
A Comprehensive Genome-Wide Association Study of Carotenoid and Capsaicinoid Contents in Capsicum chinense Germplasm


Abstract
:1. Introduction
2. Results
2.1. Carotenoids and Capsaicinoids Contents
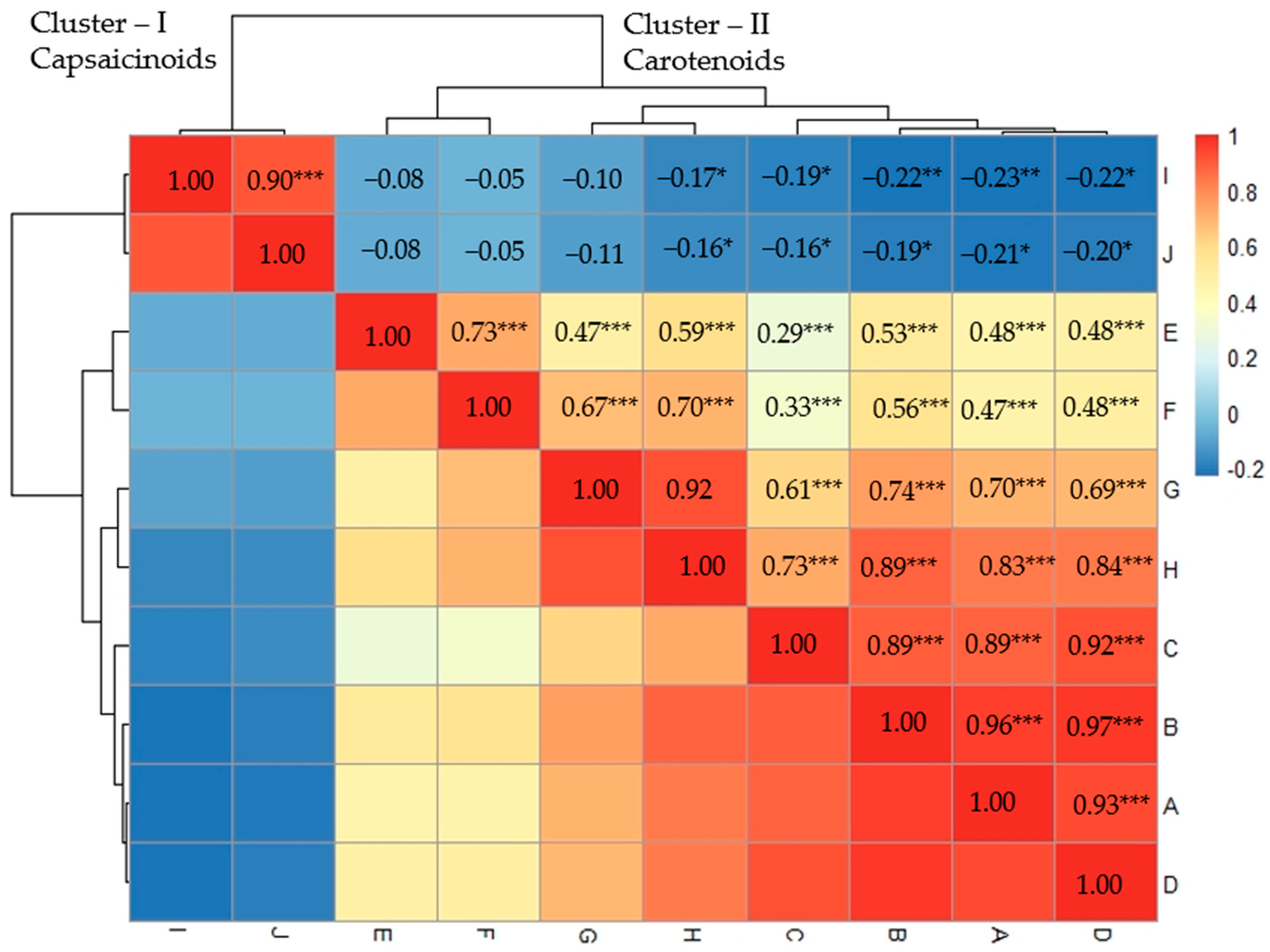
2.2. Correlation Analysis
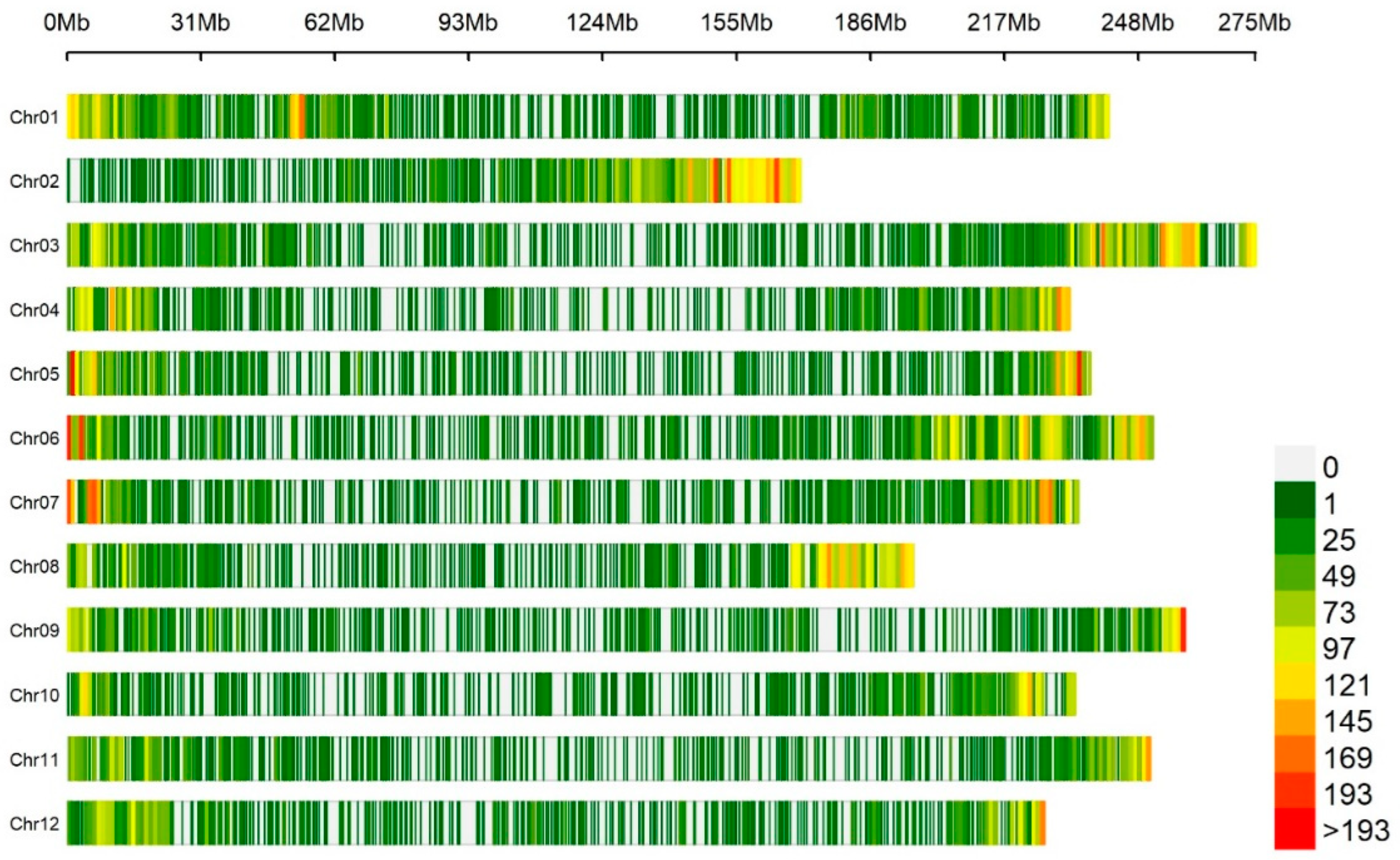
2.3. Genotyping-by-Sequencing
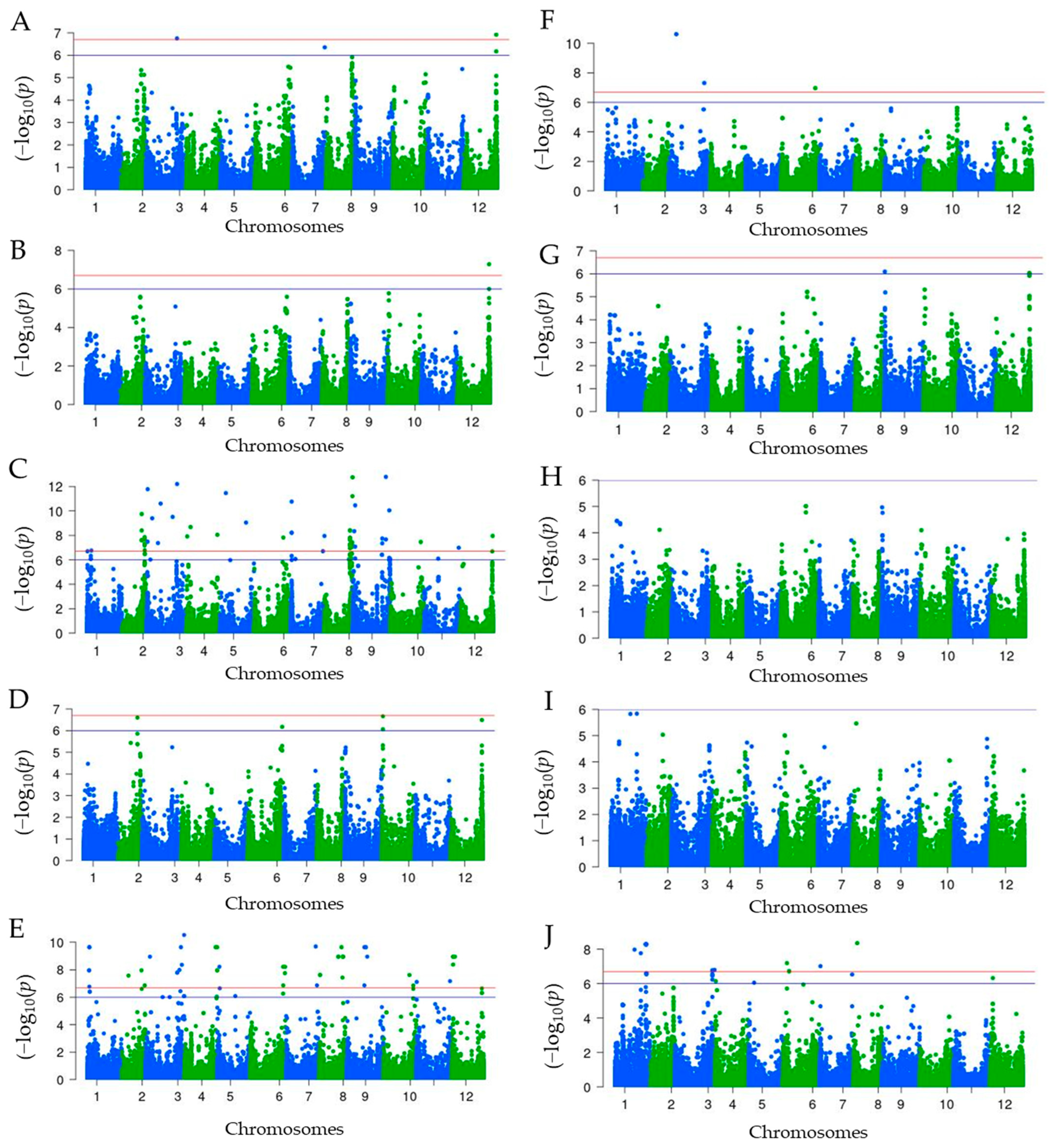
2.4. Genome-Wide Association Analysis
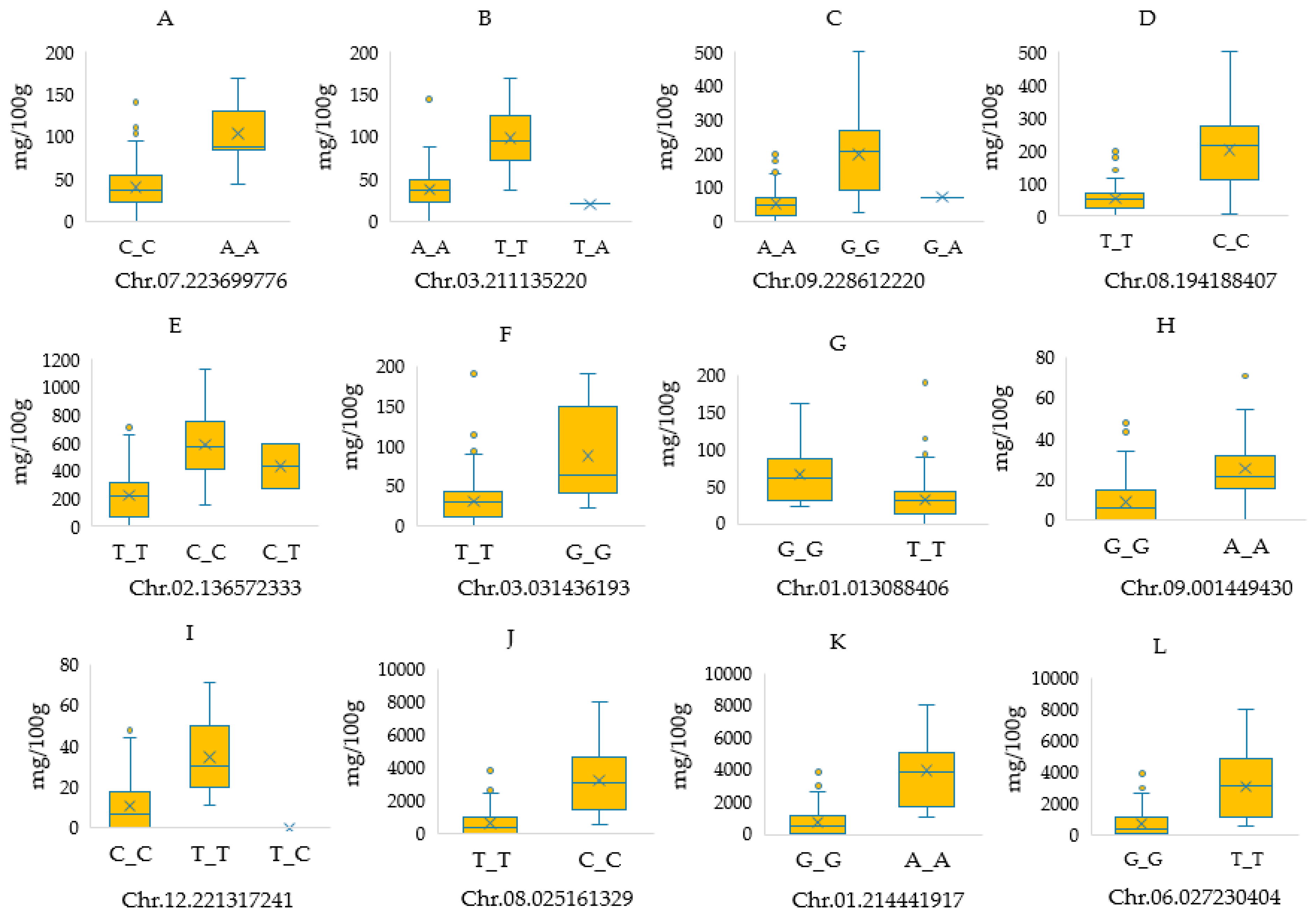
2.5. SNP Markers Showing Pleiotropic Effects
3. Discussion
3.1. Carotenoids and Capsaicinoids Contents
3.2. SNPs Associated to Carotenoids and Capsaicinoids
4. Materials and Methods
4.1. Chemicals and Plant Material
4.2. Analysis of Carotenoids
4.3. Analysis of Capsaicinoids
4.4. DNA Extraction and Genotype-by-Sequencing
4.5. SNP Calling and Filtering
4.6. Genome-Wide Association Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carrizo García, C.; Barfuss, M.H.J.; Sehr, E.M.; Barboza, G.E.; Samuel, R.; Moscone, E.A.; Ehrendorfer, F. Phylogenetic Relationships, Diversification and Expansion of Chili Peppers (Capsicum, Solanaceae). Ann. Bot. 2016, 118, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Pickersgill, B. Genetic Resources and Breeding of Capsicum Spp. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations Statistics Division. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 20 February 2023).
- de Sá Mendes, N.; de Andrade Gonçalves, É.C.B. The Role of Bioactive Components Found in Peppers. Trends Food Sci. Technol. 2020, 99, 229–243. [Google Scholar] [CrossRef]
- Conforti, F.; Statti, G.A.; Menichini, F. Chemical and Biological Variability of Hot Pepper Fruits (Capsicum annuum var. acuminatum L.) in Relation to Maturity Stage. Food Chem. 2007, 102, 1096–1104. [Google Scholar] [CrossRef]
- Gurnani, N.; Gupta, M.; Mehta, D.; Mehta, B.K. Chemical Composition, Total Phenolic and Flavonoid Contents, and in Vitro Antimicrobial and Antioxidant Activities of Crude Extracts from Red Chilli Seeds (Capsicum frutescens L.). J. Taibah Univ. Sci. 2016, 10, 462–470. [Google Scholar] [CrossRef]
- Lu, M.; Ho, C.-T.; Huang, Q. Extraction, Bioavailability, and Bioefficacy of Capsaicinoids. J. Food Drug Anal. 2017, 25, 27–36. [Google Scholar] [CrossRef]
- Van Duyn, M.A.S.; Pivonka, E. Overview of the Health Benefits of Fruit and Vegetable Consumption for the Dietetics Professional. J. Am. Diet. Assoc. 2000, 100, 1511–1521. [Google Scholar] [CrossRef]
- Cruz, J.G.; Silveira, T.; Richter, V.; Wagner, J.G.; Neitzke, R.S.; Barbieri, R.L.; Vizzotto, M. Genetic Variability of Bioactive Compounds in Capsicum chinense. Food Sci. Technol. 2022, 42, e123721. [Google Scholar] [CrossRef]
- Sarpras, M.; Chhapekar, S.S.; Ahmad, I.; Abraham, S.K.; Ramchiary, N. Analysis of Bioactive Components in Ghost Chili (Capsicum chinense) for Antioxidant, Genotoxic, and Apoptotic Effects in Mice. Drug Chem. Toxicol. 2020, 43, 182–191. [Google Scholar] [CrossRef]
- Sharma, J.; Sharma, P.; Sharma, B.; Chaudhary, P. In-Vitro Estimation of Antioxidant Activity in Green Chilli (Capsicum annuum) and Yellow Lantern Chilli (Capsicum chinense). Int. J. Res. Rev. 2017, 4, 54–61. [Google Scholar]
- Sherova, G.; Pavlov, A.; Georgiev, V. Polyphenols Profiles and Antioxidant Activities of Extracts from Capsicum chinense in Vitro Plants and Callus Cultures. FSAB 2019, 2, 30. [Google Scholar] [CrossRef]
