
A Systematic Review and Meta-Analysis of Free Triiodothyronine (FT3) Levels in Humans Depending on Seasonal Air Temperature Changes: Is the Variation in FT3 Levels Related to Nonshivering Thermogenesis?

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Methods
2.1. Sources of Information and Search Strategy
2.2. Eligibility Criteria
2.3. Selection Process
2.4. Quality Control
2.5. Data Analysis
2.6. Subgroups
3. Results
3.1. Systematic Review and Meta-Analysis of Variations in FT3 Levels in Relation to Seasonal Air Temperatures
Seasonal Variations of FT3 Levels Relation with Air Temperature, Depression, and Photoperiodism
4. Discussion
4.1. The Dynamics of T3 Levels in Response to Cold Exposure
4.2. T3 and Brown Adipose Tissue
4.3. T3 and Adaptive and “Activated” Thermogenesis
4.4. The T3 Stimulation of the Adaptive Thermogenesis Pathways under the Influence of Different Winter Temperature
4.5. The Regulation by the TSH of the T3 Levels under the Adaptive and “Mild” Adaptive Thermogenesis
4.6. The Relationship of Thyroid Hormones with Basal Metabolic Rate under Cold Exposure
4.7. Perspective and Further Study
4.8. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chiellini, G.; Apriletti, J.W.; Yoshihara, H.A.; Baxter, J.D.; Ribeiro, R.C.; Scanlan, T.S. A High-Affinity Subtype-Selective Agonist Ligand for the Thyroid Hormone Receptor. Chem. Biol. 1998, 5, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Oetting, A.; Yen, P.M. New Insights into Thyroid Hormone Action. Best. Pract. Res. Clin. Endocrinol. Metab. 2007, 21, 193–208. [Google Scholar] [CrossRef]
- Halperin, Y.; Shapiro, L.E.; Surks, M.I. Medium 3,5,3’-Triiodo-L-Thyronine (T3) and T3 Generated from L-Thyroxine Are Exchangeable in Cultured GC Cells. Endocrinology 1990, 127, 1050–1056. [Google Scholar] [CrossRef]
- Sapin, R.; Schlienger, J.-L.; Goichot, B.; Gasser, F.; Grucker, D. Evaluation of Elecsys® Free Triiodothyronine Assay: Relevance of Age-Related Reference Ranges. Clin. Biochem. 1998, 31, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Pirahanchi, Y.; Toro, F.; Jialal, I. Physiology, Thyroid Stimulating Hormone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- St Germain, D.L.; Galton, V.A.; Hernandez, A. Minireview: Defining the Roles of the Iodothyronine Deiodinases: Current Concepts and Challenges. Endocrinology 2009, 150, 1097–1107. [Google Scholar] [CrossRef]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamus-Pituitary-Thyroid Axis. Compr. Physiol. 2016, 6, 1387–1428. [Google Scholar] [CrossRef]
- Maes, M.; Mommen, K.; Hendrickx, D.; Peeters, D.; D’Hondt, P.; Ranjan, R.; De Meyer, F.; Scharpe, S. Components of Biological Variation, Including Seasonality, in Blood Concentrations of TSH, TT3, FT4, PRL, Cortisol and Testosterone in Healthy Volunteers. Clin. Endocrinol. 1997, 46, 587–598. [Google Scholar] [CrossRef]
- Hassi, J.; Sikkila, K.; Ruokonen, A.; Leppaluoto, J. The Pituitary-Thyroid Axis in Healthy Men Living under Subarctic Climatological Conditions. J. Endocrinol. 2001, 169, 195–203. [Google Scholar] [CrossRef]
- Jang, Y.-Y.; Kim, C.-Y.; Hwang, T.-Y.; Kim, K.-D.; Lee, C.-H. Reference Interval of Serum Thyroid Hormones in Healthy Korean Adults. J. Prev. Med. Public Health 2008, 41, 128. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Leonard, W.R.; Tarskaia, L.A.; Klimova, T.M.; Fedorova, V.I.; Baltakhinova, M.E.; Krivoshapkin, V.G.; Snodgrass, J.J. Seasonal and Socioeconomic Influences on Thyroid Function among the Yakut (Sakha) of Eastern Siberia: Seasonal Thyroid Changes in the Yakut. Am. J. Hum. Biol. 2013, 25, 814–820. [Google Scholar] [CrossRef]
- Chen, N.; Wu, Q.; Li, H.; Zhang, T.; Xu, C. Different Adaptations of Chinese Winter-over Expeditioners during Prolonged Antarctic and Sub-Antarctic Residence. Int. J. Biometeorol. 2016, 60, 737–747. [Google Scholar] [CrossRef]
- Wang, D.; Cheng, X.; Yu, S.; Qiu, L.; Lian, X.; Guo, X.; Hu, Y.; Lu, S.; Yang, G.; Liu, H. Data Mining: Seasonal and Temperature Fluctuations in Thyroid-Stimulating Hormone. Clin. Biochem. 2018, 60, 59–63. [Google Scholar] [CrossRef]
- Yoshihara, A.; Noh, J.Y.; Watanabe, N.; Iwaku, K.; Kunii, Y.; Ohye, H.; Suzuki, M.; Matsumoto, M.; Suzuki, N.; Sugino, K.; et al. Seasonal Changes in Serum Thyrotropin Concentrations Observed from Big Data Obtained During Six Consecutive Years from 2010 to 2015 at a Single Hospital in Japan. Thyroid 2018, 28, 429–436. [Google Scholar] [CrossRef]
