
Persistence of Coronavirus on Surface Materials and Its Control Measures Using Nonthermal Plasma and Other Agents


 ,
,  ,
, 
Abstract
:1. Introduction
1.1. Virus Transmission through Fomites
1.1.1. Porosity
1.1.2. Adsorption
1.1.3. Evaporation
1.1.4. Isoelectric Point
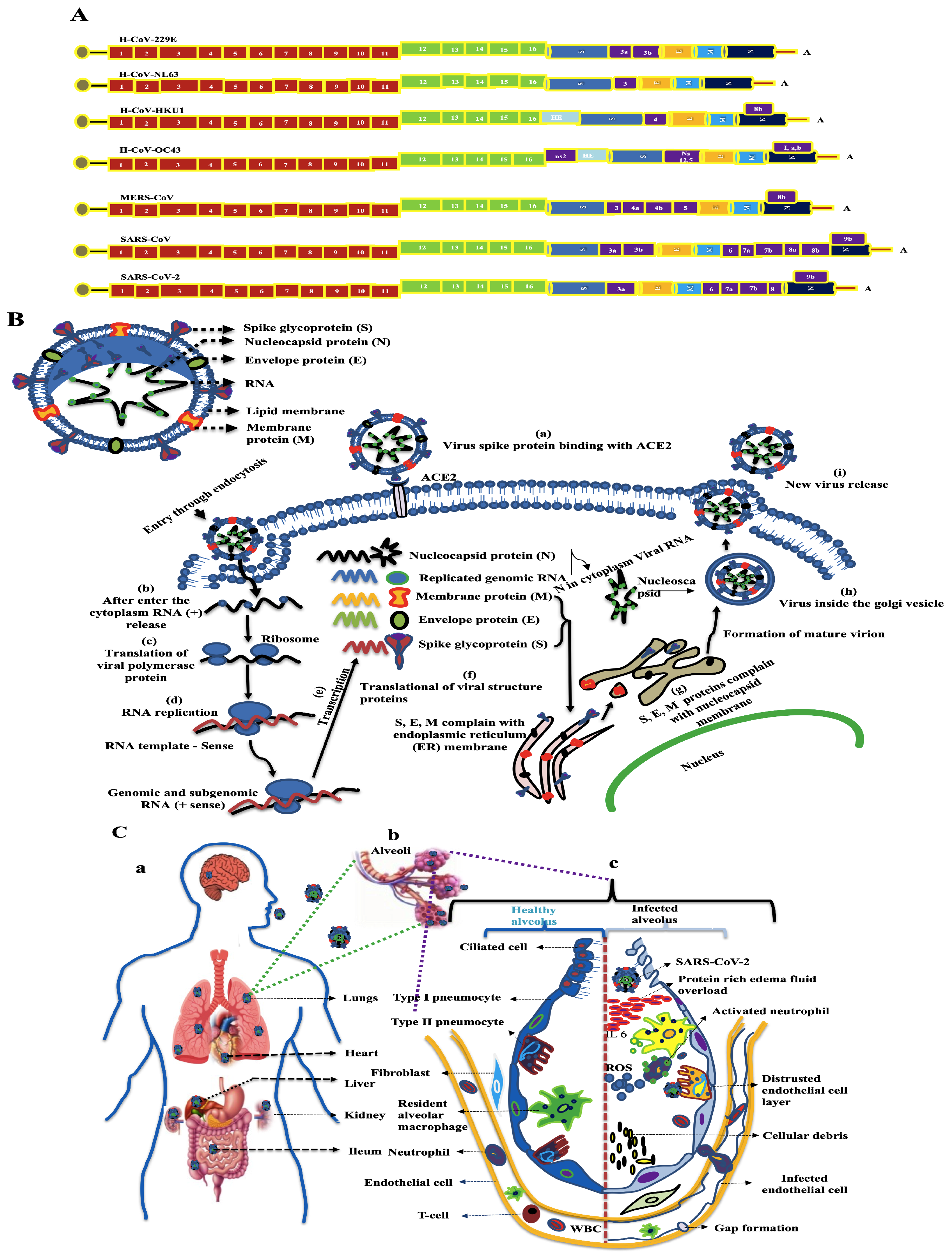
2. Coronavirus in Environment
2.1. Coronavirus-Contaminated Surface Materials
2.1.1. Porous Materials
Paper
Wood
Mask
2.1.2. Non-Porous Materials
Plastic
Glass
Stainless Steel
Copper
2.2. Environmental Factors Involved in Coronavirus Inactivity
2.2.1. Role of Temperature
2.2.2. Role of pH
2.2.3. Role of Humidity
2.3. Virus Inactivation in Chemical and Physical Treatment
2.3.1. Effective Chemicals Control Coronaviruses
Alcohols
Surfactants
Hypochlorite
Peroxide
2.3.2. Physical Methods
Ultraviolet (UV)
Gamma Radiation
X-rays
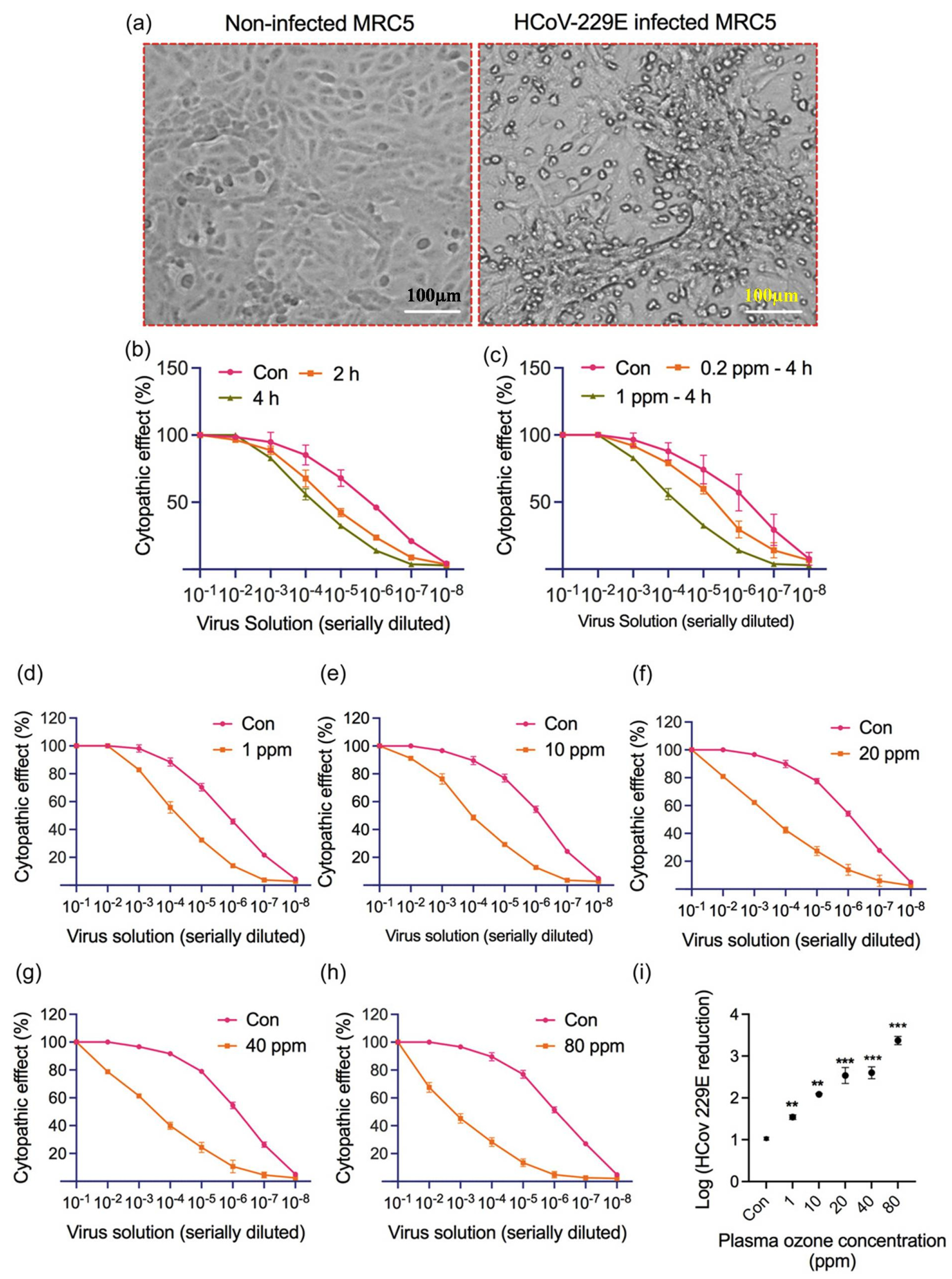
Ozone
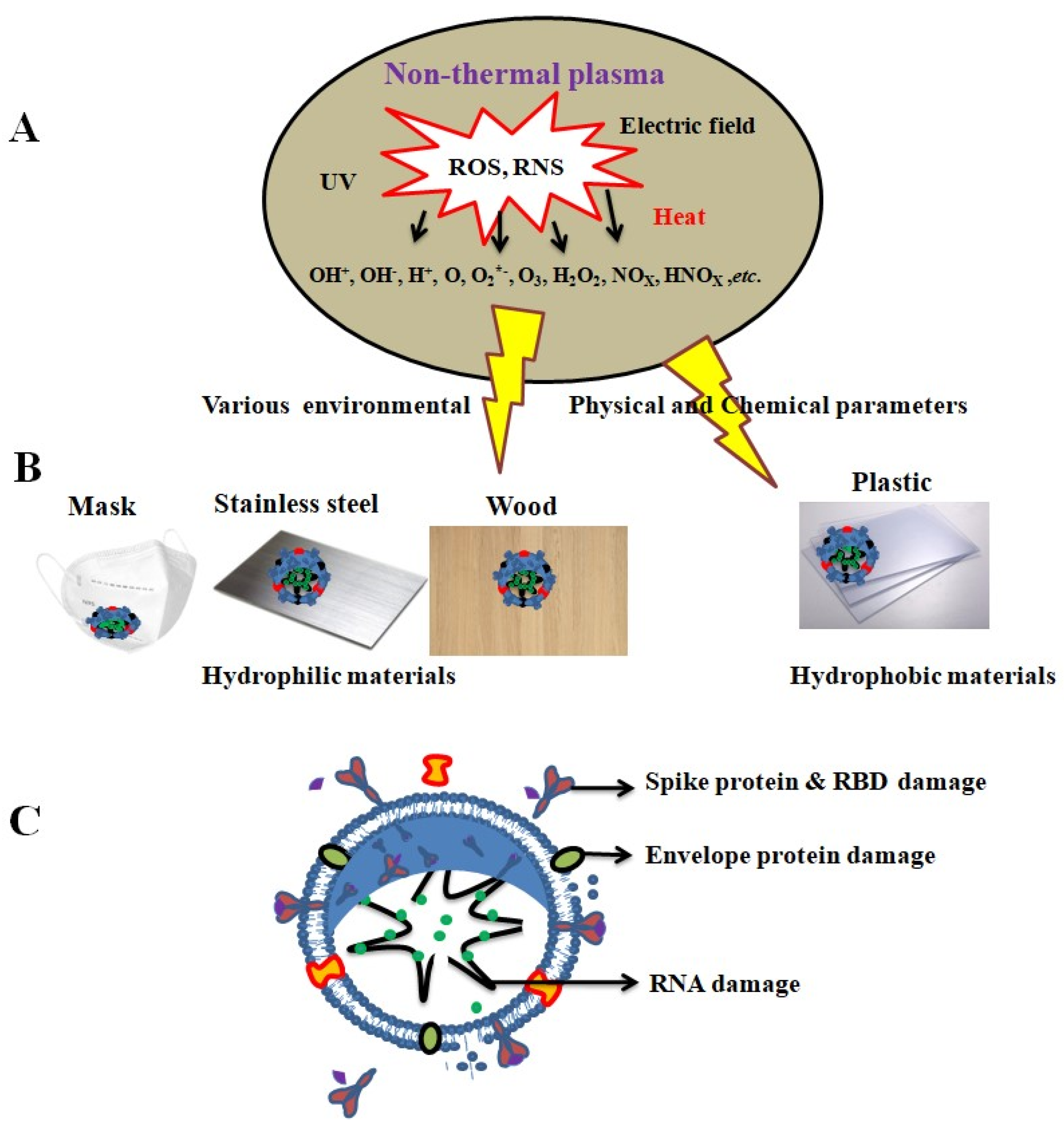
Non-Thermal Plasma (NTP)
2.4. Virus Persistence and Inactivation Mechanisms
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Information Consent Statement
Conflicts of Interest
References
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-Cov-2 Spike Glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, L.; Dai, G.; Chen, R.A.; Liu, D.X.; Fung, T.S. Regulation of the ER Stress Response by the Ion Channel Activity of the Infectious Bronchitis Coronavirus Envelope Protein Modulates Virion Release, Apoptosis, Viral Fitness, and Pathogenesis. Front. Microbiol. 2020, 10, 3022. [Google Scholar] [CrossRef]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus Membrane Fusion Mechanism Offers a Potential Target for Antiviral Development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ning, Z.; Chen, Y.; Guo, M.; Liu, Y.; Gali, N.K.; Sun, L.; Duan, Y.; Cai, J.; Westerdahl, D.; et al. Aerodynamic analysis of SARS-CoV-2 in two Wuhan hospitals. Nature 2020, 582, 557–560. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex immune dysregulation in COVID-19 patients with severe respiratory failure. Cell Host Microbe 2020, 27, 992–1000. [Google Scholar] [CrossRef]
