
Knocking Out OsAAP11 to Improve Rice Grain Quality Using CRISPR/Cas9 System
Abstract
:1. Introduction
2. Results
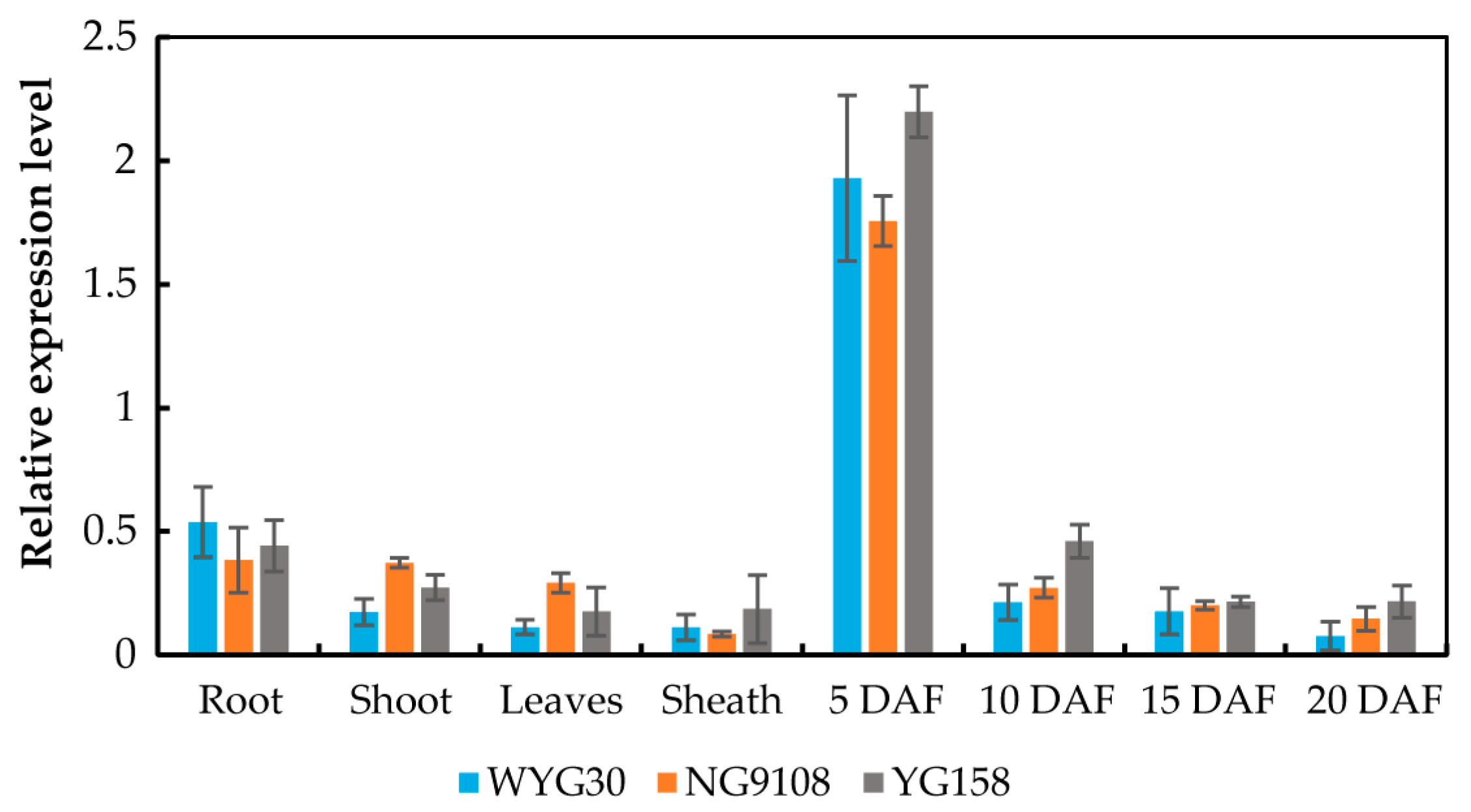
2.1. RT-qPCR Analysis of OsAAP11
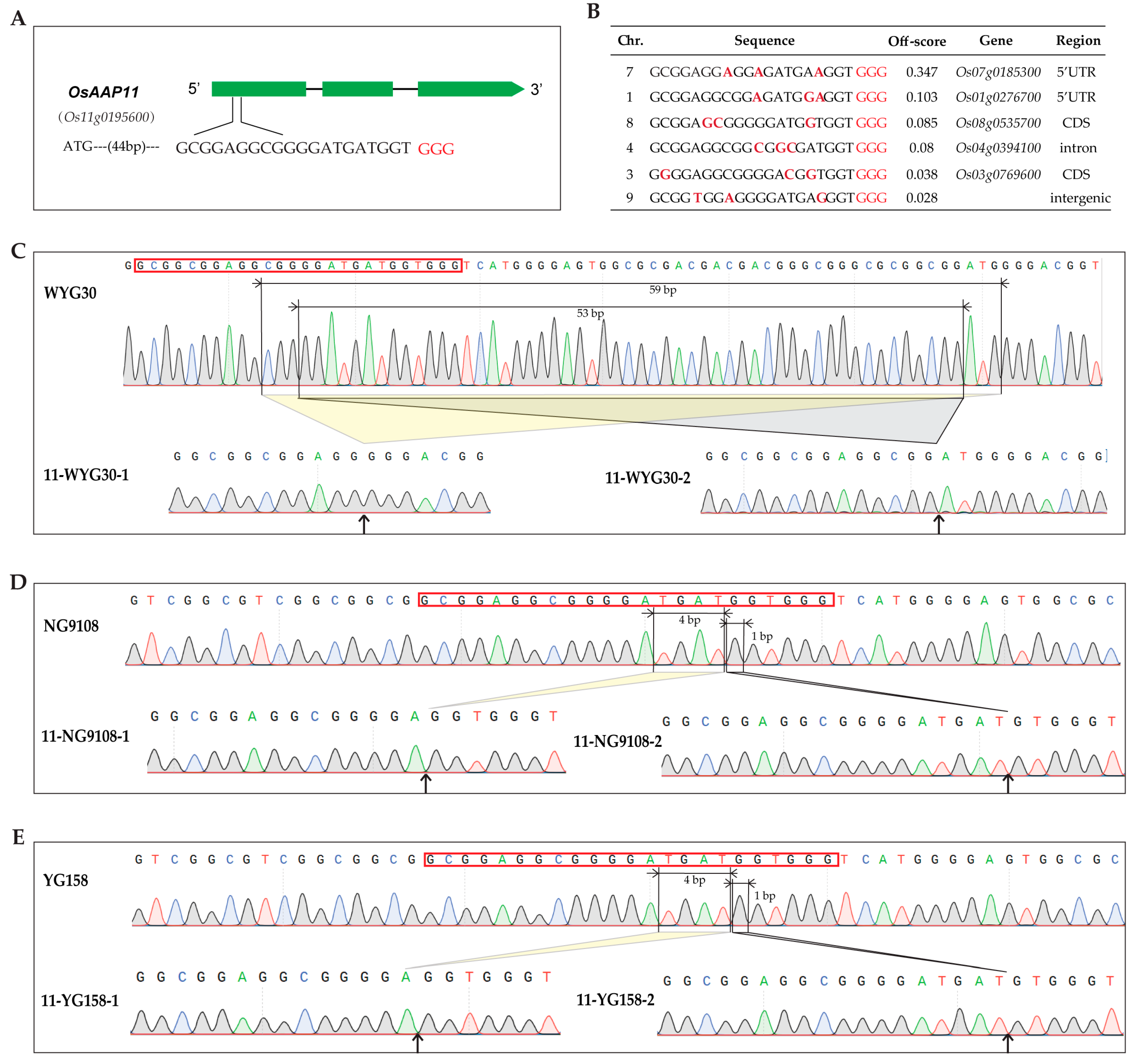
2.2. Generation of osaap11 Mutants
2.3. Agronomic Traits in osaap11 Mutants
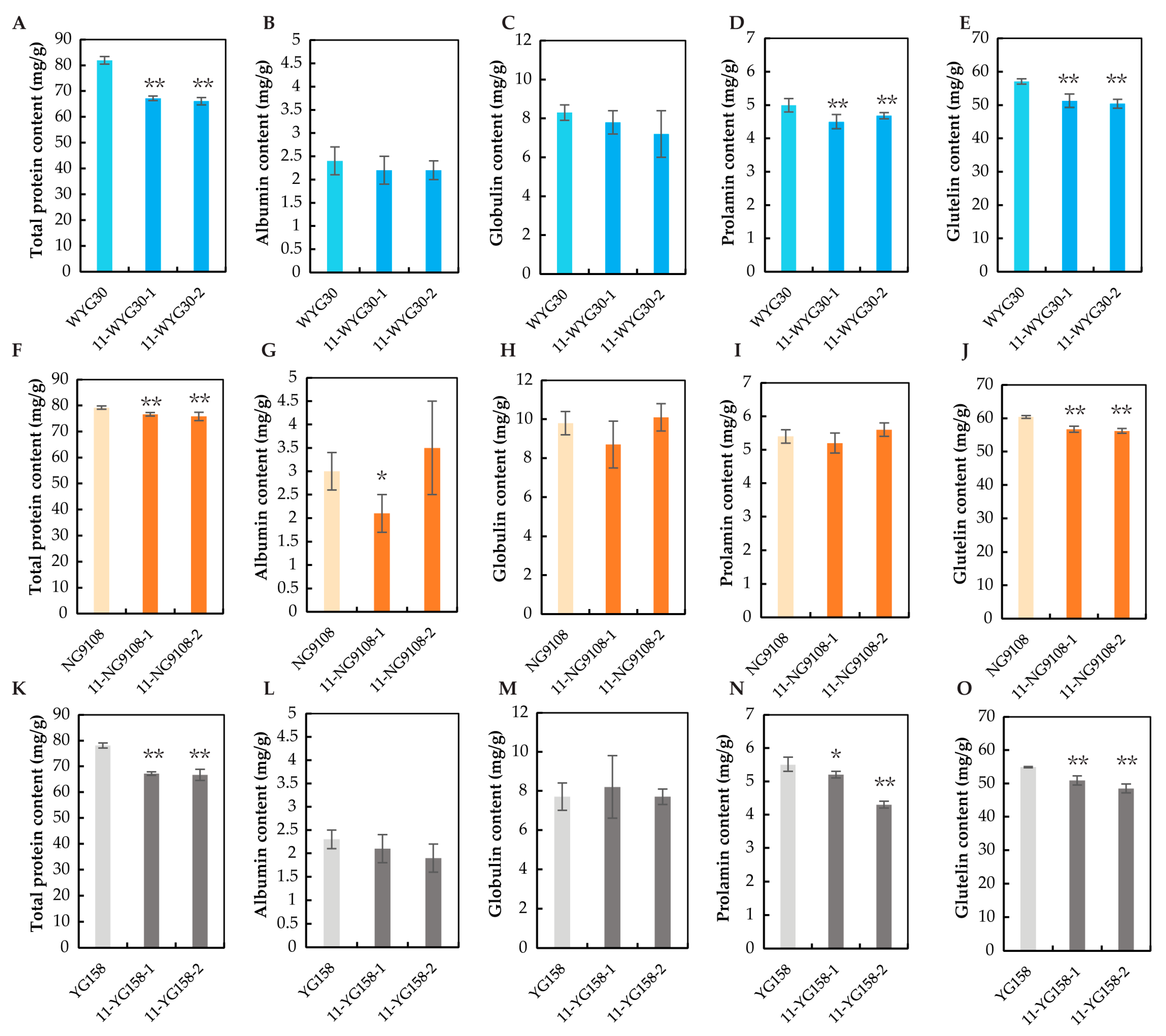
2.4. Grain Protein Content in osaap11 Mutants
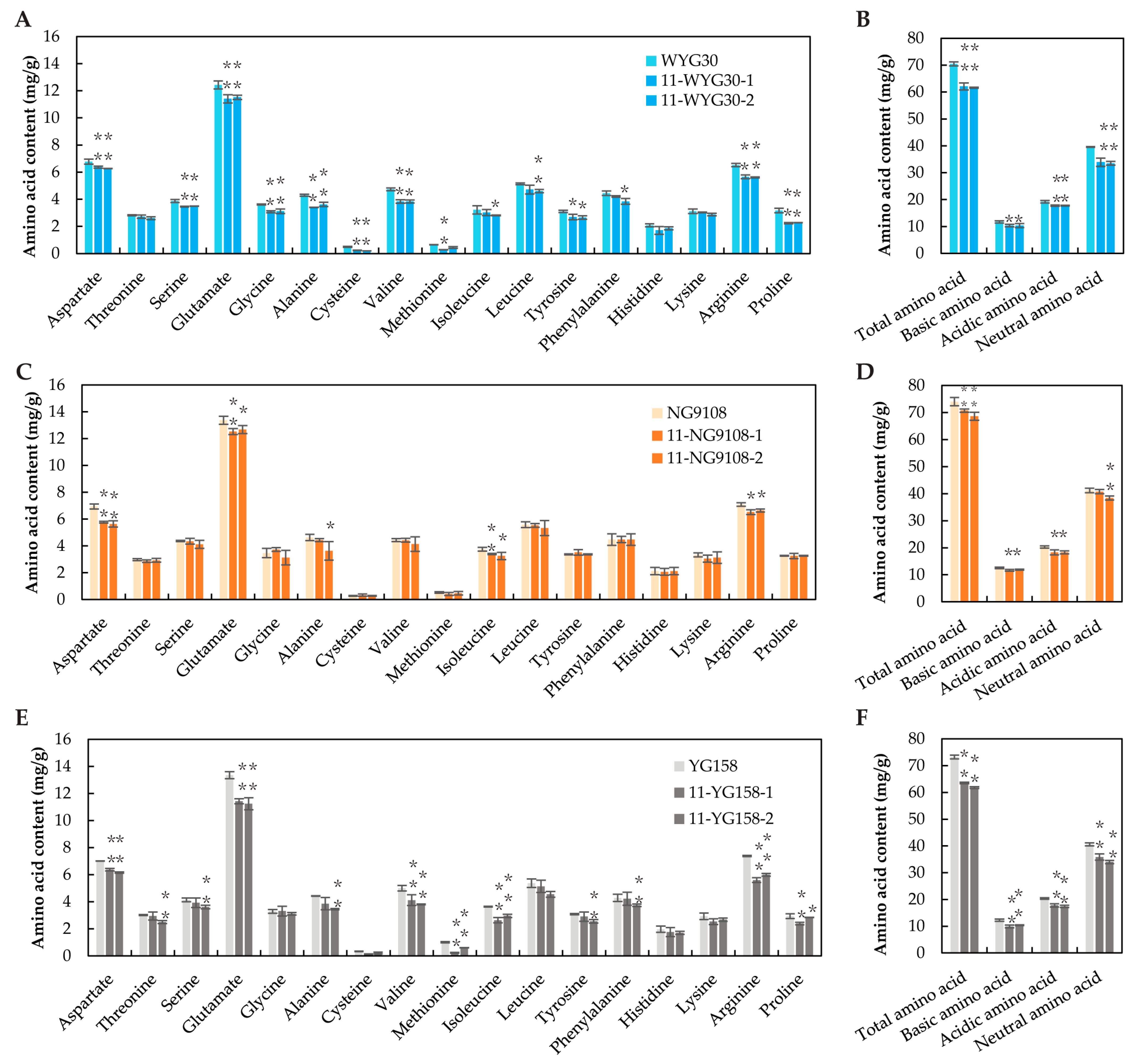
2.5. Amino Acid Content in osaap11 Mutants
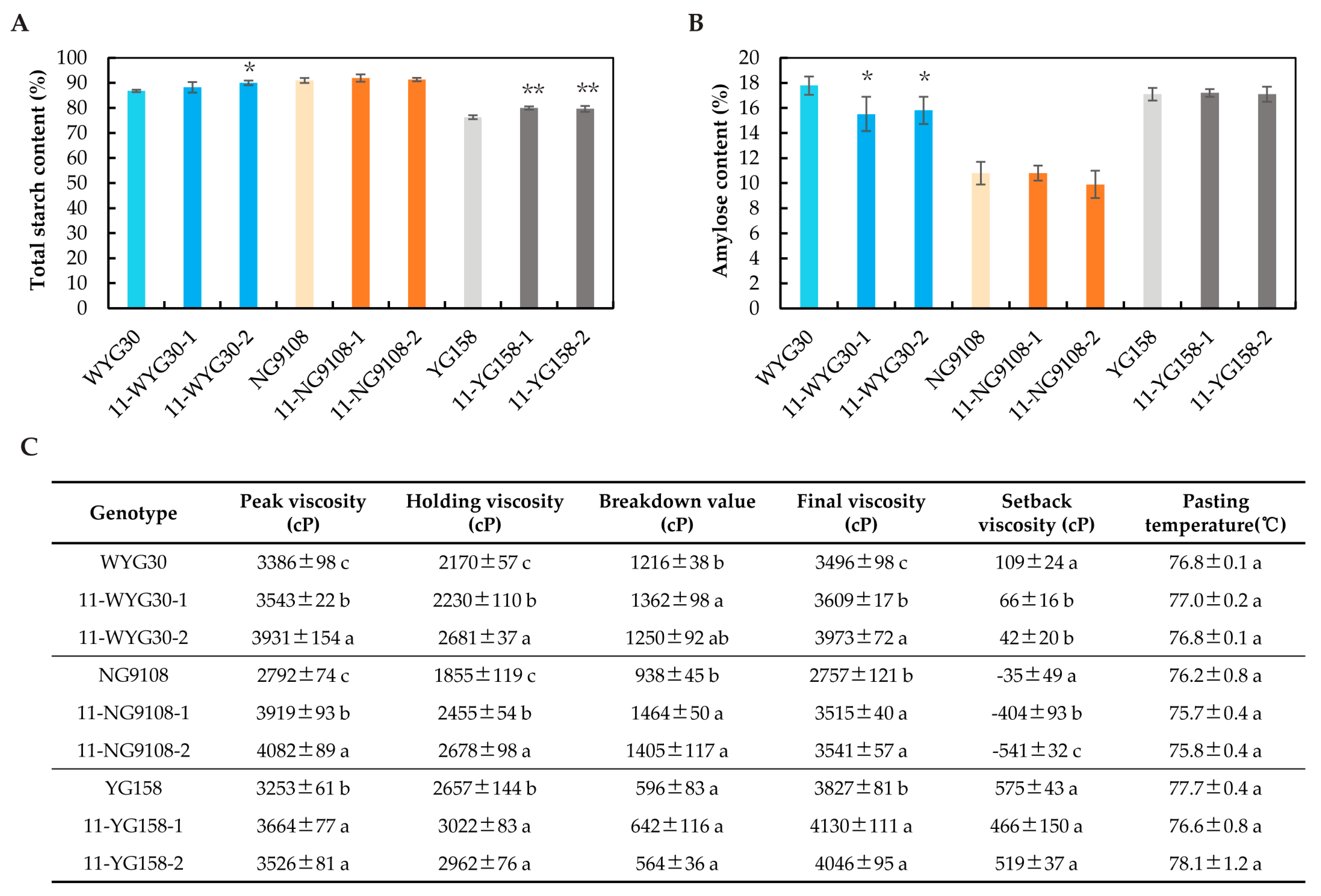
2.6. Starch Content and Physicochemical Properties of osaap11 Mutants
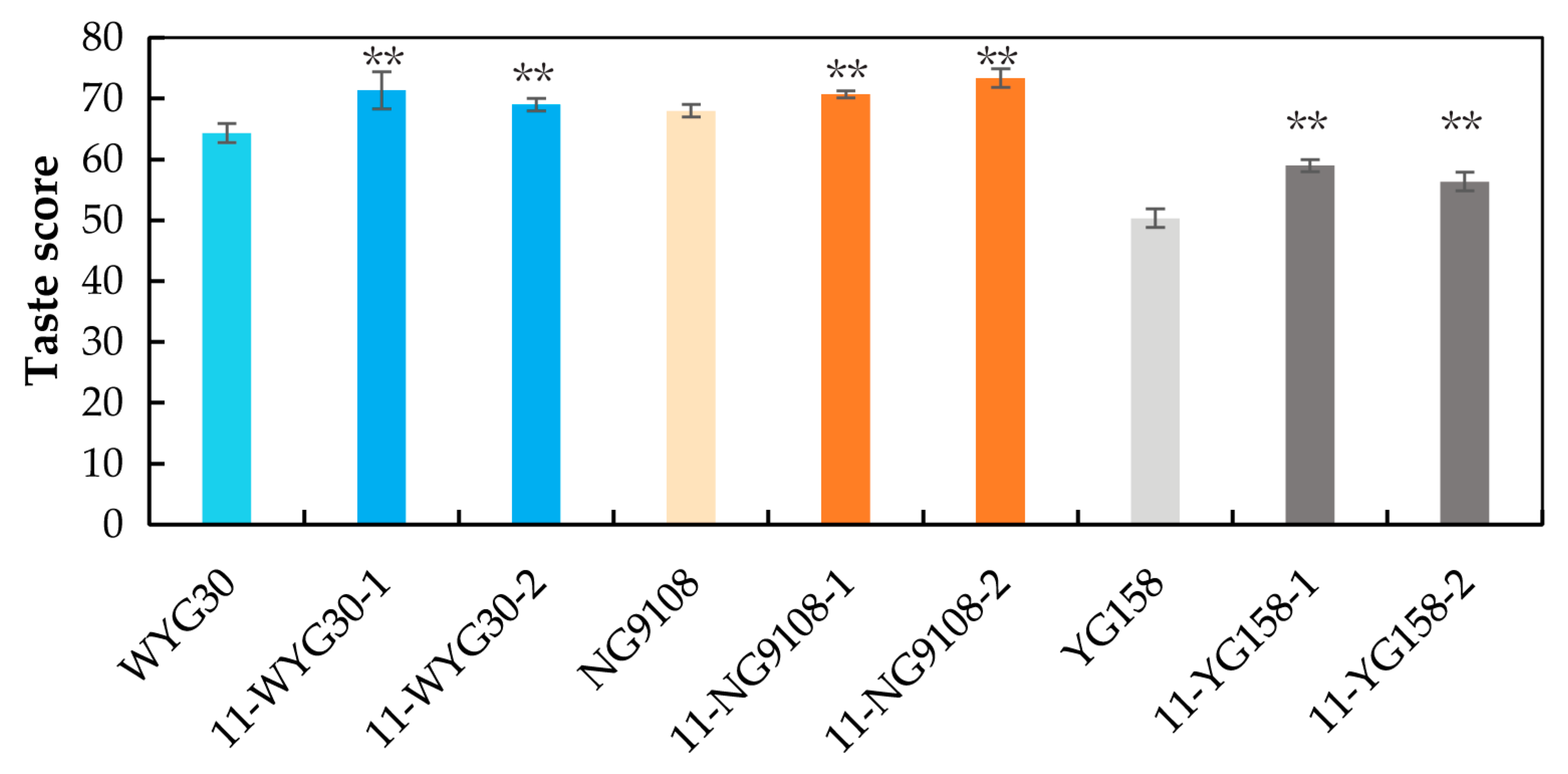
2.7. Taste Scores of osaap11 Mutants
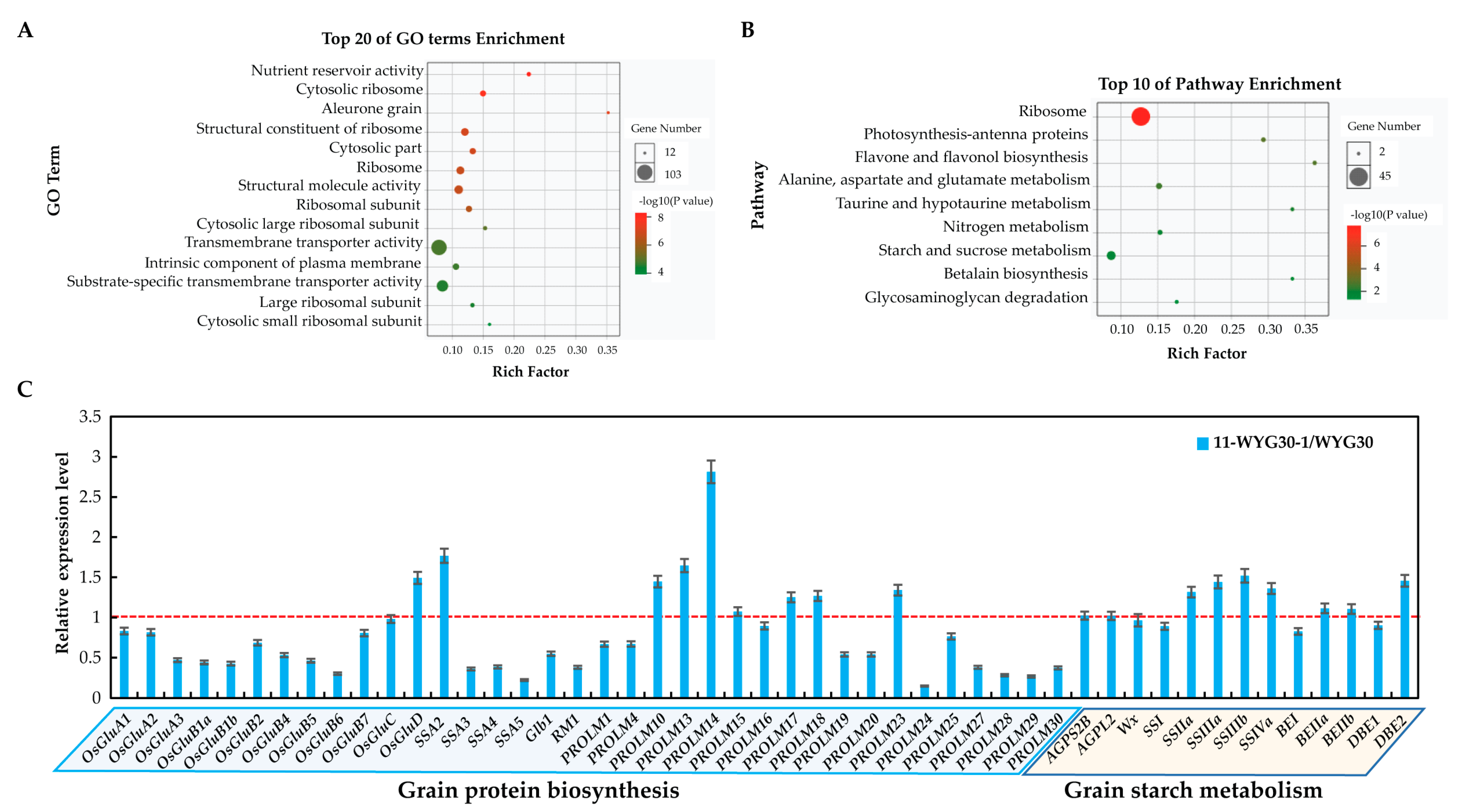
2.8. Transcriptomic Analysis of Genes Related to Protein and Starch Biosynthesis in osaap11 Mutants
3. Discussion
4. Materials and Methods
4.1. Experimental Materials and Plant Method
4.2. Target Site Design and Mutation Detection
4.3. Transgenic Component Detection
4.4. Analysis of Protein Contents and Amino Acid Contents in Rice Grains
4.5. Analysis of Amylose and Total Starch Contents in Rice Grains
4.6. Analysis of Starch Viscosity Characteristics
4.7. Analysis of Rice Taste Scores
4.8. Investigation of Main Agronomic Characteristics of Rice Plants
4.9. Transcriptome Analysis
4.10. RNA Extraction, cDNA Preparation, and qRT-PCR