- Salehi, B.; Hernández-Álvarez, A.J.; Del Mar Contreras, M.; Martorell, M.; Ramírez-Alarcón, K.; Melgar-Lalanne, G.; Matthews, K.R.; Sharifi-Rad, M.; Setzer, W.N.; Nadeem, M.; et al. Potential Phytopharmacy and Food Applications of Capsicum Spp.: A Comprehensive Review. Nat. Prod. Commun. 2018, 13, 1934578X1801301. [Google Scholar] [CrossRef]
- da Silveira Agostini-Costa, T.; da Silva Gomes, I.; de Melo, L.A.M.P.; Reifschneider, F.J.B.; da Costa Ribeiro, C.S. Carotenoid and Total Vitamin C Content of Peppers from Selected Brazilian Cultivars. J. Food Compos. Anal. 2017, 57, 73–79. [Google Scholar] [CrossRef]
- Rodríguez-Burruezo, A.; González-Mas, M.D.C.; Nuez, F. Carotenoid Composition and Vitamin A Value in Ají (Capsicum Baccatum L.) and Rocoto (C. pubescens R. & P.), 2 Pepper Species from the Andean Region. J. Food Sci. 2010, 75, S446–S453. [Google Scholar] [CrossRef]
- Carvalho, A.V.; de Andrade Mattietto, R.; de Oliveira Rios, A.; de Almeida Maciel, R.; Moresco, K.S.; de Souza Oliveira, T.C. Bioactive Compounds and Antioxidant Activity of Pepper (Capsicum sp.) Genotypes. J. Food Sci. Technol. 2015, 52, 7457–7464. [Google Scholar] [CrossRef]
- Thuphairo, K.; Sornchan, P.; Suttisansanee, U. Bioactive Compounds, Antioxidant Activity and Inhibition of Key Enzymes Relevant to Alzheimer’s Disease from Sweet Pepper (Capsicum annuum) Extracts. Prev. Nutr. Food Sci. 2019, 24, 327–337. [Google Scholar] [CrossRef]
- Mohd Hassan, N.; Yusof, N.A.; Yahaya, A.F.; Mohd Rozali, N.N.; Othman, R. Carotenoids of Capsicum Fruits: Pigment Profile and Health-Promoting Functional Attributes. Antioxidants 2019, 8, 469. [Google Scholar] [CrossRef]
- Bae, H.; Jayaprakasha, G.K.; Crosby, K.; Yoo, K.S.; Leskovar, D.I.; Jifon, J.; Patil, B.S. Ascorbic Acid, Capsaicinoid, and Flavonoid Aglycone Concentrations as a Function of Fruit Maturity Stage in Greenhouse-Grown Peppers. J. Food Compos. Anal. 2014, 33, 195–202. [Google Scholar] [CrossRef]
- Dias, A.L.B.; Arroio Sergio, C.S.; Santos, P.; Barbero, G.F.; Rezende, C.A.; Martínez, J. Effect of Ultrasound on the Supercritical CO2 Extraction of Bioactive Compounds from Dedo de Moça Pepper (Capsicum baccatum L. var. pendulum). Ultrason. Sonochem. 2016, 31, 284–294. [Google Scholar] [CrossRef]
- Menichini, F.; Tundis, R.; Bonesi, M.; Loizzo, M.; Conforti, F.; Statti, G.; Decindio, B.; Houghton, P.; Menichini, F. The Influence of Fruit Ripening on the Phytochemical Content and Biological Activity of Capsicum chinense Jacq. cv Habanero. Food Chem. 2009, 114, 553–560. [Google Scholar] [CrossRef]
- Collins, M.D.; Wasmund, L.M.; Bosland, P.W. Improved Method for Quantifying Capsaicinoids in Capsicum Using High-Performance Liquid Chromatography. HortScience 1995, 30, 137–139. [Google Scholar] [CrossRef]
- Peña-Alvarez, A.; Ramírez-Maya, E.; Alvarado-Suárez, L.Á. Analysis of Capsaicin and Dihydrocapsaicin in Peppers and Pepper Sauces by Solid Phase Microextraction–Gas Chromatography–Mass Spectrometry. J. Chromatogr. A 2009, 1216, 2843–2847. [Google Scholar] [CrossRef]
