
Early Pregnancy Markers in the Serum of Ewes Identified via Proteomic and Metabolomic Analyses
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Serum Collection and Pregnancy Diagnosis
2.2. Metabolomic Analysis of Serum
2.3. Differential Metabolites Detected between Pregnant and Non-Pregnant Ewes
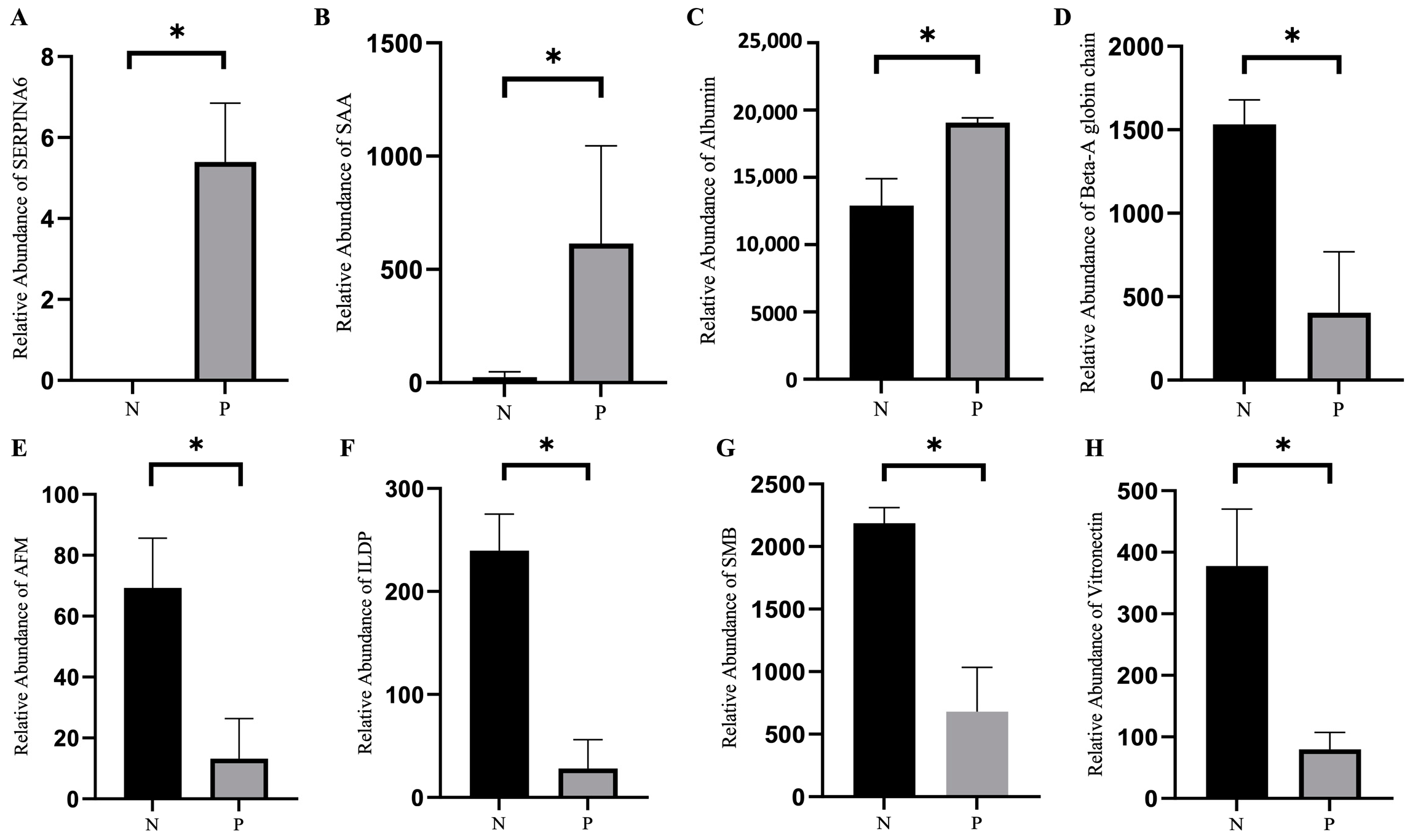
2.4. Proteomic Analysis of Serum
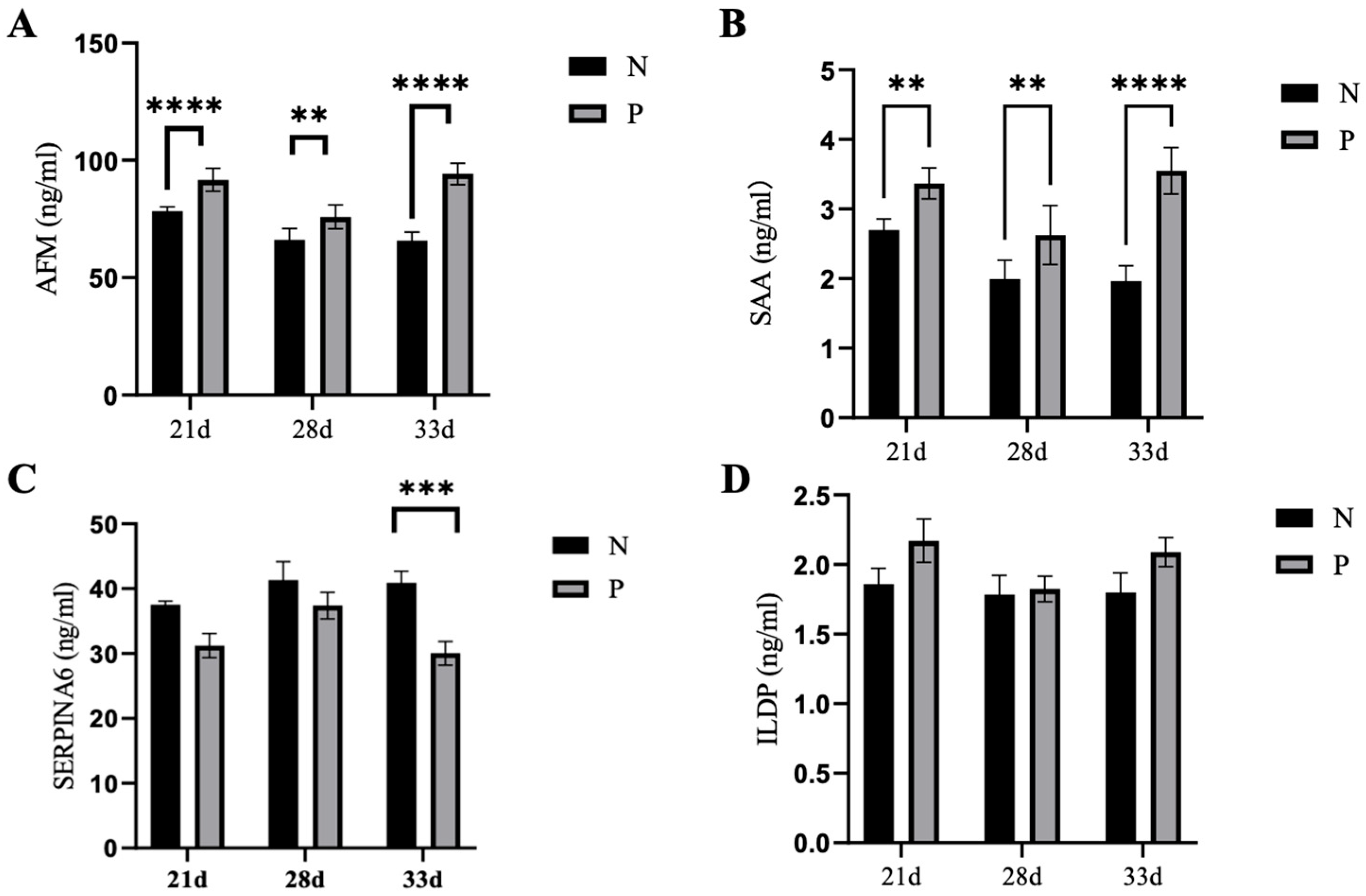
2.5. Six Levels of Serum Proteins in Pregnant and Non-Pregnant Ewes at Different Stages of Pregnancy as Determined using ELISA
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Collection of Blood Samples from Ewes
4.3. Serum Metabolomic Analysis
4.3.1. Serum Samples Selection and Quality Control
4.3.2. LC-MS/MS Analysis of the Serum Samples
4.3.3. Data Preprocessing and Annotation
4.3.4. Differential Metabolites Analysis
4.4. Serum Proteomic Analysis
4.4.1. Serum Samples Selection and Quality Control
4.4.2. Proteolytic Hydrolysis and Peptide Quantification
4.4.3. DIA Mass Detection of a Single Sample
4.4.4. Protein Identification
4.4.5. Statistical and Bioinformatics Analysis
4.5. Evaluation of Protein Levels in Sera of Ewes at Different Time Points
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Jones, A.K.; Gately, R.E.; McFadden, K.K.; Zinn, S.A.; Govoni, K.E.; Reed, S.A. Transabdominal ultrasound for detection of pregnancy, fetal and placental landmarks, and fetal age before Day 45 of gestation in the sheep. Theriogenology 2016, 85, 939−945.e1. [Google Scholar] [CrossRef] [PubMed]
- Andueza, D.; Alabart, J.L.; Lahoz, B.; Muñoz, F.; Folch, J. Early pregnancy diagnosis in sheep using near-infrared spectroscopy on blood plasma. Theriogenology 2014, 81, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Petrujkić, B.T.; Cojkić, A.; Petrujkić, K.; Jeremić, I.; Mašulović, D.; Dimitrijević, V.; Savić, M.; Pešić, M.; Beier, R.C. Transabdominal and transrectal ultrasonography of fetuses in Württemberg ewes: Correlation with gestational age. Anim. Sci. J. 2016, 87, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Hamilton, W.C.; Sasser, R.G.; Ruder, C.A.; Hass, G.M.; Williams, R.J. Detection and partial characterization of two bovine pregnancy-specific proteins. Biol. Reprod. 1982, 26, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Zoli, A.P.; Beckers, J.F.; Wouters-Ballman, P.; Closset, J.; Falmagne, P.; Ectors, F. Purification and characterization of a bovine pregnancy-associated glycoprotein. Biol. Reprod. 1991, 45, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N.M.; Ayad, A.; Beckers, J.F.; Gajewski, Z. Pregnancy-associated glycoproteins (PAG) as pregnancy markers in the ruminants. J. Physiol. Pharmacol. 2006, 57 (Suppl. S8), 153–171. [Google Scholar] [PubMed]
