
Physiological and Morphological Implications of Using Composts with Different Compositions in the Production of Cucumber Seedlings

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characteristics of Physical and Chemical Properties of Substrates
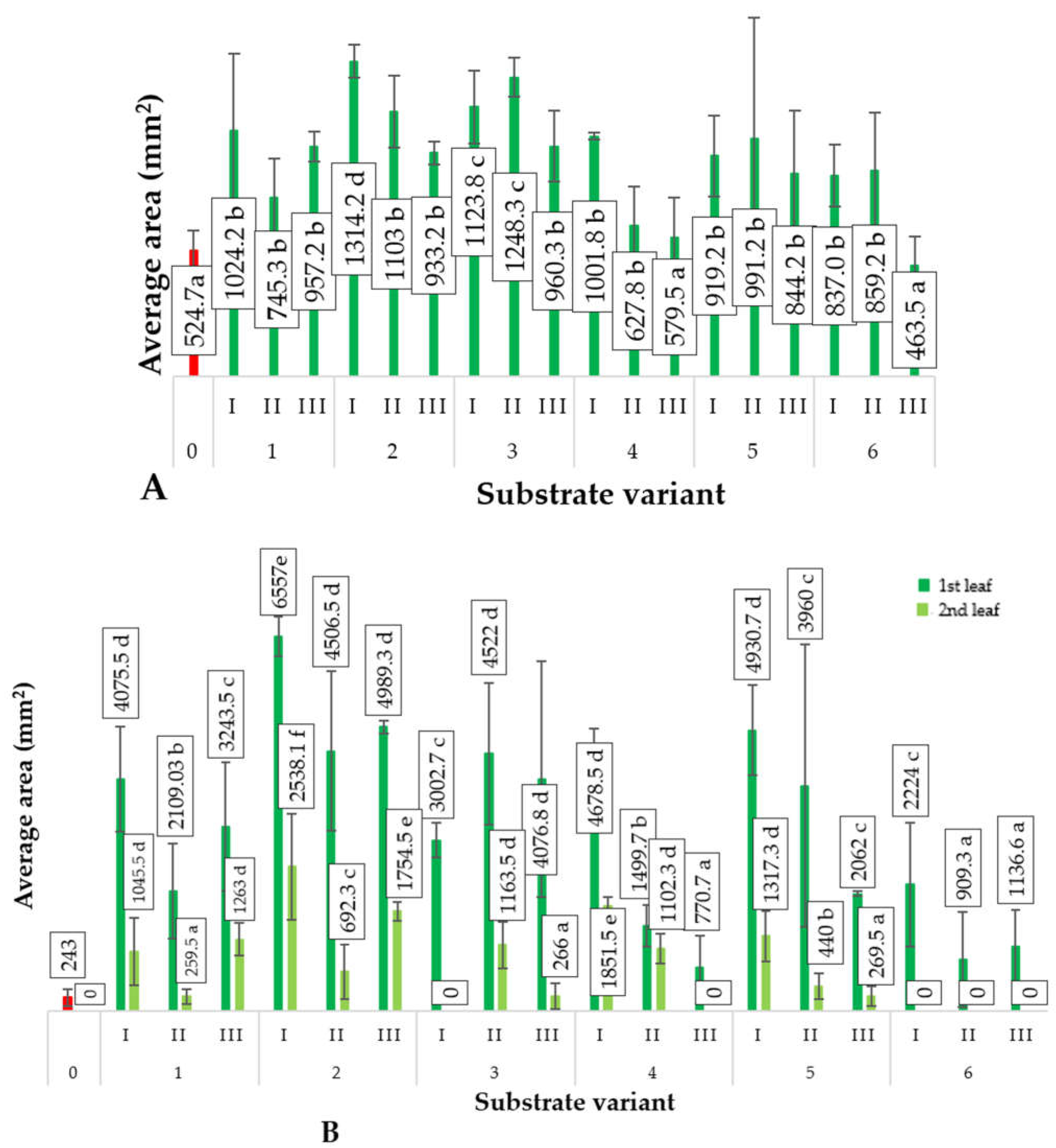