- Zeng, Y.; He, H.; Wang, X.; Zhang, M.; An, Z. Climate and Air Pollution Exposure Are Associated with Thyroid Function Parameters: A Retrospective Cross-Sectional Study. J. Endocrinol. Investig. 2021, 44, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenko, N.V.; Tsyrlin, V.A.; Pliss, M.G.; Galagudza, M.M. Seasonal Variations in Levels of Human Thyroid-Stimulating Hormone and Thyroid Hormones: A Meta-Analysis. Chronobiol. Int. 2021, 38, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Evseeva, S.A.; Burtseva, T.E.; Klimova, T.M.; Danilov, N.A.; Egorova, V.B.; Chasnyk, V.G. Seasonal changes in the pituitary-thyroid system in children of the Arctic region of Yakutia. Yakut Med. J. 2021, 71, 89–91. [Google Scholar] [CrossRef]
- Yamada, S.; Horiguchi, K.; Akuzawa, M.; Sakamaki, K.; Shimomura, Y.; Kobayashi, I.; Andou, Y.; Yamada, M. Seasonal Variation in Thyroid Function in Over 7000 Healthy Subjects in an Iodine-Sufficient Area and Literature Review. J. Endocr. Soc. 2022, 6, bvac054. [Google Scholar] [CrossRef]
- Santi, D.; Spaggiari, G.; Brigante, G.; Setti, M.; Tagliavini, S.; Trenti, T.; Simoni, M. Semi-Annual Seasonal Pattern of Serum Thyrotropin in Adults. Sci. Rep. 2019, 9, 10786. [Google Scholar] [CrossRef]
- Tendler, A.; Bar, A.; Mendelsohn-Cohen, N.; Karin, O.; Korem Kohanim, Y.; Maimon, L.; Milo, T.; Raz, M.; Mayo, A.; Tanay, A.; et al. Hormone Seasonality in Medical Records Suggests Circannual Endocrine Circuits. Proc. Natl. Acad. Sci. USA 2021, 118, e2003926118. [Google Scholar] [CrossRef]
- Reed, H.L.; Silverman, E.D.; Shakir, K.M.M.; Dons, R.; Burman, K.D.; O’Brian, J.T. Changes in Serum Triiodothyronine (T3) Kinetics after Prolonged Antarctic Residence: The Polar T3 Syndrome. J. Clin. Endocrinol. Metab. 1990, 70, 965–974. [Google Scholar] [CrossRef]
- Reed, H.L.; Brice, D.; Shakir, K.M.; Burman, K.D.; D’Alesandro, M.M.; O’Brian, J.T. Decreased Free Fraction of Thyroid Hormones after Prolonged Antarctic Residence. J. Appl. Physiol. 1990, 69, 1467–1472. [Google Scholar] [CrossRef] [PubMed]
- Reed, H.L.; Ferreiro, J.A.; Mohamed Shakir, K.M.; Burman, K.D.; O’Brian, J.T. Pituitary and Peripheral Hormone Responses to T3 Administration during Antarctic Residence. Am. J. Physiol.-Endocrinol. Metab. 1988, 254, E733–E739. [Google Scholar] [CrossRef] [PubMed]
- Del Ponte, A.; Guagnano, M.T.; Sensi, S. Time-Related Behaviour of Endocrine Secretion: Circannual Variations of FT3, Cortisol, HGH and Serum Basal Insulin in Healthy Subjects. Chronobiol. Int. 1984, 1, 297–300. [Google Scholar] [CrossRef]
- Gullo, D.; Latina, A.; Frasca, F.; Squatrito, S.; Belfiore, A.; Vigneri, R. Seasonal Variations in TSH Serum Levels in Athyreotic Patients under L-thyroxine Replacement Monotherapy. Clin. Endocrinol. 2017, 87, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Mahwi, T.O.; Abdulateef, D.S. Relation of Different Components of Climate with Human Pituitary-Thyroid Axis and FT3/FT4 Ratio: A Study on Euthyroid and SCH Subjects in Two Different Seasons. Int. J. Endocrinol. 2019, 2019, 2762978. [Google Scholar] [CrossRef]
- Pasquali, R.; Baraldi, G.; Casimirri, F.; Mattioli, L.; Capelli, M.; Melchionda, N.; Capani, F.; Labò, G. Seasonal Variations of Total and Free Thyroid Hormones in Healthy Men: A Chronobiological Study. Acta Endocrinol. 1984, 107, 42–48. [Google Scholar] [CrossRef]
- Kanikowska, D.; Sato, M.; Iwase, S.; Shimizu, Y.; Nishimura, N.; Inukai, Y.; Sugenoya, J. Effects of Living at Two Ambient Temperatures on 24-h Blood Pressure and Neuroendocrine Function among Obese and Non-Obese Humans: A Pilot Study. Int. J. Biometeorol. 2013, 57, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhang, G.; Xu, P.; Guo, R.; Li, J.; Guan, H.; Li, Y. Seasonal Changes of Thyroid Function Parameters in Women of Reproductive Age Between 2012 and 2018: A Retrospective, Observational, Single-Center Study. Front. Endocrinol. 2021, 12, 719225. [Google Scholar] [CrossRef] [PubMed]
- Case, H.S.; Reed, H.L.; Palinkas, L.A.; Reedy, K.R.; Van Do, N.; Finney, N.S.; Seip, R. Resting and Exercise Energy Use in Antarctica: Effect of 50% Restriction in Temperate Climate Energy Requirements. Clin. Endocrinol. 2006, 65, 257–264. [Google Scholar] [CrossRef]
- Moreno-Reyes, R.; Carpentier, Y.A.; Macours, P.; Gulbis, B.; Corvilain, B.; Glinoer, D.; Goldman, S. Seasons but Not Ethnicity Influence Urinary Iodine Concentrations in Belgian Adults. Eur. J. Nutr. 2011, 50, 285–290. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.; Shea, B.; O’connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (Nos) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses; Review Manager (RevMan) [Computer Program]. Version 5.4; Ottawa Hospital Research Institute: Ottawa, ON, Canada; The Cochrane Collaboration: London, UK, 2020. [Google Scholar]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. A Basic Introduction to Fixed-Effect and Random-Effects Models for Meta-Analysis. Res. Synth. Method 2010, 1, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Li, T.; Deeks, J.J. Chapter 6: Choosing effect measures and computing estimates of effect. In Cochrane Handbook for Systematic Reviews of Interventions Version 6.3; Higgins, J.P.T., Thomas, J., Chandler, J., Cumpston, M., Li, T., Page, M.J., Welch, V.A., Eds.; Cochrane: London, UK, 2022. [Google Scholar]
