
The Local Anaesthetic Procaine Prodrugs ProcCluster® and Procaine Hydrochloride Impair SARS-CoV-2 Replication and Egress In Vitro


Abstract
:1. Introduction
2. Results
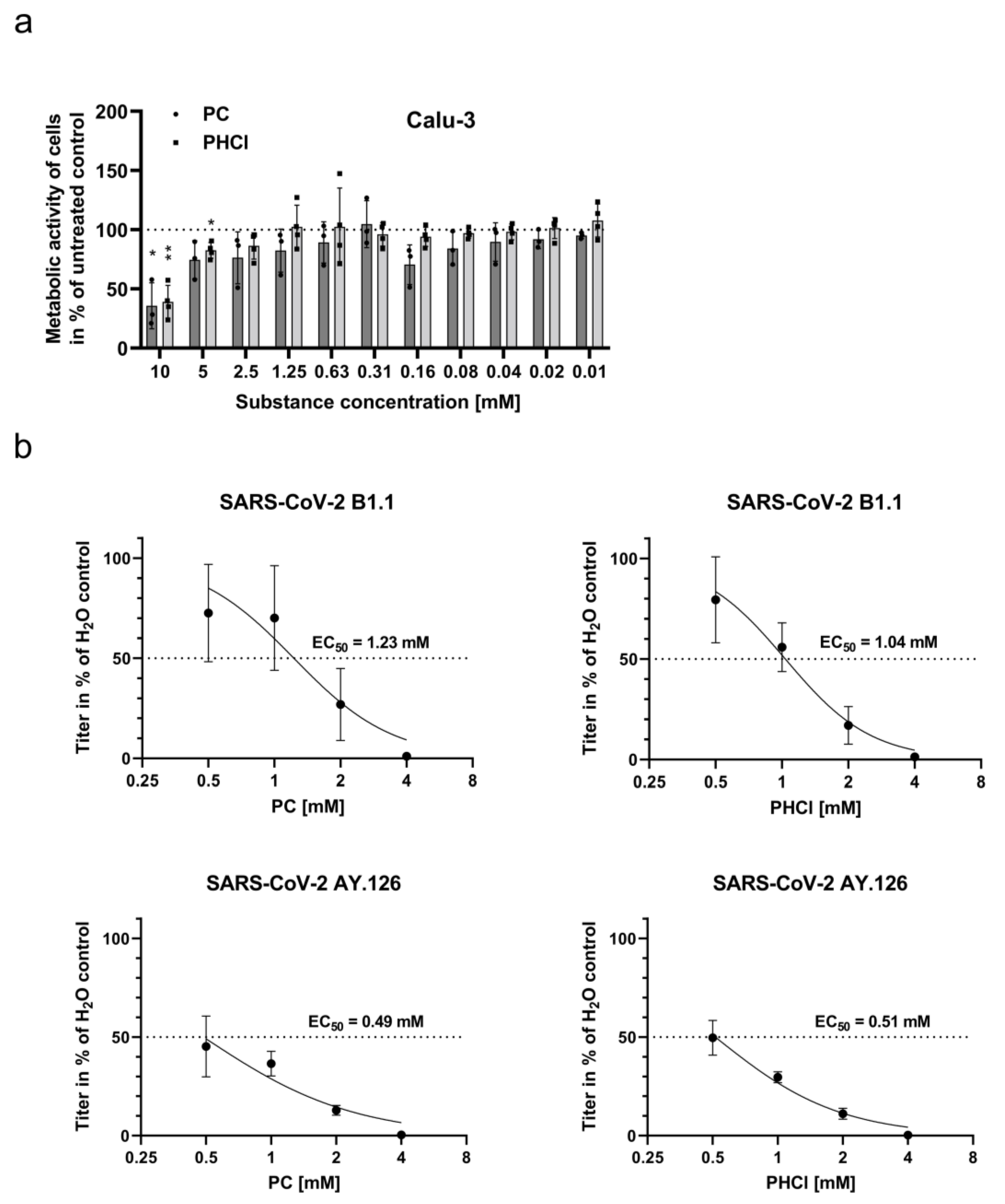
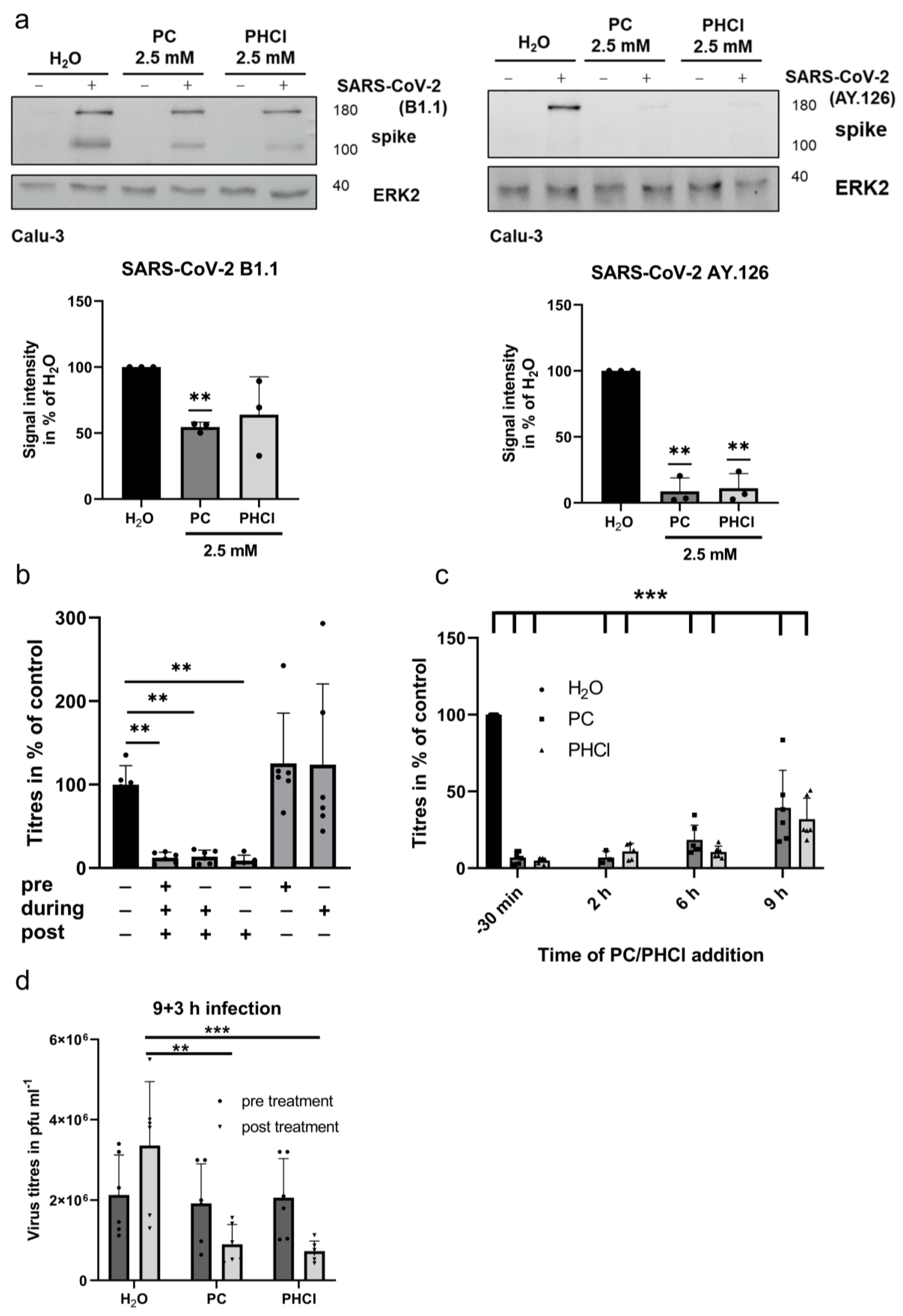
2.1. ProcCluster® and Procaine Hydrochloride Treatment Results in Reduced Replication Efficacy of SARS-CoV-2 In Vitro
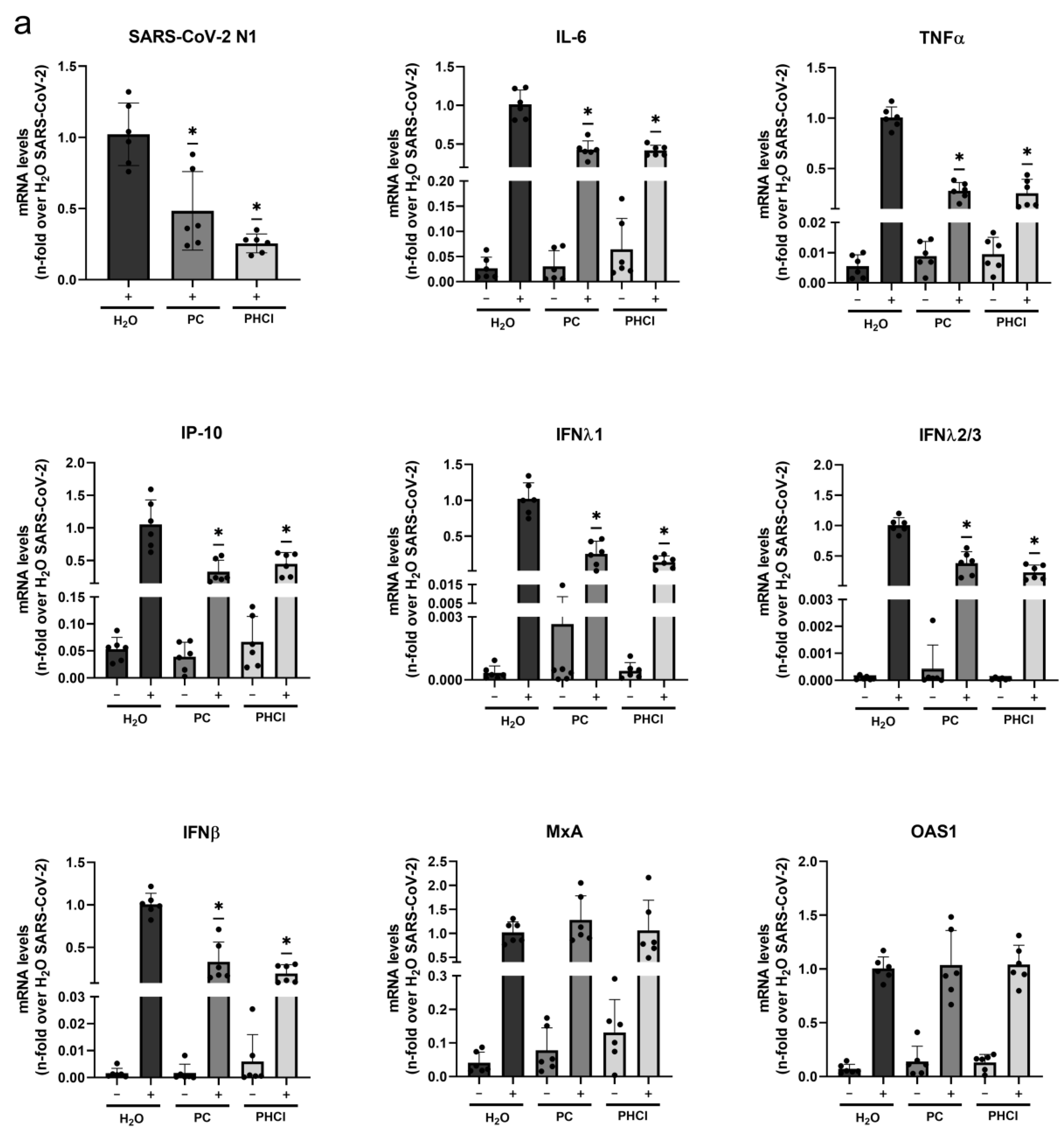
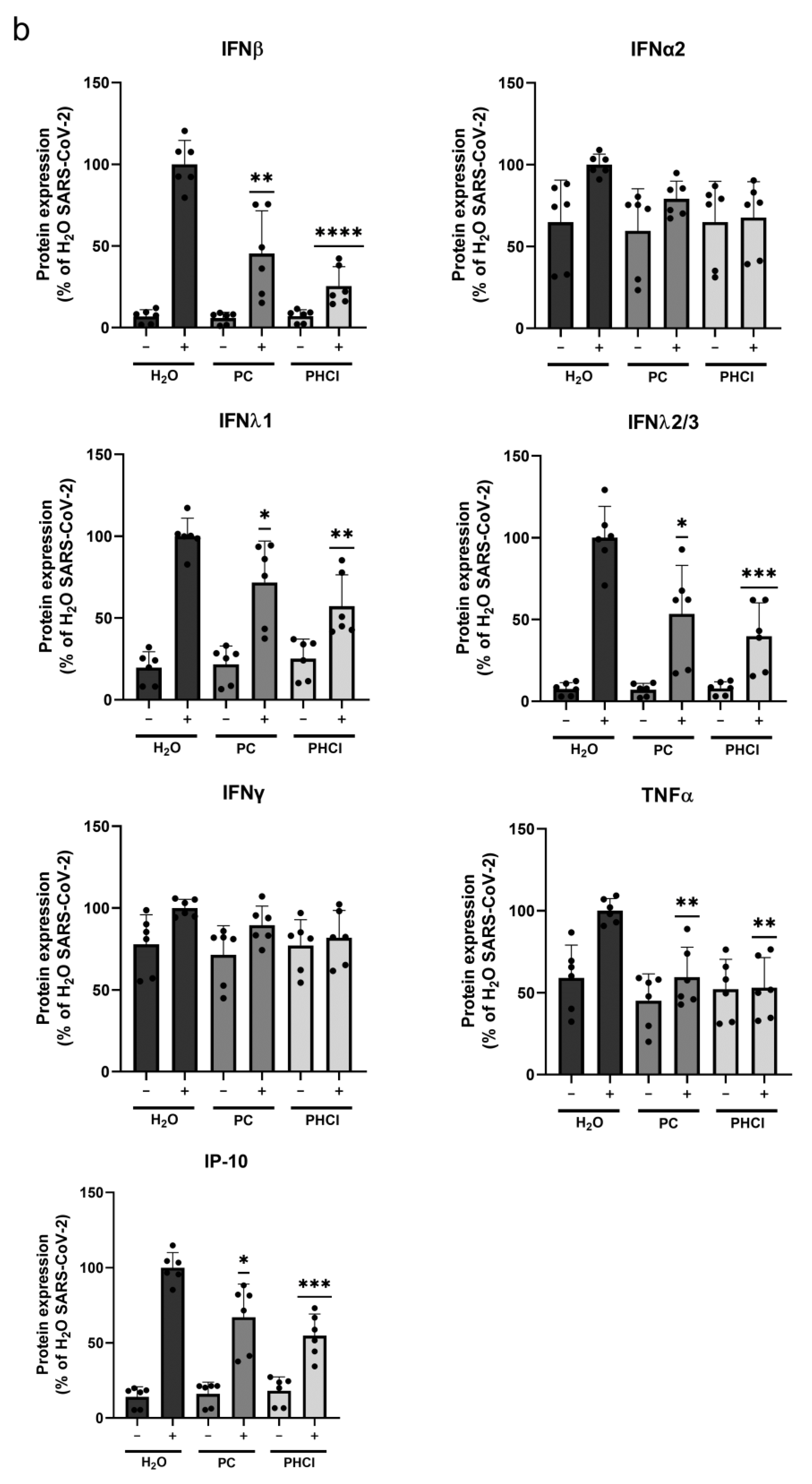
2.2. The SARS-CoV-2-Induced Interferon and Cytokine Response Is Reduced in Procaine-Treated Cells
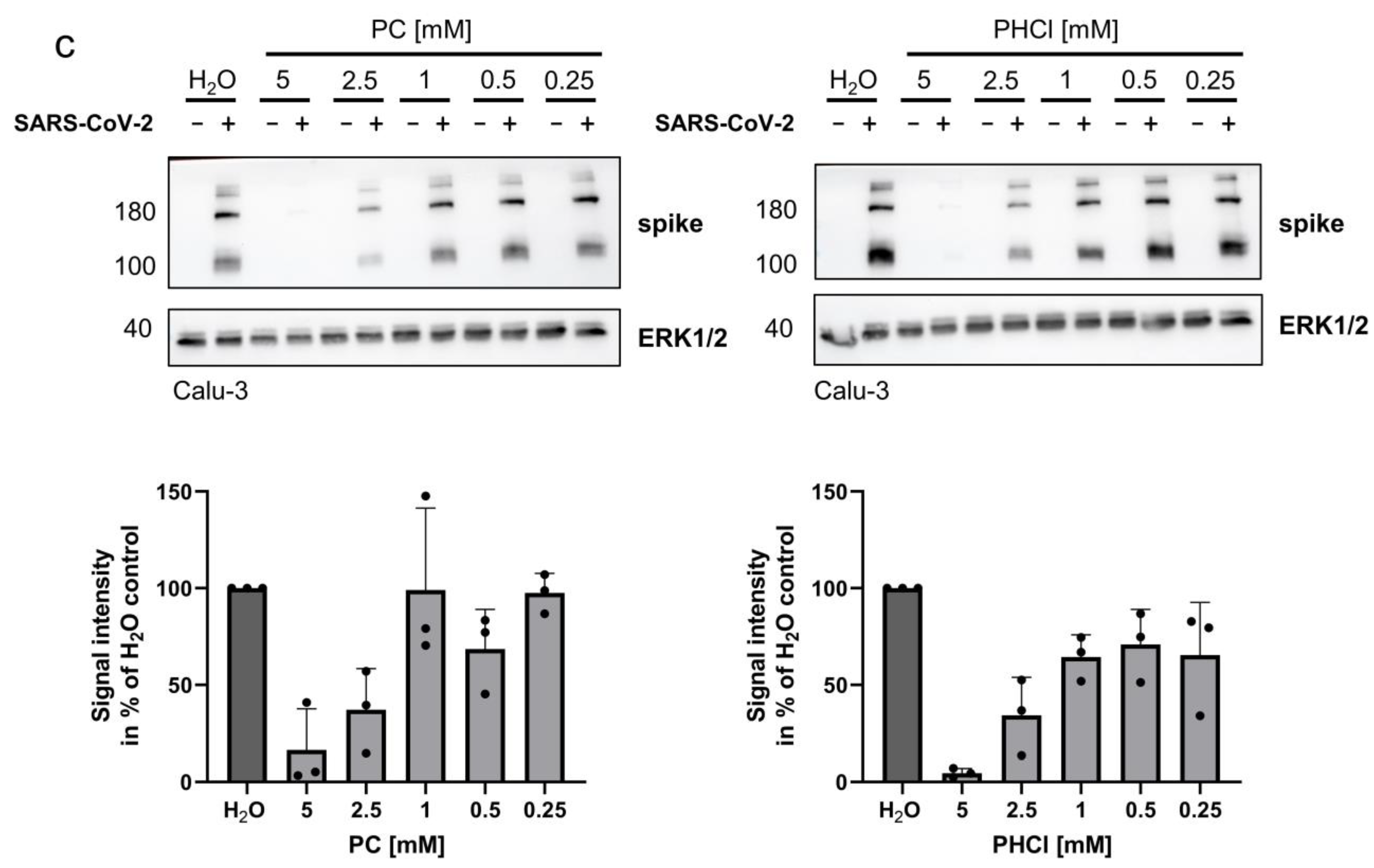
2.3. ProcCluster® Treatment Inhibits a Late Stage of the SARS-CoV-2 Replication Cycle
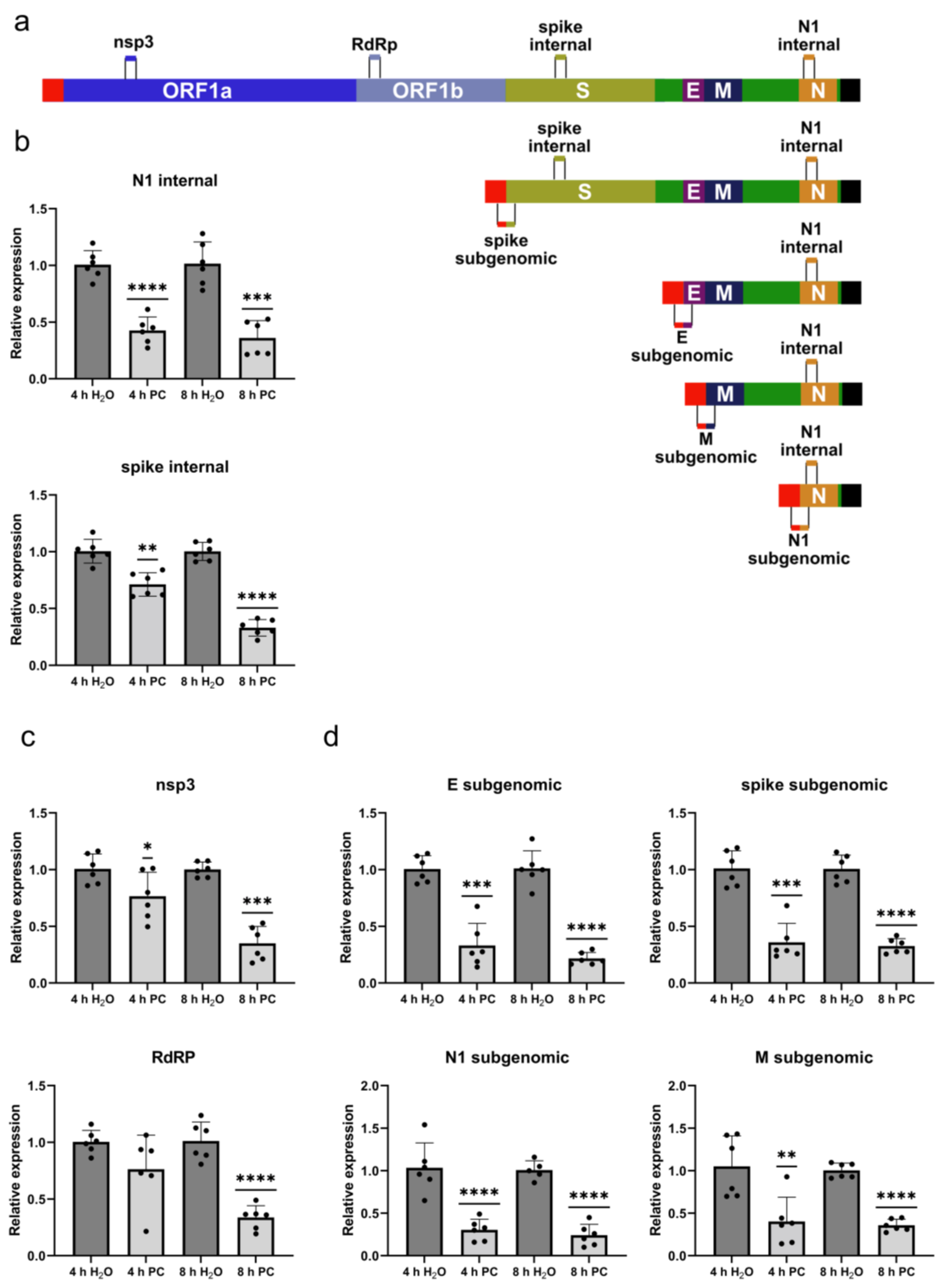
2.4. ProcCluster® Treatment Differentially Affects SARS-CoV-2 Replication and Transcription
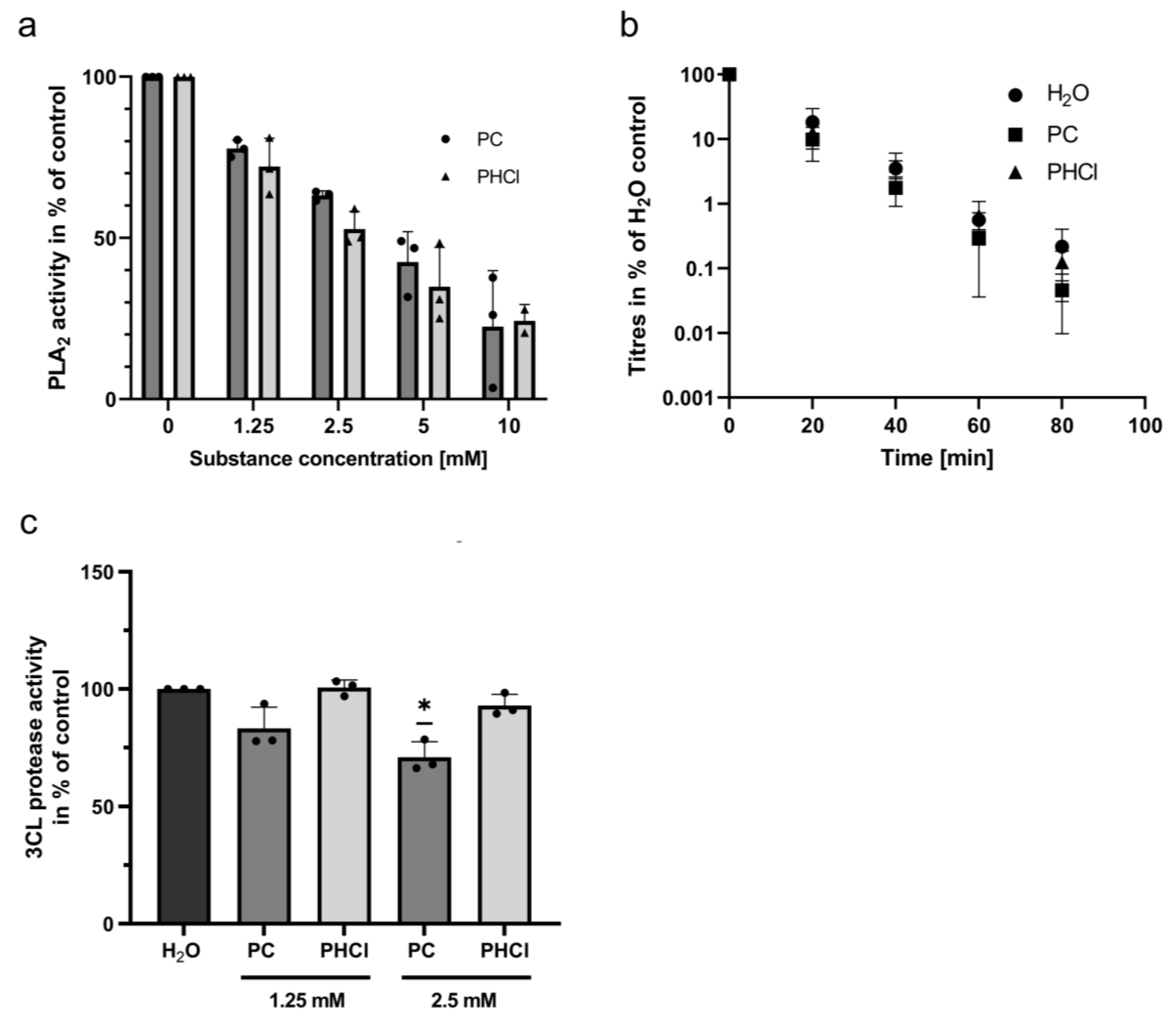
2.5. Procaine Treatment Inhibits Phospholipase A2
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Virus Cultivation
4.2. Pharmacological Substances
4.3. MTT Assay
4.4. EC50 Determination
4.5. Infection of Cell Culture
4.6. Plaque Assay
4.7. qRT-PCR
4.8. Cytokine Measurements
4.9. Western Blot
4.10. Phospholipase A2 Assay
4.11. 3CL Protease Assay
4.12. Software
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Alfajaro, M.M.; DeWeirdt, P.C.; Hanna, R.E.; Lu-Culligan, W.J.; Cai, W.L.; Strine, M.S.; Zhang, S.M.; Graziano, V.R.; Schmitz, C.O.; et al. Genome-wide CRISPR Screens Reveal Host Factors Critical for SARS-CoV-2 Infection. Cell 2021, 184, 76–91.e13. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L plays a key role in SARS-CoV-2 infection in humans and humanized mice and is a promising target for new drug development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef]
- Bayati, A.; Kumar, R.; Francis, V.; McPherson, P.S. SARS-CoV-2 infects cells after viral entry via clathrin-mediated endocytosis. J. Biol. Chem. 2021, 296, 100306. [Google Scholar] [CrossRef]
- Romano, M.; Ruggiero, A.; Squeglia, F.; Maga, G.; Berisio, R. A Structural View of SARS-CoV-2 RNA Replication Machinery: RNA Synthesis, Proofreading and Final Capping. Cells 2020, 9, 1267. [Google Scholar] [CrossRef]
- Roingeard, P.; Eymieux, S.; Burlaud-Gaillard, J.; Hourioux, C.; Patient, R.; Blanchard, E. The double-membrane vesicle (DMV): A virus-induced organelle dedicated to the replication of SARS-CoV-2 and other positive-sense single-stranded RNA viruses. Cell Mol. Life Sci. 2022, 79, 425. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, Y.G.; Zhang, H. Endomembrane remodeling in SARS-CoV-2 infection. Cell Insight 2022, 1, 100031. [Google Scholar] [CrossRef]
- Scutigliani, E.M.; Kikkert, M. Interaction of the innate immune system with positive-strand RNA virus replication organelles. Cytokine Growth Factor. Rev. 2017, 37, 17–27. [Google Scholar] [CrossRef]