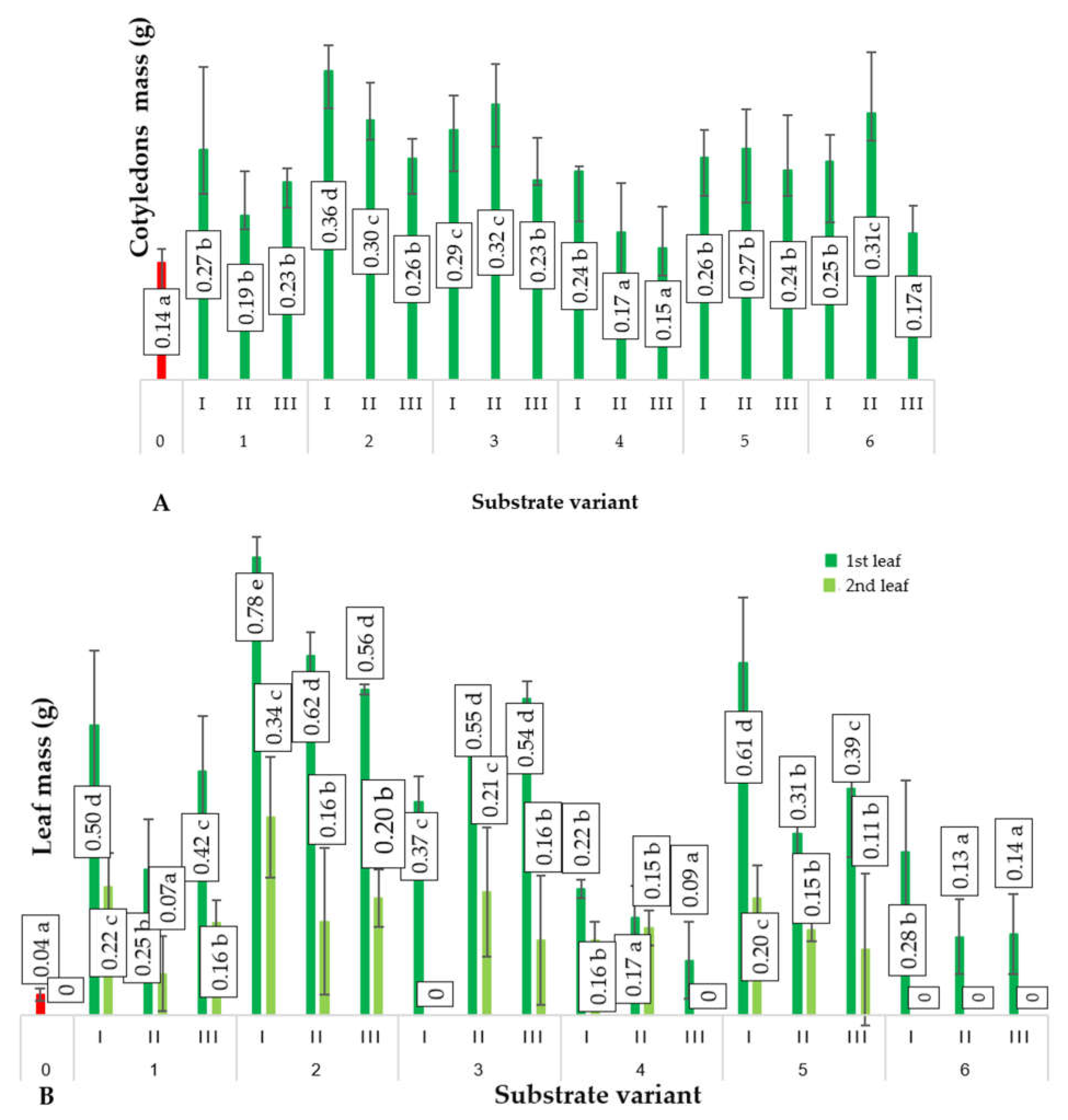
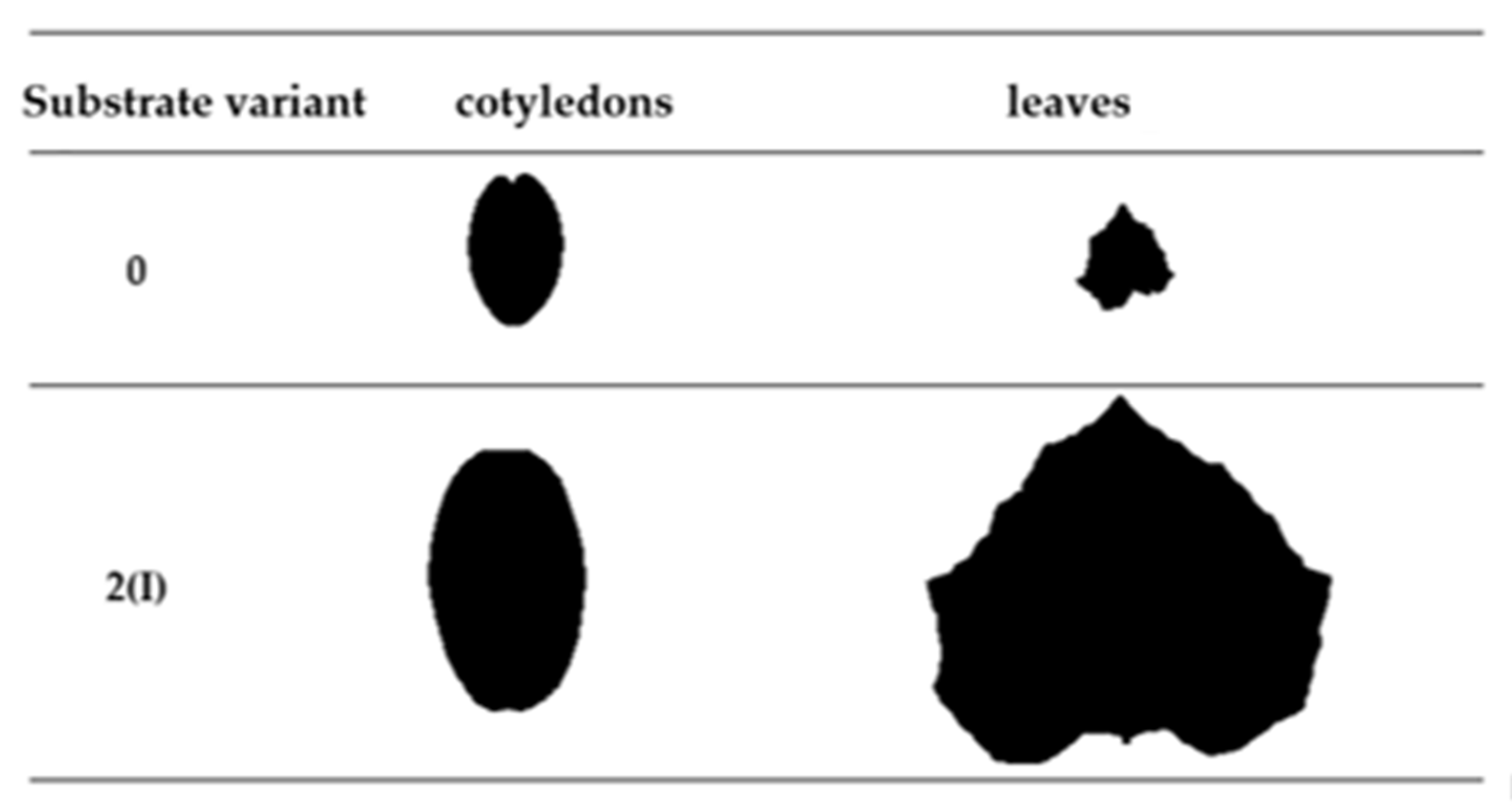
2.2. Biometric Parameters of Plants
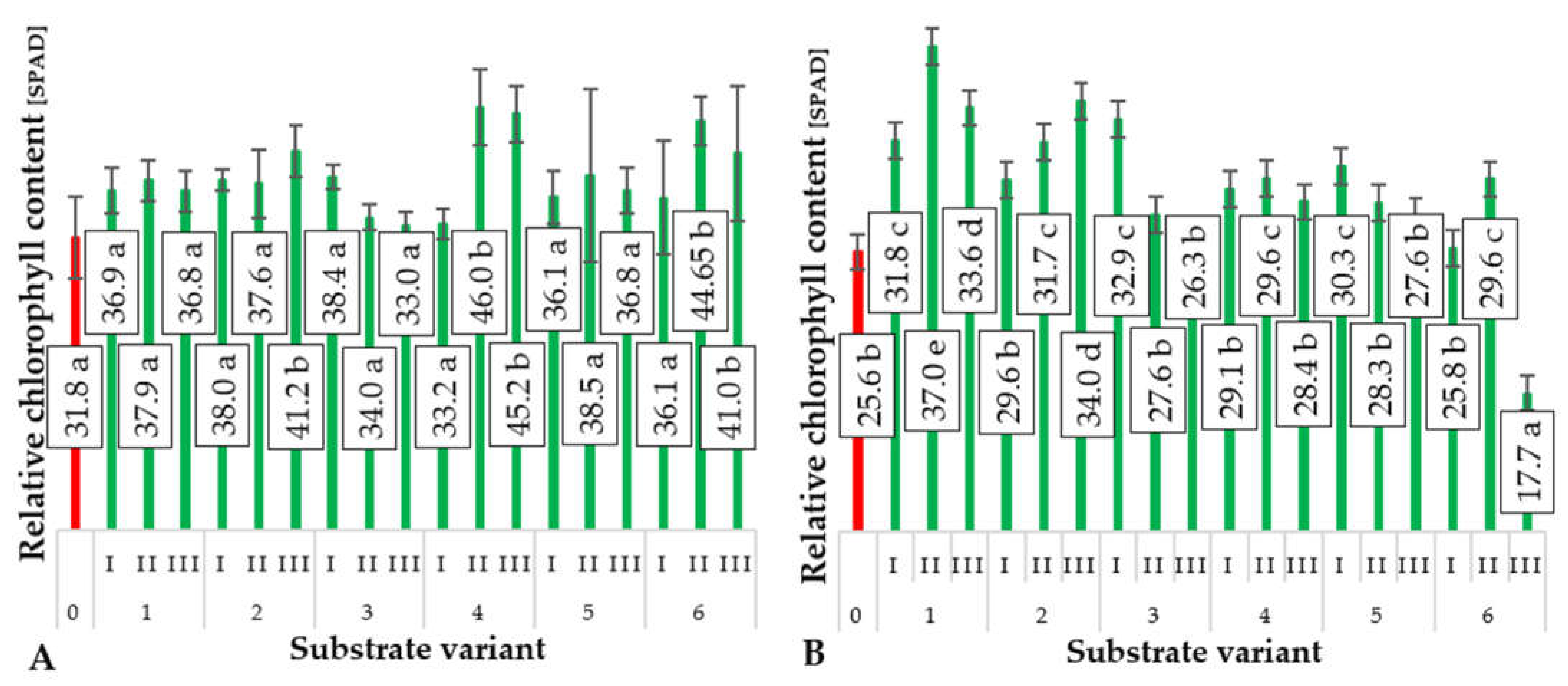
2.3. Relative Chlorophyll Content (SPAD)
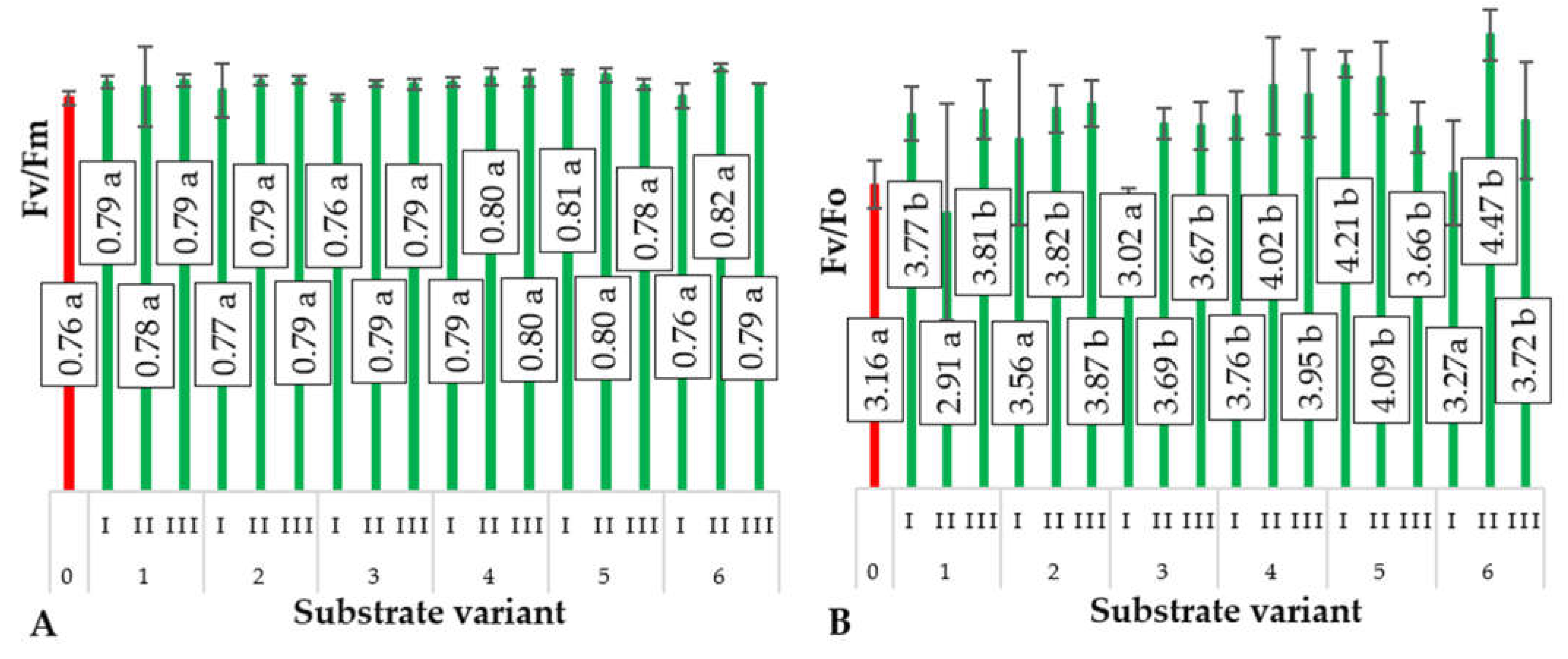
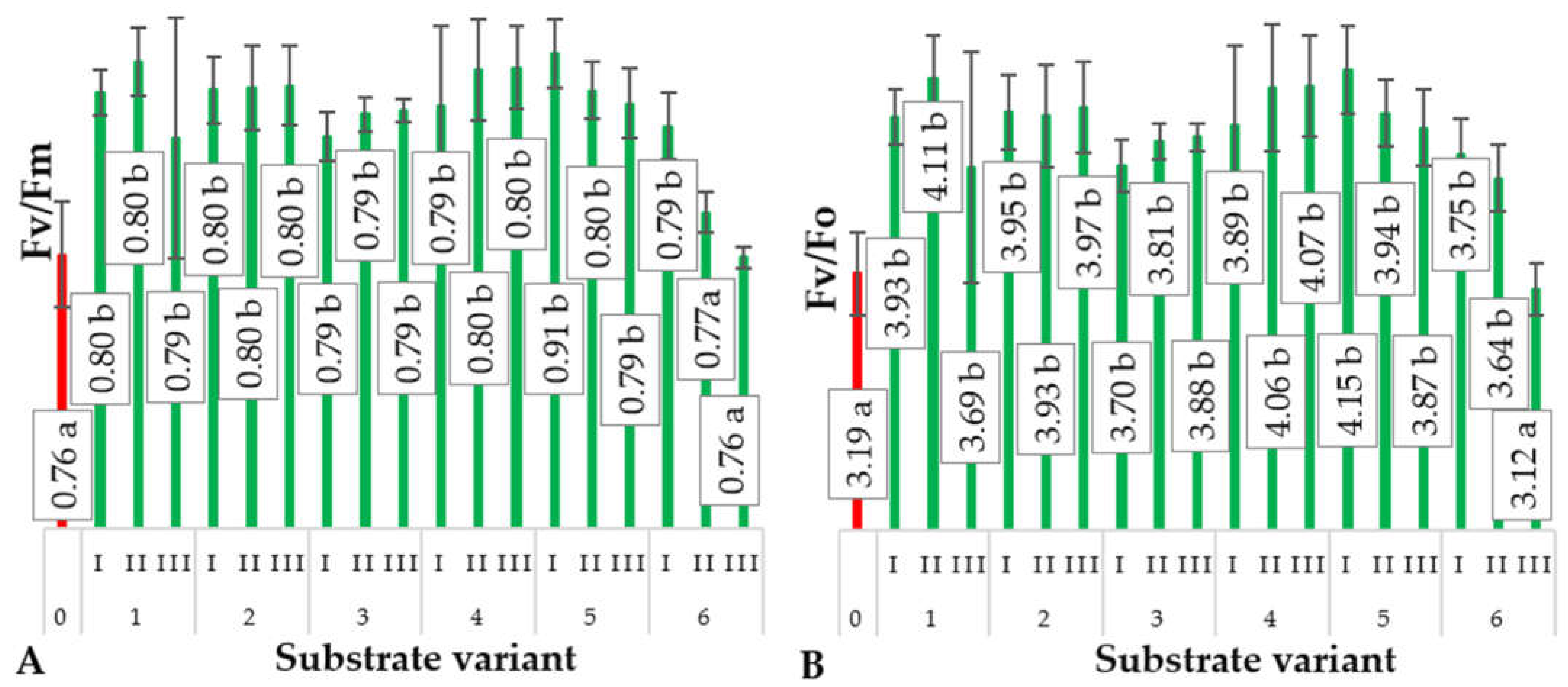
2.4. Chlorophyll Fluorescence Parameters
