
Evaluation of Anticancer and Anti-Inflammatory Activities of Some Synthetic Rearranged Abietanes

 , ,
, ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
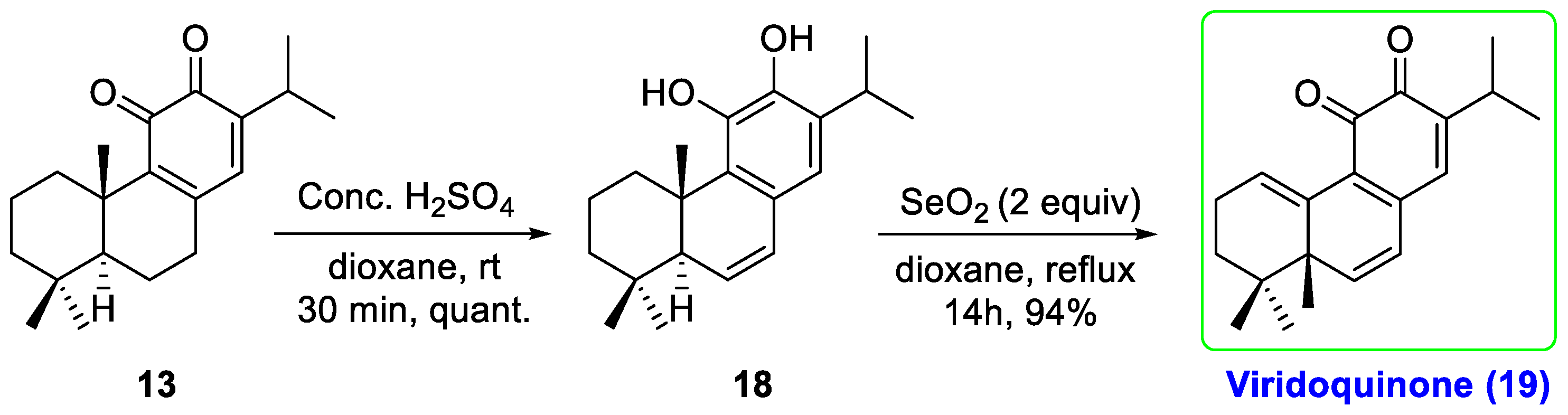
2.1. Chemistry
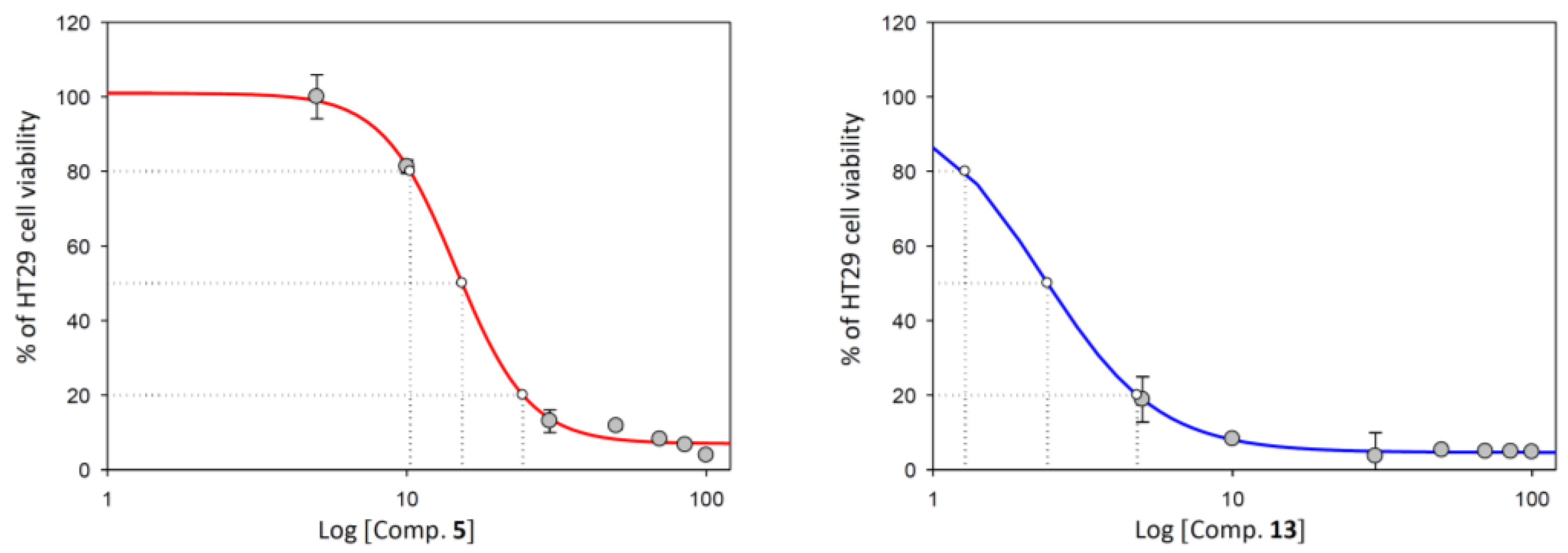
2.2. Anticancer Activity
2.2.1. Cell Viability Assay
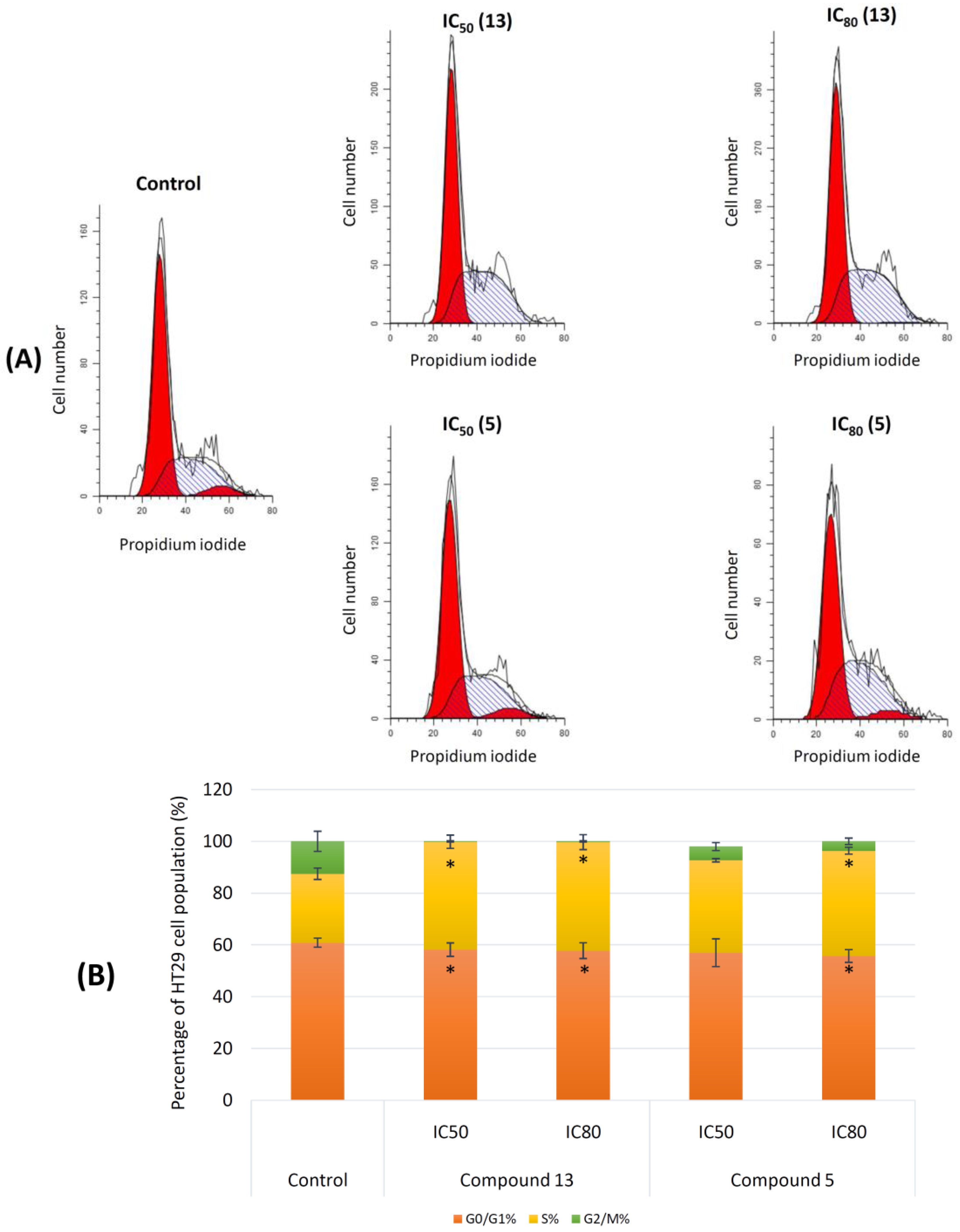
2.2.2. Cell Cycle Arrest and Distribution
2.2.3. Induction of Apoptosis in Tumor Cell Lines
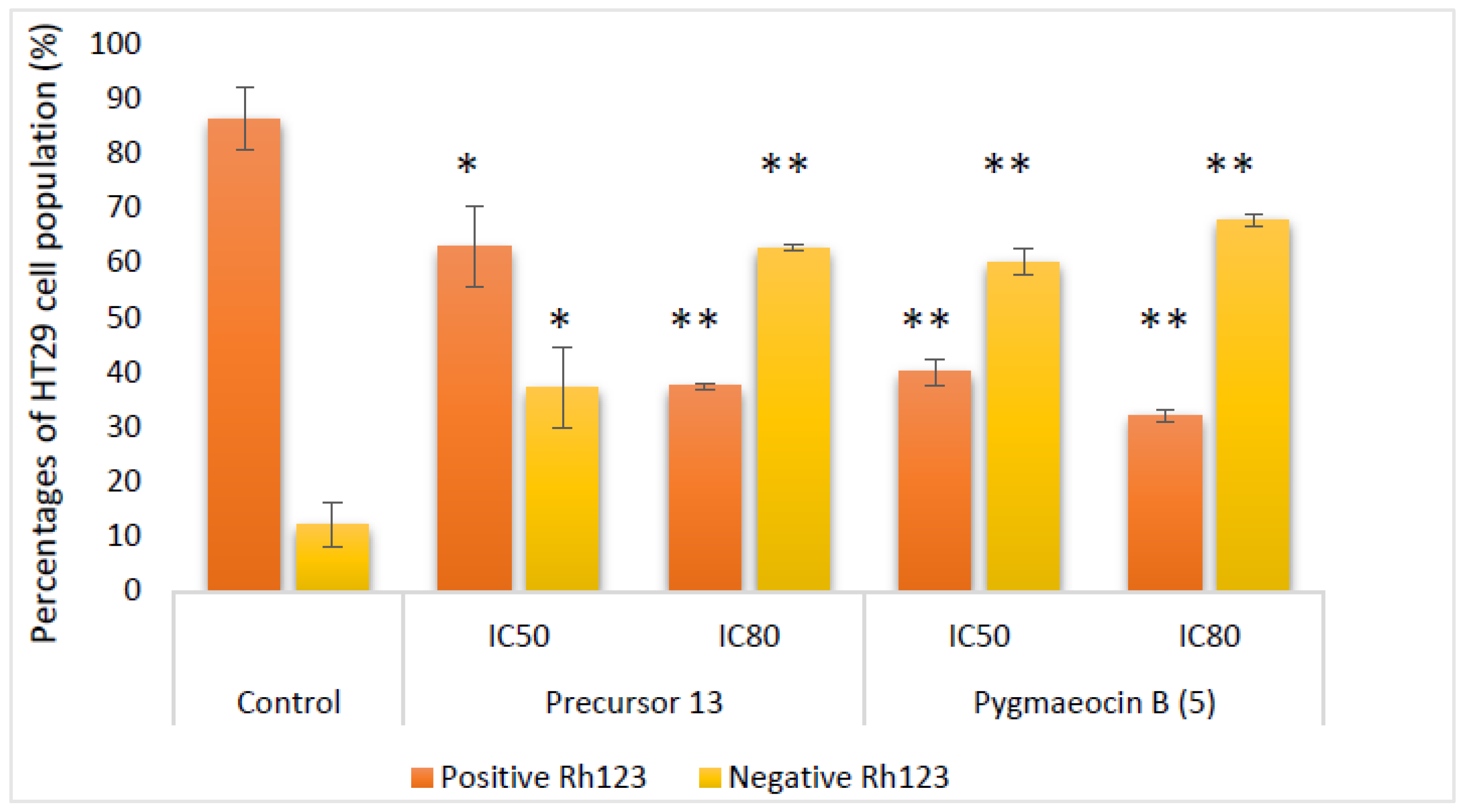
2.2.4. Effects on Mitochondrial Membrane Potential (MMP)
2.2.5. Morphological Changes during Apoptosis Hoechst Staining
2.3. Anti-Inflammatory Activity
2.3.1. RAW 264.7 Cell Viability
2.3.2. Inhibition of Nitric Oxide (NO) Production
3. Discussion
4. Materials and Methods
4.1. Chemistry
Experimental Procedures
4.2. Biological Experimental Procedures
4.2.1. Materials
4.2.2. Test Compounds
4.2.3. Cell Culture and Viability Assay
4.2.4. Measurement of Nitric Oxide Concentration
4.2.5. HT29 Cell Cycle Analysis
4.2.6. Annexin V-FITC/Propidium Iodide Flow Cytometry Analysis
4.2.7. Flow Cytometry Analysis of the Mitochondrial Membrane Potential
4.2.8. Hoechst-Stained Fluorescence Microscopy
4.2.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Song, M.; Giovannucci, E. Preventable incidence and mortality of carcinoma associated with lifestyle factors among white adults in the United States. JAMA Oncol. 2016, 2, 1154–1161. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.-J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect. 2021, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the 30 Years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Le, T.Q.; Oh, C.H. Recent Advances in Abietane/Icetexane Synthesis. Tetrahedron 2022, 108, 154133. [Google Scholar] [CrossRef]