- Leonard, W.R.; Levy, S.B.; Tarskaia, L.A.; Klimova, T.M.; Fedorova, V.I.; Baltakhinova, M.E.; Krivoshapkin, V.G.; Snodgrass, J.J. Seasonal Variation in Basal Metabolic Rates among the Yakut (Sakha) of Northeastern Siberia: Seasonality in BMR in the Yakut. Am. J. Hum. Biol. 2014, 26, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Reed, H.L.; Reedy, K.R.; Palinkas, L.A.; Van Do, N.; Finney, N.S.; Case, H.S.; LeMar, H.J.; Wright, J.; Thomas, J. Impairment in Cognitive and Exercise Performance during Prolonged Antarctic Residence: Effect of Thyroxine Supplementation in the Polar Triiodothyronine Syndrome. J. Clin. Endocrinol. Metab. 2001, 86, 110–116. [Google Scholar] [CrossRef]
- Palinkas, L.A.; Reed, H.L.; Reedy, K.R.; Do, N.V.; Case, H.S.; Finney, N.S. Circannual Pattern of Hypothalamic-Pituitary-Thyroid (HPT) Function and Mood during Extended Antarctic Residence. Psychoneuroendocrinology 2001, 26, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Nagata, H.; Izumiyama, T.; Kamata, K.; Kono, S.; Yukimura, Y. An Increase of Plasma Triiodothyronine Concentration in Man in a Cold Environment. J. Clin. Endocrinol. Metab. 1976, 43, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Do, N.V.; LeMar, H.; Reed, H.L. Thyroid hormone responses to environmental cold exposure and seasonal change: A proposed model. Endocrinol. Metab. 1996, 3, 7–16. [Google Scholar]
- Kovaničová, Z.; Kurdiová, T.; Baláž, M.; Štefanička, P.; Varga, L.; Kulterer, O.C.; Betz, M.J.; Haug, A.R.; Burger, I.A.; Kiefer, F.W.; et al. Cold Exposure Distinctively Modulates Parathyroid and Thyroid Hormones in Cold-Acclimatized and Non-Acclimatized Humans. Endocrinology 2020, 161, bqaa051. [Google Scholar] [CrossRef]
- Andersen, S.; Kleinschmidt, K.; Hvingel, B.; Laurberg, P. Thyroid Hyperactivity with High Thyroglobulin in Serum despite Sufficient Iodine Intake in Chronic Cold Adaptation in an Arctic Inuit Hunter Population. Eur. J. Endocrinol. 2012, 166, 433–440. [Google Scholar] [CrossRef]
- Freeman, M.K.; Adunlin, G.A.; Mercadel, C.; Danzi, S.; Klein, I. Hypothyroid Symptoms in Levothyroxine-Treated Patients. Innov. Pharm. 2019, 10, 19. [Google Scholar] [CrossRef]
- Ettleson, M.D.; Bianco, A.C. Individualized Therapy for Hypothyroidism: Is T4 Enough for Everyone? J. Clin. Endocrinol. Metab. 2020, 105, e3090–e3104. [Google Scholar] [CrossRef] [PubMed]
- Skarulis, M.C.; Celi, F.S.; Mueller, E.; Zemskova, M.; Malek, R.; Hugendubler, L.; Cochran, C.; Solomon, J.; Chen, C.; Gorden, P. Thyroid Hormone Induced Brown Adipose Tissue and Amelioration of Diabetes in a Patient with Extreme Insulin Resistance. J. Clin. Endocrinol. Metab. 2010, 95, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Broeders, E.P.M.; Vijgen, G.H.E.J.; Havekes, B.; Bouvy, N.D.; Mottaghy, F.M.; Kars, M.; Schaper, N.C.; Schrauwen, P.; Brans, B.; van Marken Lichtenbelt, W.D. Thyroid Hormone Activates Brown Adipose Tissue and Increases Non-Shivering Thermogenesis-A Cohort Study in a Group of Thyroid Carcinoma Patients. PLoS ONE 2016, 11, e0145049. [Google Scholar] [CrossRef] [PubMed]
- Maushart, C.I.; Loeliger, R.; Gashi, G.; Christ-Crain, M.; Betz, M.J. Resolution of Hypothyroidism Restores Cold-Induced Thermogenesis in Humans. Thyroid 2019, 29, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Triandafillou, J.; Gwilliam, C.; Himms-Hagen, J. Role of Thyroid Hormone in Cold-Induced Changes in Rat Brown Adipose Tissue Mitochondria. Can. J. Biochem. 1982, 60, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.E. Thermogenic Mechanisms and Their Hormonal Regulation. Physiol. Rev. 2006, 86, 435–464. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.E. Physiological Importance and Control of Non-Shivering Facultative Thermogenesis. Front. Biosci. 2011, 3, 352–371. [Google Scholar] [CrossRef]
- Sentis, S.C.; Oelkrug, R.; Mittag, J. Thyroid Hormones in the Regulation of Brown Adipose Tissue Thermogenesis. Endocr. Connect. 2021, 10, R106–R115. [Google Scholar] [CrossRef]
- Yau, W.W.; Singh, B.K.; Lesmana, R.; Zhou, J.; Sinha, R.A.; Wong, K.A.; Wu, Y.; Bay, B.-H.; Sugii, S.; Sun, L.; et al. Thyroid Hormone (T3) Stimulates Brown Adipose Tissue Activation via Mitochondrial Biogenesis and MTOR-Mediated Mitophagy. Autophagy 2018, 15, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.W.; Yen, P.M. Thermogenesis in Adipose Tissue Activated by Thyroid Hormone. Int. J. Mol. Sci. 2020, 21, 3020. [Google Scholar] [CrossRef]