- Guo, G.; Ye, L.; Pan, K.; Chen, Y.; Xing, D.; Yan, K.; Chen, Z.; Ding, N.; Li, W.; Huang, H.; et al. New Insights of Emerging SARS-CoV-2: Epidemiology, Etiology, Clinical Features, Clinical Treatment, and Prevention. Front. Cell Dev. Biol. 2020, 8, 410. [Google Scholar] [CrossRef]
- Killingley, B.; Nguyen-Van-Tam, J. Routes of influenza transmission. Influ. Other Respir. Viruses 2013, 7, 42–51. [Google Scholar] [CrossRef]
- Fathizadeh, H.; Maroufi, P.; Momen-Heravi, M.; Dao, S.; Köse, Ş.; Ganbarov, K.; Pagliano, P.; Esposito, S.; Kafil, H.S. Protection and disinfection policies against SARS-CoV-2 (COVID-19). Inf. Med. 2020, 28, 185–191. [Google Scholar]
- Kutter, J.S.; Spronken, M.I.; Fraaij, P.L.; Fouchier, R.A.; Herfst, S. Transmission routes of respiratory viruses among humans. Curr. Opin. Virol. 2018, 28, 142–151. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Natural Ventilation for Infection Control in Health-Care Settings (World Health Organization Guidelines). 2009. Available online: https://apps.who.int/iris/handle/10665/44167 (accessed on 1 January 2009).
- Yu, I.T.; Li, Y.; Wong, T.W.; Tam, W.; Chan, A.T.; Lee, J.H.; Leung, D.Y.; Ho, T. Evidence of airborne transmission of the severe acute respiratory syndrome virus. N. Engl. J. Med. 2004, 350, 1731–1739. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef] [PubMed]
- van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and Surface Stability of SARS-CoV-2 as Compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef] [PubMed]
- Kasloff, S.B.; Leung, A.; Strong, J.E.; Funk, D.; Cutts, T. Stability of SARS-CoV-2 on critical personal protective equipment. Sci. Rep. 2021, 11, 984. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Murallidharan, J.S.; Agrawal, A.; Bhardwaj, R. Why coronavirus survives longer on impermeable than porous surfaces. Phys. Fluids 2021, 33, 021701. [Google Scholar] [CrossRef]
- Dika, C.; Ly-Chatain, M.H.; Francius, G.; Duval, J.F.L.; Gantzer, C. Non-DLVO adhesion of F-specific RNA bacteriophages toabiotic surfaces: Importance of surface roughness, hydrophobic and electrostatic interactions. Colloids Surf. A 2013, 435, 178–187. [Google Scholar] [CrossRef]
