

ETHE1 Accelerates Triple-Negative Breast Cancer Metastasis by Activating GCN2/eIF2α/ATF4 Signaling
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
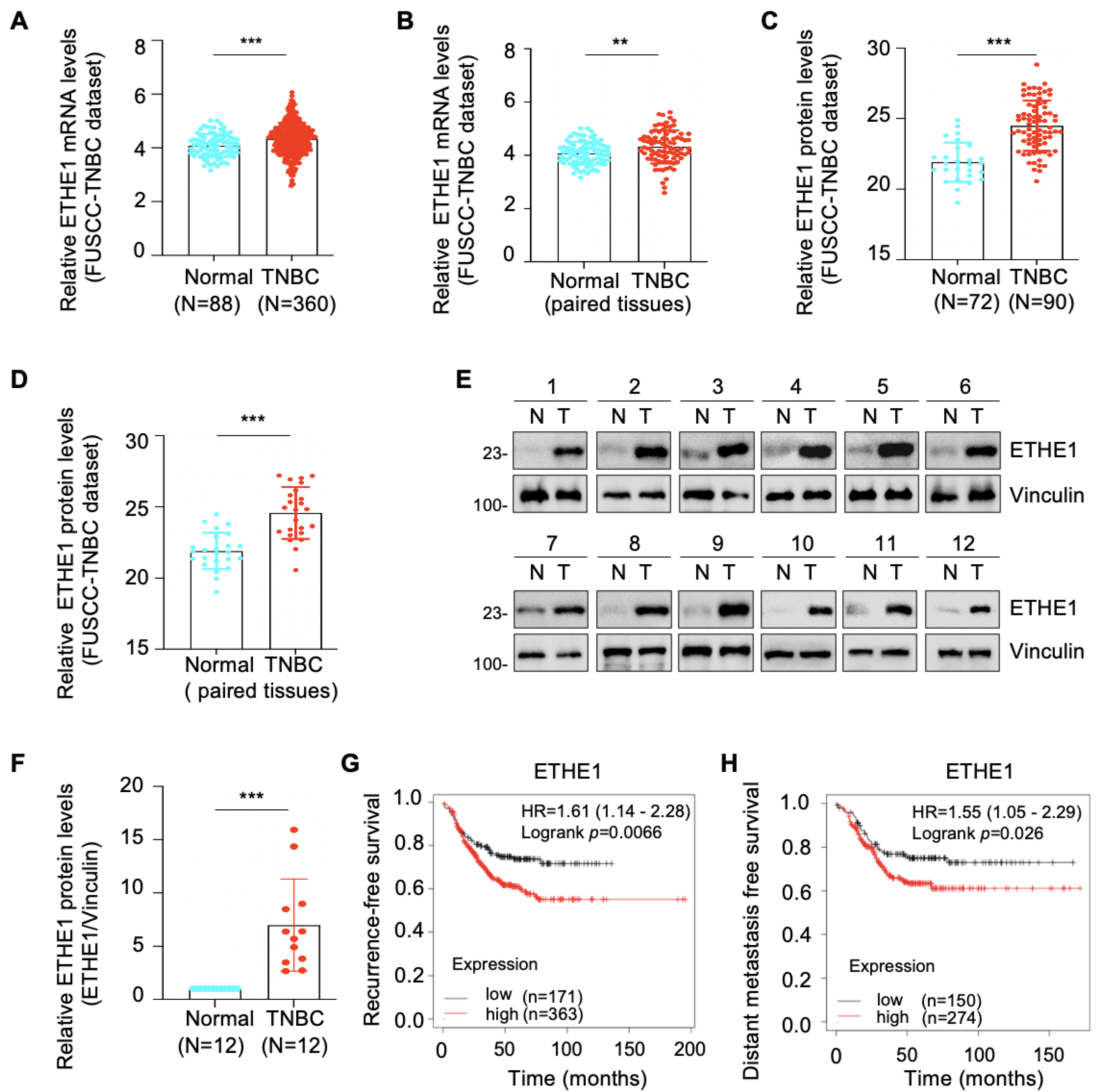
2.1. ETHE1 Is Upregulated in TNBC Tissues and Its High Expression Predicts Poor TNBC Prognosis
2.2. ETHE1 Does Not Affect TNBC Cell Proliferation or Xenograft Tumor Growth
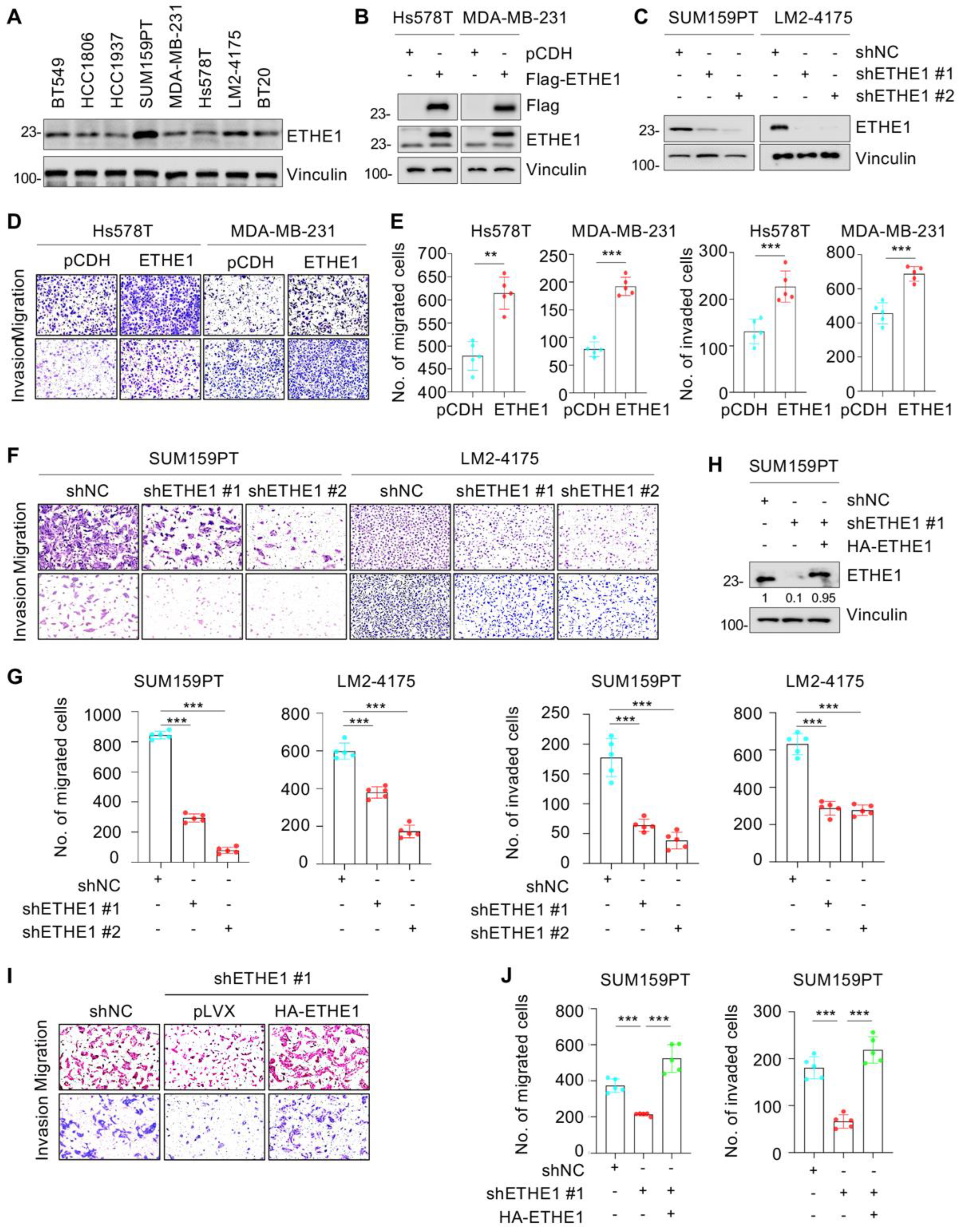
2.3. ETHE1 Promotes TNBC Cell Migration and Invasion, and the Noted Effects Are Independent of Its Enzymic Activity
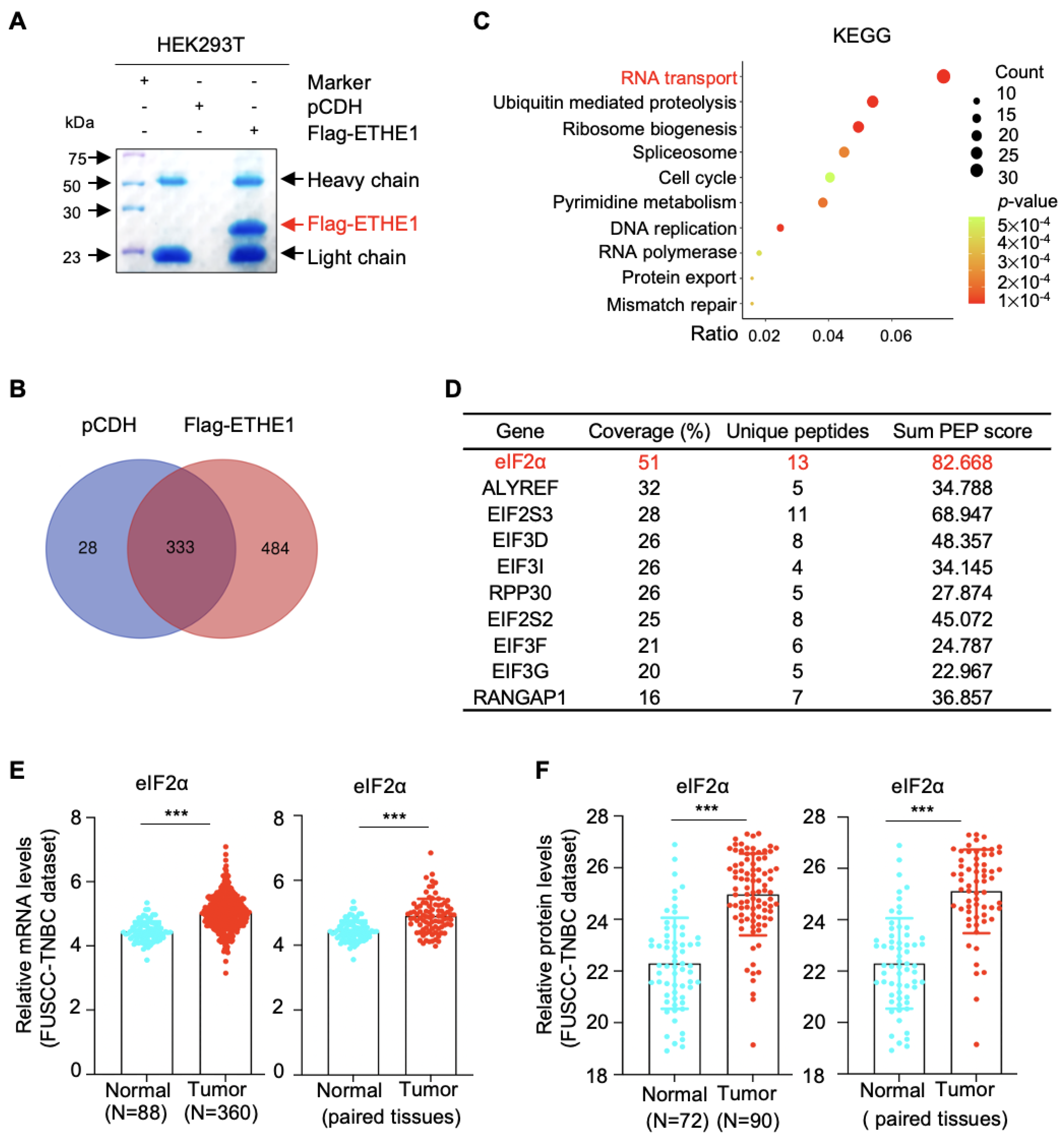
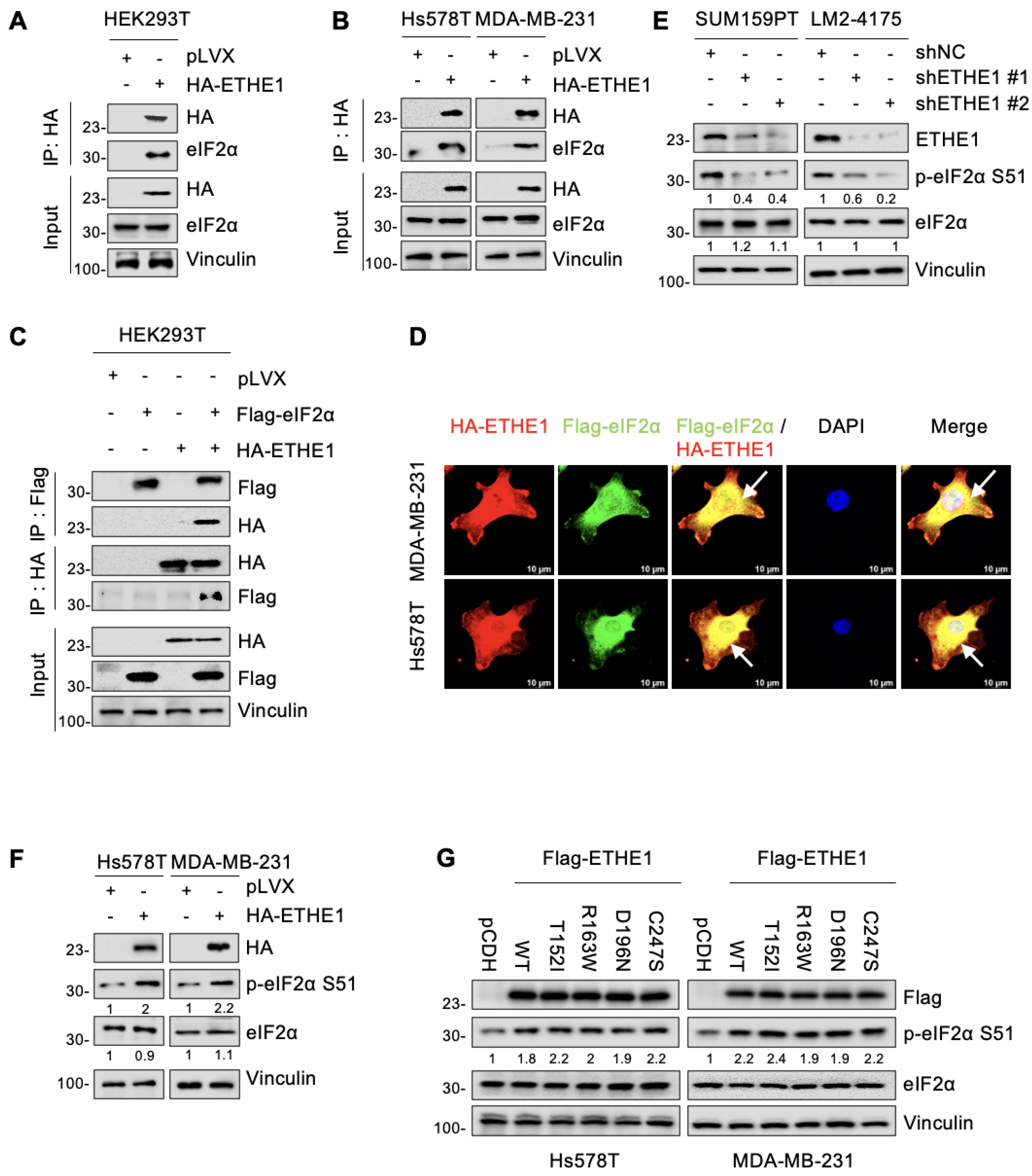
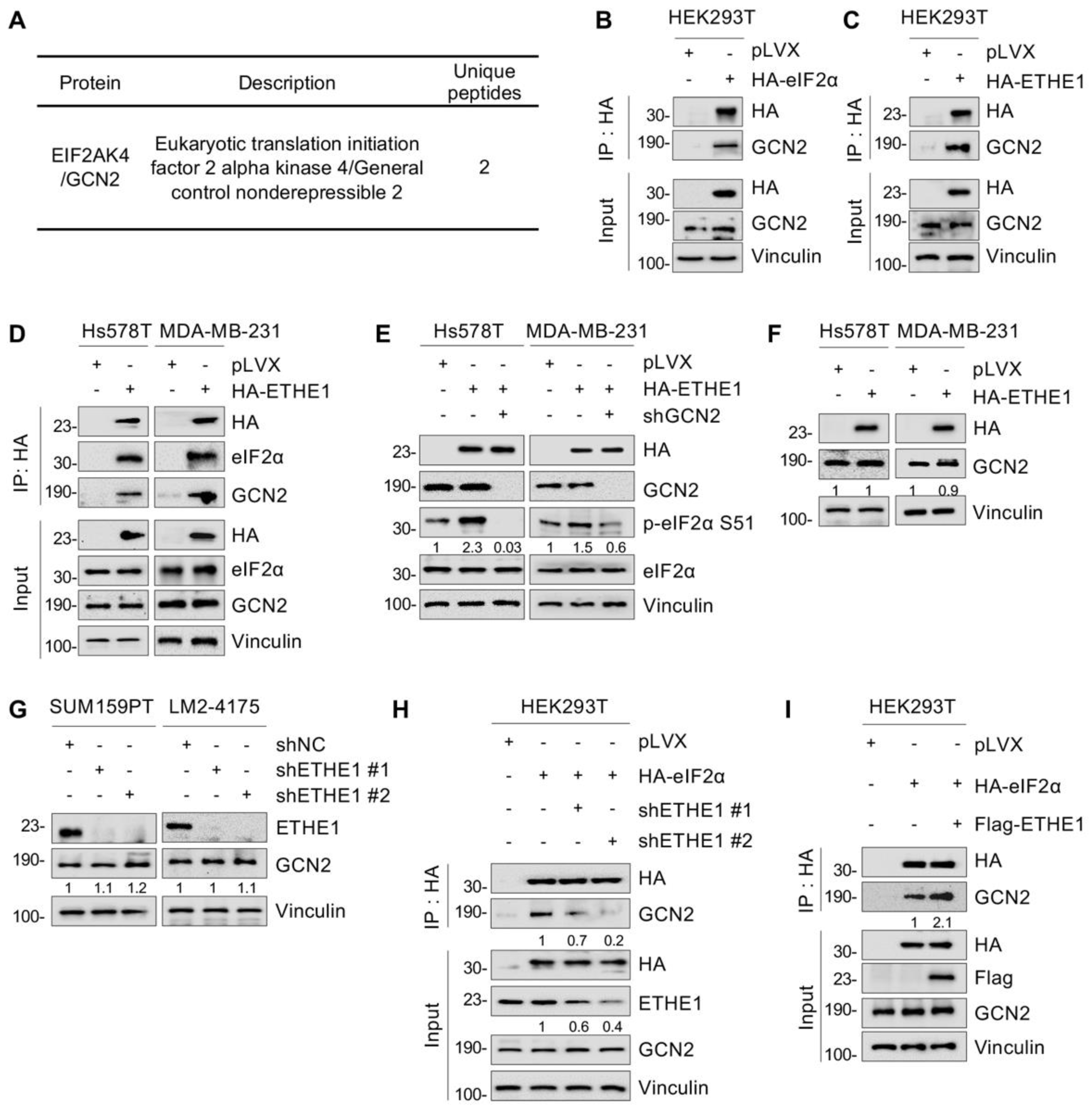
2.4. ETHE1 Interacts with eIF2α
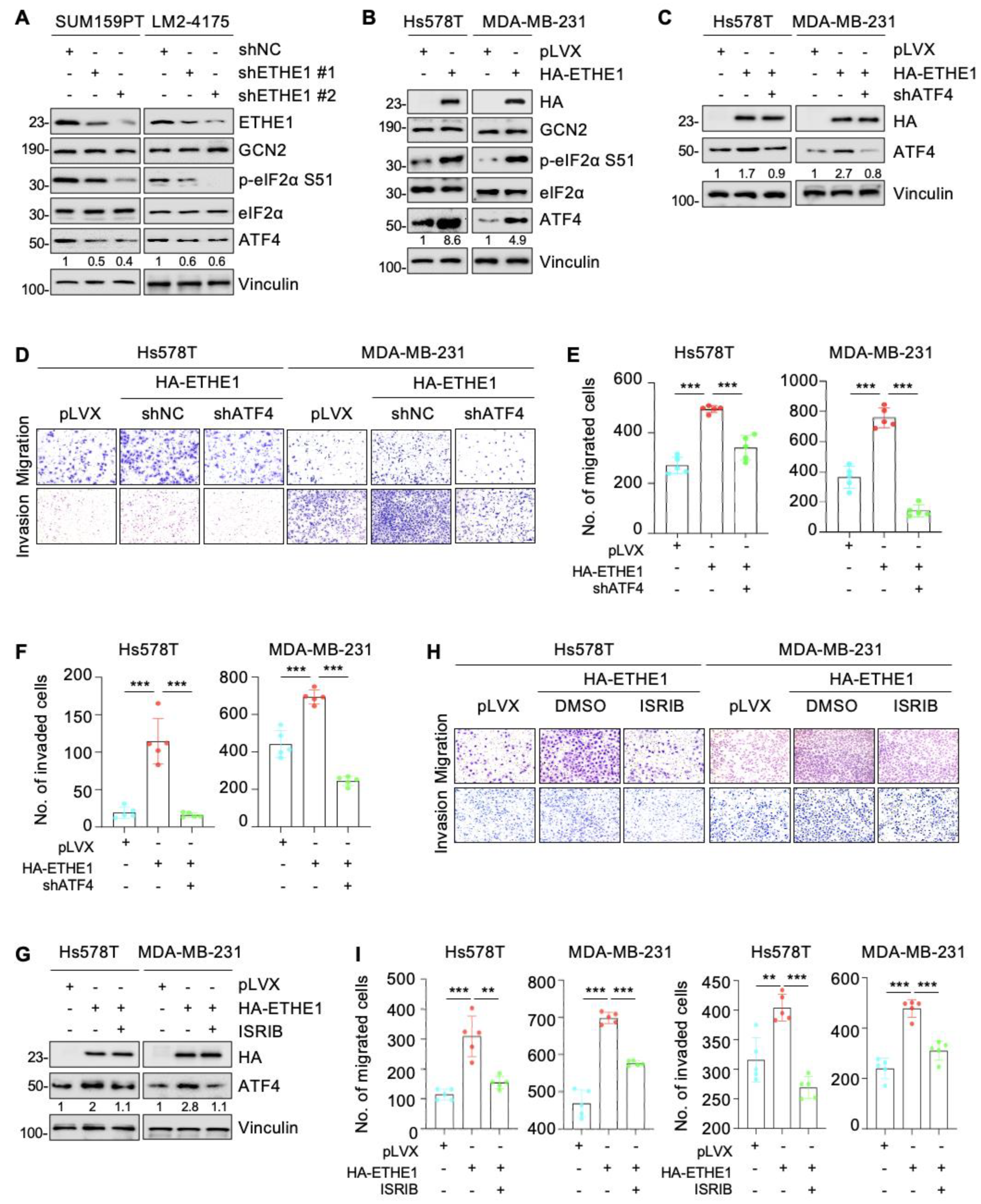
2.5. ETHE1 Enhances eIF2α Phosphorylation
2.6. ETHE1 Enhances eIF2α Phosphorylation by Promoting the Interaction between eIF2α and GCN2
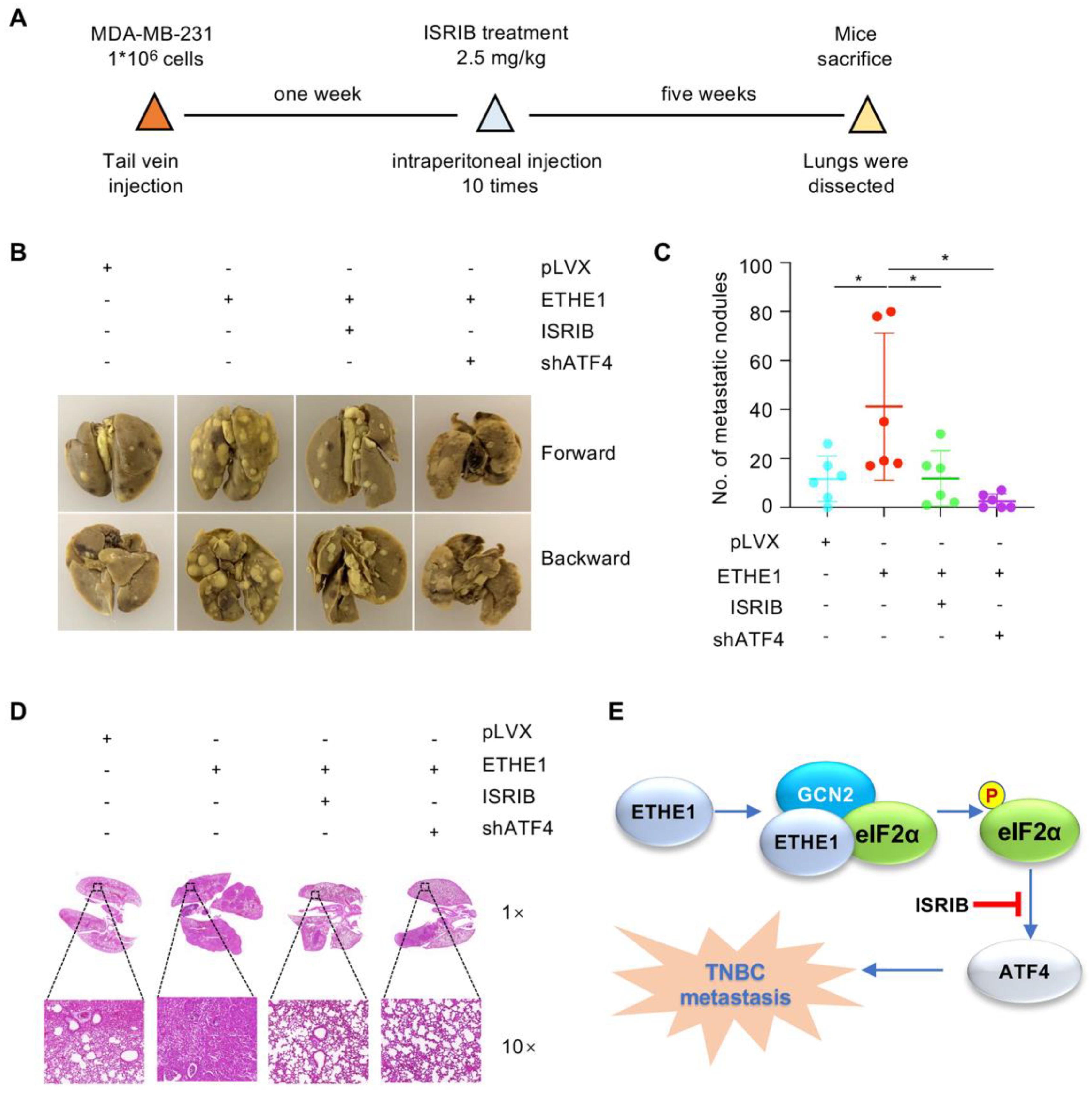
2.7. ETHE1 Promotes Lung Metastasis of TNBC Cells through eIF2α Phosphorylation–ATF4 Signaling
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals
4.2. Clinical Samples and Data
4.3. Expression Vectors
4.4. Plasmid Transfection and Lentiviral Infection
4.5. Antibodies, Immunoblotting, and Immunoprecipitation (IP)
4.6. Immunofluorescence
4.7. Cell Viability and Colony Formation Assays
4.8. Cell Migration and Invasion Assays
4.9. Proteomic Analysis
4.10. Xenograft Tumors in Nude Mice