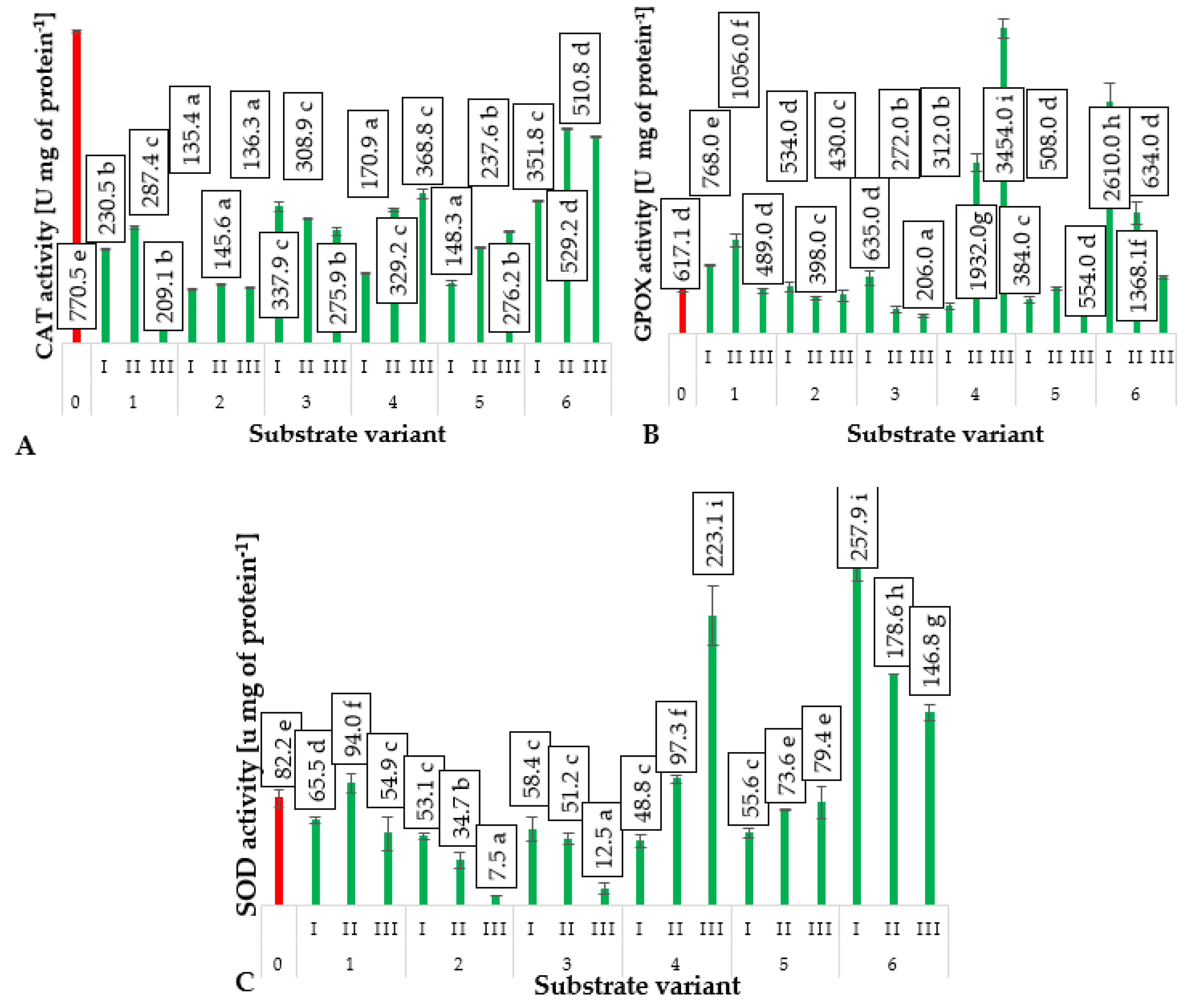
2.5. Antioxidant Enzymes Activity
3. Materials and Methods
3.1. Seed Material
3.2. Substrate Preparation Procedure
- A.
- Sewage sludge (80%) + Sawdust (20%);
- B.
- Sewage sludge (40%) + Sawdust (10%) + Biodegradable garden and park waste (50%);
- C.
- Biodegradable garden and park waste (90%) + Sawdust (10%);
- D.
- Sewage sludge (80%) + Sawdust (20%) + Eisenia fetida;
- E.
- Sewage sludge (40%) + Sawdust10%) + Biodegradable garden and park waste (50%) + Eisenia fetida;
- F.
- Biodegradable garden and park waste (90%) + Sawdust (10%) + Eisenia fetida.
- 1.