4.11. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ren, D.; Ding, C.; Qian, Q. Molecular bases of rice grain size and quality for optimized productivity. Sci. Bull. 2023, 68, 314–350. [Google Scholar] [CrossRef] [PubMed]
- Gong, D.; Zhang, X.; He, F.; Chen, Y.; Li, R.; Yao, J.; Zhang, M.; Zheng, W.; Yu, G. Genetic Improvements in Rice Grain Quality: A Review of Elite Genes and Their Applications in Molecular Breeding. Agronomy 2023, 13, 1375. [Google Scholar] [CrossRef]
- Fang, L.; Ma, L.; Zhao, S.; Cao, R.; Jiao, G.; Hu, P.; Wei, X. Alanine aminotransferase (OsAlaAT1) modulates nitrogen utilization, grain yield, and quality in rice. J. Genet. Genom. 2022, 49, 510–513. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wang, W.; Chen, J.; Liu, Y.; Chu, C. Genetic improvement toward nitrogen-use efficiency in rice: Lessons and perspectives. Mol. Plant 2023, 16, 11. [Google Scholar] [CrossRef]
- Salmenkallio, M.; Sopanen, T. Amino Acid and Peptide uptake in the scutella of germinating grains of barley, wheat, rice, and maize. Plant Physiol. 1989, 89, 1285–1291. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Lonhienne, T.; Rentsch, D.; Robinson, N.; Christie, M.; Webb, R.; Gmage, H.; Carroll, B.; Schenk, P.; Schmidt, S. Plants can use protein as a nitrogen source without assistance from other organisms. Proc. Natl. Acad. Sci. USA 2008, 105, 4524–4529. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Bush, D. Proton-coupled sugar and amino acid transporters in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 513–542. [Google Scholar] [CrossRef]
- Tegeder, M.; Rentsch, D. Uptake and partitioning of amino acids and peptides. Mol. Plant 2010, 3, 997–1011. [Google Scholar] [CrossRef]
- Mechthild, T. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar]
- Zhao, H.; Ma, H.; Yu, L.; Wang, X.; Zhao, J. Genome-wide survey and expression analysis of amino acid transporter gene family in rice (Oryza sativa L.). PLoS ONE 2012, 7, e49210. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, S.; Wipf, D.; Frommer, W. Transport mechanisms for organic forms of carbon and nitrogen between source and sink. Annu. Rev. Plant Biol. 2004, 55, 341–372. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Zhang, S.; Gu, M.; Xu, G. Function, transport, and regulation of amino acids: What is missing in rice? Crop J. 2021, 3, 530–542. [Google Scholar] [CrossRef]
- Taylor, M.; Reinders, A.; Ward, J. Transport function of rice amino acid permeases (AAPs). Plant Cell Physiol. 2015, 56, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Huang, W.; Wu, B.; Fang, Z.; Wang, X. The amino acid transporter AAP1 mediates growth and grain yield by regulating neutral amino acid uptake and reallocation in Oryza sativa. J. Exp. Bot. 2020, 71, 4763–4777. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Wu, B.; Wang, J.; Zhu, W.; Nie, H.; Qian, J.; Huang, W.; Fang, Z. Blocking amino acid transporter OsAAP3 improves grain yield by promoting outgrowth buds and increasing tiller number in rice. Plant Bio. J. 2018, 16, 1710–1722. [Google Scholar] [CrossRef]
- Fang, Z.; Wu, B.; Ji, Y. The Amino Acid Transporter OsAAP4 Contributes to Rice Tillering and Grain Yield by Regulating Neutral Amino Acid Allocation through Two Splicing Variants. Rice 2021, 14, 2. [Google Scholar] [CrossRef]
- Wang, J.; Wu, B.; Lu, K.; Wei, Q.; Qian, J.; Chen, Y.; Fang, Z. The Amino Acid Permease 5 (OsAAP5) Regulates Tiller Number and Grain Yield in Rice. Plant Physiol. 2019, 180, 1031–1045. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef]
- Wang, S.; Yang, Y.; Guo, M.; Zhong, C.; Yan, C.; Sun, S. Targeted mutagenesis of amino acid transporter genes for rice quality improvement using the CRISPR/Cas9 system. Crop J. 2020, 8, 457–464. [Google Scholar] [CrossRef]
- Liu, D.; Gong, W.; Bai, Y.; Luo, J.; Zhu, Y. OsHT, a rice gene encoding for a plasma-membrane localized histidine transporter. J. Integr. Plant Biol. 2005, 47, 92–99. [Google Scholar] [CrossRef]