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. Triple-negative breast cancer: Challenges and opportunities of a heterogeneous disease. Nat. Rev. Clin. Oncol. 2016, 13, 674–690. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; De Angelis, C.; Licata, L.; Gianni, L. Treatment landscape of triple-negative breast cancer—Expanded options, evolving needs. Nat. Rev. Clin. Oncol. 2022, 19, 91–113. [Google Scholar] [CrossRef]
- Pettinati, I.; Brem, J.; Lee, S.Y.; McHugh, P.J.; Schofield, C.J. The Chemical Biology of Human Metallo-beta-Lactamase Fold Proteins. Trends Biochem. Sci. 2016, 41, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Higashitsuji, H.; Higashitsuji, H.; Masuda, T.; Liu, Y.; Itoh, K.; Fujita, J. Enhanced deacetylation of p53 by the anti-apoptotic protein HSCO in association with histone deacetylase 1. J. Biol. Chem. 2007, 282, 13716–13725. [Google Scholar] [CrossRef]
- Tiranti, V.; Viscomi, C.; Hildebrandt, T.; Di Meo, I.; Mineri, R.; Tiveron, C.; Levitt, M.D.; Prelle, A.; Fagiolari, G.; Rimoldi, M.; et al. Loss of ETHE1, a mitochondrial dioxygenase, causes fatal sulfide toxicity in ethylmalonic encephalopathy. Nat. Med. 2009, 15, 200–205. [Google Scholar] [CrossRef]
- Higashitsuji, H.; Higashitsuji, H.; Nagao, T.; Nonoguchi, K.; Fujii, S.; Itoh, K.; Fujita, J. A novel protein overexpressed in hepatoma accelerates export of NF-kappa B from the nucleus and inhibits p53-dependent apoptosis. Cancer Cell 2002, 2, 335–346. [Google Scholar] [CrossRef]