- Joonaki, E.; Hassanpouryouzband, A.; Heldt, C.L.; Areo, O. Surface Chemistry Can Unlock Drivers of Surface Stability of SARS-CoV-2 in a Variety of Environmental Conditions. Chem 2020, 6, 2135–2146. [Google Scholar] [CrossRef]
- Gomez-Mendoza, M.; Banyasz, A.; Douki, T.; Markovitsi, D.; Ravanat, J.L. Direct oxidative damage of naked DNA generated upon absorption of UV radiation by nucleobases. J. Phys. Chem. Lett. 2016, 7, 3945–3948. [Google Scholar] [CrossRef]
- Shi, H.; Tarabara, V.V. Charge, Size distribution and hydrophobicity of viruses: Effect of propagation and purification methods. J. Virol. Methods 2018, 256, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Sakoda, A.; Sakai, Y.; Hayakawa, K.; Suzuki, M. Adsorption of viruses in water environment onto solid surfaces. Water Sci. Technol. 1997, 35, 107–114. [Google Scholar] [CrossRef]
- Zhou, J.; Wei, J.; Choy, K.T.; Sia, S.F.; Rowlands, D.K.; Yu, D.; Wu, C.Y.; Lindsley, W.G.; Cowling, B.J.; McDevitt, J.; et al. Defining the sizes of airborne particles that mediate influenza transmission in ferrets. Proc. Natl. Acad. Sci. USA 2018, 115, E2386–E2392. [Google Scholar] [CrossRef] [PubMed]
- Aaron, M.; Kevin, Z.; Joe, B.; Mausumi, B.; Margo, A.B.; Federico, C.; Matthew, J.H.; Christine, E.S. Zika virus RNA persistence in sewage. Environ. Sci. Technol. Lett. 2020, 7, 659–664. [Google Scholar]
- Yuan, J.; Yun, H.; Lan, W.; Wang, W.; Sullivan, S.G.; Jia, S.; Bittles, A.H. A climatologic investigation of the SARS-CoV outbreak in Beijing, China. Am. J. Infect. Control 2006, 34, 234–236. [Google Scholar] [CrossRef]
- Amuakwa-Mensah, F.; Marbuah, G.; Mubanga, M. Climate variability and infectious diseases nexus: Evidence from Sweden. Infect. Dis. Modell. 2017, 2, 203–217. [Google Scholar] [CrossRef] [PubMed]
- WHO. Transmission of SARS-CoV-2: Implications for Infection Prevention Precautions: Scientific Brief. 2020. Available online: https://www.who.int/news-room/commentaries/detail/transmission-of-sars-cov-2-implications-for-infection-prevention-precautions (accessed on 22 September 2020).