- Usman, M.; Rafii, M.; Ismail, M.; Malek, M.; Latif, M. Capsaicin and Dihydrocapsaicin Determination in Chili Pepper Genotypes Using Ultra-Fast Liquid Chromatography. Molecules 2014, 19, 6474–6488. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired Nociception and Pain Sensation in Mice Lacking the Capsaicin Receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The Capsaicin Receptor: A Heat-Activated Ion Channel in the Pain Pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.J.; Huang, S.M.; De Petrocellis, L.; Bisogno, T.; Ewing, S.A.; Miller, J.D.; Zipkin, R.E.; Daddario, N.; Appendino, G.; Di Marzo, V.; et al. N-Oleoyldopamine, a Novel Endogenous Capsaicin-like Lipid That Produces Hyperalgesia. J. Biol. Chem. 2003, 278, 13633–13639. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Yolton, R.L.; Yolton, D.P.; Schnider, C.; Janin, M.L. Personal Defense Sprays: Effects and Management of Exposure. J. Am. Optom. Assoc. 1996, 67, 548–560. [Google Scholar]
- Korte, A.; Farlow, A. The Advantages and Limitations of Trait Analysis with GWAS: A Review. Plant Methods 2013, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-Wide Association Studies. Nat. Rev. Methods Primers 2021, 1, 59. [Google Scholar] [CrossRef]
- Iquira, E.; Humira, S.; François, B. Association Mapping of QTLs for Sclerotinia Stem Rot Resistance in a Collection of Soybean Plant Introductions Using a Genotyping by Sequencing (GBS) Approach. BMC Plant Biol. 2015, 15, 5. [Google Scholar] [CrossRef]
- Lin, M.; Cai, S.; Wang, S.; Liu, S.; Zhang, G.; Bai, G. Genotyping-by-Sequencing (GBS) Identified SNP Tightly Linked to QTL for Pre-Harvest Sprouting Resistance. Theor. Appl. Genet. 2015, 128, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Owens, B.F.; Lipka, A.E.; Magallanes-Lundback, M.; Tiede, T.; Diepenbrock, C.H.; Kandianis, C.B.; Kim, E.; Cepela, J.; Mateos-Hernandez, M.; Buell, C.R.; et al. A Foundation for Provitamin A Biofortification of Maize: Genome-Wide Association and Genomic Prediction Models of Carotenoid Levels. Genetics 2014, 198, 1699–1716. [Google Scholar] [CrossRef] [PubMed]
- Poland, J.A.; Brown, P.J.; Sorrells, M.E.; Jannink, J.-L. Development of High-Density Genetic Maps for Barley and Wheat Using a Novel Two-Enzyme Genotyping-by-Sequencing Approach. PLoS ONE 2012, 7, e32253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Kong, W.; Robertson, J.; Goff, V.H.; Epps, E.; Kerr, A.; Mills, G.; Cromwell, J.; Lugin, Y.; Phillips, C.; et al. Genetic Analysis of Inflorescence and Plant Height Components in Sorghum (Panicoidae) and Comparative Genetics with Rice (Oryzoidae). BMC Plant Biol. 2015, 15, 107. [Google Scholar] [CrossRef] [PubMed]
- Hamed, M.; Kalita, D.; Bartolo, M.E.; Jayanty, S.S. Capsaicinoids, Polyphenols and Antioxidant Activities of Capsicum annuum: Comparative Study of the Effect of Ripening Stage and Cooking Methods. Antioxidants 2019, 8, 364. [Google Scholar] [CrossRef]
- Rodríguez-Rodríguez, E.; Sánchez-Prieto, M.; Olmedilla-Alonso, B. Assessment of Carotenoid Concentrations in Red Peppers (Capsicum annuum) under Domestic Refrigeration for Three Weeks as Determined by HPLC-DAD. Food Chem. X 2020, 6, 100092. [Google Scholar] [CrossRef]
- Elvira-Torales, L.I.; García-Alonso, J.; Periago-Castón, M.J. Nutritional Importance of Carotenoids and Their Effect on Liver Health: A Review. Antioxidants 2019, 8, 229. [Google Scholar] [CrossRef]