- Friedrich, M.; Holtz, W. Establishment of an ELISA for measuring bovine pregnancy-associated glycoprotein in serum or milk and its application for early pregnancy detection. Reprod. Domest. Anim. 2010, 45, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Steckeler, P.; Weber, F.; Zerbe, H.; Rieger, A.; Voigt, K. Evaluation of a bovine visual pregnancy test for the detection of pregnancy-associated glycoproteins in sheep. Reprod. Domest. Anim. 2019, 54, 280–288. [Google Scholar] [CrossRef]
- Karen, A.; Amiri, B.E.; Beckers, J.F.; Sulon, J.; Taverne, M.A.; Szenci, O. Comparison of accuracy of transabdominal ultrasonography, progesterone and pregnancy-associated glycoproteins tests for discrimination between single and multiple pregnancy in sheep. Theriogenology 2006, 66, 314–322. [Google Scholar] [CrossRef]
- Barbato, O.; Sousa, N.M.; Debenedetti, A.; Canali, C.; Todini, L.; Beckers, J.F. Validation of a new pregnancy-associated glycoprotein radioimmunoassay method for the detection of early pregnancy in ewes. Theriogenology 2009, 72, 993–1000. [Google Scholar] [CrossRef]
- Bianchi, S.; Nottola, S.A.; Torge, D.; Palmerini, M.G.; Necozione, S.; Macchiarelli, G. Association between Female Reproductive Health and Mancozeb: Systematic Review of Experimental Models. Int. J. Environ. Res. Public Health 2020, 17, 2580. [Google Scholar] [CrossRef] [PubMed]
- Domon, B.; Aebersold, R. Mass spectrometry and protein analysis. Science 2006, 312, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Lippolis, J.D.; Reinhardt, T.A. Centennial paper: Proteomics in animal science. J. Anim. Sci. 2008, 86, 2430–2441. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Heyman, H.M. Mass Spectrometry-Based Metabolomics. Methods Mol. Biol. 2018, 1775, 107–118. [Google Scholar] [PubMed]