- Azuma, K.; Kagi, N.; Kim, H.; Hayashi, M. Impact of climate and ambient air pollution on the epidemic growth during COVID-19 outbreak in Japan. Environ. Res. 2020, 190, 110042. [Google Scholar] [CrossRef]
- Nishiura, H.; Oshitani, H.; Kobayashi, T.; Saito, T.; Sunagawa, T.; Matsui, T.; Wakita, T.; MHLW COVID-19 Response Team; Suzuki, M. Closed environments facilitate secondary transmission of coronavirus disease 2019 (COVID-19). medRxiv 2020, 2. [Google Scholar] [CrossRef]
- Otter, J.A.; Donskey, C.; Yezli, S.; Douthwaite, S.; Goldenberg, S.D.; Weber, D.J. Transmission of SARS and MERS coronaviruses and influenza virus in healthcare settings: The possible role of dry surface contamination. J. Hosp. Infect. 2016, 92, 235e50. [Google Scholar] [CrossRef]
- Sehulster, L.; Chinn, R.Y.; Hicpac, C.D.C. Guidelines for environmental infection control in health-care facilities: Recommendations of the CDC and the Healthcare Infection Control Practices Advisory Committee (HICPAC). MMWR Morb. Mortal. Wkly. Rep. 2003, 52, 145–200. [Google Scholar]
- Huslage, K.; Rutala, W.A.; Sickbert-Bennett, E.; Weber, D.J. A quantitative approach to defining “high-touch” surfaces in hospitals. Infect. Control Hosp. Epidemiol. 2010, 31, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chang, S.Y.; Sung, M.; Park, J.H.; Bin Kim, H.; Lee, H.; Choi, J.P.; Choi, W.S.; Min, J.Y. Extensive Viable Middle East Respiratory Syndrome (MERS) Coronavirus Contamination in Air and Surrounding Environment in MERS Isolation Wards. Clin. Infect. Dis. 2016, 63, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Little, Z.R.; Keevil, C.W. Human coronavirus 229E remains infectious on common touch surface materials. mBio 2015, 6, e01697-15. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.W.H.; Chu, J.T.S.; Perera, M.R.A.; Hui, K.P.Y.; Yen, H.L.; Chan, M.C.W.; Peiris, M.; Poon, L.L.M. Stability of SARS-CoV-2 in different environmental conditions. Lancet Microbe 2020, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Grinshpun, S.A.; Haruta, H.; Eninger, R.M.; Reponen, T.; McKay, R.T.; Lee, S.A. Performance of an N95 filtering facepiece particulate respirator and a surgical mask during human breathing: Two pathways for particle penetration. J. Occup. Environ. Hyg. 2009, 6, 593–603. [Google Scholar] [CrossRef]
- Monge, F.A.; Jagadesan, P.; Bondu, V.; Donabedian, P.L.; Ista, L.; Chi, E.Y.; Schanze, K.S.; Whitten, D.G.; Kell, A.M. Highly Effective Inactivation of SARS-CoV-2 by Conjugated Polymers and Oligomers. ACS Appl. Mater. Interfaces 2020, 12, 55688–55695. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Joshi, R.; Yasir, M.; Adhikari, A.; Chughtai, A.A.; Heslop, D.; Bull, R.; Willcox, M.; Macintyre, C.R. Graphene- and Nanoparticle-Embedded Antimicrobial and Biocompatible Cotton/Silk Fabrics for Protective Clothing. ACS Appl. Bio Mater. 2021, 4, 6175–6185. [Google Scholar] [CrossRef]
- Kasbe, P.S.; Gade, H.; Liu, S.; Chase, G.G.; Xu, W. Ultrathin Polydopamine-Graphene Oxide Hybrid Coatings on Polymer Filters with Improved Filtration Performance and Functionalities. ACS Appl. Bio Mater. 2021, 4, 5180. [Google Scholar] [CrossRef]
- Jung, S.; Yang, J.Y.; Byeon, E.Y.; Kim, D.G.; Lee, D.G.; Ryoo, S.; Lee, S.; Shin, C.W.; Jang, H.W.; Kim, H.J.; et al. Copper-Coated Polypropylene Filter Face Mask with SARS-COV-2 Antiviral Ability. Polymers 2021, 13, 1367. [Google Scholar] [CrossRef]
- Rosilo, H.; McKee, J.R.; Kontturi, E.; Koho, T.; Hytönen, V.P.; Ikkala, O.; Kostiainen, M.A. Cationic polymer brush-modified cellulose nanocrystals for high-affinity virus binding. Nanoscale 2014, 6, 11871–11881. [Google Scholar] [CrossRef]
- Pandey, L.M. Surface engineering of personal protective equipment (PPEs) to prevent the contagious infections of SARS-CoV-2. Surf. Eng. 2020, 36, 901–907. [Google Scholar] [CrossRef]
- Dika, C.; Duval, J.F.; Francius, G.; Perrin, A.; Gantzer, C. Isoelectric point is an inadequate descriptor of MS2, Phi X 174 and PRD1 phages adhesion on abiotic surfaces. J. Colloid. Interface Sci. 2015, 446, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Bonny, T.S.; Yezli, S.; Lednicky, J.A. Isolation and identification of human coronavirus 229E from frequently touched environmental surfaces of a university classroom that is cleaned daily. Am. J. Infect. Control 2018, 46, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Schilling-Loeffler, K.; Falkenhagen, A.; Johne, R. Coronaviruses are stable on glass, but are eliminated by manual dish washing procedures. Food Microbiol. 2022, 106, 104036. [Google Scholar] [CrossRef]
- Saeed, B.; Alex, C.; Mohsen, H.; Leo, P.; William, A.D. A Surface Coating that Rapidly Inactivates SARS-CoV-2. Appl. Mater. Interfaces 2020, 12, 34723–34727. [Google Scholar] [CrossRef]