- Grings, M.; Seminotti, B.; Karunanidhi, A.; Ghaloul-Gonzalez, L.; Mohsen, A.W.; Wipf, P.; Palmfeldt, J.; Vockley, J.; Leipnitz, G. ETHE1 and MOCS1 deficiencies: Disruption of mitochondrial bioenergetics, dynamics, redox homeostasis and endoplasmic reticulum-mitochondria crosstalk in patient fibroblasts. Sci. Rep. 2019, 9, 12651. [Google Scholar] [CrossRef]
- Goudarzi, S.; Babicz, J.T., Jr.; Kabil, O.; Banerjee, R.; Solomon, E.I. Spectroscopic and Electronic Structure Study of ETHE1: Elucidating the Factors Influencing Sulfur Oxidation and Oxygenation in Mononuclear Nonheme Iron Enzymes. J. Am. Chem. Soc. 2018, 140, 14887–14902. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Di Meo, I.; Zeviani, M.; Viscomi, C.; Braun, H.P. Proteome adaptations in Ethe1-deficient mice indicate a role in lipid catabolism and cytoskeleton organization via post-translational protein modifications. Biosci. Rep. 2013, 33, e00052. [Google Scholar] [CrossRef]
- Grings, M.; Wajner, M.; Leipnitz, G. Mitochondrial Dysfunction and Redox Homeostasis Impairment as Pathomechanisms of Brain Damage in Ethylmalonic Encephalopathy: Insights from Animal and Human Studies. Cell Mol. Neurobiol. 2022, 42, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Horton, A.; Hong, K.M.; Pandithan, D.; Allen, M.; Killick, C.; Goergen, S.; Springer, A.; Phelan, D.; Marty, M.; Halligan, R.; et al. Ethylmalonic encephalopathy masquerading as meningococcemia. Mol. Case Stud. 2022, 8, a006193. [Google Scholar] [CrossRef] [PubMed]
