
Transcriptomic Analysis of Metarhizium anisopliae-Induced Immune-Related Long Non-Coding RNAs in Polymorphic Worker Castes of Solenopsis invicta

 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Sequencing Analysis and Quality Assessment
2.2. Genomic Characterization and Identification of lncRNAs
2.3. M. anisopliae-Responsive lncRNAs in Major and Minor Worker Ants
2.4. Functional Analysis of M. anisopliae-Responsive lncRNA Target Genes
2.5. Detection of S. invicta lncRNAs as Pre-miRNAs
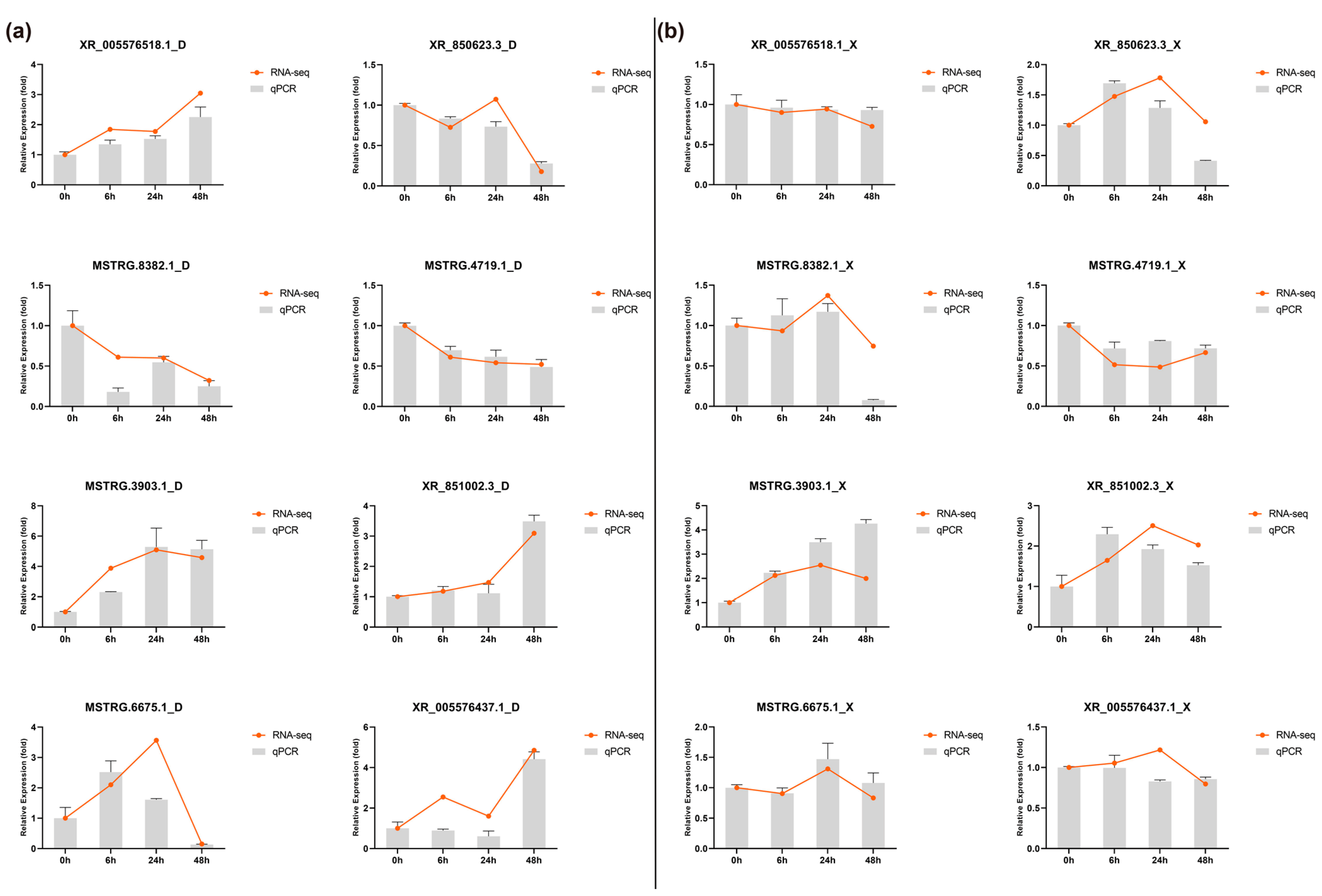
2.6. Real-Time Quantitative PCR Analysis
3. Discussion
4. Materials and Methods
4.1. Insects
4.2. M. anisopliae Infection and RNA Sequencing
4.3. Read Filters and Transcript Assembly
4.4. Identification, Annotation and Quantification of lncRNAs
4.5. Prediction of Pre-miRNAs and Target Genes of lncRNAs
4.6. Real-Time Quantitative PCR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and abundance analysis of human lncRNAs at single-cell and single-molecule resolution. Genom. Biol. 2015, 16, 20. [Google Scholar]
- Knauss, J.L.; Sun, T. Regulatory mechanisms of long noncoding RNAs in vertebrate central nervous system development and function. Neuroscience 2013, 235, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Mulvey, B.B.; Olcese, U.; Cabrera, J.R.; Horabin, J.I. An interactive network of long non-coding RNAs facilitates the Drosophila sex determination decision. Biochim. Biophys. Acta 2014, 1839, 773–784. [Google Scholar] [CrossRef]
- Sebastian-delaCruz, M.; Gonzalez-Moro, I.; Olazagoitia-Garmendia, A.; Castellanos-Rubio, A.; Santin, I. The Role of lncRNAs in Gene Expression Regulation through mRNA Stabilization. Noncoding RNA 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, 17–29. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, biology and functioning. In Non-coding RNAs in Colorectal Cancer; Springer: Berlin/Heidelberg, Germany, 2016; pp. 3–17. [Google Scholar]
- Zhang, G.; Lan, Y.; Xie, A.; Shi, J.; Zhao, H.; Xu, L.; Zhu, S.; Luo, T.; Zhao, T.; Xiao, Y. Comprehensive analysis of long noncoding RNA (lncRNA)-chromatin interactions reveals lncRNA functions dependent on binding diverse regulatory elements. J. Biol. Chem. 2019, 294, 15613–15622. [Google Scholar] [CrossRef]
- Pisignano, G.; Ladomery, M. Epigenetic regulation of alternative splicing: How LncRNAs Tailor the message. Noncoding RNA 2021, 7, 21. [Google Scholar] [CrossRef]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucleic Acids Res. 2018, 46, 2169–2184. [Google Scholar]
- Kim, M.; Faucillion, M.-L.; Larsson, J. RNA-on-X 1 and 2 in Drosophila melanogaster fulfill separate functions in dosage compensation. PLoS Genet. 2018, 14, e1007842. [Google Scholar] [CrossRef]