- Nedergaard, J.; Bengtsson, T.; Cannon, B. Unexpected Evidence for Active Brown Adipose Tissue in Adult Humans. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E444–E452. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Okamatsu-Ogura, Y.; Matsushita, M.; Watanabe, K.; Yoneshiro, T.; Nio-Kobayashi, J.; Iwanaga, T.; Miyagawa, M.; Kameya, T.; Nakada, K.; et al. High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans. Diabetes 2009, 58, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.-J.; Enerbäck, S.; et al. Functional Brown Adipose Tissue in Healthy Adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Zingaretti, M.C.; Crosta, F.; Vitali, A.; Guerrieri, M.; Frontini, A.; Cannon, B.; Nedergaard, J.; Cinti, S. The Presence of UCP1 Demonstrates That Metabolically Active Adipose Tissue in the Neck of Adult Humans Truly Represents Brown Adipose Tissue. FASEB J. 2009, 23, 3113–3120. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.C.; Silva, J.E. Cold Exposure Rapidly Induces Virtual Saturation of Brown Adipose Tissue Nuclear T3 Receptors. Am. J. Physiol. 1988, 255, E496–E503. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.D.; Kimura, E.T.; Bianco, A.C.; Silva, J.E. Central Role of Brown Adipose Tissue Thyroxine 5’-Deiodinase on Thyroid Hormone-Dependent Thermogenic Response to Cold. Endocrinology 1991, 128, 2149–2159. [Google Scholar] [CrossRef]
- Rabelo, R.; Schifman, A.; Rubio, A.; Sheng, X.; Silva, J.E. Delineation of Thyroid Hormone-Responsive Sequences within a Critical Enhancer in the Rat Uncoupling Protein Gene. Endocrinology 1995, 136, 1003–1013. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Takahashi, N.; Yasubuchi, M.; Kim, Y.-I.; Hashizaki, H.; Kim, M.-J.; Sakamoto, T.; Goto, T.; Kawada, T. Triiodothyronine Induces UCP-1 Expression and Mitochondrial Biogenesis in Human Adipocytes. Am. J. Physiol. Cell Physiol. 2012, 302, C463–C472. [Google Scholar] [CrossRef]
- Laurberg, P.; Andersen, S.; Karmisholt, J. Cold Adaptation and Thyroid Hormone Metabolism. Horm. Metab. Res. 2005, 37, 545–549. [Google Scholar] [CrossRef]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef]
- Silva, J.E.; Larsen, P.R. Adrenergic Activation of Triiodothyronine Production in Brown Adipose Tissue. Nature 1983, 305, 712–713. [Google Scholar] [CrossRef]
- Ma, Y.; Shen, S.; Yan, Y.; Zhang, S.; Liu, S.; Tang, Z.; Yu, J.; Ma, M.; Niu, Z.; Li, Z.; et al. Adipocyte Thyroid Hormone β Receptor-Mediated Hormone Action Fine-Tunes Intracellular Glucose and Lipid Metabolism and Systemic Homeostasis. Diabetes 2023, 72, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Petito, G.; Cioffi, F.; Magnacca, N.; de Lange, P.; Senese, R.; Lanni, A. Adipose Tissue Remodeling in Obesity: An Overview of the Actions of Thyroid Hormones and Their Derivatives. Pharmaceuticals 2023, 16, 572. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J. The “Skinny” on Brown Fat, Obesity, and Bone. Am. J. Phys. Anthropol. 2015, 156 (Suppl. S59), 98–115. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Heeren, J. Adipose Tissue Browning and Metabolic Health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Roth, C.L.; Molica, F.; Kwak, B.R. Browning of White Adipose Tissue as a Therapeutic Tool in the Fight against Atherosclerosis. Metabolites 2021, 11, 319. [Google Scholar] [CrossRef]
- Dempersmier, J.; Sambeat, A.; Gulyaeva, O.; Paul, S.M.; Hudak, C.S.S.; Raposo, H.F.; Kwan, H.-Y.; Kang, C.; Wong, R.H.F.; Sul, H.S. Cold-Inducible Zfp516 Activates UCP1 Transcription to Promote Browning of White Fat and Development of Brown Fat. Mol. Cell 2015, 57, 235–246. [Google Scholar] [CrossRef]
- Wang, Q.A.; Tao, C.; Gupta, R.K.; Scherer, P.E. Tracking Adipogenesis during White Adipose Tissue Development, Expansion and Regeneration. Nat. Med. 2013, 19, 1338–1344. [Google Scholar] [CrossRef]
- Schosserer, M.; Grillari, J.; Wolfrum, C.; Scheideler, M. Age-Induced Changes in White, Brite, and Brown Adipose Depots: A Mini-Review. Gerontology 2018, 64, 229–236. [Google Scholar] [CrossRef]
- Zoico, E.; Rubele, S.; De Caro, A.; Nori, N.; Mazzali, G.; Fantin, F.; Rossi, A.; Zamboni, M. Brown and Beige Adipose Tissue and Aging. Front. Endocrinol. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed]