- Horgan, R.P.; Clancy, O.H.; Myers, J.E.; Baker, P.N. An overview of proteomic and metabolomic technologies and their application to pregnancy research. BJOG 2009, 116, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2021, 53, e13711. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Ye, N.; Zhao, S.; Wang, G.; Zhang, Y.; Wang, T.; Zou, P.; Wang, J.; Yao, T.; Chen, M.; et al. Studies on Novel Diagnostic and Predictive Biomarkers of Intrahepatic Cholestasis of Pregnancy through Metabolomics and Proteomics. Front. Immunol. 2021, 12, 733225. [Google Scholar] [CrossRef]
- Xin, H.; Meng, F.; Li, B.; Wang, S.; Hu, B.; Li, J. Global proteomic analysis of serum during early pregnancy in the pig using data-independent acquisition mass spectrometry with verification by parallel reaction monitoring. Reprod. Fertil. Dev. 2022, 34, 1115–1127. [Google Scholar] [CrossRef]
- De, A.; Ali, M.A.; Chutia, T.; Onteru, S.K.; Behera, P.; Kalita, G.; Kumar, S.; Gali, J.M. Comparative serum proteome analysis reveals potential early pregnancy-specific protein biomarkers in pigs. Reprod. Fertil. Dev. 2019, 31, 613–631. [Google Scholar] [CrossRef]
- Yang, P.; Chen, X.; Tian, X.; Zhou, Z.; Zhang, Y.; Tang, W.; Fu, K.; Zhao, J.; Ruan, Y. A Proteomic Study of the Effect of N-acetylcysteine on the Regulation of Early Pregnancy in Goats. Animals 2022, 12, 2439. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, J.Y.; Kim, H.R.; Shin, H.Y.; Lin, T.; Jin, D.I. Proteomic Analysis of Bovine Pregnancy-specific Serum Proteins by 2D Fluorescence Difference Gel Electrophoresis. Asian-Australas. J. Anim. Sci. 2015, 28, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Panda, B.S.K.; Mohapatra, S.K.; Chaudhary, D.; Alhussien, M.N.; Kapila, R.; Dang, A.K. Proteomics and transcriptomics study reveals the utility of ISGs as novel molecules for early pregnancy diagnosis in dairy cows. J. Reprod. Immunol. 2020, 140, 103148. [Google Scholar] [CrossRef] [PubMed]
- Ceciliani, F.; Lecchi, C.; Urh, C.; Sauerwein, H. Proteomics and metabolomics characterizing the pathophysiology of adaptive reactions to the metabolic challenges during the transition from late pregnancy to early lactation in dairy cows. J. Proteom. 2018, 178, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Han, Y.; Tang, C.; Liao, Q.; Li, Z. Label-Free Mass Spectrometry-Based Quantitative Proteomics Analysis of Serum Proteins during Early Pregnancy in Jennies (Equus asinus). Front. Vet. Sci. 2020, 7, 569587. [Google Scholar] [CrossRef] [PubMed]