- Giuffrida, D.; Dugo, P.; Torre, G.; Bignardi, C.; Cavazza, A.; Corradini, C.; Dugo, G. Characterization of 12 Capsicum Varieties by Evaluation of Their Carotenoid Profile and Pungency Determination. Food Chem. 2013, 140, 794–802. [Google Scholar] [CrossRef]
- Ben-Chaim, A.; Borovsky, Y.; Falise, M.; Mazourek, M.; Kang, B.-C.; Paran, I.; Jahn, M. QTL Analysis for Capsaicinoid Content in Capsicum. Theor. Appl. Genet. 2006, 113, 1481–1490. [Google Scholar] [CrossRef]
- Zewdie, Y.; Bosland, P.W. Evaluation of Genotype, Environment, and Genotype-by-Environment Interaction for Capsaicinoids in Capsicum annuum L. Euphytica 2000, 111, 185–190. [Google Scholar] [CrossRef]
- Kosuge, S.; Furuta, M. Studies on the Pungent Principle of Capsicum. Agric. Biol. Chem. 1970, 34, 248–256. [Google Scholar] [CrossRef]
- Antonious, G.F.; Kochhar, T.S.; Jarret, R.L.; Snyder, J.C. Antioxidants in Hot Pepper: Variation Among Accessions. J. Environ. Sci. Health Part B 2006, 41, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- González-Zamora, A.; Sierra-Campos, E.; Luna-Ortega, J.; Pérez-Morales, R.; Ortiz, J.; García-Hernández, J. Characterization of Different Capsicum Varieties by Evaluation of Their Capsaicinoids Content by High Performance Liquid Chromatography, Determination of Pungency and Effect of High Temperature. Molecules 2013, 18, 13471–13486. [Google Scholar] [CrossRef]
- Mori, A.; Lehmann, S.; O’Kelly, J.; Kumagai, T.; Desmond, J.C.; Pervan, M.; McBride, W.H.; Kizaki, M.; Koeffler, H.P. Capsaicin, a Component of Red Peppers, Inhibits the Growth of Androgen-Independent, P53 Mutant Prostate Cancer Cells. Cancer Res. 2006, 66, 3222–3229. [Google Scholar] [CrossRef] [PubMed]
- Laskaridou-Monnerville, A. Determination of Capsaicin and Dihydrocapsaicin by Micellar Electrokinetic Capillary Chromatography and Its Application to Various Species of Capsicum, Solanaceae. J. Chromatogr. A 1999, 838, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Ro, N.; Haile, M.; Ko, H.-C.; Cho, G.-T.; Lee, J.; Kim, B.; Lee, S.; Kim, S.-H. Genome-Wide Association Study of Phenolic Content and Antioxidant Properties in Eggplant Germplasm. Genes 2023, 14, 1315. [Google Scholar] [CrossRef]
- Sukhumsirichart, W. Polymorphisms. In Genetic Diversity and Disease Susceptibility; Liu, Y., Ed.; InTech: London, UK, 2018; ISBN 978-1-78984-201-2. [Google Scholar]
- Greenbaum, J.; Wu, K.; Zhang, L.; Shen, H.; Zhang, J.; Deng, H.-W. Increased Detection of Genetic Loci Associated with Risk Predictors of Osteoporotic Fracture Using a Pleiotropic CFDR Method. Bone 2017, 99, 62–68. [Google Scholar] [CrossRef]
- Reddy, U.K.; Lopez-Ortiz, C.; Talavera-Caro, A.G.; Natarajan, P.; Tomason, Y.; Alaparthi, S.; Levi, A.; Nimmakayala, P. GWAS Resolves Molecular Mechanisms Underlying Natural Variation for Carotenoids in Cucurbita Maxima Duchesne. Sci. Hortic. 2023, 312, 111881. [Google Scholar] [CrossRef]
- Kang, C.; He, S.; Zhai, H.; Li, R.; Zhao, N.; Liu, Q. A Sweetpotato Auxin Response Factor Gene (IbARF5) Is Involved in Carotenoid Biosynthesis and Salt and Drought Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1307. [Google Scholar] [CrossRef]
- Zhang, Q.; Gong, M.; Xu, X.; Li, H.; Deng, W. Roles of Auxin in the Growth, Development, and Stress Tolerance of Horticultural Plants. Cells 2022, 11, 2761. [Google Scholar] [CrossRef]