- Zhang, S.; Yin, H.; Shen, M.; Huang, H.; Hou, Q.; Zhang, Z.; Zhao, W.; Guo, X.; Wu, P. Analysis of lncRNA-mediated gene regulatory network of Bombyx mori in response to BmNPV infection. J. Invertebr. Pathol. 2020, 170, 107323. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.M.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, T.; He, W.; Shen, X.; Zhao, Q.; Bai, J.; You, M. Genome-wide identification and characterization of putative lncRNAs in the diamondback moth, Plutella xylostella (L.). Genomics 2018, 110, 35–42. [Google Scholar] [CrossRef]
- Chang, Z.X.; Ajayi, O.E.; Guo, D.Y.; Wu, Q.F. Genome-wide characterization and developmental expression profiling of long non-coding RNAs in Sogatella furcifera. Insect Sci. 2020, 27, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Shi, T.; Qi, L.; Su, X.; Wang, D.; Dong, J.; Huang, Z.Y. lncRNA profile of Apis mellifera and its possible role in behavioural transition from nurses to foragers. BMC Genom. 2019, 20, 393. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, Y.W.; Lu, Y.Y.; Li, B.; Chen, K.P.; Li, C.J. Genome-wide identification and characterization of long non-coding RNAs in Tribolium castaneum. Insect Sci. 2021, 28, 1262–1276. [Google Scholar] [CrossRef]
- Wu, Y.; Cheng, T.; Liu, C.; Liu, D.; Zhang, Q.; Long, R.; Zhao, P.; Xia, Q. Systematic identification and characterization of long non-coding RNAs in the silkworm, Bombyx mori. PLoS ONE 2016, 11, e0147147. [Google Scholar] [CrossRef]
- Liu, W.; Cheng, P.; Zhang, K.; Gong, M.; Zhang, Z.; Zhang, R. Systematic identification and characterization of long noncoding RNAs (lncRNAs) during Aedes albopictus development. PLoS Negl. Trop. Dis. 2022, 16, e0010245. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, Y.; Yang, S.; Zhang, H.; Wu, Q.; Chen, F. An integrated evolutionary analysis of miRNA–lncRNA in mammals. Mol. Biol. Rep. 2014, 41, 201–207. [Google Scholar] [CrossRef]
- Wang, H.-L.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. In Long Non Coding RNA Biology; Rao, M.R.S., Ed.; Springer: Singapore, 2017; pp. 133–154. [Google Scholar]
- Ma, C.; Zhao, S.; Yan, S.; Wang, Y.; Lin, Q.; Fang, Y.; Liu, H.; Wu, P. Analysis on new occurrence records of red imported fire ant (Solenopsis invicta Buren) in China in 2021. Entomol. Res. 2023, 53, 55–65. [Google Scholar] [CrossRef]
- Wetterer, J.K. Exotic spread of Solenopsis invicta buren (Hymenoptera: Formicidae) beyond North america. Sociobiology 2013, 60, 50–55. [Google Scholar] [CrossRef]
- Vinson, S.B. Invasion of the red imported fire ant (Hymenoptera: Formicidae): Spread, biology, and impact. Am. Entomol. 1997, 43, 23–39. [Google Scholar] [CrossRef]
- Wetterer, J.K.; Davis, L.R. Solenopsis invicta (Hymenoptera: Formicidae) in the Lesser Antilles. Fla. Entomol. 2010, 93, 128–129, 122. [Google Scholar] [CrossRef]
- Sutherst, R.W.; Maywald, G. A Climate Model of the Red Imported Fire Ant, Solenopsis invicta Buren (Hymenoptera: Formicidae): Implications for Invasion of New Regions, Particularly Oceania. Environ. Entomol. 2005, 34, 317–335. [Google Scholar] [CrossRef]