- Baggen, J.; Vanstreels, E.; Jansen, S.; Daelemans, D. Cellular host factors for SARS-CoV-2 infection. Nat. Microbiol. 2021, 6, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Grellet, E.; L’Hote, I.; Goulet, A.; Imbert, I. Replication of the coronavirus genome: A paradox among positive-strand RNA viruses. J. Biol. Chem. 2022, 298, 101923. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, C.; Mulroney, L.; Leger, A.; Castelli, M.; Criscuolo, E.; Williamson, M.K.; Davidson, A.D.; Almuqrin, A.; Giambruno, R.; Jain, M.; et al. Nanopore ReCappable sequencing maps SARS-CoV-2 5′ capping sites and provides new insights into the structure of sgRNAs. Nucleic Acids Res. 2022, 50, 3475–3489. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Jiang, A.; Feng, J.; Li, G.; Guo, D.; Sajid, M.; Wu, K.; Zhang, Q.; Ponty, Y.; Will, S.; et al. The SARS-CoV-2 subgenome landscape and its novel regulatory features. Mol. Cell 2021, 81, 2135–2147.e2135. [Google Scholar] [CrossRef]
- Sawicki, S.G.; Sawicki, D.L.; Siddell, S.G. A contemporary view of coronavirus transcription. J. Virol. 2007, 81, 20–29. [Google Scholar] [CrossRef]
- Ghosh, S.; Dellibovi-Ragheb, T.A.; Kerviel, A.; Pak, E.; Qiu, Q.; Fisher, M.; Takvorian, P.M.; Bleck, C.; Hsu, V.W.; Fehr, A.R.; et al. beta-Coronaviruses Use Lysosomes for Egress Instead of the Biosynthetic Secretory Pathway. Cell 2020, 183, 1520–1535.e1514. [Google Scholar] [CrossRef]
- Chen, D.; Zheng, Q.; Sun, L.; Ji, M.; Li, Y.; Deng, H.; Zhang, H. ORF3a of SARS-CoV-2 promotes lysosomal exocytosis-mediated viral egress. Dev. Cell 2021, 56, 3250–3263.e3255. [Google Scholar] [CrossRef]
- Deinhardt-Emmer, S.; Wittschieber, D.; Sanft, J.; Kleemann, S.; Elschner, S.; Haupt, K.F.; Vau, V.; Haring, C.; Rodel, J.; Henke, A.; et al. Early postmortem mapping of SARS-CoV-2 RNA in patients with COVID-19 and the correlation with tissue damage. eLife 2021, 10, e60361. [Google Scholar] [CrossRef]
- Taoufik, Y.; de Goer de Herve, M.G.; Corgnac, S.; Durrbach, A.; Mami-Chouaib, F. When Immunity Kills: The Lessons of SARS-CoV-2 Outbreak. Front. Immunol. 2021, 12, 692598. [Google Scholar] [CrossRef]
- Maiuolo, J.; Mollace, R.; Gliozzi, M.; Musolino, V.; Carresi, C.; Paone, S.; Scicchitano, M.; Macri, R.; Nucera, S.; Bosco, F.; et al. The Contribution of Endothelial Dysfunction in Systemic Injury Subsequent to SARS-Cov-2 Infection. Int. J. Mol. Sci. 2020, 21, 9309. [Google Scholar] [CrossRef]
- Manne, B.K.; Denorme, F.; Middleton, E.A.; Portier, I.; Rowley, J.W.; Stubben, C.; Petrey, A.C.; Tolley, N.D.; Guo, L.; Cody, M.; et al. Platelet gene expression and function in patients with COVID-19. Blood 2020, 136, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Gustine, J.N.; Jones, D. Immunopathology of Hyperinflammation in COVID-19. Am. J. Pathol. 2021, 191, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.Y.; Day-Lewis, M.; Henderson, L.A.; Friedman, K.G.; Lo, J.; Roberts, J.E.; Lo, M.S.; Platt, C.D.; Chou, J.; Hoyt, K.J.; et al. Distinct clinical and immunological features of SARS-CoV-2-induced multisystem inflammatory syndrome in children. J. Clin. Investig. 2020, 130, 5942–5950. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Wang, R.; Chen, J.; Hozumi, Y.; Yin, C.; Wei, G.W. Emerging Vaccine-Breakthrough SARS-CoV-2 Variants. ACS Infect. Dis. 2022, 8, 546–556. [Google Scholar] [CrossRef]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.E.; Tuekprakhon, A.; Nutalai, R.; et al. Evidence of escape of SARS-CoV-2 variant B.1.351 from natural and vaccine-induced sera. Cell 2021, 184, 2348–2361.e2346. [Google Scholar] [CrossRef]