- (I): A (75%) + O(25%); (II): A (50%) + O (50%); (III): A(25%) + O (75%);
- 2.
- (I): B (75%) + O(25%); (II): B (50%) + O (50%); (III): B(25%) + O (75%);
- 3.
- (I): C (75%) + O(25%); (II): C (50%) + O (50%); (III): C(25%) + O (75%);
- 4.
- (I): D (75%) + O(25%); (II): D (50%) + O (50%); (III): D(25%) + O (75%);
- 5.
- (I): E (75%) + O(25%); (II): E (50%) + O (50%); (III): E(25%) + O (75%);
- 6.
- (I): F (75%) + O(25%); (II): F (50%) + O (50%); (III): F(25%) + O (75%).
3.3. Experimental Design
3.4. Analysis of Physicochemical Properties of Substrates
3.5. Physiological Parameters of Plants
3.5.1. Relative Chlorophyll Content
3.5.2. Chlorophyll Fluorescence
3.6. Biometric Parameters of Cotyledons and Leaves
3.7. Biochemical Analyzes of Plant Biomass
3.7.1. Enzymes Extraction
3.7.2. Activity of Superoxide Dismutase, Catalase and Guaiacol Peroxidase
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the performance of cucumber (Cucumis sativus L.) seedlings by LED illumination. Sci. Rep. 2022, 12, 852. [Google Scholar] [CrossRef]
- Sawan, O.M.; Eissa, A.M.; Abou-Hadid, A.F. The effect of different growing media on cucumber seedling production, fruit yield and quality under greenhouse conditions. Int. Symp. Greenh. Manag. Better Yield Qual. Mild Winter Clim. 1997, 491, 369–376. [Google Scholar] [CrossRef]
- Sallaku, G.; Babaj, I.; Kaciu, S.; Balliu, A. The influence of vermicompost on plant growth characteristics of cucumber (Cucumis sativus L.) seedlings under saline conditions. J. Food Agric. Environ. 2009, 7, 869–872. [Google Scholar]
- Abdel-Razzak, H.; Alkoaik, F.; Rashwan, M.; Fulleros, R.; Ibrahim, M. Tomato waste compost as an alternative substrate to peat moss for the production of vegetable seedlings. J. Plant Nutr. 2019, 42, 287–295. [Google Scholar] [CrossRef]
- Bayoumi, Y.A.; El-Henawy, A.S.; Abdelaal, K.A.; Elhawat, N. Grape fruit waste compost as a nursery substrate ingredient for high-quality cucumber (Cucumis sativus L.) seedlings production. Compos. Sci. Util. 2019, 27, 205–216. [Google Scholar] [CrossRef]
- Szostek, M.; Kosowski, P.; Szpunar-Krok, E.; Jańczak-Pieniążek, M.; Matłok, N.; Skrobacz, K.; Pieniążek, R.; Balawejder, M. The Usefulness of Ozone-Stabilized Municipal Sewage Sludge for Fertilization of Maize (Zea mays L.). Agriculture 2022, 12, 387. [Google Scholar] [CrossRef]
- Koutroubas, S.D.; Antoniadis, V.; Damalas, C.A.; Fotiadis, S. Municipal Sewage Sludge Effects on Maize Yield, Nitrogen Use Efficiency, and Soil Properties. J. Soil Sci. Plant Nutr. 2023, 23, 1209–1221. [Google Scholar] [CrossRef]
- Usman, K.; Khan, S.; Ghulam, S.; Khan, M.U.; Khan, N.; Khan, M.A.; Khalil, S.K. Sewage Sludge: An Important Biological Resource for Sustainable Agriculture and Its Environmental Implications. Am. J. Plant Sci. 2012, 3, 1708–1721. [Google Scholar] [CrossRef]
- Singh, R.; Agrawal, M. Effects of sewage sludge amendment on heavy metal accumulation and consequent responses of Beta vulgaris plants. Chemosphere 2007, 67, 2229–2240. [Google Scholar] [CrossRef]
- European Council. Council Directive 86/278/EEC of 12 June 1986 of the European Parliament and of the Council on the Protection of the Environment, and in Particular of the Soil, when Sewage Sludge Is Used in Agriculture; Official Journal of the European Communities: Luxembourg City, Luxembourg, 1986; p. 181.
- Herrera, F.; Castillo, J.E.; Chica, A.F.; López Bellido, L. Use of municipal solid waste compost (MSWC) as a growing medium in the nursery production of tomato plants. Bioresour. Technol. 2008, 99, 287–296. [Google Scholar] [CrossRef]
- Hicklenton, P.R.; Rodd, V.; Warman, P.R. The effectiveness and consistency of source-separated municipal solid waste and bark composts as components of container growing media. Sci. Hortic. 2001, 91, 365–378. [Google Scholar] [CrossRef]
- Stewart-Wade, S.M. Efficacy of organic amendments used in containerized plant production: Part 1—Compost-based amendments. Sci. Hortic. 2020, 266, 108856. [Google Scholar] [CrossRef]
- Act of 14 December 2012. On waste. Journal of Laws 2013, item 21. In polish: Dz. U. 2013 poz. 21—Ustawa o odpadach z dnia 14 grudnia 2012. Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20130000021/T/D20130021L.pdf (accessed on 29 June 2023).
- Regulation of the Minister of the Environment of 1 August 2002 on Municipal Sewage Sludge. Journal of Laws 2002, item. 1140. In polish: Dz. U. 2002 nr 134 poz. 1140—Rozporządzenie Ministra Środowiska z dnia 1 sierpnia 2002 r. w sprawie komunalnych osadów ściekowych. Available online: https://isap.sejm.gov.pl/isap.nsf/download.xsp/WDU20021341140/O/D20021140.pdf (accessed on 29 June 2023).