- Wylie, R.; Jennings, C.; McNaught, M.K.; Oakey, J.; Harris, E.J. Eradication of two incursions of the Red Imported Fire Ant in Queensland, Australia. Ecol. Manag. Restor. 2016, 17, 22–32. [Google Scholar] [CrossRef]
- Ujiyama, S.; Tsuji, K. Controlling invasive ant species: A theoretical strategy for efficient monitoring in the early stage of invasion. Sci. Rep. 2018, 8, 8033. [Google Scholar] [CrossRef]
- Lyu, D.P.; Lee, H.S. The red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae: Myrmicinae) discovered in Busan sea port, Korea. Korean J. Appl. Entomol. 2017, 56, 437–438. [Google Scholar]
- Zeng, L.; Lu, Y.; He, X.; Zhang, W.; Liang, G. Identification of red imported fire ant, Solenopsis invicta, to invade mainland China and infestation in Wuchuan, Guangdong. Chin. Bull. Entomol. 2005, 42, 144–148. [Google Scholar]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.; Keller, L. Recent human history governs global ant invasion dynamics. Nat. Ecol. Evol. 2017, 1, 0184. [Google Scholar] [CrossRef]
- Lard, C.F.; Schmidt, J.; Morris, B.; Estes, L.; Ryan, C.; Bergquist, D. An Economic Impact of Imported Fire Ants in the United States of America; Texas A & M University, Department of Agricultural Economics, Texas Agricultural Experiment Station: College Station, TX, USA, 2006. [Google Scholar]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; Volume 12. [Google Scholar]
- Fan, Y.; Pereira, R.M.; Kilic, E.; Casella, G.; Keyhani, N.O. Pyrokinin β-neuropeptide affects necrophoretic behavior in fire ants (S. invicta), and expression of β-NP in a mycoinsecticide increases its virulence. PLoS ONE 2012, 7, e26924. [Google Scholar] [CrossRef]
- Wang, L.; Zeng, L.; Xu, Y.; Lu, Y. Prevalence and management of Solenopsis invicta in China. NeoBiota 2020, 54, 89–124. [Google Scholar] [CrossRef]
- Du, Y.; Grodowitz, M.J.; Chen, J. Insecticidal and enzyme inhibitory activities of isothiocyanates against red imported fire ants, Solenopsis invicta. Biomolecules 2020, 10, 716. [Google Scholar] [CrossRef]
- Zafar, J.; Freed, S.; Khan, B.A.; Farooq, M. Effectiveness of Beauveria bassiana against cotton whitefly, Bemisia tabaci (Gennadius)(Aleyrodidae: Homoptera) on different host plants. Pak. J. Zool. 2016, 48, 91–99. [Google Scholar]
- Shoukat, R.F.; Hassan, B.; Shakeel, M.; Zafar, J.; Li, S.; Freed, S.; Xu, X.; Jin, F. Pathogenicity and transgenerational effects of Metarhizium anisopliae on the demographic parameters of Aedes albopictus (Culicidae: Diptera). J. Med. Entomol. 2020, 57, 677–685. [Google Scholar] [CrossRef]
- Mwamburi, L.A. Endophytic fungi, Beauveria bassiana and Metarhizium anisopliae, confer control of the fall armyworm, Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae), in two tomato varieties. Egypt J. Biol. Pest Control. 2021, 31, 7. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungus Metarhizium anisopliae. Biocontrol Sci. Technol. 2007, 17, 879–920. [Google Scholar] [CrossRef]