- Tiranti, V.; Briem, E.; Lamantea, E.; Mineri, R.; Papaleo, E.; De Gioia, L.; Forlani, F.; Rinaldo, P.; Dickson, P.; Abu-Libdeh, B.; et al. ETHE1 mutations are specific to ethylmalonic encephalopathy. J. Med. Genet. 2006, 43, 340–346. [Google Scholar] [CrossRef]
- Witherspoon, M.; Sandu, D.; Lu, C.; Wang, K.; Edwards, R.; Yeung, A.; Gelincik, O.; Manfredi, G.; Gross, S.; Kopelovich, L.; et al. ETHE1 overexpression promotes SIRT1 and PGC1alpha mediated aerobic glycolysis, oxidative phosphorylation, mitochondrial biogenesis and colorectal cancer. Oncotarget 2019, 10, 4004–4017. [Google Scholar] [CrossRef] [PubMed]
- Ozluk, E.; Coppola, D.; Mohammad, I.Z.; Islam, T.; Ghali, G.; Kevil, C.G.; Shackelford, R.E. Ethylmalonic Encephalopathy 1 Protein Is Increased in Colorectal Adenocarcinoma. Anticancer. Res. 2021, 41, 4719–4723. [Google Scholar] [CrossRef]
- Pan, J.; Song, G.; Chen, D.; Li, Y.; Liu, S.; Hu, S.; Rosa, C.; Eichinger, D.; Pino, I.; Zhu, H.; et al. Identification of Serological Biomarkers for Early Diagnosis of Lung Cancer Using a Protein Array-Based Approach. Mol. Cell. Proteom. 2017, 16, 2069–2078. [Google Scholar] [CrossRef]
- Jiang, Y.Z.; Ma, D.; Suo, C.; Shi, J.; Xue, M.; Hu, X.; Xiao, Y.; Yu, K.D.; Liu, Y.R.; Yu, Y.; et al. Genomic and Transcriptomic Landscape of Triple-Negative Breast Cancers: Subtypes and Treatment Strategies. Cancer Cell 2019, 35, 428–440.e425. [Google Scholar] [CrossRef]