- Gerton, G.L.; Fan, X.J.; Chittams, J.; Sammel, M.; Hummel, A.; Strauss, J.F.; Barnhart, K. A serum proteomics approach to the diagnosis of ectopic pregnancy. Ann. N. Y. Acad. Sci. 2004, 1022, 306–316. [Google Scholar] [CrossRef]
- Nie, L.; Xin, S.; Zheng, J.; Luo, Y.; Zou, Y.; Liu, X.; Chen, H.; Lei, X.; Zeng, X.; Lai, H. DIA-based proteomics analysis of serum-derived exosomal proteins as potential candidate biomarkers for intrahepatic cholestasis in pregnancy. Arch. Gynecol. Obstet. 2023, 308, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Jansen, B.C.; Bondt, A.; Reiding, K.R.; Lonardi, E.; de Jong, C.J.; Falck, D.; Kammeijer, G.S.; Dolhain, R.J.; Rombouts, Y.; Wuhrer, M. Pregnancy-associated serum N-glycome changes studied by high-throughput MALDI-TOF-MS. Sci. Rep. 2016, 6, 23296. [Google Scholar] [CrossRef] [PubMed]
- Zeng, S.; Han, M.; Jiang, M.; Liu, F.; Hu, Y.; Long, Y.; Zhu, C.; Zeng, F.; Gan, Q.; Ye, W.; et al. Serum complement proteomics reveal biomarkers for hypertension disorder of pregnancy and the potential role of Clustering. Reprod. Biol. Endocrinol. 2021, 19, 56. [Google Scholar] [CrossRef]
- Vora, N.; Kalagiri, R.; Mallett, L.H.; Oh, J.H.; Wajid, U.; Munir, S.; Colon, N.; Raju, V.N.; Beeram, M.R.; Uddin, M.N. Proteomics and Metabolomics in Pregnancy-An Overview. Obstet. Gynecol. Surv. 2019, 74, 111–125. [Google Scholar] [CrossRef]
- Chen, M.; Guan, Y.; Huang, R.; Duan, J.; Zhou, J.; Chen, T.; Wang, X.; Xia, Y.; London, S.J. Associations between the Maternal Exposome and Metabolome during Pregnancy. Environ. Health Perspect. 2022, 130, 37003. [Google Scholar] [CrossRef]
- Alesi, S.; Ghelani, D.; Rassie, K.; Mousa, A. Metabolomic Biomarkers in Gestational Diabetes Mellitus: A Review of the Evidence. Int. J. Mol. Sci. 2021, 22, 5512. [Google Scholar] [CrossRef] [PubMed]
- Goldansaz, S.A.; Markus, S.; Plastow, G.; Wishart, D.S. Predictive blood biomarkers of sheep pregnancy and litter size. Sci. Rep. 2022, 12, 10307. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, C.; Wang, L.; Feng, R.; Guo, Y.; Feng, S.; Zhang, L.; Zheng, Z.; Su, G.; Fan, L.; et al. Correlation analysis of serum reproductive hormones and metabolites during multiple ovulation in sheep. BMC Vet. Res. 2022, 18, 290. [Google Scholar] [CrossRef]
- Chiaradia, E.; Avellini, L.; Tartaglia, M.; Gaiti, A.; Just, I.; Scoppetta, F.; Czentnar, Z.; Pich, A. Proteomic evaluation of sheep serum proteins. BMC Vet. Res. 2012, 8, 66. [Google Scholar] [CrossRef] [PubMed]