- Wei, Z.; Ortiz-Urquiza, A.; Keyhani, N.O. Altered Expression of Chemosensory and Odorant Binding Proteins in Response to Fungal Infection in the Red Imported Fire Ant, Solenopsis invicta. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef]
- Kafle, L.; Wu, W.J.; Kao, S.S.; Shih, C.J. Efficacy of Beauveria bassiana against the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae), in Taiwan. Pest Manag. Sci. 2011, 67, 1434–1438. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.-L.; Fox, E.G.; Qin, C.-S.; Zhao, D.-Y.; Yang, H.; Xu, J.-Z. Microcapsuled entomopathogenic fungus against fire ants, Solenopsis invicta. Biol. Control 2019, 134, 141–149. [Google Scholar] [CrossRef]
- Ugelvig, L.V.; Cremer, S. Effects of social immunity and unicoloniality on host–parasite interactions in invasive insect societies. Funct. Ecol. 2012, 26, 1300–1312. [Google Scholar] [CrossRef]
- Wang, L.; Elliott, B.; Jin, X.; Zeng, L.; Chen, J. Antimicrobial properties of nest volatiles in red imported fire ants, Solenopsis invicta (hymenoptera: Formicidae). Sci. Nat. 2015, 102, 66. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-L.; Leger, R.S. Insect Immunity to Entomopathogenic Fungi. In Advances in Genetics; Brian Lovett, R.J.S.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 94, pp. 251–285. [Google Scholar]
- Moure, U.A.E.; Tan, T.; Sha, L.; Lu, X.; Shao, Z.; Yang, G.; Wang, Y.; Cui, H. Advances in the Immune Regulatory Role of Non-Coding RNAs (miRNAs and lncRNAs) in Insect-Pathogen Interactions. Front. Immunol. 2022, 13, 856457. [Google Scholar] [CrossRef] [PubMed]
- Starkey, J.; Tamborindeguy, C. Family before work: Task reversion in workers of the red imported fire ant, Solenopsis invicta in the presence of brood. Sci. Rep. 2023, 13, 2379. [Google Scholar] [CrossRef] [PubMed]
- Power, K.T.; An, R.; Grewal, P.S. Effectiveness of Heterohabditis bacteriophora strain GPS11 applications targeted against different instars of the Japanese beetle Popillia japonica. Biol. Control 2009, 48, 232–236. [Google Scholar] [CrossRef]
- Williams, L.; Cherry, R.; Shapiro-Ilan, D. Effect of Host Size on Susceptibility of Melanotus Communis (Coleoptera: Elateridae) Wireworms to Entomopathogens. J. Nematol. 2022, 54, 20220033. [Google Scholar] [CrossRef]
- Byhrø, E.M.H.; Salmela, H.; Vitlic, A.; Wang, Y.; Münch, D.; Amdam, G.V. Different activation of immune-related genes in honey bee nurses and foragers (Apis mellifera). Apidologie 2019, 50, 463–471. [Google Scholar] [CrossRef]
- Ren, C.; Webster, P.; Finkel, S.E.; Tower, J. Increased Internal and External Bacterial Load during Drosophila Aging without Life-Span Trade-Off. Cell Metab. 2007, 6, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.L.; Zhao, C.Y.; He, Y.R. On the Molecular Basis of Division of Labor in Solenopsis invicta (Hymenoptera: Formicidae) Workers: RNA-seq Analysis. J. Insect Sci. 2017, 17. [Google Scholar] [CrossRef]