- Gong, T.Q.; Jiang, Y.Z.; Shao, C.; Peng, W.T.; Liu, M.W.; Li, D.Q.; Zhang, B.Y.; Du, P.; Huang, Y.; Li, F.F.; et al. Proteome-centric cross-omics characterization and integrated network analyses of triple-negative breast cancer. Cell Rep. 2022, 38, 110460. [Google Scholar] [CrossRef]
- Kabil, O.; Banerjee, R. Characterization of patient mutations in human persulfide dioxygenase (ETHE1) involved in H2S catabolism. J. Biol. Chem. 2012, 287, 44561–44567. [Google Scholar] [CrossRef]
- Henriques, B.J.; Lucas, T.G.; Rodrigues, J.V.; Frederiksen, J.H.; Teixeira, M.S.; Tiranti, V.; Bross, P.; Gomes, C.M. Ethylmalonic encephalopathy ETHE1 R163W/R163Q mutations alter protein stability and redox properties of the iron centre. PLoS ONE 2014, 9, e107157. [Google Scholar] [CrossRef]
- Kabil, O.; Motl, N.; Strack, M.; Seravalli, J.; Metzler-Nolte, N.; Banerjee, R. Mechanism-based inhibition of human persulfide dioxygenase by gamma-glutamyl-homocysteinyl-glycine. J. Biol. Chem. 2018, 293, 12429–12439. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Kasamatsu, S.; Matsunaga, T.; Akashi, S.; Ono, K.; Nishimura, A.; Morita, M.; Abdul Hamid, H.; Fujii, S.; Kitamura, H.; et al. Protein polysulfidation-dependent persulfide dioxygenase activity of ethylmalonic encephalopathy protein 1. Biochem. Biophys. Res. Commun. 2016, 480, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.F.; Poyry, T.A.A.; Stoneley, M.; Willis, A.E. Signaling from mTOR to eIF2alpha mediates cell migration in response to the chemotherapeutic doxorubicin. Sci. Signal. 