- Graja, A.; Gohlke, S.; Schulz, T.J. Aging of Brown and Beige/Brite Adipose Tissue. Handb. Exp. Pharmacol. 2019, 251, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Linderman, J.; Smith, S.; Brychta, R.J.; Wang, J.; Idelson, C.; Perron, R.M.; Werner, C.D.; Phan, G.Q.; Kammula, U.S.; et al. Irisin and FGF21 Are Cold-Induced Endocrine Activators of Brown Fat Function in Humans. Cell Metab. 2014, 19, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Frontini, A.; Cinti, S. Distribution and Development of Brown Adipocytes in the Murine and Human Adipose Organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Orava, J.; Nuutila, P.; Lidell, M.E.; Oikonen, V.; Noponen, T.; Viljanen, T.; Scheinin, M.; Taittonen, M.; Niemi, T.; Enerbäck, S.; et al. Different Metabolic Responses of Human Brown Adipose Tissue to Activation by Cold and Insulin. Cell Metab. 2011, 14, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Brychta, R.J.; Linderman, J.D.; Smith, S.; Courville, A.; Dieckmann, W.; Herscovitch, P.; Millo, C.M.; Remaley, A.; Lee, P.; et al. Brown Fat Activation Mediates Cold-Induced Thermogenesis in Adult Humans in Response to a Mild Decrease in Ambient Temperature. J. Clin. Endocrinol. Metab. 2013, 98, E1218–E1223. [Google Scholar] [CrossRef]
- Ikeda, K.; Yamada, T. UCP1 Dependent and Independent Thermogenesis in Brown and Beige Adipocytes. Front. Endocrinol. 2020, 11, 498. [Google Scholar] [CrossRef]
- Nirengi, S.; Stanford, K. Brown Adipose Tissue and Aging: A Potential Role for Exercise. Exp. Gerontol. 2023, 178, 112218. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, R.; Meng, Y.; Li, S.; Donelan, W.; Zhao, Y.; Qi, L.; Zhang, M.; Wang, X.; Cui, T.; et al. Irisin Stimulates Browning of White Adipocytes through Mitogen-Activated Protein Kinase P38 MAP Kinase and ERK MAP Kinase Signaling. Diabetes 2014, 63, 514–525. [Google Scholar] [CrossRef]
- Schneider, M.J.; Fiering, S.N.; Pallud, S.E.; Parlow, A.F.; St Germain, D.L.; Galton, V.A. Targeted Disruption of the Type 2 Selenodeiodinase Gene (DIO2) Results in a Phenotype of Pituitary Resistance to T4. Mol. Endocrinol. 2001, 15, 2137–2148. [Google Scholar] [CrossRef] [PubMed]
- Nikanorova, A.A.; Barashkov, N.A.; Pshennikova, V.G.; Nakhodkin, S.S.; Gotovtsev, N.N.; Romanov, G.P.; Solovyev, A.V.; Kuzmina, S.S.; Sazonov, N.N.; Fedorova, S.A. The Role of Nonshivering Thermogenesis Genes on Leptin Levels Regulation in Residents of the Coldest Region of Siberia. Int. J. Mol. Sci. 2021, 22, 4657. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E.; Murgatroyd, P.R.; Rothnie, I.; Reid, I.W.; Harvey, R. Metabolic and Thyroidal Responses to Mild Cold Are Abnormal in Obese Diabetic Women. Clin. Endocrinol. 1988, 28, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Savourey, G.; Caravel, J.P.; Barnavol, B.; Bittel, J.H. Thyroid Hormone Changes in a Cold Air Environment after Local Cold Acclimation. J. Appl. Physiol. 1994, 76, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Solter, M.; Brkic, K.; Petek, M.; Posavec, L.; Sekso, M. Thyroid Hormone Economy in Response to Extreme Cold Exposure in Healthy Factory Workers. J. Clin. Endocrinol. Metab. 1989, 68, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.M.; Bird, G.S.; Boag, L.M.; Delahaye, D.J.; Green, J.E.; Hatcher, J.D.; Page, J. Blood Volume and Basal Metabolic Rate of Eskimos. Metabolism 1954, 3, 247–254. [Google Scholar] [PubMed]
- Crile, G.W.; Quiring, D.P. Indian and Eskimo metabolism. J. Nutr. 1939, 18, 361–368. [Google Scholar] [CrossRef]
- Heinbecker, P. Studies on the metabolism of Eskimos. J. Biol. Chem. 1928, 80, 461–475. [Google Scholar] [CrossRef]
- Rabinowitch, I.M.; Smith, F.C. Metabolic studies of Eskimos in the Canadian Eastern Arctic. J. Nutr. 1936, 12, 337–356. [Google Scholar] [CrossRef]
- Roberts, D.F. Basal metabolism, race and climate. J. R. Anthropol. Inst. 1952, 82, 169–183. [Google Scholar] [CrossRef]
- Plasqui, G.; Kester, A.D.M.; Westerterp, K.R. Seasonal Variation in Sleeping Metabolic Rate, Thyroid Activity, and Leptin. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E338–E343. [Google Scholar] [CrossRef]
- Simpson, A. The Effect of Antarctic Residence on Energy Dynamics and Aerobic Fitness. Int. J. Circumpolar Health 2010, 69, 220–235. [Google Scholar] [CrossRef]
- Duncan, R. Variations in resting metabolic rate of men in Antarctica. Eur. J. Appl. Physiol. 1988, 57, 514–518. [Google Scholar] [CrossRef]
- Junghans, P.; Schrader, G.; Faust, H.; Wagner, B.; Hirschberg, K.; Reinhardt, R. Studies of the Protein and the Energy Metabolism in Man during a Wintering in Antarctica. Isot. Environ. Health Stud. 2012, 48, 208–225. [Google Scholar] [CrossRef] [PubMed]
- Verras, G.-I.; Mulita, F.; Pouwels, S.; Parmar, C.; Drakos, N.; Bouchagier, K.; Kaplanis, C.; Skroubis, G. Outcomes at 10-Year Follow-Up after Roux-En-Y Gastric Bypass, Biliopancreatic Diversion, and Sleeve Gastrectomy. J. Clin. Med. 2023, 12, 4973. [Google Scholar] [CrossRef] [PubMed]
- Haththotuwa, R.N.; Wijeyaratne, C.N.; Senarath, U. Chapter 1—Worldwide Epidemic of Obesity. In Obesity and Obstetrics, 2nd ed.; Mahmood, T.A., Arulkumaran, S., Chervenak, F.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 3–8. ISBN 978-0-12-817921-5. [Google Scholar]