- de Miranda E Silva Chaves, C.; Dias da Costa, R.L.; Roncato Duarte, K.M.; Machado, D.C.; Paro de Paz, C.C.; Beltrame, R.T. Visual ELISA for detection of pregnancy-associated glycoproteins (PAGs) in ewe serum. Theriogenology 2017, 97, 78–82. [Google Scholar] [CrossRef] [PubMed]
- de Miranda E Silva Chaves, C.; da Costa, R.L.D.; Duarte, K.M.R.; Beltrame, R.T.; Quirino, C.R. Evaluation of a cattle rapid test for early pregnancy diagnosis in sheep. Trop. Anim. Health Prod. 2020, 52, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Mauffré, V.; Grimard, B.; Eozenou, C.; Inghels, S.; Silva, L.; Giraud-Delville, C.; Capo, D.; Sandra, O.; Constant, F. Interferon stimulated genes as peripheral diagnostic markers of early pregnancy in sheep: A critical assessment. Animal 2016, 66, 1856–1863. [Google Scholar] [CrossRef] [PubMed]
- Milisits-Németh, T.; Balogh, O.G.; Egerszegi, I.; Kern, L.; Sasser, R.G.; Gábor, G. Detection of pregnancy in sheep using an ELISA for pregnancy-specific protein B. Acta Vet. Hung. 2018, 66, 329–336. [Google Scholar] [CrossRef]
- Entrican, G.; Wheelhouse, N.M. Immunity in the female sheep reproductive tract. Vet. Res. 2006, 37, 295–309. [Google Scholar] [CrossRef]
- Canisso, I.F.; Ball, B.A.; Cray, C.; Williams, N.M.; Scoggin, K.E.; Davolli, G.M.; Squires, E.L.; Troedsson, M.H. Serum amyloid A and haptoglobin concentrations are increased in plasma of mares with ascending placentitis in the absence of changes in peripheral leukocyte counts or fibrinogen concentration. Am. J. Reprod. Immunol. 2014, 72, 376–385. [Google Scholar] [CrossRef]
- Chiriac, I.E.; Vilceanu, N.; Maghiar, A.; Andrei, C.; Hanganu, B.; Daina, L.G.; Dunarintu, S.; Buhas, L.C. Maternal Serum Amyloid A as a Marker of Preterm Birth/PROM: A Systematic Review and Meta-Analysis. Medicina 2023, 59, 1025. [Google Scholar] [CrossRef] [PubMed]
- Hubalek, M.; Buchner, H.; Mörtl, M.G.; Schlembach, D.; Huppertz, B.; Firulovic, B.; Köhler, W.; Hafner, E.; Dieplinger, B.; Wildt, L.; et al. The vitamin E-binding protein afamin increases in maternal serum during pregnancy. Clin. Chim. Acta 2014, 434, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Köninger, A.; Enekwe, A.; Mach, P.; Andrikos, D.; Schmidt, B.; Frank, M.; Birdir, C.; Kimmig, R.; Gellhaus, A.; Dieplinger, H. Afamin: An early predictor of preeclampsia. Arch. Gynecol. Obstet. 2018, 298, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Lichenstein, H.S.; Lyons, D.E.; Wurfel, M.M.; Johnson, D.A.; McGinley, M.D.; Leidli, J.C.; Trollinger, D.B.; Mayer, J.P.; Wright, S.D.; Zukowski, M.M. Afamin is a new member of the albumin, alpha-fetoprotein, and vitamin D-binding protein gene family. J. Biol. Chem. 1994, 269, 18149–18154. [Google Scholar] [CrossRef] [PubMed]