- Wang, P.; Shen, L.; Guo, J.; Jing, W.; Qu, Y.; Li, W.; Bi, R.; Xuan, W.; Zhang, Q.; Zhang, W. Phosphatidic Acid Directly Regulates PINOID-Dependent Phosphorylation and Activation of the PIN-FORMED2 Auxin Efflux Transporter in Response to Salt Stress. Plant Cell 2019, 31, 250–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, T.; Wang, R.; Zhao, Y. Recent Advances in Auxin Research in Rice and Their Implications for Crop Improvement. J. Exp. Bot. 2018, 69, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Matthes, M.S.; Best, N.B.; Robil, J.M.; Malcomber, S.; Gallavotti, A.; McSteen, P. Auxin EvoDevo: Conservation and Diversification of Genes Regulating Auxin Biosynthesis, Transport, and Signaling. Mol. Plant 2019, 12, 298–320. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Ahmar, S.; Ali, B.; Saleem, M.H.; Khan, M.U.; Zhou, W.; Liu, S. The Role of Membrane Transporters in Plant Growth and Development, and Abiotic Stress Tolerance. Int. J. Mol. Sci. 2021, 22, 12792. [Google Scholar] [CrossRef]
- Harper, R.M.; Stowe-Evans, E.L.; Luesse, D.R.; Muto, H.; Tatematsu, K.; Watahiki, M.K.; Yamamoto, K.; Liscum, E. The NPH4 Locus Encodes the Auxin Response Factor ARF7, a Conditional Regulator of Differential Growth in Aerial Arabidopsis Tissue. Plant Cell 2000, 12, 757–770. [Google Scholar] [CrossRef]
- Waller, F.; Furuya, M.; Nick, P. OsARF1, an Auxin Response Factor from Rice, Is Auxin-Regulated and Classifies as a Primary Auxin Responsive Gene. Plant Mol. Biol. 2002, 50, 415–425. [Google Scholar] [CrossRef]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 Regulate Senescence and Floral Organ Abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef]
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D.; et al. Functional Genomic Analysis of the AUXIN RESPONSE FACTOR Gene Family Members in Arabidopsis thaliana: Unique and Overlapping Functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef]
- Wilmoth, J.C.; Wang, S.; Tiwari, S.B.; Joshi, A.D.; Hagen, G.; Guilfoyle, T.J.; Alonso, J.M.; Ecker, J.R.; Reed, J.W. NPH4/ARF7 and ARF19 Promote Leaf Expansion and Auxin-Induced Lateral Root Formation: ARF Proteins Regulate Auxin-Induced Root Formation. Plant J. 2005, 43, 118–130. [Google Scholar] [CrossRef]
- Wang, L.; Hua, D.; He, J.; Duan, Y.; Chen, Z.; Hong, X.; Gong, Z. Auxin Response Factor2 (ARF2) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef]
- Ren, Z.; Liu, R.; Gu, W.; Dong, X. The Solanum Lycopersicum Auxin Response Factor SlARF2 Participates in Regulating Lateral Root Formation and Flower Organ Senescence. Plant Sci. 2017, 256, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Ledent, V.; Vervoort, M. The Basic Helix-Loop-Helix Protein Family: Comparative Genomics and Phylogenetic Analysis. Genome Res. 2001, 11, 754–770. [Google Scholar] [CrossRef]
- Toledo-Ortiz, G.; Huq, E.; Quail, P.H. The Arabidopsis Basic/Helix-Loop-Helix Transcription Factor Family. Plant Cell 2003, 15, 1749–1770. [Google Scholar] [CrossRef]
- Liu, R.; Song, J.; Liu, S.; Chen, C.; Zhang, S.; Wang, J.; Xiao, Y.; Cao, B.; Lei, J.; Zhu, Z. Genome-Wide Identification of the Capsicum BHLH Transcription Factor Family: Discovery of a Candidate Regulator Involved in the Regulation of Species-Specific Bioactive Metabolites. BMC Plant Biol. 2021, 21, 262. [Google Scholar] [CrossRef]