- Wulff, R.D. Seed Size Variation in Desmodium Paniculatum: II. Effects on Seedling Growth and Physiological Performance. J. Ecol. 1986, 74, 99–114. [Google Scholar] [CrossRef]
- Zhong, Z.; Bian, F.; Zhang, X. Testing composted bamboo residues with and without added effective microorganisms as a renewable alternative to peat in horticultural production. Ind. Crop. Prod. 2018, 112, 602–607. [Google Scholar] [CrossRef]
- Mohammed, S.J.; Alkobaisy, J.S.; Saleh, J.M. Effect of Earthworm on Lettuce Production through the Recycling of Organic and Bio-Compost Production. Asian Soil Res. J. 2019, 2, 42411. [Google Scholar] [CrossRef]
- Enebe, M.C.; Erasmus, M. Vermicomposting technology—A perspective on vermicompost production technologies, limitations and prospects. J. Environ. Manag. 2023, 345, 118585. [Google Scholar] [CrossRef]
- Ratnasari, A.; Syafiuddin, A.; Mehmood, M.A.; Boopathy, R. A review of the vermicomposting process of organic and inorganic waste in soils: Additives effects, bioconversion process, and recommendations. Bioresour. Technol. Rep. 2023, 21, 101332. [Google Scholar] [CrossRef]
- Kováčik, P.; Šimanský, V.; Smoleń, S.; Neupauer, J.; Olšovská, K. The Effect of Vermicompost and Earthworms (Eisenia fetida) Application on Phytomass and Macroelement Concentration and Tetanic Ratio in Carrot. Agronomy 2022, 12, 2770. [Google Scholar] [CrossRef]
- Kováčik, P.; Olšovská, K.; Smoleń, S.; Smoleń-Ledwożyw, I. Parameters of radish phytomass (Raphanus sativus L.) determined by vermicompost and earthworms (Eisenia fetida). Folia Hortic. 2021, 33, 217–233. [Google Scholar] [CrossRef]
- Argenta, G.; Silva, P.R.F.D.; Sangoi, L. Leaf relative chlorophyll content as an indicator parameter to predict nitrogen fertilization in maize. Ciência Rural. 2004, 34, 1379–1387. [Google Scholar] [CrossRef]
- Matłok, N.; Basara, O.; Zardzewiały, M.; Gorzelany, J.; Balawejder, M. Effectiveness of a Complex Fertilisation Technology Applied to Zea mays, Assessed Based on Normalised Difference Vegetation Index (NDVI) from Terra Moderate Resolution Imaging Spectroradiometer (MODIS). Agriculture 2021, 11, 754. [Google Scholar] [CrossRef]
- Usuda, H.; Ku, M.S.B.; Edwards, G.E. Rates of photosynthesis relative to activity of photosynthetic enzymes, chlorophyll and soluble protein content among ten C4 species. Aust. J. Plant Physiol. 1984, 11, 509–517. [Google Scholar] [CrossRef]
- Matlok, N.; Piechowiak, T.; Gorzelany, J.; Zardzewiały, M.; Balawejder, M. Effect of Ozone Fumigation on Physiological Processes and Bioactive Compounds of Red-Veined Sorrel (Rumex sanguineus ssp sanguineus). Agronomy 2020, 10, 1726. [Google Scholar] [CrossRef]
- Griffin, K.L.; Seemann, J.R. Plants, CO2 and photosynthesis in the 21st century. Chem. Biol. 1996, 3, 245–254. [Google Scholar] [CrossRef]
- Dubey, R.S. Photosynthesis in plants under stressful conditions. In Handbook of Photosynthesis, 3rd ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 629–649. [Google Scholar] [CrossRef]
- Huang, Z.A.; Jiang, D.A.; Yang, Y.; Sun, J.W.; Jin, S.H. Effects of nitrogen deficiency on gas exchange, chlorophyll fluorescence, and antioxidant enzymes in leaves of rice plants. Photosynthetica 2004, 42, 357–364. [Google Scholar] [CrossRef]
- Matlok, N.; Szostek, M.; Antos, P.; Gajdek, G.; Gorzelany, J.; Bobrecka-Jamro, D.; Balawejder, M. Effect of Foliar and Soil Fertilization with New Products Based on Calcinated Bones on Selected Physiological Parameters of Maize Plants. Appl. Sci. 2020, 10, 2579. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Guo, P. Chlorophyll fluorescence: A usefull tool in barley plant breeding programs. In Photochemistry Research Progress; Sanchez, A., Gutierrez, S.J., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2008; pp. 439–463. ISBN 978-1-60456-568-3. [Google Scholar]
- Baker, N.R.; Oxborough, K. Chlorophyll Fluorescence as a Probe of Photosynthetic Productivity. In Chlorophyll a Fluorescence. Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 65–82. [Google Scholar] [CrossRef]
- Michałek, W.; Kocira, A.; Findura, P.; Szparaga, A.; Kocira, S. The Influence of Biostimulant Asahi SL on the Photosynthetic Activity of Selected Cultivars of Phaseolus vulgaris L. Annu. Set Environ. Prot. 2018, 20, 1286–1301. Available online: https://yadda.icm.edu.pl/baztech/element/bwmeta1.element.baztech-dce4de82-9c66-4f07-88c7-ad0e574bdf3a (accessed on 29 June 2023).
- Van Kooten, O.; Snel, J.F.H. The Use of Chlorophyll Fluorescence Nomenclature in Plant Stress Physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]
- Björkman, O. The effect of oxygen concentration on photosynthesis in higher plants. Physiol. Plant. 1966, 19, 618–633. [Google Scholar] [CrossRef]