- Wang, X.; Yang, G.; Shi, M.; Hao, D.; Wei, Q.; Wang, Z.; Fu, S.; Su, Y.; Xia, J. Disruption of an amino acid transporter LHT1 leads to growth inhibition and low yields in rice. BMC Plant Biol. 2019, 19, 268. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Hu, J.; Yan, M.; Luo, L.; Tegeder, M.; Xu, G. Oryza sativa Lysine-Histidine-type Transporter 1 functions in root uptake and root-to-shoot allocation of amino acids in rice. Plant J. 2020, 103, 395–411. [Google Scholar] [CrossRef]
- Guo, N.; Gu, M.; Hu, J.; Qu, H.; Xu, G. Rice OsLHT1 Functions in Leaf-to-Panicle Nitrogen Allocation for Grain Yield and Quality. Front. Plant Sci. 2020, 11, 1150. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Chen, Y.; Tang, F.; Xu, F.; Huang, Y.; Chen, H.; Bao, J. Genetic diversity of amylose content and RVA pasting parameters in 20 rice accessions grown in Hainan, China. Food Chem. 2014, 161, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Champagne, E.; Richard, O.; Bett, K.; Grimm, C.; Vinyard, B.; Webb, B.; Mcclung, A.; Barton, F.; Lyon, B.; Moldenhauer, L.; et al. Quality evaluation of U.S. medium-grain rice using a Japanese taste analyzer. Cereal Chem. 1996, 73, 290–294. [Google Scholar]
- Xie, L.; Chen, N.; Duan, B.; Zhu, Z.; Liao, X. Impact of proteins on pasting and cooking properties of waxy and non-waxy rice. J. Cereal Sci. 2008, 47, 372–379. [Google Scholar] [CrossRef]