- Riddell, S.; Goldie, S.; Hill, A.; Eagles, D.; Drew, T.W. The effect of temperature on persistence of SARS-CoV-2 on common surfaces. Virol. J. 2020, 17, 145. [Google Scholar] [CrossRef]
- Ratnesar-Shumate, S.; Williams, G.; Green, B.; Krause, M.; Holland, B.; Wood, S.; Bohannon, J.; Boydston, J.; Freeburger, D.; Hooper, I.; et al. Simulated Sunlight Rapidly Inactivates SARS-CoV-2 on Surfaces. J. Infect. Dis. 2020, 29, 214–222. [Google Scholar] [CrossRef]
- Darnell, M.E.; Subbarao, K.; Feinstone, S.M.; Taylor, D.R. Inactivation of the coronavirus that induces severe acute respiratory syndrome, SARS-CoV. J. Virol. Methods 2004, 121, 85–91. [Google Scholar] [CrossRef]
- Sizun, J.; Yu, M.W.; Talbot, P.J. Survival of human coronaviruses 229E and OC43 in suspension and after drying on surfaces: A possible source of hospital-acquired infections. J. Hosp. Infect. 2000, 46, 55–60. [Google Scholar] [CrossRef]
- vanDoremalen, N.; Bushmaker, T.; Munster, V.J. Stability of Middle East respiratory syndrome coronavirus (MERS-CoV) under different environmental conditions. Eurosurveillance 2013, 18, 20590. [Google Scholar] [CrossRef]
- Duan, S.M.; Zhao, X.S.; Wen, R.F.; Huang, J.J.; Pi, G.H.; Zhang, S.X.; Han, J.; Bi, S.L.; Ruan, L.; Dong, X.P. Stability of SARS coronavirus in human specimens and environment and its sensitivity to heating and UV irradiation. Biomed. Environ. Sci. 2003, 16, 246–255. [Google Scholar] [PubMed]
- Ben-Shmuel, A.; Brosh-Nissimov, T.; Glinert, I.; Bar-David, E.; Sittner, A.; Poni, R.; Cohen, R.; Achdout, H.; Tamir, H.; Yahalom-Ronen, Y.; et al. Detection and infectivity potential of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) environmental contamination in isolation units and quarantine facilities. Clin. Microbiol. Infect. 2020, 26, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, T.; Deng, Y.; Liu, S.; Zhang, D.; Li, H.; Wang, X.; Jia, L.; Han, J.; Bei, Z.; et al. Stability of SARS-CoV-2 on environmental surfaces and in human excreta. J. Hosp. Infect. 2021, 107, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Oxford, J.; Berezin, E.N.; Courvalin, P.; Dwyer, D.E.; Exner, M.; Jana, L.A.; Kaku, M.; Lee, C.; Letlape, K.; Low, D.E.; et al. The survival of influenza A(H1N1)pdm09 virus on 4 household surfaces. Am. J. Infect. Control 2014, 42, 423–425. [Google Scholar] [CrossRef]
- Thompson, K.A.; Bennett, A.M. Persistence of influenza on surfaces. J. Hosp. Infect. 2017, 95, 194–199. [Google Scholar] [CrossRef]
- Michen, B.; Graule, T. Isoelectric points of viruses. J. Appl. Microbiol. 2010, 109, 388–397. [Google Scholar] [CrossRef]
- Gallagher, T.M.; Escarmis, C.; Buchmeier, M.J. Alteration of the pH dependence of coronavirus-induced cell fusion: Effect of mutations in the spike glycoprotein. J. Virol. 1991, 65, 1916–1928. [Google Scholar] [CrossRef]
- Chan, K.H.; Peiris, J.S.; Lam, S.Y.; Poon, L.L.; Yuen, K.Y.; Seto, W.H. The effects of temperature and relative humidity on the viability of the SARS coronavirus. Adv. Virol. 2011, 2011, 734690. [Google Scholar] [CrossRef]
- Casanova, L.M.; Jeon, S.; Rutala, W.A.; Weber, D.J.; Sobsey, M.D. Effects of air temperature and relative humidity on coronavirus survival on surfaces. Appl. Environ. Microbiol. 2010, 76, 2712–2717. [Google Scholar] [CrossRef]
- Smither, S.J.; Eastaugh, L.S.; Findlay, J.S.; Lever, M.S. Experimental aerosol survival of SARS-CoV-2 in artificial saliva and tissue culture media at medium and high humidity. Emerg. Microbes Infect. 2020, 9, 1415–1417. [Google Scholar] [CrossRef]
- Lowen, A.C.; Steel, J.; Mubareka, S.; Palese, P. High temperature (30 °C) blocks aerosol but not contact transmission of influenza virus. J. Virol. 2008, 82, 5650–5652. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.M.; Ko, G. Effect of ultraviolet germicidal irradiation on viral aerosols. Environ. Sci. Technol. 2007, 41, 5460–5465. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.C.; Li, C.S. Inactivation of viruses on surfaces by ultraviolet germicidal irradiation. J. Occup. Environ. Hyg. 2007, 4, 400–405. [Google Scholar] [CrossRef] [PubMed]
- How, Z.T.; Kristiana, I.; Busetti, F.; Linge, K.L.; Joll, C.A. Organic chloramines in chlorine-based disinfected water systems: A critical review. J. Environ. Sci. 2017, 58, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Derraik, J.G.; Anderson, W.A.; Connelly, E.A.; Anderson, Y.C. Rapid evidence summary on SARS-CoV-2 survivorship and disinfection, and a reusable PPE protocol using a double-hit process. MedRxiv 2020, 133, 7. [Google Scholar]
- Rabenau, H.F.; Kampf, G.; Cinatl, J.; Doerr, H.W. Efficacy of various disinfectants against SARS coronavirus. J. Hosp. Infect. 2005, 61, 107–111. [Google Scholar] [CrossRef]