- Luo, F.; Cai, J.-H.; Kong, X.-M.; Zhou, Q.; Zhou, X.; Zhao, Y.-B.; Ji, S.-J. Transcriptome Profiling Reveals the Roles of Pigment Mechanisms in Postharvest Broccoli Yellowing. Hortic. Res. 2019, 6, 74. [Google Scholar] [CrossRef]
- Bouige, P.; Laurent, D.; Piloyan, L.; Dassa, E. Phylogenetic and Functional Classification of ATP-Binding Cassette (ABC) Systems. Curr. Protein Pept. Sci. 2002, 3, 541–559. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Lee, S.Y.; Chu, S.M.; Lim, S.H.; Suh, S.-C.; Lee, Y.-T.; Cho, H.S.; Ha, S.-H. Variation and Correlation Analysis of Flavonoids and Carotenoids in Korean Pigmented Rice (Oryza sativa L.) Cultivars. J. Agric. Food Chem. 2010, 58, 12804–12809. [Google Scholar] [CrossRef]
- Lee, H.-Y.; Ro, N.-Y.; Jeong, H.-J.; Kwon, J.-K.; Jo, J.; Ha, Y.; Jung, A.; Han, J.-W.; Venkatesh, J.; Kang, B.-C. Genetic Diversity and Population Structure Analysis to Construct a Core Collection from a Large Capsicum Germplasm. BMC Genet. 2016, 17, 142. [Google Scholar] [CrossRef]
- De Donato, M.; Peters, S.O.; Mitchell, S.E.; Hussain, T.; Imumorin, I.G. Genotyping-by-Sequencing (GBS): A Novel, Efficient and Cost-Effective Genotyping Method for Cattle Using Next-Generation Sequencing. PLoS ONE 2013, 8, e62137. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-Glance Quality Assessment of Illumina Second-Generation Sequencing Data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Oh, S.-K.; Lee, J.-H.; Lee, B.-M.; Jo, S.-H. Genome-Wide SNP Calling Using Next Generation Sequencing Data in Tomato. Mol. Cells 2014, 37, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Browning, S.R.; Browning, B.L. Rapid and Accurate Haplotype Phasing and Missing-Data Inference for Whole-Genome Association Studies By Use of Localized Haplotype Clustering. Am. J. Hum. Genet. 2007, 81, 1084–1097. [Google Scholar] [CrossRef] [PubMed]
- QTLmax Global. QTLmax 3.0: A Genetic Analysis Toolset for Enterprises. 2022. Katy, TX, USA. Available online: https://www.qtlmax.com (accessed on 20 February 2023).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Range (mg/100 g) | Mean (mg/100 g) | SE | SD | Count |
|---|---|---|---|---|---|
| Violaxanthin | 0–168.89 | 43.91 | 2.61 | 33.01 | 160 |
| Antheraxanthin | 0–445.35 | 127.53 | 7.64 | 96.61 | 160 |
| Capsorubin | 0–501.4 | 76.53 | 6.50 | 82.24 | 160 |
| Capsanthin | 0–1120.95 | 299.92 | 18.31 | 231.59 | 160 |
| Zeaxanthin | 0–190.36 | 34.45 | 2.47 | 31.24 | 160 |
| β-Cryptoxanthin | 0–81.81 | 8.37 | 0.85 | 10.71 | 160 |
| α-Carotene | 0–70.87 | 12.40 | 1.06 | 13.45 | 160 |
| β-Carotene | 0–184.33 | 54.09 | 2.96 | 37.42 | 160 |
| Capsaicin | 0–11,632.93 | 1836.97 | 158.68 | 2007.22 | 160 |
| Dihydrocapsaicin | 0–8038.17 | 892.58 | 91.01 | 1151.15 | 160 |
| Traits | Chr. | Reference Allele | Alternate Allele | Position (bp.) | p.Value | −Log10 (p.Value) | Feature | Minor Allele | Major Allele |
|---|---|---|---|---|---|---|---|---|---|
| Violaxanthin | 12 | G | A | 221,085,307 | 1.21 × 10−7 | 6.92 | Genic | A | G |
| 03 | T | A | 211,135,220 | 1.77 × 10−7 | 6.75 | Genic | T | A | |
| 07 | A | C | 223,699,776 | 4.43 × 10−7 | 6.35 | Intergenic | A | C | |
| 12 | C | T | 221,105,042 | 6.71 × 10−7 | 6.17 | Intergenic | T | C | |
| Antheraxanthin | 12 | G | A | 221,085,307 | 5.24 × 10−8 | 7.28 | Genic | A | G |