- Yang, Y.; Shen, Z.; Li, Y.; Xu, C.; Xia, H.; Zhuang, H.; Sun, S.; Guo, M.; Yan, C. Rapid improvement of rice eating and cooking quality through gene editing toward glutelin as target. J. Integr. Plant Biol. 2022, 64, 1860–1865. [Google Scholar] [CrossRef]
- Park, S.; Park, H.; Baek, M.; Jeong, J.; Cho, Y.; Lee, G.; Lee, C.; Suh, J.; Kim, C.; Kim, S. Improving the glossiness of cooked rice, an important component of visual rice grain quality. Rice 2019, 12, 87. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, M.; Sun, S.; Zou, Y.; Yin, S.; Liu, Y.; Tang, S.; Gu, M.; Yang, Z.; Yan, C. Natural variation of OsGluA2 is involved in grain protein content regulation in rice. Nat. Commun. 2019, 10, 1949. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Improving Plant Nitrogen Use Efficiency through Alteration of Amino Acid Transport Processes. Plant Physiol. 2017, 175, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Santiago, J.; Tegeder, M. Connecting Source with Sink: The Role of Arabidopsis AAP8 in Phloem Loading of Amino Acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tan, Q.; Lee, R.; Trethewy, A.; Lee, Y.; Tegeder, M. Altered xylem-phloem transfer of amino acids affects metabolism and leads to increased seed yield and oil content in Arabidopsis. Plant Cell 2010, 22, 3603–3620. [Google Scholar] [CrossRef] [PubMed]