- Moisan, M.; Boudam, K.; Carignan, D.; Kéroack, D.; Levif, P.; Barbeau, J.; Zorzi, W. Sterilization/disinfection of medical devices using plasma: The flowing afterglow of the reduced-pressure N2-O2 discharge as the inactivating medium. Eur. Phys. J. Appl. Phys. 2013, 63, 10001. [Google Scholar] [CrossRef]
- Asculai, S.S.; Weis, M.T.; Rancourt, M.W.; Kupferberg, A.B. Inactivation of Herpes Simplex Viruses by Nonionic Surfactants. Antimicrob. Agents Chemother. 1978, 13, 686–690. [Google Scholar] [CrossRef]
- LeChevallier, M.W.; Au, K.K. Water Treatment and Pathogen Control; IWA Publishing: London, UK, 2004. [Google Scholar]
- Wigginton, K.R.; Pecson, B.M.; Sigstam, T.; Bosshard, F.; Kohn, T. Virus inactivation mechanisms: Impact of disinfectants on virus function and structural integrity Environ. Sci. Technol. 2012, 46, 12069–12078. [Google Scholar] [CrossRef]
- Bidra, A.S.; Pelletier, J.S.; Westover, J.B.; Frank, S.; Brown, S.M.; Tessema, B. Comparison of In Vitro Inactivation of SARS CoV-2 with Hydrogen Peroxide and Povidone-Iodine Oral Antiseptic Rinses. J. Prosthodont. 2020, 29, 599–603. [Google Scholar] [CrossRef]
- Omidbakhsh, N.; Sattar, S.A. Broad-spectrum microbicidal activity, toxicologic assessment, and materials compatibility of a new generation of accelerated hydrogen peroxide-based environmental surface disinfectant. Am. J. Infect. Control 2006, 34, 251–257. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Shimoda, T.; Chikamori, N.; Usuki, S.; Kanai, Y.; Endo, T.; Katsumata, K.; Terashima, C.; Ikekita, M.; Fujishima, A.; et al. Sporicidal performance induced by photocatalytic production of organic peroxide under visible light irradiation. Sci. Rep. 2016, 6, 33715. [Google Scholar] [CrossRef]
- Eggers, M.; Eickmann, M.; Zorn, J. Rapid and Effective Virucidal Activity of Povidone-Iodine Products against Middle East Respiratory Syndrome Coronavirus (MERS-CoV) and Modified Vaccinia Virus Ankara (MVA). Infect. Dis. Ther. 2015, 4, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Guenezan, J.; Garcia, M.; Strasters, D.; Jousselin, C.; Lévêque, N.; Frasca, D.; Mimoz, O. Povidone Iodine Mouthwash, Gargle, and Nasal Spray to Reduce Nasopharyngeal Viral Load in Patients with COVID-19: A Randomized Clinical Trial. JAMA Otolaryngol. Head Neck Surg. 2021, 147, 400–401. [Google Scholar] [CrossRef] [PubMed]
- Akram, M.Z. Inanimate surfaces as potential source of 2019-nCoV spread and their disinfection with biocidal agents. Virus Dis. 2020, 31, 94–96. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, W. Ultraviolet Germicidal Irradiation Handbook; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Buonanno, M.; Randers-Pehrson, G.; Bigelow, A.W.; Trivedi, S.; Lowy, F.D.; Spotnitz, H.M.; Hammer, S.M.; Brenner, D.J. 207-nm UV Light—A Promising Tool for Safe Low-Cost Reduction of Surgical Site Infections. I: In Vitro Studies. PLoS ONE 2013, 8, e76968. [Google Scholar] [CrossRef] [PubMed]
- Buonanno, M.; Ponnaiya, B.; Welch, D.; Stanislauskas, M.; Randers-Pehrson, G.; Smilenov, L.; Lowy, F.D.; Owens, D.M.; Brenner, D.J. Germicidal Efficacy and Mammalian Skin Safety of 222-nm UV Light. Radiat. Res. 2017, 187, 493–501. [Google Scholar] [CrossRef]
- Buonanno, M.; Welch, D.; Shuryak, I.; Brenner, D.J. Far-UVC light (222 nm) efficiently and safely inactivates airborne human coronaviruses. Sci. Rep. 2020, 10, 10285. [Google Scholar] [CrossRef]
- Ravanat, J.L.; Thierry, J.C. Direct and indirect effects of UV radiation on DNA and its components. J. Photochem. Photobiol. B Biol. 2001, 63, 88–102. [Google Scholar] [CrossRef]
- Matsuura, R.; Lo, C.W.; Wada, S.; Somei, J.; Ochiai, H.; Murakami, T.; Saito, N.; Ogawa, T.; Shinjo, A.; Benno, Y.; et al. SARS-CoV-2 Disinfection of Air and Surface Contamination by TiO2 Photocatalyst-Mediated Damage to Viral Morphology, RNA, and Protein. Viruses 2021, 13, 942. [Google Scholar] [CrossRef]
- Leung, K.C.P.; Ko, T.C.S. Improper Use of the Germicidal Range Ultraviolet Lamp for Household Disinfection Leading to Phototoxicity in COVID-19 Suspects. Cornea 2021, 40, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Iida, Y.; Matsuda, A.; Kuwabara, M. Damage induced by hydroxyl radicals generated in the hydration layer of gamma-irradiated frozen aqueous solution of DNA. J. Radiat. Res. 1996, 37, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Lowy, R.J.; Vavrina, G.A.; LaBarre, D.D. Comparison of gamma and neutron radiation inactivation of influenza A virus. Antivir. Res. 2001, 52, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, F.; Shupert, W.L.; Haddock, E.; Twardoski, B.; Feldmann, H. Gamma Irradiation as an Effective Method for Inactivation of Emerging Viral Pathogens. Am. J. Trop. Med. Hyg. 2019, 100, 1275–1277. [Google Scholar] [CrossRef]
- Hume, A.J.; Ames, J.; Rennick, L.J.; Duprex, W.P.; Marzi, A.; Tonkiss, J.; Mühlberger, E. Inactivation of RNA Viruses by Gamma Irradiation: A Study on Mitigating Factors. Viruses 2016, 8, 204. [Google Scholar] [CrossRef]