- Nagata, H.; Miyazawa, N.; Ogasawara, K. Tandem Single-Step Construction of Chiral Hexahydrophenanthrenes: A Concise Route to (+)-Ferruginol. Org. Lett. 2001, 3, 1737–1740. [Google Scholar] [CrossRef]
- Lv, C.; Zeng, H.-W.; Wang, J.-X.; Yuan, X.; Zhang, C.; Fang, T.; Yang, P.-M.; Wu, T.; Zhou, Y.-D.; Nagle, D.G.; et al. The Antitumor Natural Product Tanshinone IIA Inhibits Protein Kinase C and Acts Synergistically with 17-AAG. Cell Death Dis. 2018, 9, 165. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Zappavigna, S.; Cossu, A.M.; Grimaldi, A.; Bocchetti, M.; Ferraro, G.A.; Nicoletti, G.F.; Filosa, R.; Caraglia, M. Anti-inflammatory drugs as anticancer agents. Int. J. Mol. Sci. 2020, 21, 2605. [Google Scholar] [CrossRef]
- Galisteo, A.; Jannus, F.; García-García, A.; Aheget, H.; Rojas, S.; Lupiañez, J.A.; Rodríguez-Diéguez, A.; Reyes-Zurita, F.J.; Quílez del Moral, J.F. Diclofenac N-derivatives as therapeutic agents with anti-inflammatory and anti-cancer effect. Inter. J. Mol. Sci. 2021, 22, 5067. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-H.; Fang, J.-M.; Cheng, Y.-S. Diterpenes and Related Cycloadducts from Taiwania cryptomerioides. Phytochemistry 1996, 42, 1657–1663. [Google Scholar] [CrossRef]
- Majetich, G.; Zou, G. Total Synthesis of (−)-Barbatusol, (+)-Demethylsalvicanol, (−)-Brussonol, and (+)-Grandione. Org. Lett. 2008, 10, 81–83. [Google Scholar] [CrossRef]
- Chen, W.; Meng, Q.; Piantini, U.; Hesse, M. Two Novel Diterpenoids from Pygmaeopremna herbacea. J. Nat. Prod. 1989, 52, 581–587. [Google Scholar] [CrossRef]
- Monache, F.D.; Monache, G.D.; Gacs-Baitz, E.; De Coelho, J.S.; De Albuquerque, I.L.; De Chiappeta, A.; De Mello, J.F. Um-brosone, An Ortho-Quinone from Hyptis umbrosa. Phytochemistry 1990, 29, 3971–3972. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Blaskó, G.; Cordell, G.A. Diterpenes of Salvia prionitis. Phytochemistry 1989, 28, 177–181. [Google Scholar] [CrossRef]