2019, 12, eaaw6763. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, N.; Gorman, A.M.; Gupta, S.; Samali, A. The eIF2alpha kinases: Their structures and functions. Cell. Mol. Life Sci. 2013, 70, 3493–3511. [Google Scholar] [CrossRef]
- Nagelkerke, A.; Bussink, J.; Mujcic, H.; Wouters, B.G.; Lehmann, S.; Sweep, F.C.; Span, P.N. Hypoxia stimulates migration of breast cancer cells via the PERK/ATF4/LAMP3-arm of the unfolded protein response. Breast Cancer Res. 2013, 15, R2. [Google Scholar] [CrossRef]
- Zeng, P.; Sun, S.; Li, R.; Xiao, Z.X.; Chen, H. HER2 Upregulates ATF4 to Promote Cell Migration via Activation of ZEB1 and Downregulation of E-Cadherin. Int. J. Mol. Sci. 2019, 20, 2223. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, A.; Munoz-Muela, E.; Marchal, J.A.; Cara, F.E.; Molina, M.P.; Cruz-Lozano, M.; Jimenez, G.; Verma, A.; Ramirez, A.; Qian, W.; et al. Activating Transcription Factor 4 Modulates TGFbeta-Induced Aggressiveness in Triple-Negative Breast Cancer via SMAD2/3/4 and mTORC2 Signaling. Clin. Cancer Res. 2018, 24, 5697–5709. [Google Scholar] [CrossRef]
- Bao, J.; Qian, Z.; Liu, L.; Hong, X.; Che, H.; Wu, X. Pharmacological Disruption of Phosphorylated Eukaryotic Initiation Factor-2alpha/Activating Transcription Factor 4/Indian Hedgehog Protects Intervertebral Disc Degeneration via Reducing the Reactive Oxygen Species and Apoptosis of Nucleus Pulposus Cells. Front. Cell Dev. Biol. 2021, 9, 675486. [Google Scholar] [CrossRef]