- Rolletschek, H.; Hosein, F.; Miranda, M.; Heim, U.; Gotz, K.; Schlereth, A.; Borisjuk, L.; Saalbach, I.; Wobus, U.; Weber, H. Ectopic Expression of an Amino Acid Transporter (VfAAP1) in Seeds of Vicia narbonensis and Pea Increases Storage Proteins. Plant Physiol. 2005, 137, 1236–1249. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Tiller Number | Grain Number/Panicle | Fertility (%) | Grain Length (cm) | Grain Width (cm) | Grain Weight/100 (g) | |
|---|---|---|---|---|---|---|---|
| WYG30 | 100.9 ± 1.3a | 10.0 ± 0.9a | 192.2 ± 16.0a | 93.1 ± 1.4a | 7.21 ± 0.05bc | 3.65 ± 0.04a | 2.77 ± 0.14a |
| 11-WYG30-1 | 100.9 ± 2.7a | 10.0 ± 1.0a | 194.3 ± 14.0a | 89.2 ± 0.9a | 7.12 ± 0.05c | 3.70 ± 0.02a | 2.66 ± 0.10a |
| 11-WYG30-2 | 100.2 ± 1.9a | 8.3 ± 1.0a | 187.0 ± 20.5a | 88.9 ± 2.4a | 7.26 ± 0.10a | 3.68 ± 0.05a | 2.70 ± 0.12a |
| NG9108 | 94.1 ± 2.1a | 10.5 ± 1.4a | 176.8 ± 10.7a | 90.5 ± 1.7a | 7.11 ± 0.10a | 3.85 ± 0.03a | 2.78 ± 0.07a |
| 11-NG9108-1 | 85.0 ± 3.3b | 9.7 ± 0.6a | 183.3 ± 5.9a | 88.2 ± 2.5a | 7.08 ± 0.03b | 3.72 ± 0.07a | 2.71 ± 0.10ab |
| 11-NG9108-2 | 93.1 ± 1.8a | 10.7 ± 2.1a | 174.3 ± 11.1a | 88.7 ± 1.9a | 7.11 ± 0.10a | 3.79 ± 0.06a | 2.67 ± 0.05b |
| YG158 | 76.9 ± 3.5a | 8.3 ± 0.6a | 130.2 ± 6.9a | 91.7 ± 2.0a | 7.06 ± 0.29b | 3.64 ± 0.04b | 2.78 ± 0.12ab |
| 11-YG158-1 | 70.9 ± 2.2b | 8.7 ± 1.5a | 121.3 ± 9.0a | 89.3 ± 1.4a | 7.09 ± 0.06b | 3.64 ± 0.05b | 2.71 ± 0.06b |