- Zhou, Q.-Z.; Zhang, B.; Yu, Q.-Y.; Zhang, Z. BmncRNAdb: A comprehensive database of non-coding RNAs in the silkworm, Bombyx mori. BMC Bioinform. 2016, 17, 370. [Google Scholar] [CrossRef]
- Liu, F.; Guo, D.; Yuan, Z.; Chen, C.; Xiao, H. Genome-wide identification of long non-coding RNA genes and their association with insecticide resistance and metamorphosis in diamondback moth, Plutella xylostella. Sci. Rep. 2017, 7, 15870. [Google Scholar] [CrossRef]
- Wills, B.D.; Powell, S.; Rivera, M.D.; Suarez, A.V. Correlates and Consequences of Worker Polymorphism in Ants. Annu. Rev. Entomol. 2018, 63, 575–598. [Google Scholar] [CrossRef]
- Zafar, J.; Huang, J.; Xu, X.; Jin, F. Analysis of Long Non-Coding RNA-Mediated Regulatory Networks of Plutella xylostella in Response to Metarhizium anisopliae Infection. Insects 2022, 13, 916. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhou, Y.; Li, H. LncRNA, miRNA and lncRNA-miRNA interaction in viral infection. Virus Res. 2018, 257, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, K.; Vilcinskas, A. Development and immunity-related microRNAs of the lepidopteran model host Galleria mellonella. BMC Genom. 2014, 15, 705. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Asgari, S. Conserved microRNA miR-8 blocks activation of the Toll pathway by upregulating Serpin 27 transcripts. RNA Biol. 2013, 10, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.J.; Hyun, S. Multiple targets of the microRNA miR-8 contribute to immune homeostasis in Drosophila. Dev. Comp. Immunol. 2014, 45, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, X.-Y.; Tang, Q.-B.; Lei, C.-L.; Huang, Q.-Y. The Mechanisms of Social Immunity Against Fungal Infections in Eusocial Insects. Toxins 2019, 11, 244. [Google Scholar] [CrossRef]
- Agliano, F.; Rathinam, V.A.; Medvedev, A.E.; Vanaja, S.K.; Vella, A.T. Long Noncoding RNAs in Host-Pathogen Interactions. Trends Immunol. 2019, 40, 492–510. [Google Scholar] [CrossRef]
- Shields, E.J.; Sheng, L.; Weiner, A.K.; Garcia, B.A.; Bonasio, R. High-Quality Genome Assemblies Reveal Long Non-coding RNAs Expressed in Ant Brains. Cell Rep. 2018, 23, 3078–3090. [Google Scholar] [CrossRef]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, discovery, and classification of lncRNAs. Long Non Coding RNA Biol. 2017, 1008, 1–46. [Google Scholar]
- Valadkhan, S.; Valencia-Hipólito, A. lncRNAs in Stress Response. In Long Non-Coding RNAs in Human Disease; Morris, K.V., Ed.; Springer International Publishing: Cham, Switzerland, 2016; Volume 394, pp. 203–236. [Google Scholar]
- Mao, W.; Zeng, Q.; She, L.; Yuan, H.; Luo, Y.; Wang, R.; She, Y.; Wang, W.; Wang, C.; Pan, X. Wolbachia Utilizes lncRNAs to Activate the Anti-Dengue Toll Pathway and Balance Reactive Oxygen Species Stress in Aedes aegypti through a Competitive Endogenous RNA Network. Front. Cell. Infect. Microbiol. 2022, 11, 823403. [Google Scholar] [CrossRef]