- Tramontana, A.; Dieplinger, B.; Stangl, G.; Hafner, E.; Dieplinger, H. First trimester serum afamin concentrations are associated with the development of pre-eclampsia and gestational diabetes mellitus in pregnant women. Clin. Chim. Acta 2018, 476, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Dieplinger, H.; Dieplinger, B. Afamin—A pleiotropic glycoprotein involved in various disease states. Clin. Chim. Acta 2015, 446, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Utge, S.; Räikkönen, K.; Kajantie, E.; Lipsanen, J.; Andersson, S.; Strandberg, T.; Reynolds, R.M.; Eriksson, J.G.; Lahti, J. Polygenic risk score of SERPINA6/SERPINA1 associates with diurnal and stress-induced HPA axis activity in children. Psychoneuroendocrinology 2018, 93, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vashchenko, G.; Das, S.; Moon, K.M.; Rogalski, J.C.; Taves, M.D.; Soma, K.K.; Van Petegem, F.; Foster, L.J.; Hammond, G.L. Identification of Avian Corticosteroid-binding Globulin (SerpinA6) Reveals the Molecular Basis of Evolutionary Adaptations in SerpinA6 Structure and Function as a Steroid-binding Protein. J. Biol. Chem. 2016, 291, 11300–11312. [Google Scholar] [CrossRef]
- Anderson, L.N.; Briollais, L.; Atkinson, H.C.; Marsh, J.A.; Xu, J.; Connor, K.L.; Matthews, S.G.; Pennell, C.E.; Lye, S.J. Investigation of genetic variants, birthweight and hypothalamic-pituitary-adrenal axis function suggests a genetic variant in the SERPINA6 gene is associated with corticosteroid binding globulin in the western Australia pregnancy cohort (Raine) study. PLoS ONE 2014, 9, e92957. [Google Scholar] [CrossRef]
- Lin, H.Y.; Underhill, C.; Lei, J.H.; Helander-Claesson, A.; Lee, H.Y.; Gardill, B.R.; Muller, Y.A.; Wang, H.; Hammond, G.L. High frequency of SERPINA6 polymorphisms that reduce plasma corticosteroid-binding globulin activity in Chinese subjects. J. Clin. Endocrinol. Metab. 2012, 97, 678–686. [Google Scholar] [CrossRef]