- Fischer, W.A., 2nd; Eron, J.J., Jr.; Holman, W.; Cohen, M.S.; Fang, L.; Szewczyk, L.J.; Sheahan, T.P.; Baric, R.; Mollan, K.R.; Wolfe, C.R.; et al. A phase 2a clinical trial of molnupiravir in patients with COVID-19 shows accelerated SARS-CoV-2 RNA clearance and elimination of infectious virus. Sci. Transl. Med. 2022, 14, eabl7430. [Google Scholar] [CrossRef]
- Chen, P.J.; Chao, C.M.; Lai, C.C. Clinical efficacy and safety of favipiravir in the treatment of COVID-19 patients. J. Infect. 2021, 82, 186–230. [Google Scholar] [CrossRef]
- Chuah, C.H.; Chow, T.S.; Hor, C.P.; Cheng, J.T.; Ker, H.B.; Lee, H.G.; Lee, K.S.; Nordin, N.; Ng, T.K.; Zaid, M.; et al. Efficacy of Early Treatment with Favipiravir on Disease Progression among High Risk COVID-19 Patients: A Randomized, Open-Label Clinical Trial. Clin. Infect. Dis. 2021, 75, e432–e439. [Google Scholar] [CrossRef]
- Lamb, Y.N. Nirmatrelvir Plus Ritonavir: First Approval. Drugs 2022, 82, 585–591. [Google Scholar] [CrossRef]
- Ansems, K.; Grundeis, F.; Dahms, K.; Mikolajewska, A.; Thieme, V.; Piechotta, V.; Metzendorf, M.I.; Stegemann, M.; Benstoem, C.; Fichtner, F. Remdesivir for the treatment of COVID-19. Cochrane Database Syst. Rev. 2021, 8, CD014962. [Google Scholar] [CrossRef]
- Huang, J.; Zhou, C.; Deng, J.; Zhou, J. JAK inhibition as a new treatment strategy for patients with COVID-19. Biochem. Pharmacol. 2022, 202, 115162. [Google Scholar] [CrossRef]
- Elahi, R.; Karami, P.; Heidary, A.H.; Esmaeilzadeh, A. An updated overview of recent advances, challenges, and clinical considerations of IL-6 signaling blockade in severe coronavirus disease 2019 (COVID-19). Int. Immunopharmacol. 2022, 105, 108536. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M.; O’Meara, M.J.; Rezelj, V.V.; Guo, J.Z.; Swaney, D.L.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Sadegh, S.; Matschinske, J.; Blumenthal, D.B.; Galindez, G.; Kacprowski, T.; List, M.; Nasirigerdeh, R.; Oubounyt, M.; Pichlmair, A.; Rose, T.D.; et al. Exploring the SARS-CoV-2 virus-host-drug interactome for drug repurposing. Nat. Commun. 2020, 11, 3518. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.F.; McQueen, D.S.; Seckl, J.R. Local anaesthesia prevents acute inflammatory changes in neuropeptide messenger RNA expression in rat dorsal root ganglia neurons. Neurosci. Lett. 1994, 175, 111–113. [Google Scholar] [CrossRef]
- Hollmann, M.W.; Durieux, M.E. Local anesthetics and the inflammatory response: A new therapeutic indication? Anesthesiology 2000, 93, 858–875. [Google Scholar] [CrossRef]