- Ulubelen, A.; Topçu, G.; Chai, H.-B.; Pezzuto, J.M. Cytotoxic Activity of Diterpenoids Isolated from Salvia hypargeia. Pharm. Biol. 1999, 37, 148–151. [Google Scholar] [CrossRef]
- Naman, C.B.; Gromovsky, A.D.; Vela, C.M.; Fletcher, J.N.; Gupta, G.; Varikuti, S.; Zhu, X.; Zywot, M.; Chai, H.; Werbovetz, K.A.; et al. Antileishmanial and Cytotoxic Activity of Some Highly Oxidized Abietane Diterpenoids from the Bald Cypress, Taxodium distichum. J. Nat. Prod. 2016, 79, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Luo, D.; Wang, T.; Ji, X.; Xu, G. Vasorelaxant 4,5-Seco-Abietane Diterpenoids with Diverse 6/6/6, 6/6/7, and 6/6/8 Architectures from Salvia prattii Hemsl. Fitoterapia 2020, 142, 104521. [Google Scholar] [CrossRef]
- Zadali, R.; Nejad-Ebrahimi, S.; Hadjiakhoondi, A.; Fiengo, L.; D’Ambola, M.; De Vita, S.; Tofighi, Z.; Chini, M.G.; Bifulco, G.; De Tommasi, N. Diterpenoids from Zhumeria majdae Roots as Potential Heat Shock Protein 90 (HSP90) Modulators. Phytochemistry 2021, 185, 112685. [Google Scholar] [CrossRef]
- Zhao, S.-M.; Chou, G.-X.; Yang, Q.-S.; Wang, W.; Zhou, J.-L. Abietane Diterpenoids from Caryopteris incana (Thunb.) Miq. Org. Biomol. Chem. 2016, 14, 3510–3511. [Google Scholar] [CrossRef]
- Ait El Had, M.; Guardia, J.J.; Ramos, J.M.; Taourirte, M.; Chahboun, R.; Alvarez-Manzaneda, E. Bioinspired Synthesis of Pygmaeocins and Related Rearranged Abietane Diterpenes: Synthesis of Viridoquinone. Org. Lett. 2018, 20, 5666–5670. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Tanaka, Y.; Terao, H.; Takeda, Y.; Wada, M. The Synthesis of Salvinolone, Saprorthoquinone, and 4-Hydroxysapriparaquinone from (+)-Dehydroabietic Acid. Bull. Chem. Soc. Jpn. 1993, 66, 3053–3057. [Google Scholar] [CrossRef]