- Chen, D.; Chen, H.; Du, Y.; Zhou, D.; Geng, S.; Wang, H.; Wan, J.; Xiong, C.; Zheng, Y.; Guo, R. Genome-wide identification of long non-coding RNAs and their regulatory networks involved in Apis mellifera ligustica response to Nosema ceranae infection. Insects 2019, 10, 245. [Google Scholar] [CrossRef]
- Li, B.; Hou, L.; Zhu, D.; Xu, X.; An, S.; Wang, X. Identification and caste-dependent expression patterns of DNA methylation associated genes in Bombus terrestris. Sci. Rep. 2018, 8, 2332. [Google Scholar] [CrossRef] [PubMed]
- Scharf, M.E.; Wu-Scharf, D.; Pittendrigh, B.R.; Bennett, G.W. Caste-and development-associated gene expression in a lower termite. Genom. Biol. 2003, 4, R62. [Google Scholar] [CrossRef]
- Husseneder, C.; Simms, D.M. Effects of caste on the expression of genes associated with septic injury and xenobiotic exposure in the Formosan subterranean termite. PLoS ONE 2014, 9, e105582. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.M.; Kingwell, C.J.; Wcislo, W.T.; Robinson, G.E. Caste-biased gene expression in a facultatively eusocial bee suggests a role for genetic accommodation in the evolution of eusociality. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162228. [Google Scholar] [CrossRef] [PubMed]
- Mitaka, Y.; Kobayashi, K.; Matsuura, K. Caste-, sex-, and age-dependent expression of immune-related genes in a Japanese subterranean termite, Reticulitermes speratus. PLoS ONE 2017, 12, e0175417. [Google Scholar] [CrossRef]
- Koch, S.I.; Groh, K.; Vogel, H.; Hannson, B.S.; Kleineidam, C.J.; Grosse-Wilde, E. Caste-specific expression patterns of immune response and chemosensory related genes in the leaf-cutting ant, Atta vollenweideri. PLoS ONE 2013, 8, e81518. [Google Scholar] [CrossRef]
- Feldmeyer, B.; Elsner, D.; Foitzik, S. Gene expression patterns associated with caste and reproductive status in ants: Worker-specific genes are more derived than queen-specific ones. Mol. Ecol. 2014, 23, 151–161. [Google Scholar] [CrossRef]
- Ometto, L.; Shoemaker, D.; Ross, K.G.; Keller, L. Evolution of Gene Expression in Fire Ants: The Effects of Developmental Stage, Caste, and Species. Mol. Biol. Evol. 2010, 28, 1381–1392. [Google Scholar] [CrossRef]
- Chouvenc, T.; Su, N.-Y.; Robert, A. Cellular encapsulation in the eastern subterranean termite, Reticulitermes flavipes (Isoptera), against infection by the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2009, 101, 234–241. [Google Scholar] [CrossRef]
- Sun, Y.-X.; Chen, C.; Xu, W.-J.; Abbas, M.N.; Mu, F.-F.; Ding, W.-J.; Zhang, H.-J.; Li, J. Functions of Bombyx mori cathepsin L-like in innate immune response and anti-microbial autophagy. Dev. Comp. Immunol. 2021, 116, 103927. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sun, Y.; Chang, M.; Zhang, Y.; Qiao, H.; Huang, S.; Kan, Y.; Yao, L.; Li, D.; Ayra-Pardo, C. RNA Interference-Mediated Knockdown of Bombyx mori Haemocyte-Specific Cathepsin L (Cat L)-Like Cysteine Protease Gene Increases Bacillus thuringiensis kurstaki Toxicity and Reproduction in Insect Cadavers. Toxins 2022, 14, 394. [Google Scholar] [CrossRef] [PubMed]