- Cruz, F.F.; Rocco, P.R.; Pelosi, P. Anti-inflammatory properties of anesthetic agents. Crit. Care 2017, 21, 67. [Google Scholar] [CrossRef]
- Cassuto, J.; Sinclair, R.; Bonderovic, M. Anti-inflammatory properties of local anesthetics and their present and potential clinical implications. Acta Anaesthesiol. Scand. 2006, 50, 265–282. [Google Scholar] [CrossRef]
- Taylor, A.; McLeod, G. Basic pharmacology of local anaesthetics. BJA Educ. 2020, 20, 34–41. [Google Scholar] [CrossRef]
- Hollmann, M.W.; Durieux, M.E.; Graf, B.M. Novel local anaesthetics and novel indications for local anaesthetics. Curr. Opin. Anaesthesiol. 2001, 14, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Hollmann, M.W.; Herroeder, S.; Kurz, K.S.; Hoenemann, C.W.; Struemper, D.; Hahnenkamp, K.; Durieux, M.E. Time-dependent inhibition of G protein-coupled receptor signaling by local anesthetics. Anesthesiology 2004, 100, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, S.; Li, X.; Li, C.; Ma, L. Procaine Inhibits the Proliferation and Migration of Colon Cancer Cells Through Inactivation of the ERK/MAPK/FAK Pathways by Regulation of RhoA. Oncol. Res. 2018, 26, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, S.; Hrincius, E.R.; Boergeling, Y. The Two Sides of the Same Coin-Influenza Virus and Intracellular Signal Transduction. Cold Spring Harb. Perspect. Med. 2021, 11, a038513. [Google Scholar] [CrossRef]
- Yanguez, E.; Hunziker, A.; Dobay, M.P.; Yildiz, S.; Schading, S.; Elshina, E.; Karakus, U.; Gehrig, P.; Grossmann, J.; Dijkman, R.; et al. Phosphoproteomic-based kinase profiling early in influenza virus infection identifies GRK2 as antiviral drug target. Nat. Commun. 2018, 9, 3679. [Google Scholar] [CrossRef]
- Bouhaddou, M.; Memon, D.; Meyer, B.; White, K.M.; Rezelj, V.V.; Correa Marrero, M.; Polacco, B.J.; Melnyk, J.E.; Ulferts, S.; Kaake, R.M.; et al. The Global Phosphorylation Landscape of SARS-CoV-2 Infection. Cell 2020, 182, 685–712.e619. [Google Scholar] [CrossRef]
- Fuchs, P.; Levanon, A. Inhibition of adsorption of West-Nile and herpes simplex viruses by procaine. Arch. Virol. 1978, 56, 163–168. [Google Scholar] [CrossRef]
- Castilla, V.; Mersich, S.E.; Candurra, N.A.; Damonte, E.B. The entry of Junin virus into Vero cells. Arch. Virol. 1994, 136, 363–374. [Google Scholar] [CrossRef]
- Deinhardt-Emmer, S.; Böttcher, S.; Häring, C.; Giebeler, L.; Henke, A.; Zell, R.; Jungwirth, J.; Jordan, P.M.; Werz, O.; Hornung, F.; et al. SARS-CoV-2 Causes Severe Epithelial Inflammation and Barrier Dysfunction. J. Virol. 2021, 95, e00110-21. [Google Scholar] [CrossRef]
- Schneider, M.; Ackermann, K.; Stuart, M.; Wex, C.; Protzer, U.; Schätzl, H.M.; Gilch, S. Severe acute respiratory syndrome coronavirus replication is severely impaired by MG132 due to proteasome-independent inhibition of M-calpain. J. Virol. 2012, 86, 10112–10122. [Google Scholar] [CrossRef]