- Jannus, F.; Medina-O’Donnell, M.; Rivas, F.; Díaz-Ruiz, L.; Rufino-Palomares, E.E.; Lupiáñez, J.A.; Parra, A.; Reyes-Zurita, F.J. A diamine-PEGylated oleanolic acid derivative induced efficient apoptosis through a death receptor and mitochondrial apoptotic pathway in HepG2 human hepatoma cells. Biomolecules 2020, 10, 1375. [Google Scholar] [CrossRef] [PubMed]
- González, M.A. Aromatic abietane diterpenoids: Their biological activity and synthesis. Nat. Prod. Rep. 2015, 32, 684–704. [Google Scholar] [CrossRef]
- Kuroda, Y.; Nicacio, K.J.; da Silva, I.A., Jr.; Leger, P.R.; Chang, S.; Gubiani, J.R.; Deflon, V.M.; Nagashima, N.; Rode, A.; Blackford, K.; et al. Isolation, synthesis and bioactivity studies of phomactin terpenoids. Nat. Chem. 2018, 10, 938–945. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Emami, S.A.; Tayarani-Najaran, Z.; Iranshahi, M.; Shakeri, A.; Hohmann, J.; Asili, J. Cytotoxic diterpene quinones from Salvia tebesana Bunge. Fitoterapia 2018, 128, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Xu, J.; Li, M.; Zhao, M.; Ding, J.; Zhang, J.S. Novel cytotoxic seco-abietane rearranged diterpenoids from Salvia prionitis. Planta Med. 2005, 71, 861–866. [Google Scholar] [CrossRef]
- Zheng, X.; Kadir, A.; Zheng, G.; Jin, P.; Qin, D.; Maiwulanjiang, M.; Aisa, H.A.; Yao, G. Antiproliferative abietane quinone diterpenoids from the roots of Salvia deserta. Bioorg. Chem. 2020, 104, 104261. [Google Scholar] [CrossRef]
- Chen, X.; Ding, J.; Ye, Y.M.; Zhang, J.S. Bioactive Abietane and s eco-Abietane Diterpenoids from Salvia prionitis. J. Nat. Prod. 2002, 65, 1016–1020. [Google Scholar] [CrossRef]
- Nazemosadat-Arsanjani, Z.; Moein, M.; Yousuf, S.; Firuzi, O.; Choudhary, M.I. Reassessing the molecular structures of some previously isolated abietane diterpenoids with a naphthalene moiety and the structure-activity relationship (SAR) of quinone diterpenoids. Phytochemistry 2022, 204, 113433. [Google Scholar] [CrossRef]