- Akpinar, B.A.; Muslu, T.; Reddy, G.V.P.; Dogramaci, M.; Budak, H. Wheat Long Noncoding RNAs from Organelle and Nuclear Genomes Carry Conserved microRNA Precursors Which May Together Comprise Intricate Networks in Insect Responses. Int. J. Mol. Sci. 2023, 24, 2226. [Google Scholar] [CrossRef] [PubMed]
- Fullaondo, A.; Lee, S.Y. Identification of putative miRNA involved in Drosophila melanogaster immune response. Dev. Comp. Immunol. 2012, 36, 267–273. [Google Scholar] [CrossRef]
- Shi, T.-F.; Wang, Y.-F.; Liu, F.; Qi, L.; Yu, L.-S. Influence of the Neonicotinoid Insecticide Thiamethoxam on miRNA Expression in the Honey Bee (Hymenoptera: Apidae). J. Insect Sci. 2017, 17, 96. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, Y.; Lei, Y.; Shi, Q.; Qi, G.; He, Y.; Lyu, L. Role of the foraging gene in worker behavioral transition in the red imported fire ant, Solenopsis invicta (Hymenoptera: Formicidae). Pest Manag. Sci. 2022, 78, 2964–2975. [Google Scholar] [CrossRef]
- Qiu, H.-L.; Lu, L.-H.; Shi, Q.-X.; He, Y.-R. Fungus Exposed Solenopsis invicta Ants Benefit from Grooming. J. Insect Behav. 2014, 27, 678–691. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genom. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wei, B.; Liu, H.; Li, T.; Rayner, S. MiRPara: A SVM-based software tool for prediction of most probable microRNA coding regions in genome scale sequences. BMC Bioinform. 2011, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Gil, N.; Ulitsky, I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet. 2020, 21, 102–117. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA | Target Gene ID | Gene Strand | Target Description | Abundant Expression (Major/Minor Ants) |
|---|---|---|---|---|
| M6hD vs. M6hX | ||||
| XR_850979.3 | LOC113004832 | - | Cytochrome | Major |
| XR_005575015.1 | LOC105207451 | + | ||
| XR_005575418.1 | LOC105193078 | - | AChE | Minor |
| XR_005576519.1 | LOC113004380 | - | Serine protease | Major |
| LOC120359735 | - | |||
| LOC105196739 | - | CLIP protease 2 | ||
| LOC105204892 | - | |||
| MSTRG.1438.10 | LOC105201965 | + | FAF1 | Major |
| MSTRG.3312.1 | LOC105208238 | + | Cathepsin L | Minor |
| MSTRG.4719.1 | LOC105204661 | + | Cytochrome P450 | Minor |
| MSTRG.4919.3 | LOC105204695 | + | AhR | Major |
| MSTRG.6347.1 | LOC105200144 | + | TRAF | Minor |
| MSTRG.8317.1 | LOC105205263 | - | Aminopeptidase N | Minor |
| MSTRG.16558.2 | LOC105206459 | + | Cytochrome P450 | Major |
| MSTRG.16671.16 | LOC120359317 | + | Major | |
| M24hD vs. M24hX | ||||
| XR_850979.3 | LOC113004832 | - | Cytochrome P450 | Major |
| MSTRG.19934.21 | LOC105202834 | + | ||
| LOC105202818 | + | |||
| MSTRG.4719.1 | LOC105204661 | + | Minor | |
| MSTRG.18135.1 | LOC105201710 | + | Minor | |
| XR_005575418.1 | LOC105193078 | - | AChE | Minor |
| MSTRG.10978.9 | LOC105206581 | + | Caspase | Minor |