- Snijder, E.J.; van der Meer, Y.; Zevenhoven-Dobbe, J.; Onderwater, J.J.; van der Meulen, J.; Koerten, H.K.; Mommaas, A.M. Ultrastructure and origin of membrane vesicles associated with the severe acute respiratory syndrome coronavirus replication complex. J. Virol. 2006, 80, 5927–5940. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, K.A.; Thiel, V.; Dobbe, J.C.; van der Meer, Y.; Snijder, E.J.; Ziebuhr, J. Multiple enzymatic activities associated with severe acute respiratory syndrome coronavirus helicase. J. Virol. 2004, 78, 5619–5632. [Google Scholar] [CrossRef]
- Dagotto, G.; Mercado, N.B.; Martinez, D.R.; Hou, Y.J.; Nkolola, J.P.; Carnahan, R.H.; Crowe, J.E.; Baric, R.S.; Barouch, D.H. Comparison of Subgenomic and Total RNA in SARS-CoV-2-Challenged Rhesus Macaques. J. Virol. 2021, 95, e02370-20. [Google Scholar] [CrossRef] [PubMed]
- Kahl, S.; Mayer, J.; Schuette, K.; Peuravuori, H.; Pross, M.; Schulz, H.U.; Nevalainen, T.J.; Malfertheiner, P. Effect of Procainhydrochloride on Phospholipase A2 Catalytic Activity in Sodium Taurocholate-Induced Acute Experimental Pancreatitis in Rats. Dig. Dis. 2010, 28, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, G.J.; Flower, R.J.; Nijkamp, F.P.; Vane, J.R. Phospholipase A2 activity of guinea-pig isolated perfused lungs: Stimulation, and inhibition by anti-inflammatory steroids. Br. J. Pharmacol. 1978, 62, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Urakami, M.; Takanaka, K. Effects of Various Drugs on Superoxide Generation, Arachidonic Acid Release and Phospholipase A2 in Polymorphonuclear Leukocytes. Jpn. J. Pharmacol. 1988, 46, 275–284. [Google Scholar] [CrossRef]
- Kunze, H.; Nahas, N.; Traynor, J.R.; Wurl, M. Effects of local anaesthetics on phospholipases. Biochim. et Biophys. Acta (BBA)—Lipids Lipid Metab. 1976, 441, 93–102. [Google Scholar] [CrossRef]
- Hendrickson, H.S.; van Dam-Mieras, M.C. Local anesthetic inhibition of pancreatic phospholipase A2 action on lecithin monolayers. J. Lipid Res. 1976, 17, 399–405. [Google Scholar] [CrossRef]
- Brown, W.J.; Chambers, K.; Doody, A. Phospholipase A2 (PLA2) enzymes in membrane trafficking: Mediators of membrane shape and function. Traffic 2003, 4, 214–221. [Google Scholar] [CrossRef]
- Gayda, R.C.; Henderson, G.W.; Markovitz, A. Neuroactive drugs inhibit trypsin and outer membrane protein processing in Escherichia coli K-12. Proc. Natl. Acad. Sci. USA 1979, 76, 2138–2142. [Google Scholar] [CrossRef]
- Prete, M.; Favoino, E.; Catacchio, G.; Racanelli, V.; Perosa, F. SARS-CoV-2 Inflammatory Syndrome. Clinical Features and Rationale for Immunological Treatment. Int. J. Mol. Sci. 2020, 21, 3377. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.; Wong, P.; Klein, J.; Castro, T.B.R.; Silva, J.; Sundaram, M.; Ellingson, M.K.; Mao, T.; Oh, J.E.; Israelow, B.; et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature 2020, 584, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e1039. [Google Scholar] [CrossRef]