- Hernandez-Perez, M.; Rabanal, R.M.; Arias, A.; de La Torre, M.C.; Rodriguez, B. Aethiopinone, an antibacterial and cytotoxic agent from Salvia aethiopis roots. Pharm. Biol. 1999, 37, 17–21. [Google Scholar] [CrossRef]
- Różalski, M.; Kuźma, Ł.; Wysokińska, H.; Krajewska, U. Cytotoxic and proapoptotic activity of diterpenoids from in vitro cultivated Salvia sclarea roots. Studies on the leukemia cell lines. Z. Naturforsch. C J. Biosci. 2006, 61, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, H.; Niu, T.; Tang, M.L.; Chang, J. Discovery of novel indoleamine 2, 3-dioxygenase 1 (IDO1) and histone deacetylase 1 (HDAC1) dual inhibitors derived from the natural product saprorthoquinone. Molecules 2020, 25, 4494. [Google Scholar] [CrossRef]
- Netto, C.D.; Santos, E.S.; Castro, C.P.; da Silva, A.J.; Rumjanek, V.M.; Costa, P.R. (±)-3, 4-Dihydroxy-8, 9-methylenedioxypterocarpan and derivatives: Cytotoxic effect on human leukemia cell lines. Eur. J. Med. Chem. 2009, 44, 920–925. [Google Scholar] [CrossRef] [PubMed]
- Barnum, K.J.; O’Connell, M.J. Cell cycle regulation by checkpoints. Cell Cycle Control Mech. Protoc. 2014, 1170, 29–40. [Google Scholar]
- Karunagaran, D.; Rashmi, R.; Kumar, T.R. Induction of apoptosis by curcumin and its implications for cancer therapy. Curr. Cancer Drug Targets 2005, 5, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, I.; Grösch, S.; Schmidtko, A.; Häussler, A.; Schmidt, H.; Niederberger, E.; Scholich, K.; Geisslinger, G. G protein-independent G1 cell cycle block and apoptosis with morphine in adenocarcinoma cells: Involvement of p53 phosphorylation. Cancer Res. 2003, 63, 1846–1852. [Google Scholar] [PubMed]