- Genchi, V.A.; Palma, G.; Sorice, G.P.; D’Oria, R.; Caccioppoli, C.; Marrano, N.; Biondi, G.; Caruso, I.; Cignarelli, A.; Natalicchio, A.; et al. Pharmacological Modulation of Adaptive Thermogenesis: New Clues for Obesity Management? J. Endocrinol. Investig. 2023, 1–24. [Google Scholar] [CrossRef]
- Jagtap, U.; Paul, A. UCP1 Activation: Hottest Target in the Thermogenesis Pathway to Treat Obesity Using Molecules of Synthetic and Natural Origin. Drug Discov. Today 2023, 28, 103717. [Google Scholar] [CrossRef]
- Kang, Y.; Chen, L. Structural basis for the binding of DNP and purine nucleotides onto UCP1. Nature 2023, 620, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Muzik, O.; Mangner, T.J.; Leonard, W.R.; Kumar, A.; Granneman, J.G. Sympathetic Innervation of Cold-Activated Brown and White Fat in Lean Young Adults. J. Nucl. Med. 2017, 58, 799–806. [Google Scholar] [CrossRef]
- Paulo, E.; Wu, D.; Wang, Y.; Zhang, Y.; Wu, Y.; Swaney, D.L.; Soucheray, M.; Jimenez-Morales, D.; Chawla, A.; Krogan, N.J.; et al. Sympathetic Inputs Regulate Adaptive Thermogenesis in Brown Adipose Tissue through CAMP-Salt Inducible Kinase Axis. Sci. Rep. 2018, 8, 11001. [Google Scholar] [CrossRef]
- Wu, L.; Xia, M.; Duan, Y.; Zhang, L.; Jiang, H.; Hu, X.; Yan, H.; Zhang, Y.; Gu, Y.; Shi, H.; et al. Berberine Promotes the Recruitment and Activation of Brown Adipose Tissue in Mice and Humans. Cell Death Dis. 2019, 10, 468. [Google Scholar] [CrossRef]
- Hardie, D.G. AMPK: A Target for Drugs and Natural Products with Effects on Both Diabetes and Cancer. Diabetes 2013, 62, 2164–2172. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, L.; Fernández-Quintela, A.; Arias, N.; Portillo, M.P. Resveratrol: Anti-Obesity Mechanisms of Action. Molecules 2014, 19, 18632–18655. [Google Scholar] [CrossRef]
- Tauriainen, E.; Luostarinen, M.; Martonen, E.; Finckenberg, P.; Kovalainen, M.; Huotari, A.; Herzig, K.-H.; Lecklin, A.; Mervaala, E. Distinct Effects of Calorie Restriction and Resveratrol on Diet-Induced Obesity and Fatty Liver Formation. J. Nutr. Metab. 2011, 2011, 525094. [Google Scholar] [CrossRef]
- Takeda, Y.; Dai, P. Capsaicin Directly Promotes Adipocyte Browning in the Chemical Compound-Induced Brown Adipocytes Converted from Human Dermal Fibroblasts. Sci. Rep. 2022, 12, 6612. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A. Capsaicin for Weight Control: “Exercise in a Pill” (or Just Another Fad)? Pharmaceuticals 2022, 15, 851. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Yu, H.; Zhao, Z.; Luo, Z.; Chen, J.; Ni, Y.; Jin, R.; Ma, L.; Wang, P.; Zhu, Z.; et al. Activation of the Cold-Sensing TRPM8 Channel Triggers UCP1-Dependent Thermogenesis and Prevents Obesity. J. Mol. Cell Biol. 2012, 4, 88–96. [Google Scholar] [CrossRef]
- Pei, Y.; Otieno, D.; Gu, I.; Lee, S.-O.; Parks, J.S.; Schimmel, K.; Kang, H.W. Effect of Quercetin on Nonshivering Thermogenesis of Brown Adipose Tissue in High-Fat Diet-Induced Obese Mice. J. Nutr. Biochem. 2021, 88, 108532. [Google Scholar] [CrossRef]
- Zheng, Z.; Liu, X.; Zhao, Q.; Zhang, L.; Li, C.; Xue, Y. Regulation of UCP1 in the Browning of Epididymal Adipose Tissue by Β3-Adrenergic Agonist: A Role for MicroRNAs. Int. J. Endocrinol. 2014, 2014, 530636. [Google Scholar] [CrossRef]
- Liu, J.; Lin, L. Small Molecules for Fat Combustion: Targeting Thermosensory and Satiety Signals in the Central Nervous System. Drug Discov. Today 2019, 24, 300–306. [Google Scholar] [CrossRef]
- Riis-Vestergaard, M.J.; Richelsen, B.; Bruun, J.M.; Li, W.; Hansen, J.B.; Pedersen, S.B. Beta-1 and Not Beta-3 Adrenergic Receptors May Be the Primary Regulator of Human Brown Adipocyte Metabolism. J. Clin. Endocrinol. Metab. 2020, 105, dgz298. [Google Scholar] [CrossRef] [PubMed]
- Blondin, D.P.; Nielsen, S.; Kuipers, E.N.; Severinsen, M.C.; Jensen, V.H.; Miard, S.; Jespersen, N.Z.; Kooijman, S.; Boon, M.R.; Fortin, M.; et al. Human Brown Adipocyte Thermogenesis Is Driven by Β2-AR Stimulation. Cell Metab. 2020, 32, 287–300.e7. [Google Scholar] [CrossRef] [PubMed]
- O’Mara, A.E.; Johnson, J.W.; Linderman, J.D.; Brychta, R.J.; McGehee, S.; Fletcher, L.A.; Fink, Y.A.; Kapuria, D.; Cassimatis, T.M.; Kelsey, N.; et al. Chronic Mirabegron Treatment Increases Human Brown Fat, HDL Cholesterol, and Insulin Sensitivity. J. Clin. Investig. 2020, 130, 2209–2219. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.M.; Duntas, L.; Fadeyev, V.; Nygaard, B.; Vanderpump, M.P.J. 2012 ETA Guidelines: The Use of L-T4 + L-T3 in the Treatment of Hypothyroidism. Eur. Thyroid. J. 2012, 1, 55–71. [Google Scholar] [CrossRef]
- Kim, H.-L.; Jung, Y.; Park, J.; Youn, D.-H.; Kang, J.; Lim, S.; Lee, B.S.; Jeong, M.-Y.; Choe, S.-K.; Park, R.; et al. Farnesol Has an Anti-Obesity Effect in High-Fat Diet-Induced Obese Mice and Induces the Development of Beige Adipocytes in Human Adipose Tissue Derived-Mesenchymal Stem Cells. Front. Pharmacol. 2017, 8, 654. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Suh, J.M.; Reilly, S.M.; Yu, E.; Osborn, O.; Lackey, D.; Yoshihara, E.; Perino, A.; Jacinto, S.; Lukasheva, Y.; et al. Intestinal FXR Agonism Promotes Adipose Tissue Browning and Reduces Obesity and Insulin Resistance. Nat. Med. 2015, 21, 159–165. [Google Scholar] [CrossRef]