- Lowery, S.A.; Sariol, A.; Perlman, S. Innate immune and inflammatory responses to SARS-CoV-2: Implications for COVID-19. Cell Host Microbe 2021, 29, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Galani, I.E.; Rovina, N.; Lampropoulou, V.; Triantafyllia, V.; Manioudaki, M.; Pavlos, E.; Koukaki, E.; Fragkou, P.C.; Panou, V.; Rapti, V.; et al. Untuned antiviral immunity in COVID-19 revealed by temporal type I/III interferon patterns and flu comparison. Nat. Immunol. 2021, 22, 32–40. [Google Scholar] [CrossRef]
- Zhou, Z.; Ren, L.; Zhang, L.; Zhong, J.; Xiao, Y.; Jia, Z.; Guo, L.; Yang, J.; Wang, C.; Jiang, S.; et al. Heightened Innate Immune Responses in the Respiratory Tract of COVID-19 Patients. Cell Host Microbe 2020, 27, 883–890.e882. [Google Scholar] [CrossRef]
- Katsura, H.; Sontake, V.; Tata, A.; Kobayashi, Y.; Edwards, C.E.; Heaton, B.E.; Konkimalla, A.; Asakura, T.; Mikami, Y.; Fritch, E.J.; et al. Human Lung Stem Cell-Based Alveolospheres Provide Insights into SARS-CoV-2-Mediated Interferon Responses and Pneumocyte Dysfunction. Cell Stem Cell 2020, 27, 890–904.e898. [Google Scholar] [CrossRef]
- Trouillet-Assant, S.; Viel, S.; Gaymard, A.; Pons, S.; Richard, J.C.; Perret, M.; Villard, M.; Brengel-Pesce, K.; Lina, B.; Mezidi, M.; et al. Type I IFN immunoprofiling in COVID-19 patients. J. Allergy Clin. Immunol. 2020, 146, 206–208.e202. [Google Scholar] [CrossRef]
- Youk, J.; Kim, T.; Evans, K.V.; Jeong, Y.I.; Hur, Y.; Hong, S.P.; Kim, J.H.; Yi, K.; Kim, S.Y.; Na, K.J.; et al. Three-Dimensional Human Alveolar Stem Cell Culture Models Reveal Infection Response to SARS-CoV-2. Cell Stem Cell 2020, 27, 905–919.e910. [Google Scholar] [CrossRef]
- Long, S. SARS-CoV-2 Subgenomic RNAs: Characterization, Utility, and Perspectives. Viruses 2021, 13, 1923. [Google Scholar] [CrossRef]
- Lee, J.Y.; Wing, P.A.C.; Gala, D.S.; Noerenberg, M.; Jarvelin, A.I.; Titlow, J.; Zhuang, X.; Palmalux, N.; Iselin, L.; Thompson, M.K.; et al. Absolute quantitation of individual SARS-CoV-2 RNA molecules provides a new paradigm for infection dynamics and variant differences. eLife 2022, 11, e74153. [Google Scholar] [CrossRef] [PubMed]
- Cortese, M.; Lee, J.Y.; Cerikan, B.; Neufeldt, C.J.; Oorschot, V.M.J.; Kohrer, S.; Hennies, J.; Schieber, N.L.; Ronchi, P.; Mizzon, G.; et al. Integrative Imaging Reveals SARS-CoV-2-Induced Reshaping of Subcellular Morphologies. Cell Host Microbe 2020, 28, 853–866.e855. [Google Scholar] [CrossRef] [PubMed]
- Prydz, K.; Saraste, J. The life cycle and enigmatic egress of coronaviruses. Mol. Microbiol. 2022, 117, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Scherer, K.M.; Mascheroni, L.; Carnell, G.W.; Wunderlich, L.C.S.; Makarchuk, S.; Brockhoff, M.; Mela, I.; Fernandez-Villegas, A.; Barysevich, M.; Stewart, H.; et al. SARS-CoV-2 nucleocapsid protein adheres to replication organelles before viral assembly at the Golgi/ERGIC and lysosome-mediated egress. Sci. Adv. 2022, 8, eabl4895. [Google Scholar] [CrossRef]
- Saraste, J.; Prydz, K. Assembly and Cellular Exit of Coronaviruses: Hijacking an Unconventional Secretory Pathway from the Pre-Golgi Intermediate Compartment via the Golgi Ribbon to the Extracellular Space. Cells 2021, 10, 503. [Google Scholar] [CrossRef]