- Senwar, K.R.; Sharma, P.; Reddy, T.S.; Jeengar, M.K.; Nayak, V.L.; Naidu, V.G.; Kamal, A.; Shankaraiah, N. Spirooxindole-derived morpholine-fused-1, 2, 3-triazoles: Design, synthesis, cytotoxicity and apoptosis inducing studies. Eur. J. Med. Chem. 2015, 102, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Zentar, H.; Jannus, F.; Gutierrez, P.; Medina-O’Donnell, M.; Lupiáñez, J.A.; Reyes-Zurita, F.J.; Alvarez-Manzaneda, E.; Chahboun, R. Semisynthesis and Evaluation of Anti-Inflammatory Activity of the Cassane-Type Diterpenoid Taepeenin F and of Some Synthetic Intermediates. J. Nat. Prod. 2022, 85, 2372–2384. [Google Scholar] [CrossRef]
- Xie, J.; Liu, J.; Liu, H.; Liang, S.; Lin, M.; Gu, Y.; Liu, T.; Wang, D.; Ge, H.; Mo, S.L. The antitumor effect of tanshinone IIA on anti-proliferation and decreasing VEGF/VEGFR2 expression on the human non-small cell lung cancer A549 cell line. Acta Pharm. Sin. B 2015, 5, 554–563. [Google Scholar] [CrossRef]
- Hu, Y.P.; Tan, Z.J.; Wu, X.S.; Liu, T.Y.; Jiang, L.; Bao, R.F.; Shu, Y.J.; Li, M.L.; Weng, H.; Ding, Q.; et al. Triptolide induces s phase arrest and apoptosis in gallbladder cancer cells. Molecules 2014, 19, 2612–2628. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Silva, C.O.; Monteiro, C.M.; Nicolai, M.; Viana, A.; Andrade, J.M.; Barasoain, I.; Stankovic, T.; Quintana, J.; Hernández, I.; et al. Anticancer properties of the abietane diterpene 6, 7-dehydroroyleanone obtained by optimized extraction. Future Med. Chem. 2018, 10, 1177–1189. [Google Scholar] [CrossRef]
- Cheng, A.C.; Lee, M.F.; Tsai, M.L.; Lai, C.S.; Lee, J.H.; Ho, C.T.; Pan, M.H. Rosmanol potently induces apoptosis through both the mitochondrial apoptotic pathway and death receptor pathway in human colon adenocarcinoma COLO 205 cells. Food Chem. Toxicol. 2011, 49, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and colon cancer. Gastroenterology 2010, 138, 2101–2114. [Google Scholar] [CrossRef]
- Maione, F.; Cantone, V.; Chini, M.G.; De Feo, V.; Mascolo, N.; Bifulco, G. Molecular mechanism of tanshinone IIA and cryptotanshinone in platelet anti-aggregating effects: An integrated study of pharmacology and computational analysis. Fitoterapia 2015, 100, 174–178. [Google Scholar] [CrossRef]