- Kong, X.; Guo, Z.; Yao, Y.; Xia, L.; Liu, R.; Song, H.; Zhang, S. Acetic acid alters rhizosphere microbes and metabolic composition to improve willows drought resistance. Sci. Total Environ. 2022, 844, 157132. [Google Scholar] [CrossRef]
- Li, C.; Al-Dalali, S.; Zhou, H.; Xu, B. Influence of curing on the metabolite profile of water-boiled salted duck. Food. Chem. 2022, 397, 133752. [Google Scholar] [CrossRef]
- Graves, P.R.; Haystead, T.A. Molecular biologist’s guide to proteomics. Microbiol. Mol. Biol. Rev. 2002, 66, 39–63. [Google Scholar] [CrossRef]
- Mun, D.G.; Vanderboom, P.M.; Madugundu, A.K.; Garapati, K.; Chavan, S.; Peterson, J.A.; Saraswat, M.; Pandey, A. DIA-Based Proteome Profiling of Nasopharyngeal Swabs from COVID-19 Patients. J. Proteome Res. 2021, 20, 4165–4175. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Regulate | p-Value | FC (P/N) |
|---|---|---|---|
| Tiglic acid | up | 0.008503 | 1.0083 |
| 4-Acetamido-2-aminobutanoic acid | up | 0.0003752 | 1.0192 |
| (3R,4R)-3-Amino-1-hydroxy-4-methylpyrrolidin-2-one | up | 0.03014 | 1.0213 |
| Glycylglycylglycine | up | 0.02539 | 1.0668 |
| N4-Acetylcytidine | up | 0.02476 | 1.0778 |
| 3,4-Dimethyl-5-pentyl-2-furanpropanoic acid | up | 0.04541 | 1.0456 |
| DL-2-Aminooctanoic acid | up | 0.01569 | 1.0753 |
| Capsidiol | up | 0.01163 | 1.0407 |
| Polidocanol | down | 0.02161 | 0.9714 |
| 4,5-Dihydrovomifoliol | up | 0.04924 | 1.0791 |
| PC(18:4(6Z,9Z,12Z,15Z)/20:2(11Z,14Z)) | up | 0.01659 | 1.01 |
| 1-Octen-3-yl glucoside | down | 0.01561 | 0.9823 |
| Indolophenanthridine | down | 0.04359 | 0.8634 |
| D-Cathine | up | 0.006031 | 1.0305 |
| Imexon | up | 0.03142 | 1.0665 |
| 2-Propylglutaric acid | up | 0.01513 | 1.015 |
| 1-Oleoylglycerophosphoserine | down | 0.01433 | 0.9535 |
| Thromboxane | down | 0.01514 | 0.9073 |
| (2R,3S)-2-Octyl-3-pentyloxane | down | 0.0355 | 0.9217 |
| PE-NMe(22:2(13Z,16Z)/16:1(9Z)) | up | 0.03717 | 1.0413 |
| Dodecyl hydrogen sulfate | down | 0.04624 | 0.9879 |
| PS(15:0/24:1(15Z)) | up | 0.01613 | 1.0313 |
| (+)-gamma-Hydroxy-L-homoarginine | up | 0.0239 | 1.0124 |
| 2-Mercaptobenzothiazole | up | 0.04465 | 1.1744 |
| Ecgonine | up | 0.03409 | 1.1418 |
| MJDBISSN00000004 | up | 0.04705 | 1.0085 |
| Cyclopentenyl cytosine | up | 0.02362 | 1.0617 |
| 3-Methyl-2-oxovaleric acid | down | 0.03653 | 0.9819 |
| D-Galactose | down | 0.02608 | 0.9901 |
| Accession Number | Description | Relative Abundance (P) | Relative Abundance (N) | FC (P/N) | p-Value (P/N) |