- Mendonca, L.; Howe, A.; Gilchrist, J.B.; Sheng, Y.; Sun, D.; Knight, M.L.; Zanetti-Domingues, L.C.; Bateman, B.; Krebs, A.S.; Chen, L.; et al. Correlative multi-scale cryo-imaging unveils SARS-CoV-2 assembly and egress. Nat. Commun. 2021, 12, 4629. [Google Scholar] [CrossRef]
- Tian, D.; Sun, Y.; Zhou, J.; Ye, Q. The Global Epidemic of the SARS-CoV-2 Delta Variant, Key Spike Mutations and Immune Escape. Front. Immunol. 2021, 12, 751778. [Google Scholar] [CrossRef]
- Mukherjee, R.; Satardekar, R. Why are some coronavirus variants more infectious? J. Biosci. 2021, 46, 101. [Google Scholar] [CrossRef]
- Klein, S.; Cortese, M.; Winter, S.L.; Wachsmuth-Melm, M.; Neufeldt, C.J.; Cerikan, B.; Stanifer, M.L.; Boulant, S.; Bartenschlager, R.; Chlanda, P. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nat. Commun. 2020, 11, 5885. [Google Scholar] [CrossRef]
- Barreto-Vieira, D.F.; da Silva, M.A.N.; de Almeida, A.L.T.; Rasinhas, A.D.C.; Monteiro, M.E.; Miranda, M.D.; Motta, F.C.; Siqueira, M.M.; Girard-Dias, W.; Archanjo, B.S.; et al. SARS-CoV-2: Ultrastructural Characterization of Morphogenesis in an In Vitro System. Viruses 2022, 14, 201. [Google Scholar] [CrossRef]
- Müller, C.; Hardt, M.; Schwudke, D.; Neuman, B.W.; Pleschka, S.; Ziebuhr, J. Inhibition of Cytosolic Phospholipase A(2)α Impairs an Early Step of Coronavirus Replication in Cell Culture. J. Virol. 2018, 92, 10–128. [Google Scholar] [CrossRef]
- Nakano, T.; Inoue, I.; Shinozaki, R.; Matsui, M.; Akatsuka, T.; Takahashi, S.; Tanaka, K.; Akita, M.; Seo, M.; Hokari, S.; et al. A possible role of lysophospholipids produced by calcium-independent phospholipase A2 in membrane-raft budding and fission. Biochim. et Biophys. Acta (BBA)—Biomembr. 2009, 1788, 2222–2228. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 94. [Google Scholar] [CrossRef] [PubMed]
- Seifen, A.B.; Ferrari, A.A.; Seifen, E.E.; Thompson, D.S.; Chapman, J. Pharmacokinetics of intravenous procaine infusion in humans. Anesth. Analg. 1979, 58, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Usubiaga, J.E.; Wikinski, J.; Ferrero, R.; Usubiaga, L.E.; Wikinski, R. Local anesthetic-induced convulsions in man--an electroencephalographic study. Anesth. Analg. 1966, 45, 611–620. [Google Scholar] [CrossRef] [PubMed]
- Jungwirth, J.; Häring, C.; König, S.; Giebeler, L.; Doshi, H.; Brandt, C.; Deinhardt-Emmer, S.; Löffler, B.; Ehrhardt, C. D,L-Lysine-Acetylsalicylate + Glycine (LASAG) Reduces SARS-CoV-2 Replication and Shows an Additive Effect with Remdesivir. Int. J. Mol. Sci. 2022, 23, 6880. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Source |
|---|---|---|
| SARS-CoV-2 N1 fw | 5′-GACCCCAAAATCAGCGAAAT-3′ | |
| SARS-CoV-2 N1 rv | 5′-TCTGGTTACTGCCAGTTGAATCTG-3′ | |
| Human GAPDH fw | 5′-CTCTGCTCCTCCTGTTCGAC-3′ | |
| Human GAPDH rv | 5′-CAATACGACCAAATCCGTTGAC-3′ | |
| Human IFNβ fw | 5′-ATGACCAACAAGTGTCTCCTCC-3′ | |
| Human IFNβ rv | 5′-GGAATCCAAGCAAGTTGTAGCTC-3′ | |