- Dong, J.; Jin, S.; Guo, J.; Yang, R.; Tian, D.; Xue, H.; Xiao, L.; Guo, Q.; Wang, R.; Xu, M.; et al. Pharmacological inhibition of eIF2alpha phosphorylation by integrated stress response inhibitor (ISRIB) ameliorates vascular calcification in rats. Physiol. Res. 2022, 71, 379–388. [Google Scholar] [CrossRef]
- Di Rocco, M.; Caruso, U.; Briem, E.; Rossi, A.; Allegri, A.E.; Buzzi, D.; Tiranti, V. A case of ethylmalonic encephalopathy with atypical clinical and biochemical presentation. Mol. Genet. Metab. 2006, 89, 395–397. [Google Scholar] [CrossRef]
- You, K.; Wang, L.; Chou, C.H.; Liu, K.; Nakata, T.; Jaiswal, A.; Yao, J.; Lefkovith, A.; Omar, A.; Perrigoue, J.G.; et al. QRICH1 dictates the outcome of ER stress through transcriptional control of proteostasis. Science 2021, 371, eabb6896. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Cevallos, R.C.; Jan, E. An upstream open reading frame regulates translation of GADD34 during cellular stresses that induce eIF2alpha phosphorylation. J. Biol. Chem. 2009, 284, 6661–6673. [Google Scholar] [CrossRef]
- Lei, D.; Chen, Y.; Zhou, Y.; Hu, G.; Luo, F. A Starvation-Based 9-mRNA Signature Correlates With Prognosis in Patients With Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 716757. [Google Scholar] [CrossRef]