|---|---|---|---|---|---|
| A0A835ZZL2 | SERPIN domain-containing protein | 2825.4179 | 3124.8457 | 0.9042 | 0.04336 |
| A0A452ELD4 | SMB domain-containing protein | 679.4033 | 2186.5892 | 0.3107 | 0.01607 |
| A0A452DX18 | Complement C3 | 864.3877 | 1408.2509 | 0.6138 | 0.03305 |
| Q1KYZ7 | Beta-A globin chain β | 404.1686 | 1532.0683 | 0.2638 | 0.04558 |
| W5Q9D5 | Vitronectin | 79.6122 | 377.7781 | 0.2107 | 0.03671 |
| A0A6P3YFZ8 | Uncharacterized protein | 132.6988 | 176.5994 | 0.7514 | 0.004184 |
| W5NYF4 | Peptidoglycan recognition protein 2 | 56.3539 | 89.8142 | 0.6274 | 0.01077 |
| A0A452FXE6 | Insulin-like growth factor-binding protein 3 | 51.4896 | 66.5762 | 0.7734 | 0.01531 |
| A0A452F0G8 | Leukotriene A (4) hydrolase | 9.4593 | 16.1675 | 0.5851 | 0.04163 |
| A0A452FW61 | Ig-like domain-containing protein | 0 | 239.5522 | 1.00 × 10−5 | 0 |
| W5PXI3 | Afamin | 0 | 69.3152 | 1.00 × 10−5 | 0 |
| W5PWE9 | Albumin | 19,073.4655 | 12,902.1035 | 1.478 | 0.03812 |
| A0A835ZMI7 | Serum amyloid A protein | 614.5201 | 0 | 424.2 | 0 |
| A0A8C2R1B8 | Serpin family A member 66 | 5.3973 | 0 | 424.2 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, Y.; Xia, F.; Shi, L.; Ma, W.; Lv, X.; Sun, W.; Ji, P.; Gao, S.; Machaty, Z.; Liu, G.; et al. Early Pregnancy Markers in the Serum of Ewes Identified via Proteomic and Metabolomic Analyses. Int. J. Mol. Sci. 2023, 24, 14054. https://doi.org/10.3390/ijms241814054
Zhai Y, Xia F, Shi L, Ma W, Lv X, Sun W, Ji P, Gao S, Machaty Z, Liu G, et al. Early Pregnancy Markers in the Serum of Ewes Identified via Proteomic and Metabolomic Analyses. International Journal of Molecular Sciences. 2023; 24(18):14054. https://doi.org/10.3390/ijms241814054
Chicago/Turabian StyleZhai, Yaying, Fan Xia, Luting Shi, Wenkui Ma, Xiaoyang Lv, Wei Sun, Pengyun Ji, Shuai Gao, Zoltan Machaty, Guoshi Liu, and et al. 2023. "Early Pregnancy Markers in the Serum of Ewes Identified via Proteomic and Metabolomic Analyses" International Journal of Molecular Sciences 24, no. 18: 14054. https://doi.org/10.3390/ijms241814054
APA StyleZhai, Y., Xia, F., Shi, L., Ma, W., Lv, X., Sun, W., Ji, P., Gao, S., Machaty, Z., Liu, G., & Zhang, L. (2023). Early Pregnancy Markers in the Serum of Ewes Identified via Proteomic and Metabolomic Analyses. International Journal of Molecular Sciences, 24(18), 14054. https://doi.org/10.3390/ijms241814054