| Human IL-6 fw | 5′-CAGCCCTGAGAAAGGAGACATG-3′ | |
| Human-IL-6 rv | 5′-GCATCCATCTTTTTCAGCCATC-3′ | |
| Human MxA fw | 5′-GAAGGGCAACTCCTGACAG-3′ | |
| Human MxA rv | 5′-GTTTCCGAAGTGGACATCGCA-3‘ | |
| Human IP10 fw | 5′-CCAGAATCGAAGGCCATCAA-3′ | |
| Human IP10 rv | 5′-TTTCCTTGCTAACTGCTTTCAG-3´ | |
| Human IFNλ1 fw | 5′-CGCCTTGGAAGAGTCACTCA-3′ | |
| Human IFNλ1 rv | 5′-GAAGCCTCAGGTCCCAATTC-3′ | |
| Human IFNλ2/3 fw | 5′-AGTTCCGGGCCTGTATCCAG-3′ | |
| Human IFNλ2/3 rv | 5′-GAGCCGGTACAGCCAATGGT-3′ | |
| Human TNFα fw | 5′-GGAGAAGGGTGACCGACTCA-3′ | |
| Human TNFα rv | 5′-CTGCCCAGACTCGGCAA-3′ | |
| Human TRAIL fw | 5′-GTCTCTCTGTGTGGCTGTAACTTACG-3′ | |
| Human TRAIL rv | 5′-AAACAAGCAATGCCACTTTTGG-3′ | |
| Human OAS fw | 5′-GATCTCAGAAATACCCCAGCCA-3′ | |
| Human OAS rv | 5′-AGCTACCTCGGAAGCACCTT-3′ | |
| Human ACE2 fw | 5′-TTCCTGCTCAAACAAGCAC-3′ | |
| Human ACE2 rv | 5′-TTCCACCACCCCAACTATC-3′ | |
| Human TMPRSS2 fw | 5′-CCTCTAACTGGTGTGATGGCGT-3′ | |
| Human TMPRSS2 rv | 5′-TGCCAGGACTTCCTCTGAGATG-3′ | |
| SARS-CoV-2 nsp3 fw | 5′-TTCTGCTGCTCTTCAACCTGA-3′ | |
| SARS-CoV-2 nsp3 rv | 5′-ATAGTCTGAACAACTGGTGTAAGT-3′ | |
| SARS-CoV-2 RdRp fw | 5′-ACGCTCAAAGCTACTGAGGAGAC-3′ | OriGene #HP234774 |
| SARS-CoV-2 RdRp rv | 5′-GGTCTAGGTTTACCAACTTCCC-3′ | |
| SARS-CoV-2 Spike fw | 5′-CAACTGAAATCTATCAGGCCG-3′ | OriGene #HP234776 |
| SARS-CoV-2 Spike rv | 5′-ACCAACACCATTAGTGGGTTG-3′ | |
| SARS-CoV-2 lead fw | 5′-CGATCTCTTGTAGATCTGTTCTC-3′ | [53] |
| Spike subgenomic rv | 5′-GAATTAGTGTATGCAGGGGGTAA-3′ | |
| E subgenomic rv | 5′-ATATTGCAGCAGTACGCACACA-3′ | [53] |
| M subgenomic rv | 5′-CAAATCCATGTAAGGAATAGGAAACC-3′ | |
| N1 subgenomic rv | 5′-TCTGGTTACTGCCAGTTGAATCTG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Häring, C.; Jungwirth, J.; Schroeder, J.; Löffler, B.; Engert, B.; Ehrhardt, C. The Local Anaesthetic Procaine Prodrugs ProcCluster® and Procaine Hydrochloride Impair SARS-CoV-2 Replication and Egress In Vitro. Int. J. Mol. Sci. 2023, 24, 14584. https://doi.org/10.3390/ijms241914584
Häring C, Jungwirth J, Schroeder J, Löffler B, Engert B, Ehrhardt C. The Local Anaesthetic Procaine Prodrugs ProcCluster® and Procaine Hydrochloride Impair SARS-CoV-2 Replication and Egress In Vitro. International Journal of Molecular Sciences. 2023; 24(19):14584. https://doi.org/10.3390/ijms241914584
Chicago/Turabian StyleHäring, Clio, Johannes Jungwirth, Josefine Schroeder, Bettina Löffler, Beatrice Engert, and Christina Ehrhardt. 2023. "The Local Anaesthetic Procaine Prodrugs ProcCluster® and Procaine Hydrochloride Impair SARS-CoV-2 Replication and Egress In Vitro" International Journal of Molecular Sciences 24, no. 19: 14584. https://doi.org/10.3390/ijms241914584
APA StyleHäring, C., Jungwirth, J., Schroeder, J., Löffler, B., Engert, B., & Ehrhardt, C. (2023). The Local Anaesthetic Procaine Prodrugs ProcCluster® and Procaine Hydrochloride Impair SARS-CoV-2 Replication and Egress In Vitro. International Journal of Molecular Sciences, 24(19), 14584. https://doi.org/10.3390/ijms241914584