- Guild, W.R. The inactivation of bacteriophage and catalase by soft X-rays. Arch. Biochem. Biophys. 1952, 40, 402–415. [Google Scholar] [CrossRef]
- von Sonntag, C.; von Gunten, U. Chemistry of Ozone in Water and Wastewater Treatment: From Basic Principles to Applications; IWA Publishing: London, UK, 2012. [Google Scholar] [CrossRef]
- Clavo, B.; Cordoba-Lanús, E.; Rodríguez-Esparragon, F.; Cazorla-Rivero, S.E.; GarcíaP’erez, O.; Pinero, J.E.; Villar, J.; Blanco, A.; Torres-Ascension, C.; Martín-Barrasa, J.L.; et al. Effects of ozone treatment on personal protective equipment contaminated with sars-cov-2. Antioxidants 2020, 9, 1222. [Google Scholar] [CrossRef]
- Yano, H.; Nakano, R.; Suzuki, Y.; Nakano, A.; Kasahara, K.; Hosoi, H. Inactivation of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) by gaseous ozone treatment. J. Hosp. Infect. 2020, 106, 837–838. [Google Scholar] [CrossRef]
- Choi, E.H.; Uhm, H.S.; Kaushik, N.K. Plasma bioscience and its application to medicine. AAPPS Bull. 2021, 31, 10. [Google Scholar] [CrossRef]
- Augusto, G.A. Ozone—An emerging technology for the seafood industry. Braz. Arch. Biol. Technol. 2009, 52, 15271539. [Google Scholar] [CrossRef]
- Han, I.; Mumtaz, S.; Choi, E.H. Nonthermal Biocompatible Plasma Inactivation of Coronavirus SARS-CoV-2: Prospects for Future Antiviral Applications. Viruses 2022, 14, 2685. [Google Scholar] [CrossRef] [PubMed]
- Neha, K.; Sarmistha, M.; Eun, J.B.; Linh, N.N.; Pradeep, B.; June, H.K.; Eun, H.C.; Nagendra, K.K. The inactivation and destruction of viruses by reactive oxygen species generated through physical and cold atmospheric plasma techniques: Current status and perspectives. J. Adv. Res. 2023, 43, 59–71. [Google Scholar] [CrossRef]
- Gaens, W.V.; Bogaerts, A. Kinetic Modelling for an Atmospheric Pressure Argon Plasma Jet in Humid Air. J. Phys. D Appl. Phys. 2013, 46, 275201. [Google Scholar] [CrossRef]
- Reuter, S.; von Woedtke, T.; Weltmann, K.D. The kINPen—A Review on Physics and Chemistry of the Atmospheric Pressure Plasma Jet and its Applications. J. Phys. D Appl. Phys. 2018, 51, 233001. [Google Scholar] [CrossRef]
- Graves, D.B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar] [CrossRef]
- Wende, K.; Williams, P.; Dalluge, J.; Gaens, W.V.; Aboubakr, H.; Bischof, J.; von Woedtke, T.; Goyal, S.M.; Weltmann, K.D.; Bogaerts, A.; et al. Identification of the biologically active liquid chemistry induced by a nonthermal atmospheric pressure plasma jet. Biointerphases 2015, 10, 029518. [Google Scholar] [CrossRef]
- Guo, L.; Yao, Z.; Yang, L.; Zhang, H.; Qi, Y.; Gou, L.; Xi, W.; Liu, D.; Zhang, L.; Cheng, Y.; et al. Plasma-activated Water: An Alternative Disinfectant for S Protein Inactivation to Prevent SARS-CoV-2 Infection. Chem. Eng. J. 2020, 421, 127742. [Google Scholar] [CrossRef]
- Zimmermann, J.L.; Dumler, K.; Shimizu, T.; Morfill, G.E.; Wolf, A.; Boxhammer, V.; Schlegel, J.; Gansbacher, B.; Anton, M. Effects of Cold Atmospheric Plasmas on Adenoviruses in Solution. J. Phys. D Appl. Phys. 2011, 44, 505201. [Google Scholar] [CrossRef]
- Bisag, A.; Isabelli, P.; Laurita, R.; Bucci, C.; Capelli, F.; Dirani, G.; Gherardi, M.; Laghi, G.; Paglianti, A.; Sambri, V.; et al. Cold Atmospheric Plasma Inactivation of Aerosolized Microdroplets Containing Bacteria and Purified SARS-CoV-2 RNA to Contrast Airborne Indoor Transmission. Plasma Process. Polym. 2020, 17, 2000154. [Google Scholar] [CrossRef]
- Attri, P.; Koga, K.; Shiratani, M. Possible Impact of Plasma Oxidation on the Structure of the C-Terminal Domain of SARS-CoV-2 Spike Protein: A Computational Study. Appl. Phys. Express. 2021, 14, 027002. [Google Scholar] [CrossRef]
- Chen, Z.; Garcia, G.J.; Arumugaswami, V.; Richard, E.W. Cold atmospheric plasma for SARS-CoV-2 inactivation. Phys. Fluids 2020, 32, 111702. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Bong, C.; Bae, P.K.; Abafogi, A.T.; Baek, S.H.; Shin, Y.-B.; Bak, M.S.; Park, S. Fast and easy disinfection of coronavirus-1 contaminated face masks using ozone gas produced by a dielectric barrier discharge plasma generator. Environ. Sci. Technol. Lett. 2021, 8, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, P.; Lim, J.S.; Kaushik, N.; Shaik, A.M.; Shin, Y.O.; Kaushik, N.K.; Choi, E.H. Nonthermal plasma-generated ozone inhibits human coronavirus 229E infectivity on glass surface. Plasma Process. Polym. 2022, 19, e2200054. [Google Scholar] [CrossRef]
- Yamashiro, R.; Misawa, T.; Sakudo, A. Key role of singlet oxygen and peroxynitrite in viral RNA damage during virucidal effect of plasma torch on feline calicivirus. Sci. Rep. 2018, 8, 17947. [Google Scholar] [CrossRef]