- Hamamura, K.; Minami, K.; Tanjung, N.; Wan, Q.; Koizumi, M.; Matsuura, N.; Na, S.; Yokota, H. Attenuation of malignant phenotypes of breast cancer cells through eIF2alpha-mediated downregulation of Rac1 signaling. Int. J. Oncol. 2014, 44, 1980–1988. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-Y.; Liao, L.; Hu, S.-Y.; Deng, L.; Andriani, L.; Zhang, T.-M.; Zhang, Y.-L.; Ma, X.-Y.; Zhang, F.-L.; Liu, Y.-Y.; et al. ETHE1 Accelerates Triple-Negative Breast Cancer Metastasis by Activating GCN2/eIF2α/ATF4 Signaling. Int. J. Mol. Sci. 2023, 24, 14566. https://doi.org/10.3390/ijms241914566
Yang S-Y, Liao L, Hu S-Y, Deng L, Andriani L, Zhang T-M, Zhang Y-L, Ma X-Y, Zhang F-L, Liu Y-Y, et al. ETHE1 Accelerates Triple-Negative Breast Cancer Metastasis by Activating GCN2/eIF2α/ATF4 Signaling. International Journal of Molecular Sciences. 2023; 24(19):14566. https://doi.org/10.3390/ijms241914566
Chicago/Turabian StyleYang, Shao-Ying, Li Liao, Shu-Yuan Hu, Ling Deng, Lisa Andriani, Tai-Mei Zhang, Yin-Ling Zhang, Xiao-Yan Ma, Fang-Lin Zhang, Ying-Ying Liu, and et al. 2023. "ETHE1 Accelerates Triple-Negative Breast Cancer Metastasis by Activating GCN2/eIF2α/ATF4 Signaling" International Journal of Molecular Sciences 24, no. 19: 14566. https://doi.org/10.3390/ijms241914566
APA StyleYang, S. -Y., Liao, L., Hu, S. -Y., Deng, L., Andriani, L., Zhang, T. -M., Zhang, Y. -L., Ma, X. -Y., Zhang, F. -L., Liu, Y. -Y., & Li, D. -Q. (2023). ETHE1 Accelerates Triple-Negative Breast Cancer Metastasis by Activating GCN2/eIF2α/ATF4 Signaling. International Journal of Molecular Sciences, 24(19), 14566. https://doi.org/10.3390/ijms241914566