| 12 | C | T | 221,105,042 | 1.02 × 10−6 | 6.00 | Intergenic | T | C | |
| Capsorubin | 09 | G | A | 228,612,220 | 1.65 × 10−13 | 12.78 | Intergenic | G | A |
| 08 | C | T | 194,188,407 | 1.77 × 10−13 | 12.75 | Genic | C | T | |
| 03 | T | A | 211,135,220 | 6.32 × 10−13 | 12.20 | Genic | T | A | |
| 03 | C | T | 7,882,123 | 1.67 × 10−12 | 11.78 | Genic | C | T | |
| 05 | G | G | 41,033,949 | 3.47 × 10−12 | 11.46 | Genic | G | C | |
| 08 | C | A | 193,486,347 | 6.22 × 10−12 | 11.21 | Intergenic | C | A | |
| 07 | C | A | 7,431,933 | 1.73 × 10−11 | 10.76 | Genic | A | C | |
| 03 | A | G | 97,513,286 | 2.51 × 10−11 | 10.60 | Intergenic | A | G | |
| 09 | C | T | 16,302,649 | 3.53 × 10−11 | 10.45 | Intergenic | C | T | |
| 09 | G | C | 252,016,648 | 9.07 × 10−11 | 10.04 | Genic | G | C | |
| 02 | C | T | 136,572,333 | 1.79 × 10−10 | 9.75 | Genic | C | T | |
| 03 | A | G | 180,540,684 | 3.08 × 10−10 | 9.51 | Intergenic | A | G | |
| 03 | T | A | 39,034,826 | 4.04 × 10−10 | 9.39 | Genic | T | A | |
| 05 | T | C | 180,424,789 | 9.00 × 10−10 | 9.05 | Genic | T | C | |
| 04 | A | G | 29,271,696 | 2.06 × 10−9 | 8.69 | Genic | A | G | |
| 08 | A | G | 176,546,501 | 3.91 × 10−9 | 8.41 | Genic | A | G | |
| 08 | A | G | 176,546,659 | 3.91 × 10−9 | 8.41 | Genic | A | G | |
| 02 | A | G | 136,573,788 | 3.94 × 10−9 | 8.40 | Intergenic | A | G | |
| 09 | T | C | 10,001,201 | 4.58 × 10−9 | 8.34 | Genic | T | C | |
| 07 | C | T | 7,389,025 | 6.12 × 10−9 | 8.21 | Intergenic | T | C | |
| 07 | T | C | 7,389,133 | 6.12 × 10−9 | 8.21 | Intergenic | C | T | |
| 04 | A | C | 214,280,150 | 8.99 × 10−9 | 8.05 | Intergenic | A | C | |
| 12 | A | G | 223,949,835 | 1.11 × 10−8 | 7.95 | Intergenic | A | G | |
| 07 | C | A | 233,954,670 | 1.12 × 10−8 | 7.95 | Genic | C | A | |
| Capsanthin | 10 | T | C | 8,626,608 | 2.17 × 10−7 | 6.66 | Intergenic | C | T |
| 02 | C | T | 136,572,333 | 2.49 × 10−7 | 6.60 | Genic | C | T | |
| 12 | G | A | 221,085,307 | 3.24 × 10−7 | 6.49 | Genic | A | G | |
| 06 | G | A | 240,758,527 | 6.70 × 10−7 | 6.17 | Genic | A | G | |
| 10 | A | G | 8,625,878 | 8.68 × 10−7 | 6.06 | Intergenic | G | A | |
| Zeaxanthin | 03 | T | C | 273,399,119 | 2.99 × 10−11 | 10.52 | Genic | C | T |
| 07 | C | T | 209,703,812 | 2.03 × 10−10 | 9.69 | Intergenic | T | C | |
| 01 | C | C | 13,088,352 | 2.29 × 10−10 | 9.64 | Intergenic | T | C | |
| 01 | T | T | 13,088,406 | 2.29 × 10−10 | 9.64 | Intergenic | G | T | |
| 03 | T | C | 250,691,656 | 2.29 × 10−10 | 9.64 | Genic | C | T | |
| 04 | A | C | 221,648,109 | 2.29 × 10−10 | 9.64 | Genic | C | A | |
| 04 | G | A | 232,075,190 | 2.29 × 10−10 | 9.64 | Genic | A | G | |
| 08 | A | G | 158,075,413 | 2.29 × 10−10 | 9.64 | Intergenic | G | A | |
| 09 | A | C | 124,336,652 | 2.29 × 10−10 | 9.64 | Intergenic | C | A | |
| 09 | A | G | 135,166,449 | 2.29 × 10−10 | 9.64 | Genic | G | A | |
| 03 | T | G | 31,436,193 | 1.10 × 10−9 | 8.96 | Genic | G | T | |
| 08 | T | C | 135,871,114 | 1.10 × 10−9 | 8.96 | Genic | C | T | |
| 08 | G | A | 135,910,583 | 1.10 × 10−9 | 8.96 | Genic | A | G | |
| 08 | G | A | 167,703,221 | 1.10 × 10−9 | 8.96 | Genic | A | G | |
| 08 | T | A | 167,703,365 | 1.10 × 10−9 | 8.96 | Genic | A | T | |
| 08 | C | T | 170,330,854 | 1.10 × 10−9 | 8.96 | Genic | T | C | |