- Becker, K.; Schwaiger, S.; Waltenberger, B.; Fuchs, D.; Pezzei, C.K.; Schennach, H.; Stuppner, H.; Gostner, J.M. Immunomodulatory effects of diterpene quinone derivatives from the roots of Horminum pyrenaicum in human PBMC. Oxid. Med. Cell Longev. 2018, 2018, 2018. [Google Scholar] [CrossRef]
- Alizadeh, Z.; Farimani, M.M.; Parisi, V.; Marzocco, S.; Ebrahimi, S.N.; De Tommasi, N. Nor-abietane diterpenoids from Perovskia abrotanoides roots with anti-inflammatory potential. J. Nat. Prod. 2021, 84, 1185–1197. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Connelly, L.; Palacios-Callender, M.; Ameixa, C.; Moncada, S.; Hobbs, A.J. Biphasic regulation of NF-κB activity underlies the pro-and anti-inflammatory actions of nitric oxide. J. Immunol. 2001, 166, 3873–3881. [Google Scholar] [CrossRef]
- Bryan, N.S.; Grisham, M.B. Methods to Detect Nitric Oxide and Its Metabolites in Biological Samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef]
- Vega-Granados, K.; Medina-O’Donnell, M.; Rivas, F.; Reyes-Zurita, F.J.; Martinez, A.; Alvarez de Cienfuegos, L.; Lupiañez, J.A.; Parra, A. Synthesis and biological activity of triterpene–coumarin conjugates. J. Nat. Prod. 2021, 84, 1587–1597. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Compound | IC50 (μg/mL) | IC80 (μg/mL) |
|---|---|---|---|
| HT29 | Pygmaeocin B (5) | 6.69 ± 1.18 | 9.73 ± 1.18 |
| Saprorthoquinone (7) | 7.82 ± 0.17 | 10.62 ± 0.28 | |
| Pygmaeocin C (17) | 15.28 ± 0.71 | 24.36 ± 2.65 | |
| Viridoquinone (19) | 60.52 ± 0.28 | 74.65 ± 1.53 | |
| Precursor (13) | 2.67 ± 0.82 | 5.19 ± 0.68 | |
| HepG2 | Pygmaeocin B (5) | 8.99 ± 1.12 | 11.41 ± 1.08 |
| Saprorthoquinone (7) | 21.16 ± 2.26 | 29.12 ± 0.21 | |
| Pygmaeocin C (17) | 38.75 ± 1.55 | 73.53 ± 2.06 | |