- Qin, H.; Qiu, H.; He, S.T.; Hong, B.; Liu, K.; Lou, F.; Li, M.; Hu, P.; Kong, X.; Song, Y.; et al. Efficient disinfection of SARS-CoV-2-like coronavirus, pseudotyped SARS-CoV-2 and other coronaviruses using cold plasma induces spike protein damage. J. Hazard. Mater. 2022, 430, 128414. [Google Scholar] [CrossRef]
- Guo, L.; Xu, R.; Gou, L.; Liu, Z.; Zhao, Y.; Liu, D.; Zhang, L.; Chen, H.; Kong, M.G. Mechanism of virus inactivation by cold atmospheric-pressure plasma and plasma-activated water. Appl. Environ. Microbiol. 2018, 84, e00726-18. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Walsh, K.; Willcox, M.; Morgan, P.; Nichols, J. The COVID-19 pandemic: Important considerations for contact lens practitioners. Cont. Lens. Anterior. Eye 2020, 43, 196–203. [Google Scholar] [CrossRef]
- Choi, E.H.; Kaushik, N.K.; Hong, Y.J.; Lim, J.S.; Choi, J.S.; Han, I. Plasma bioscience for medicine, agriculture and hygiene applications. J. Korean Phys. Soc. 2022, 80, 817–851. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Tang, W.; Yu, L.; Liu, Z.; Liu, Y.; Xia, H.; Zhang, H.; Chen, S.; Wu, J.; Cui, X.; et al. Inactivating SARS-CoV-2 by electrochemical oxidation. Sci. Bull. 2021, 66, 720–726. [Google Scholar] [CrossRef]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Akerström, S.; Mousavi-Jazi, M.; Klingström, J.; Leijon, M.; Lundkvist, A.; Mirazimi, A. Nitric oxide inhibits the replication cycle of severe acute respiratory syndrome coronavirus. J. Virol. 2005, 79, 1966–1969. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viral Strain and Cell | Source of Materials | Inoculum Amount | Condition | Period | TCID50/mL | Reference |
|---|---|---|---|---|---|---|
| Human Coronavirus 229E, OC43 and embryonic lung cell line L132 | Aluminum and Latex surgical gloves | 10 µL of a ×105 TCID50 | 21 °C | 5 d | 90% removed | [51] |
| Coronavirus 229E and human lung cell MRC-5 | Polyfluorotetraethylene (Teflon; PTFE) | ×103 | 21 °C | 5 d | ND | [35] |
| Polyvinyl chloride (PVC) | ||||||
| Ceramic tiles | ||||||
| Glass | ||||||
| Stainless steel | 90% removed | |||||
| MERS | Plastic | 100 µL of a ×106 TCID50 | 30 °C | 48 h | 80% | [52] |
| Steel | ||||||
| Human Coronavirus SARS and Vero E6 | Wood board | 300 µL of a ×106 TCID50 | Room temperature | 60 h | <20% | [53] |
| Glass | 48 h | 51–75% | ||||
| Metal | 72 h | 26–50% | ||||
| Cloth | 72 h | 26–50% | ||||
| Filter paper | 72 h | 26–50% | ||||
| Plastic | 60 h | 26–50% | ||||
| SARS-CoV-1 and Vero E6 | Plastic | 103.4 TCID50 | 21–23 °C | 72 h | 100.7 | [16] |
| Stainless steel | 103.6 TCID50 | 48 h | 100.6 | |||
| SARS-CoV-2 and Vero E6 | Plastic | 103.7 TCID50 | 72 h | 100.6 | ||
| Stainless steel | 103.7 TCID50 | 48 h | 100.6 | |||
| SARS-CoV-2 and Vero E6 | Steel | 200 µL × 106 PFU | Room temperature | 5 ds | [54] | |
| SARS-CoV-2 and Vero cells | Plastic | 50 µL of a 1.5 × 106 TCID50 | Room temperature | 168 h | 2 log10 | [55] |
| Stainless steel | ||||||
| Glass | ||||||
| Wood | ||||||
| Surgical mask | ||||||
| Latex gloves | 1.8 log10 | |||||
| Cotton | 1.5 log10 | |||||
| Paper | ||||||
| Ceramics | ||||||
| Influenza A(H1N1)pdm09 and Madin Darby Canine Kidney (MDCK) cell | Wood | 500 µL of 5.5 log10 | Room temperature | 48 h | 2.8 | [56] |
| Plastic | 24 h | 2.6 | ||||
| Stainless steel | 2.7 | |||||
| Cloth | 8 h | 1.5 | ||||
| Influenza Cal/7/09/H1N1 and Madin Darby Canine Kidney cells (MDCKs) | Cotton | 10 µL of a 3.3 × 104 pfu/mL | Room temperature | 17.7 h | 1.2 × 103 | [57] |
| Microfibre | 34.3 h | 1.2 × 103 | ||||
| Stainless steel | 174.9 h | 1.9 × 103 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashokkumar, S.; Kaushik, N.K.; Han, I.; Uhm, H.S.; Park, J.S.; Cho, G.S.; Oh, Y.-J.; Shin, Y.O.; Choi, E.H. Persistence of Coronavirus on Surface Materials and Its Control Measures Using Nonthermal Plasma and Other Agents. Int. J. Mol. Sci. 2023, 24, 14106. https://doi.org/10.3390/ijms241814106
Ashokkumar S, Kaushik NK, Han I, Uhm HS, Park JS, Cho GS, Oh Y-J, Shin YO, Choi EH. Persistence of Coronavirus on Surface Materials and Its Control Measures Using Nonthermal Plasma and Other Agents. International Journal of Molecular Sciences. 2023; 24(18):14106. https://doi.org/10.3390/ijms241814106
Chicago/Turabian StyleAshokkumar, Sekar, Nagendra Kumar Kaushik, Ihn Han, Han Sup Uhm, Jang Sick Park, Gyu Seong Cho, Young-Jei Oh, Yung Oh Shin, and Eun Ha Choi. 2023. "Persistence of Coronavirus on Surface Materials and Its Control Measures Using Nonthermal Plasma and Other Agents" International Journal of Molecular Sciences 24, no. 18: 14106. https://doi.org/10.3390/ijms241814106
APA StyleAshokkumar, S., Kaushik, N. K., Han, I., Uhm, H. S., Park, J. S., Cho, G. S., Oh, Y. -J., Shin, Y. O., & Choi, E. H. (2023). Persistence of Coronavirus on Surface Materials and Its Control Measures Using Nonthermal Plasma and Other Agents. International Journal of Molecular Sciences, 24(18), 14106. https://doi.org/10.3390/ijms241814106