- Matłok, N.; Piechowiak, T.; Kapusta, I.; Królikowski, K.; Balawejder, M. Induction of Biosynthesis Antioxidant Molecules in Young Barley Plants by Trioxygen. Molecules 2022, 27, 7195. [Google Scholar] [CrossRef]
- Slesak, I.; Libik, M.; Karpinska, B.; Karpinski, S.; Miszalski, Z. The role of hydrogen peroxide in regulation of plant metabolism and cellular signalling in response to environmental stresses. Acta Biochim. Pol. 2007, 54, 39–50. Available online: https://pubmed.ncbi.nlm.nih.gov/17325747/ (accessed on 29 June 2023). [CrossRef] [PubMed]
- Luna, C.M.; Pastori, G.M.; Driscoll, S.; Groten, K.; Bernard, S.; Foyer, C.H. Drought controls on H2O2 accumulation, catalase (CAT) activity and CAT gene expression in wheat. J. Exp. Bot. 2005, 56, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Goggin, F.L.; Fischer, H.D. Reactive Oxygen Species in Plant Interactions with Aphids. Front. Plant Sci. 2022, 12, 811105. Available online: https://www.frontiersin.org/articles/10.3389/fpls.2021.811105/full (accessed on 29 June 2023). [CrossRef]
- Niu, L.; Liao, W. Hydrogen Peroxide Signaling in Plant Development and Abiotic Responses: Crosstalk with Nitric Oxide and Calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef]
- Krustev, S.; Angelova, V.; Ivanov, K.; Zaprajnova, P. Methods for preparation of soil samples for determination of trace elments. World Academy of Science, Engineering and Technolog. Int. J. Geol. Environ. Eng. 2017, 11, 7. [Google Scholar] [CrossRef]
- Egner, H.; Riehm, H.; Domingo, W.R. Untersuchungen über die chemische Bodenanalyse als Grundlage für die Beurteilung des Nahrstoffzustandes der Boden, II: Chemische Extractionsmetoden zu Phosphor-und Kaliumbestimmung. K. Lantbrukshügskolans Ann. 1960, 26, 199–215. [Google Scholar]
- Balawejder, M.; Matłok, N.; Piechowiak, T.; Szostek, M.; Kapusta, I.; Niemiec, M.; Komorowska, M.; Wróbel, M.; Mudryk, K.; Szeląg-Sikora, A.; et al. The Modification of Substrate in the Soilless Cultivation of Raspberries (Rubus idaeus L.) as a Factor Stimulating the Biosynthesis of Selected Bioactive Compounds in Fruits. Molecules 2023, 28, 118. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Variant | Compost Variant + Deacidified Peat (O) | pH | EC | TOC | Ntotal | C:N | Available Forms of Nutrients Calculated as Oxides | Total Forms of Nutrients | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P2O5 | K2O | Mg | Ca | ||||||||
| (%) | (H2O) | (µS cm−1) | (%) | (%) | (mg∙100 g−1) | (g∙kg−1) | |||||
| 0 | (Control) | O (100%) | 6.97 ± 0.21 | 136 ± 11 | 55.0 ± 2.6 | 2.30 ± 0.11 | 23.9 ± 1.21 | 46.9 ± 3.5 | 33.0 ± 2.7 | 1.37 ± 0.18 | 2.13 ± 0.11 |
| 1 | (I) | A (25%) + O (75%) | 6.63 ± 0.32 | 560 ± 24 | 52.1 ± 3.2 | 2.73 ± 0.17 | 19.1 ± 0.97 | 325 ± 11 | 123 ± 14 | 1.83 ± 0.23 | 5.85 ± 0.18 |
| (II) | A (50%) + O (50%) | 7.05 ± 0.41 | 774 ± 31 | 49.1 ± 2.9 | 3.16 ± 0.19 | 15.5 ± 0.86 | 650 ± 34 | 247 ± 23 | 2.29 ± 0.17 | 9.57 ± 0.64 | |
| (III) | A (75%) + O (25%) | 6.89 ± 0.37 | 1134 ± 38 | 46.2 ± 3.9 | 3.60 ± 0.09 | 12.8 ± 0.54 | 975 ± 45 | 370 ± 34 | 2.74 ± 0.21 | 13.28 ± 0.76 | |
| 2 | (I) | B (25%) + O (75%) | 6.85 ± 0.19 | 570 ± 17 | 47.5 ± 4.2 | 2.19 ± 0.13 | 21.7 ± 0.42 | 273 ± 27 | 172 ± 18 | 1.60 ± 0.16 | 5.40 ± 0.54 |
| (II) | B (50%) + O (50%) | 6.82 ± 0.25 | 988 ± 29 | 40.0 ± 1.9 | 2.06 ± 0.08 | 19.4 ± 0.38 | 546 ± 36 | 344 ± 26 | 1.84 ± 0.29 | 8.67 ± 0.48 | |
| (III) | B (75%) + O (25%) | 7.05 ± 0.26 | 1315 ± 28 | 32.4 ± 1.6 | 1.95 ± 0.14 | 16.6 ± 0.19 | 818 ± 43 | 516 ± 34 | 2.07 ± 0.18 | 11.93 ± 0.52 | |
| 3 | (I) | C (25%)+ O (75%) | 6.91 ± 0.36 | 556 ± 19 | 46.8 ± 2.4 | 1.98 ± 0.17 | 23.6 ± 0.21 | 50 ± 2.6 | 116 ± 12 | 1.48 ± 0.24 | 6.47 ± 0.37 |
| (II) | C (50%)+ O (50%) | 7.08 ± 0.29 | 1104 ± 21 | 38.5 ± 2.9 | 1.66 ± 0.12 | 23.2 ± 0.26 | 101 ± 12 | 232 ± 27 | 1.59 ± 0.16 | 10.82 ± 0.46 | |
| (III) | C (75%)+ O (25%) | 7.28 ± 0.29 | 1469 ± 32 | 30.3 ± 1.7 | 1.34 ± 0.11 | 22.6 ± 0.27 | 151 ± 16 | 347 ± 36 | 1.69 ± 0.19 | 15.16 ± 0.57 | |
| 4 | (I) | D (25%)+ O (75%) | 6.27 ± 0.17 | 1398 ± 36 | 51.8 ± 2.4 | 2.38 ± 0.28 | 21.8 ± 0.19 | 236 ± 32 | 127 ± 16 | 1.88 ± 0.11 | 6.92 ± 0.34 |
| (II) | D (50%)+ O (50%) | 6.09 ± 0.11 | 1552 ± 27 | 48.6 ± 2.9 | 2.45 ± 0.20 | 19.8 ± 0.25 | 471 ± 43 | 255 ± 21 | 2.39 ± 0.27 | 11.72 ± 0.47 | |
| (III) | D (75%) + O (25%) | 5.90 ± 0.10 | 1958 ± 32 | 45.4 ± 3.2 | 2.52 ± 0.19 | 18.0 ± 0.34 | 707 ± 25 | 382 ± 31 | 2.89 ± 0.31 | 16.51 ± 0.57 | |
| 5 | (I) | E (25%) + O (75%) | 6.69 ± 0.23 | 465 ± 18 | 47.0 ± 2.8 | 2.20 ± 0.10 | 21.4 ± 0.19 | 222 ± 10 | 191 ± 23 | 1.78 ± 0.18 | 5.70 ± 0.32 |