| Viridoquinone (19) | 76.27 ± 2.14 | 100.57 ± 3.47 | |
| Precursor (13) | 5.58 ± 0.93 | 7.67 ± 0.80 | |
| B16-F10 | Pygmaeocin B (5) | 13.63 ± 0.14 | 14.04 ± 0.08 |
| Saprorthoquinone (7) | 6.89 ± 0.37 | 8.02 ± 0.52 | |
| Pygmaeocin C (17) | 10.11 ± 1.81 | 26.61 ± 4.82 | |
| Viridoquinone (19) | 6.42 ± 0.35 | 9.14 ± 0.38 | |
| Precursor (13) | 4.14 ± 0.25 | 5.41 ± 0.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ait El Had, M.; Zentar, H.; Ruiz-Muñoz, B.; Sainz, J.; Guardia, J.J.; Fernández, A.; Justicia, J.; Alvarez-Manzaneda, E.; Reyes-Zurita, F.J.; Chahboun, R. Evaluation of Anticancer and Anti-Inflammatory Activities of Some Synthetic Rearranged Abietanes. Int. J. Mol. Sci. 2023, 24, 13583. https://doi.org/10.3390/ijms241713583
Ait El Had M, Zentar H, Ruiz-Muñoz B, Sainz J, Guardia JJ, Fernández A, Justicia J, Alvarez-Manzaneda E, Reyes-Zurita FJ, Chahboun R. Evaluation of Anticancer and Anti-Inflammatory Activities of Some Synthetic Rearranged Abietanes. International Journal of Molecular Sciences. 2023; 24(17):13583. https://doi.org/10.3390/ijms241713583
Chicago/Turabian StyleAit El Had, Mustapha, Houda Zentar, Blanca Ruiz-Muñoz, Juan Sainz, Juan J. Guardia, Antonio Fernández, José Justicia, Enrique Alvarez-Manzaneda, Fernando J. Reyes-Zurita, and Rachid Chahboun. 2023. "Evaluation of Anticancer and Anti-Inflammatory Activities of Some Synthetic Rearranged Abietanes" International Journal of Molecular Sciences 24, no. 17: 13583. https://doi.org/10.3390/ijms241713583
APA StyleAit El Had, M., Zentar, H., Ruiz-Muñoz, B., Sainz, J., Guardia, J. J., Fernández, A., Justicia, J., Alvarez-Manzaneda, E., Reyes-Zurita, F. J., & Chahboun, R. (2023). Evaluation of Anticancer and Anti-Inflammatory Activities of Some Synthetic Rearranged Abietanes. International Journal of Molecular Sciences, 24(17), 13583. https://doi.org/10.3390/ijms241713583