- Broeders, E.P.M.; Nascimento, E.B.M.; Havekes, B.; Brans, B.; Roumans, K.H.M.; Tailleux, A.; Schaart, G.; Kouach, M.; Charton, J.; Deprez, B.; et al. The Bile Acid Chenodeoxycholic Acid Increases Human Brown Adipose Tissue Activity. Cell Metab. 2015, 22, 418–426. [Google Scholar] [CrossRef]
- De Moura Souza, A.; Sichieri, R. Association between Serum TSH Concentration within the Normal Range and Adiposity. Eur. J. Endocrinol. 2011, 165, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Kitahara, C.M.; Platz, E.A.; Ladenson, P.W.; Mondul, A.M.; Menke, A.; Berrington de González, A. Body Fatness and Markers of Thyroid Function among U.S. Men and Women. PLoS ONE 2012, 7, e34979. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, N.; Laurberg, P.; Rasmussen, L.B.; Bülow, I.; Perrild, H.; Ovesen, L.; Jørgensen, T. Small Differences in Thyroid Function May Be Important for Body Mass Index and the Occurrence of Obesity in the Population. J. Clin. Endocrinol. Metab. 2005, 90, 4019–4024. [Google Scholar] [CrossRef]
- Bétry, C.; Challan-Belval, M.A.; Bernard, A.; Charrié, A.; Drai, J.; Laville, M.; Thivolet, C.; Disse, E. Increased TSH in Obesity: Evidence for a BMI-Independent Association with Leptin. Diabetes Metab. 2015, 41, 248–251. [Google Scholar] [CrossRef]
- Lambrinoudaki, I.; Armeni, E.; Rizos, D.; Georgiopoulos, G.; Athanasouli, F.; Triantafyllou, N.; Panoulis, K.; Augoulea, A.; Creatsa, M.; Alexandrou, A.; et al. Indices of Adiposity and Thyroid Hormones in Euthyroid Postmenopausal Women. Eur. J. Endocrinol. 2015, 173, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Asvold, B.O.; Bjøro, T.; Vatten, L.J. Association of Serum TSH with High Body Mass Differs between Smokers and Never-Smokers. J. Clin. Endocrinol. Metab. 2009, 94, 5023–5027. [Google Scholar] [CrossRef]
- Nyrnes, A.; Jorde, R.; Sundsfjord, J. Serum TSH Is Positively Associated with BMI. Int. J. Obes. 2006, 30, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Díez, J.J.; Iglesias, P. Relationship between Thyrotropin and Body Mass Index in Euthyroid Subjects. Exp. Clin. Endocrinol. Diabetes 2011, 119, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Ribaudo, M.C.; Zappaterreno, A.; Iannucci, C.V.; Leonetti, F. Relationship of Thyroid Function with Body Mass Index, Leptin, Insulin Sensitivity and Adiponectin in Euthyroid Obese Women. Clin. Endocrinol. 2005, 62, 487–491. [Google Scholar] [CrossRef]
- Rotondi, M.; Leporati, P.; La Manna, A.; Pirali, B.; Mondello, T.; Fonte, R.; Magri, F.; Chiovato, L. Raised Serum TSH Levels in Patients with Morbid Obesity: Is It Enough to Diagnose Subclinical Hypothyroidism? Eur. J. Endocrinol. 2009, 160, 403–408. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Sorice, G.P.; Mezza, T.; Prioletta, A.; Lassandro, A.P.; Pirronti, T.; Della Casa, S.; Pontecorvi, A.; Giaccari, A. High-Normal TSH Values in Obesity: Is It Insulin Resistance or Adipose Tissue’s Guilt? Obesity 2013, 21, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Du, F.-M.; Kuang, H.-Y.; Duan, B.-H.; Liu, D.-N.; Yu, X.-Y. Associations Between Thyroid Hormones Within the Euthyroid Range and Indices of Obesity in Obese Chinese Women of Reproductive Age. Metab. Syndr. Relat. Disord. 2019, 17, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Al Mohareb, O.; Al Saqaaby, M.; Ekhzaimy, A.; Hamza, M.; AlMalki, M.H.; Bamehriz, F.; Abukhater, M.; Brema, I. The Relationship between Thyroid Function and Body Composition, Leptin, Adiponectin, and Insulin Sensitivity in Morbidly Obese Euthyroid Subjects Compared to Non-Obese Subjects. Clin. Med. Insights Endocrinol. Diabetes 2021, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abdi, H.; Faam, B.; Gharibzadeh, S.; Mehran, L.; Tohidi, M.; Azizi, F.; Amouzegar, A. Determination of Age and Sex Specific TSH and FT4 Reference Limits in Overweight and Obese Individuals in an Iodine-Replete Region: Tehran Thyroid Study (TTS). Endocr. Res. 2021, 46, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Ren, R.; Jiang, X.; Zhang, X.; Guan, Q.; Yu, C.; Li, Y.; Gao, L.; Zhang, H.; Zhao, J. Association between Thyroid Hormones and Body Fat in Euthyroid Subjects. Clin. Endocrinol. 2014, 80, 585–590. [Google Scholar] [CrossRef] [PubMed]