| 11-YG158-2 | 76.6 ± 2.1a | 8.7 ± 2.1a | 134.3 ± 11.6a | 89.7 ± 4.8a | 7.31 ± 0.12a | 3.72 ± 0.03a | 2.82 ± 0.06a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Zhang, Y.; Sun, Z.; Shen, Z.; Li, Y.; Guo, Y.; Feng, Y.; Sun, S.; Guo, M.; Hu, Z.; et al. Knocking Out OsAAP11 to Improve Rice Grain Quality Using CRISPR/Cas9 System. Int. J. Mol. Sci. 2023, 24, 14360. https://doi.org/10.3390/ijms241814360
Yang Y, Zhang Y, Sun Z, Shen Z, Li Y, Guo Y, Feng Y, Sun S, Guo M, Hu Z, et al. Knocking Out OsAAP11 to Improve Rice Grain Quality Using CRISPR/Cas9 System. International Journal of Molecular Sciences. 2023; 24(18):14360. https://doi.org/10.3390/ijms241814360
Chicago/Turabian StyleYang, Yihao, Yi Zhang, Zixing Sun, Ziyan Shen, Youguang Li, Yifan Guo, Yuntong Feng, Shengyuan Sun, Min Guo, Zhi Hu, and et al. 2023. "Knocking Out OsAAP11 to Improve Rice Grain Quality Using CRISPR/Cas9 System" International Journal of Molecular Sciences 24, no. 18: 14360. https://doi.org/10.3390/ijms241814360
APA StyleYang, Y., Zhang, Y., Sun, Z., Shen, Z., Li, Y., Guo, Y., Feng, Y., Sun, S., Guo, M., Hu, Z., & Yan, C. (2023). Knocking Out OsAAP11 to Improve Rice Grain Quality Using CRISPR/Cas9 System. International Journal of Molecular Sciences, 24(18), 14360. https://doi.org/10.3390/ijms241814360