| LOC105207826 | + | |||
| LOC105194248 | + | |||
| M48hD vs. M48hX | ||||
| XR_005574228.1 | LOC105200920 | - | Serine protease | Major |
| XR_005575418.1 | LOC105193078 | - | AChE | Minor |
| MSTRG.12826.9 | LOC105194452 | + | JNK-interacting protein 1 | Minor |
| XR_005574580.1 | LOC105204409 | - | Cytochrome P450 | Minor |
| MSTRG.8353.3 | LOC105193893 | + | Minor | |
| XR_850979.3 | LOC113004832 | - | Major | |
| MSTRG.4719.1 | LOC105204661 | + | Minor | |
| MSTRG.19934.21 | LOC105202834 | + | Major | |
| LOC105202818 | + | |||
| MSTRG.16671.16 | LOC120359317 | + | Minor | |
| MSTRG.18075.1 | LOC105201710 | + | Cytochrome b5 | Major |
| + | Minor | |||
| MSTRG.18135.1 | + | |||
| XR_850838.3 | LOC105197801 | - | TRAF3 | Major |
| LOC105193099 | - | Ankyrin-1 | Major | |
| LOC105193095 | - | SR-B1 | Major | |
| XR_005576517.1 | LOC105204787 | - | Melanization protease-1 | Major |
| LOC113004380 | - | Serine protease | ||
| LOC120359735 | - | |||
| LOC105196739 | - | CLIP serine protease-2 | ||
| LOC105194356 | - | Transmembrane protease serine | ||
| XR_005576437.1 | LOC105193099 | - | Ankyrin-1 | Major |
| MSTRG.10978.9 | LOC105206581 | + | Caspase-1 | Minor |
| LOC105207826 | + | |||
| LOC105194248 | + | |||
| MSTRG.6344.3 | LOC105200144 | + | TRAF | Minor |
| MSTRG.12460.18 | LOC105198898 | - | Trypsin-1 | Major |
| MSTRG.12460.53 | - | Major | ||
| MSTRG.12460.67 | - | Major | ||
| MSTRG.14976.1 | LOC113005542 | - | Spaetzle-3 | Major |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar, J.; Wu, H.; Xu, Y.; Lin, L.; Kang, Z.; Zhang, J.; Zhang, R.; Lu, Y.; Jin, F.; Xu, X. Transcriptomic Analysis of Metarhizium anisopliae-Induced Immune-Related Long Non-Coding RNAs in Polymorphic Worker Castes of Solenopsis invicta. Int. J. Mol. Sci. 2023, 24, 13983. https://doi.org/10.3390/ijms241813983
Zafar J, Wu H, Xu Y, Lin L, Kang Z, Zhang J, Zhang R, Lu Y, Jin F, Xu X. Transcriptomic Analysis of Metarhizium anisopliae-Induced Immune-Related Long Non-Coding RNAs in Polymorphic Worker Castes of Solenopsis invicta. International Journal of Molecular Sciences. 2023; 24(18):13983. https://doi.org/10.3390/ijms241813983
Chicago/Turabian StyleZafar, Junaid, Hongxin Wu, Yating Xu, Liangjie Lin, Zehong Kang, Jie Zhang, Ruonan Zhang, Yongyue Lu, Fengliang Jin, and Xiaoxia Xu. 2023. "Transcriptomic Analysis of Metarhizium anisopliae-Induced Immune-Related Long Non-Coding RNAs in Polymorphic Worker Castes of Solenopsis invicta" International Journal of Molecular Sciences 24, no. 18: 13983. https://doi.org/10.3390/ijms241813983
APA StyleZafar, J., Wu, H., Xu, Y., Lin, L., Kang, Z., Zhang, J., Zhang, R., Lu, Y., Jin, F., & Xu, X. (2023). Transcriptomic Analysis of Metarhizium anisopliae-Induced Immune-Related Long Non-Coding RNAs in Polymorphic Worker Castes of Solenopsis invicta. International Journal of Molecular Sciences, 24(18), 13983. https://doi.org/10.3390/ijms241813983