- De Pergola, G.; Ciampolillo, A.; Paolotti, S.; Trerotoli, P.; Giorgino, R. Free Triiodothyronine and Thyroid Stimulating Hormone Are Directly Associated with Waist Circumference, Independently of Insulin Resistance, Metabolic Parameters and Blood Pressure in Overweight and Obese Women. Clin. Endocrinol. 2007, 67, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Shon, H.S.; Jung, E.D.; Kim, S.H.; Lee, J.H. Free T4 Is Negatively Correlated with Body Mass Index in Euthyroid Women. Korean J. Intern. Med. 2008, 23, 53–57. [Google Scholar] [CrossRef]
- Santini, F.; Marzullo, P.; Rotondi, M.; Ceccarini, G.; Pagano, L.; Ippolito, S.; Chiovato, L.; Biondi, B. Mechanisms in Endocrinology: The Crosstalk between Thyroid Gland and Adipose Tissue: Signal Integration in Health and Disease. Eur. J. Endocrinol. 2014, 171, R137–R152. [Google Scholar] [CrossRef] [PubMed]
- Mele, C.; Mai, S.; Cena, T.; Pagano, L.; Scacchi, M.; Biondi, B.; Aimaretti, G.; Marzullo, P. The Pattern of TSH and FT4 Levels across Different BMI Ranges in a Large Cohort of Euthyroid Patients with Obesity. Front. Endocrinol. 2022, 13, 1029376. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
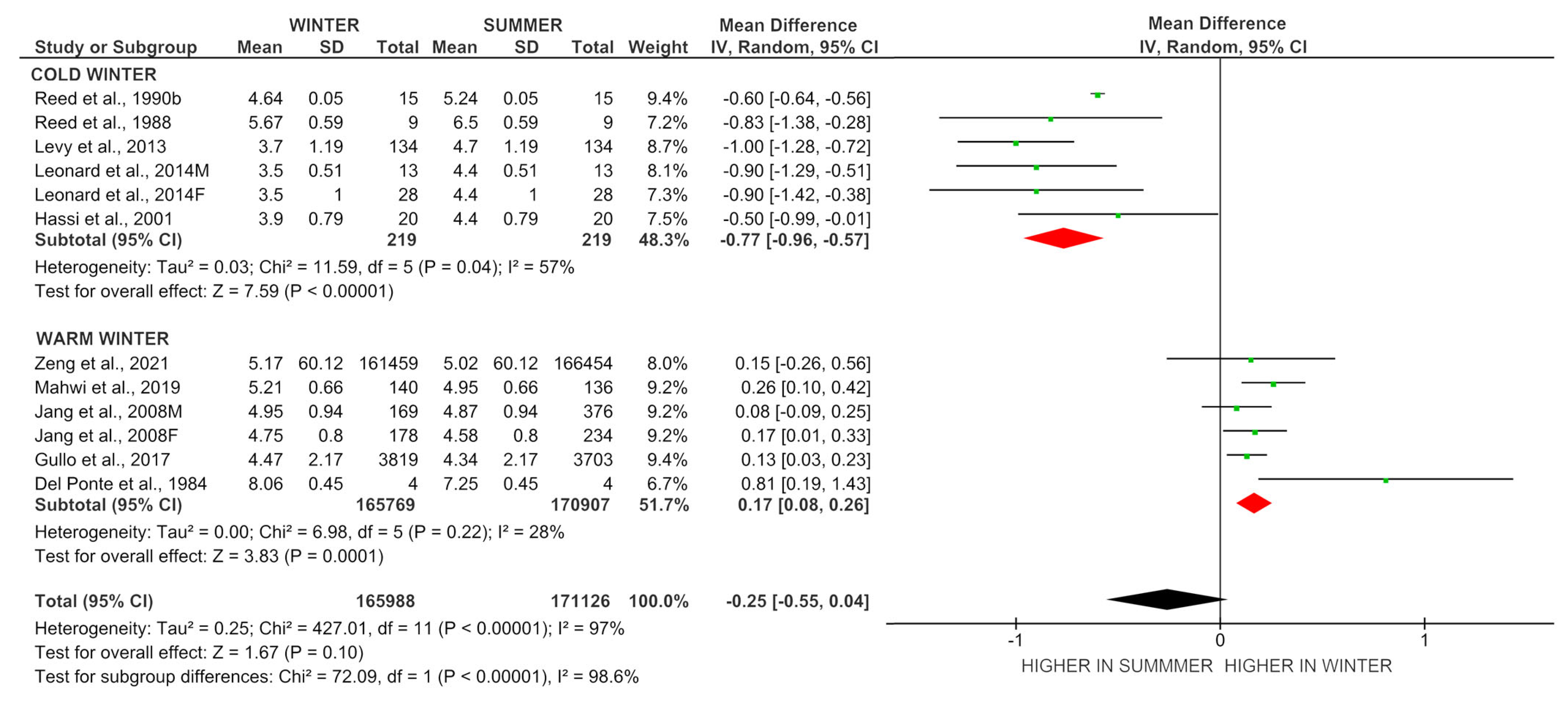
| # | Author, Year | Country | Region | Study Design | Sample Size | Age, Years | Sex | Temperature, °C | FT3, pmol/L | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Winter | Summer | Winter | Summer | |||||||||
| Cold Winter | ||||||||||||
| 1 | Reed et al., 1988 [23] | USA and Antarctica | Los Angeles and McMurdo Station | L | 9 | 25 ± 1.0 | M | −20 | +25 | 5.67 ± 0.51 | 6.5 ± 0.26 | |
| 2 | Reed et al., 1990b [22] | USA and Antarctica | Los Angeles and McMurdo Station | L | 15 | 24.5 ± 0.9 | M | −29 | +22 | 4.64 ± 0.14 | 5.24 ± 0.18 | |
| 3 | Hassi et al., 2001 [9] | Finland | province of Lapland | L | 20 | 26–40 | M | −10 | +15 | 3.9 ± 0.1 | 4.4 ± 0.2 | |
| 4 | Leonard et al., 2014 [36] | Russia | Republic of Sakha (Yakutia) | L | 13 | 18–49 | M | −34 | +18 | 3.5 ± 0.19 | 4.4 ± 0.14 | |
| Leonard et al., 2014 [36] | Russia | Republic of Sakha (Yakutia) | L | 28 | 18–49 | F | −34 | +18 | 3.5 ± 0.7 | 4.4 ± 0.7 | ||
| 5 | Levy et al., 2013 [11] | Russia | Republic of Sakha (Yakutia) | L | 134 | 19–49 | M/F | −34 | +18 | 3.7 ± 1.5 | 4.7 ± 1.1 | |
| Warm winter | ||||||||||||
| 6 | Mahwi et al., 2019 [26] | Iraq | Kurdistan Region, Sulaymaniyah | C | 140 | 34 ± 12 | M/F | +7 | +34 | 5.21 ± 0.05 | 4.95 ± 0.06 | |
| 7 | Zeng et al., 2021 [15] | China | Sichuan Province | C | 327913 | 18–90 | M/F | +5 | +33 | 5.17 ± 0.001 | 5.02 ± 0.002 | |
| 8 | Jang et al., 2008 [10] | South Korea | Gyeongsang Province | C | 545 | 18–65 | M | +7 | +28 | 4.95 ± 0.07 | 4.87 ± 0.04 | |
| Jang et al., 2008 [10] | South Korea | Gyeongsang Province | C | 412 | 18–65 | F | +7 | +28 | 4.75 ± 0.91 | 4.58 ± 0.77 | ||
| 9 | Del Ponte et al., 1984 [24] | Italy | Province of Chieti | L | 4 | 24–28 | M | +9 | +25 | 8.06 ± 0.38 | 7.25 ± 0.37 | |
| 10 | Gullo et al., 2017 [25] | Italy | City of Catania | C | 7522 | 37–61 | M/F | +10 | +25 | 4.47 ± 0.54 | 4.34 ± 0.52 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikanorova, A.A.; Barashkov, N.A.; Pshennikova, V.G.; Teryutin, F.M.; Nakhodkin, S.S.; Solovyev, A.V.; Romanov, G.P.; Burtseva, T.E.; Fedorova, S.A. A Systematic Review and Meta-Analysis of Free Triiodothyronine (FT3) Levels in Humans Depending on Seasonal Air Temperature Changes: Is the Variation in FT3 Levels Related to Nonshivering Thermogenesis? Int. J. Mol. Sci. 2023, 24, 14052. https://doi.org/10.3390/ijms241814052
Nikanorova AA, Barashkov NA, Pshennikova VG, Teryutin FM, Nakhodkin SS, Solovyev AV, Romanov GP, Burtseva TE, Fedorova SA. A Systematic Review and Meta-Analysis of Free Triiodothyronine (FT3) Levels in Humans Depending on Seasonal Air Temperature Changes: Is the Variation in FT3 Levels Related to Nonshivering Thermogenesis? International Journal of Molecular Sciences. 2023; 24(18):14052. https://doi.org/10.3390/ijms241814052
Chicago/Turabian StyleNikanorova, Alena A., Nikolay A. Barashkov, Vera G. Pshennikova, Fedor M. Teryutin, Sergey S. Nakhodkin, Aisen V. Solovyev, Georgii P. Romanov, Tatiana E. Burtseva, and Sardana A. Fedorova. 2023. "A Systematic Review and Meta-Analysis of Free Triiodothyronine (FT3) Levels in Humans Depending on Seasonal Air Temperature Changes: Is the Variation in FT3 Levels Related to Nonshivering Thermogenesis?" International Journal of Molecular Sciences 24, no. 18: 14052. https://doi.org/10.3390/ijms241814052
APA StyleNikanorova, A. A., Barashkov, N. A., Pshennikova, V. G., Teryutin, F. M., Nakhodkin, S. S., Solovyev, A. V., Romanov, G. P., Burtseva, T. E., & Fedorova, S. A. (2023). A Systematic Review and Meta-Analysis of Free Triiodothyronine (FT3) Levels in Humans Depending on Seasonal Air Temperature Changes: Is the Variation in FT3 Levels Related to Nonshivering Thermogenesis? International Journal of Molecular Sciences, 24(18), 14052. https://doi.org/10.3390/ijms241814052