| (II) | E (50%) + O (50%) | 6.90 ± 0.28 | 1106 ± 24 | 39.0 ± 2.9 | 2.08 ± 0.09 | 18.8 ± 0.18 | 445 ± 31 | 382 ± 34 | 2.19 ± 0.24 | 9.27 ± 0.31 | |
| (III) | E (75%) + O (25%) | 6.59 ± 0.17 | 1875 ± 29 | 31.0 ± 2.5 | 1.98 ± 0.13 | 15.7 ± 0.23 | 667 ± 45 | 572 ± 40 | 2.59 ± 0.23 | 12.83 ± 0.48 | |
| 6 | (I) | F (25%) + O (75%) | 7.33 ± 0.25 | 552 ± 17 | 48.1 ± 3.1 | 2.12 ± 0.15 | 22.7 ± 0.31 | 91 ± 11 | 182 ± 16 | 2.30 ± 0.17 | 7.45 ± 0.47 |
| (II) | F (50%) + O (50%) | 7.46 ± 0.23 | 863 ± 21 | 41.2 ± 1.7 | 1.93 ± 0.17 | 21.3 ± 0.19 | 182 ± 17 | 364 ± 21 | 3.24 ± 0.32 | 12.77 ± 0.38 | |
| (III) | F (75%) + O (25%) | 7.87 ± 0.31 | 1680 ± 29 | 34.3 ± 1.9 | 1.75 ± 0.23 | 19.6 ± 0.16 | 273 ± 23 | 546 ± 28 | 4.17 ± 0.29 | 18.08 ± 0.49 | |
| Substrate Variant | Compost variant + Deacidified Peat (O) | Pb | Cr | Cu | Ni | Cd | Zn | Hg | |
|---|---|---|---|---|---|---|---|---|---|
| (%) | (mg∙kg−1) | ||||||||
| 0 | (Control) | O (100%) | 1.23 ± 0.11 | 3.45 ± 0.18 | 12.35 ± 1.12 | 1.12 ± 0.09 | 0.05 ± 0.00 | 127 ± 15 | 0.01 ± 0.00 |
| 1 | (I) | A (25%) + O (75%) | 3.42 ± 0.12 | 5.11 ± 0.36 | 26.76 ± 3.21 | 2.07 ± 0.76 | 0.15 ± 0.01 | 159 ± 19 | 0.04 ± 0.00 |
| (II) | A (50%) + O (50%) | 5.62 ± 0.09 | 6.78 ± 0.46 | 41.18 ± 4.54 | 3.01 ± 0.43 | 0.25 ± 0.02 | 190 ± 21 | 0.07 ± 0.01 | |
| (III) | A (75%) + O (25%) | 7.81 ± 0.24 | 8.44 ± 0.73 | 55.59 ± 3.24 | 3.96 ± 0.23 | 0.34 ± 0.01 | 222 ± 32 | 0.10 ± 0.02 | |
| 2 | (I) | B (25%) + O (75%) | 3.42 ± 0.31 | 5.06 ± 0.68 | 23.76 ± 2.77 | 2.04 ± 0.19 | 0.14 ± 0.00 | 151 ± 17 | 0.03 ± 0.00 |
| (II) | B (50%) + O (50%) | 4.57 ± 0.29 | 6.66 ± 0.54 | 35.18 ± 3.46 | 2.96 ± 0.16 | 0.24 ± 0.01 | 175 ± 21 | 0.06 ± 0.01 | |
| (III) | B (75%) + O (25%) | 6.23 ± 0.32 | 8.27 ± 0.67 | 46.59 ± 5.23 | 3.88 ± 0.21 | 0.33 ± 0.03 | 199 ± 19 | 0.08 ± 0.01 | |
| 3 | (I) | C (25%)+ O (75%) | 2.87 ± 0.16 | 5.59 ± 0.48 | 13.26 ± 2.09 | 2.17 ± 0.11 | 0.12 ± 0.00 | 118 ± 10 | 0.01 ± 0.00 |
| (II) | C (50%)+ O (50%) | 4.52 ± 0.19 | 7.73 ± 0.38 | 14.18 ± 1.97 | 3.21 ± 0.09 | 0.20 ± 0.01 | 109 ± 11 | 0.02 ± 0.00 | |
| (III) | C (75%)+ O (25%) | 6.16 ± 0.32 | 9.86 ± 1.09 | 15.09 ± 1.34 | 4.26 ± 0.23 | 0.27 ± 0.03 | 100 ± 9 | 0.02 ± 0.00 | |
| 4 | (I) | D (25%)+ O (75%) | 2.82 ± 0.14 | 5.59 ± 0.83 | 30.26 ± 3.45 | 2.04 ± 0.24 | 0.16 ± 0.03 | 164 ± 15 | 0.05 ± 0.00 |
| (II) | D (50%)+ O (50%) | 4.42 ± 0.17 | 7.73 ± 0.65 | 48.18 ± 2.78 | 2.96 ± 0.38 | 0.28 ± 0.04 | 201 ± 21 | 0.10 ± 0.01 | |
| (III) | D (75%) + O (25%) | 6.01 ± 0.24 | 9.86 ± 0.65 | 66.09 ± 5.67 | 3.88 ± 0.27 | 0.39 ± 0.02 | 237 ± 23 | 0.14 ± 0.02 | |
| 5 | (I) | E (25%) + O (75%) | 2.77 ± 0.16 | 5.34 ± 0.43 | 20.51 ± 1.45 | 1.97 ± 0.12 | 0.13 ± 0.01 | 153 ± 24 | 0.03 ± 0.00 |
| (II) | E (50%) + O (50%) | 4.32 ± 0.27 | 7.23 ± 0.37 | 28.68 ± 4.12 | 2.81 ± 0.32 | 0.22 ± 0.01 | 180 ± 19 | 0.04 ± 0.00 | |
| (III) | E (75%) + O (25%) | 5.86 ± 0.31 | 9.11 ± 0.98 | 36.84 ± 3.65 | 3.66 ± 0.31 | 0.30 ± 0.04 | 206 ± 21 | 0.06 ± 0.01 | |
| 6 | (I) | F (25%) + O (75%) | 2.87 ± 0.16 | 5.84 ± 0.56 | 13.26 ± 2.09 | 2.12 ± 0.27 | 0.13 ± 0.01 | 115 ± 19 | 0.02 ± 0.00 |
| (II) | F (50%) + O (50%) | 4.52 ± 0.39 | 8.23 ± 0.62 | 14.18 ± 2.34 | 3.11 ± 0.19 | 0.20 ± 0.02 | 103 ± 8 | 0.03 ± 0.00 | |
| (III) | F (75%) + O (25%) | 6.16 ± 0.24 | 10.61 ± 1.04 | 15.09 ± 1.98 | 4.11 ± 0.27 | 0.28 ± 0.04 | 90 ± 9 | 0.03 ± 0.00 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zapałowska, A.; Matłok, N.; Piechowiak, T.; Szostek, M.; Puchalski, C.; Balawejder, M. Physiological and Morphological Implications of Using Composts with Different Compositions in the Production of Cucumber Seedlings. Int. J. Mol. Sci. 2023, 24, 14400. https://doi.org/10.3390/ijms241814400
Zapałowska A, Matłok N, Piechowiak T, Szostek M, Puchalski C, Balawejder M. Physiological and Morphological Implications of Using Composts with Different Compositions in the Production of Cucumber Seedlings. International Journal of Molecular Sciences. 2023; 24(18):14400. https://doi.org/10.3390/ijms241814400
Chicago/Turabian StyleZapałowska, Anita, Natalia Matłok, Tomasz Piechowiak, Małgorzata Szostek, Czesław Puchalski, and Maciej Balawejder. 2023. "Physiological and Morphological Implications of Using Composts with Different Compositions in the Production of Cucumber Seedlings" International Journal of Molecular Sciences 24, no. 18: 14400. https://doi.org/10.3390/ijms241814400
APA StyleZapałowska, A., Matłok, N., Piechowiak, T., Szostek, M., Puchalski, C., & Balawejder, M. (2023). Physiological and Morphological Implications of Using Composts with Different Compositions in the Production of Cucumber Seedlings. International Journal of Molecular Sciences, 24(18), 14400. https://doi.org/10.3390/ijms241814400