| 09 | A | C | 144,268,010 | 1.10 × 10−9 | 8.96 | Intergenic | C | A | |
| 12 | G | C | 9,887,870 | 1.10 × 10−9 | 8.96 | Genic | C | G | |
| 12 | T | C | 10,065,787 | 1.10 × 10−9 | 8.96 | Intergenic | C | T | |
| 12 | T | C | 23,244,295 | 1.10 × 10−9 | 8.96 | Intergenic | C | T | |
| 12 | T | A | 10,062,899 | 4.11 × 10−9 | 8.39 | Intergenic | A | T | |
| β-Cryptoxanthin | 03 | A | G | 50,199,834 | 2.41 × 10−11 | 10.62 | Genic | G | A |
| 03 | C | T | 234,486,991 | 4.89 × 10−8 | 7.31 | Intergenic | T | C | |
| 06 | C | T | 222,803,463 | 1.08 × 10−7 | 6.97 | Genic | T | C | |
| α-Carotene | 09 | A | G | 1,449,430 | 8.02 × 10−7 | 6.10 | Genic | A | G |
| 12 | C | T | 221,317,241 | 9.17 × 10−7 | 6.04 | genic | T | C | |
| Dihydrocapsaicin | 08 | T | C | 25,161,329 | 4.61 × 10−9 | 8.34 | Intergenic | C | T |
| 01 | A | G | 208,666,749 | 5.36 × 10−9 | 8.27 | Genic | G | A | |
| 01 | A | T | 209,615,478 | 5.36 × 10−9 | 8.27 | Genic | T | A | |
| 01 | G | A | 214,441,815 | 5.36 × 10−9 | 8.27 | Intergenic | A | G | |
| 01 | G | A | 214,441,917 | 5.36 × 10−9 | 8.27 | Intergenic | A | G | |
| 01 | C | T | 214,442,584 | 5.36 × 10−9 | 8.27 | Intergenic | T | C | |
| 01 | T | C | 132,753,120 | 1.07 × 10−8 | 7.97 | Intergenic | C | T | |
| 01 | C | A | 176,184,702 | 1.72 × 10−8 | 7.77 | Genic | A | C | |
| 06 | G | T | 27,230,404 | 6.43 × 10−8 | 7.19 | Genic | T | G | |
| 07 | A | C | 6,571,083 | 9.69 × 10−8 | 7.01 | Intergenic | C | A |
| Traits | Chr. | Allele | Positions (bp.) | −Log10 (p-Value) | Feature | Gene | Descriptions |
|---|---|---|---|---|---|---|---|
| Violaxanthin, antheraxanthin, capsorubin and capsanthin | Chr12 | A/G | 221,085,307 | 6.92 | Genic | CA12g19380 | 6,7-dimethyl-8-ribityllumazine synthase |
| Violaxanthin and antheraxanthin | Chr12 | T/C | 221,105,042 | 6.01 | Intergenic | - | - |
| Violaxanthin and capsorubin | Chr07 | A/C | 223,699,776 | 6.35 | Intergenic | - | - |
| Chr03 | A/T | 211,135,220 | 6.75 | Genic | CA03g18160 | 4-hydroxycinnamoyl-CoA ligase 2 | |
| Capsorubin and capsanthin | Chr02 | T/C | 136,572,333 | 9.75 | Genic | CA02g25010 | Detected protein of unknown function |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haile, M.; Ro, N.; Ko, H.-C.; Oh, H.; Lee, G.-A. A Comprehensive Genome-Wide Association Study of Carotenoid and Capsaicinoid Contents in Capsicum chinense Germplasm. Int. J. Mol. Sci. 2023, 24, 13885. https://doi.org/10.3390/ijms241813885
Haile M, Ro N, Ko H-C, Oh H, Lee G-A. A Comprehensive Genome-Wide Association Study of Carotenoid and Capsaicinoid Contents in Capsicum chinense Germplasm. International Journal of Molecular Sciences. 2023; 24(18):13885. https://doi.org/10.3390/ijms241813885
Chicago/Turabian StyleHaile, Mesfin, Nayoung Ro, Ho-Cheol Ko, Hyeonseok Oh, and Gi-An Lee. 2023. "A Comprehensive Genome-Wide Association Study of Carotenoid and Capsaicinoid Contents in Capsicum chinense Germplasm" International Journal of Molecular Sciences 24, no. 18: 13885. https://doi.org/10.3390/ijms241813885
APA StyleHaile, M., Ro, N., Ko, H. -C., Oh, H., & Lee, G. -A. (2023). A Comprehensive Genome-Wide Association Study of Carotenoid and Capsaicinoid Contents in Capsicum chinense Germplasm. International Journal of Molecular Sciences, 24(18), 13885. https://doi.org/10.3390/ijms241813885