, Kang H, Shin JW, Kim S, Lee SH, Jung SW, Lee T-K. Marine Bacterioplankton Community Dynamics and Potentially Pathogenic Bacteria in Seawater around Jeju Island, South Korea, via Metabarcoding. International Journal of Molecular Sciences. 2023; 24(17):13561. https://doi.org/10.3390/ijms241713561
Chicago/Turabian StyleKim, Hyun-Jung, Kang Eun Kim, Yu Jin Kim, Hangoo Kang, Ji Woo Shin, Soohyun Kim, Sang Heon Lee, Seung Won Jung, and Taek-Kyun Lee. 2023. "Marine Bacterioplankton Community Dynamics and Potentially Pathogenic Bacteria in Seawater around Jeju Island, South Korea, via Metabarcoding" International Journal of Molecular Sciences 24, no. 17: 13561. https://doi.org/10.3390/ijms241713561
APA StyleKim, H. -J., Kim, K. E., Kim, Y. J., Kang, H., Shin, J. W., Kim, S., Lee, S. H., Jung, S. W., & Lee, T. -K. (2023). Marine Bacterioplankton Community Dynamics and Potentially Pathogenic Bacteria in Seawater around Jeju Island, South Korea, via Metabarcoding. International Journal of Molecular Sciences, 24(17), 13561. https://doi.org/10.3390/ijms241713561