


Proteome Profiling of Canine Epididymal Fluid: In Search of Protein Markers of Epididymal Sperm Motility

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Epididymal Spermatozoa Quality Assessment
2.2. Epididymal Fluid Protein Mass Spectrometry Analysis
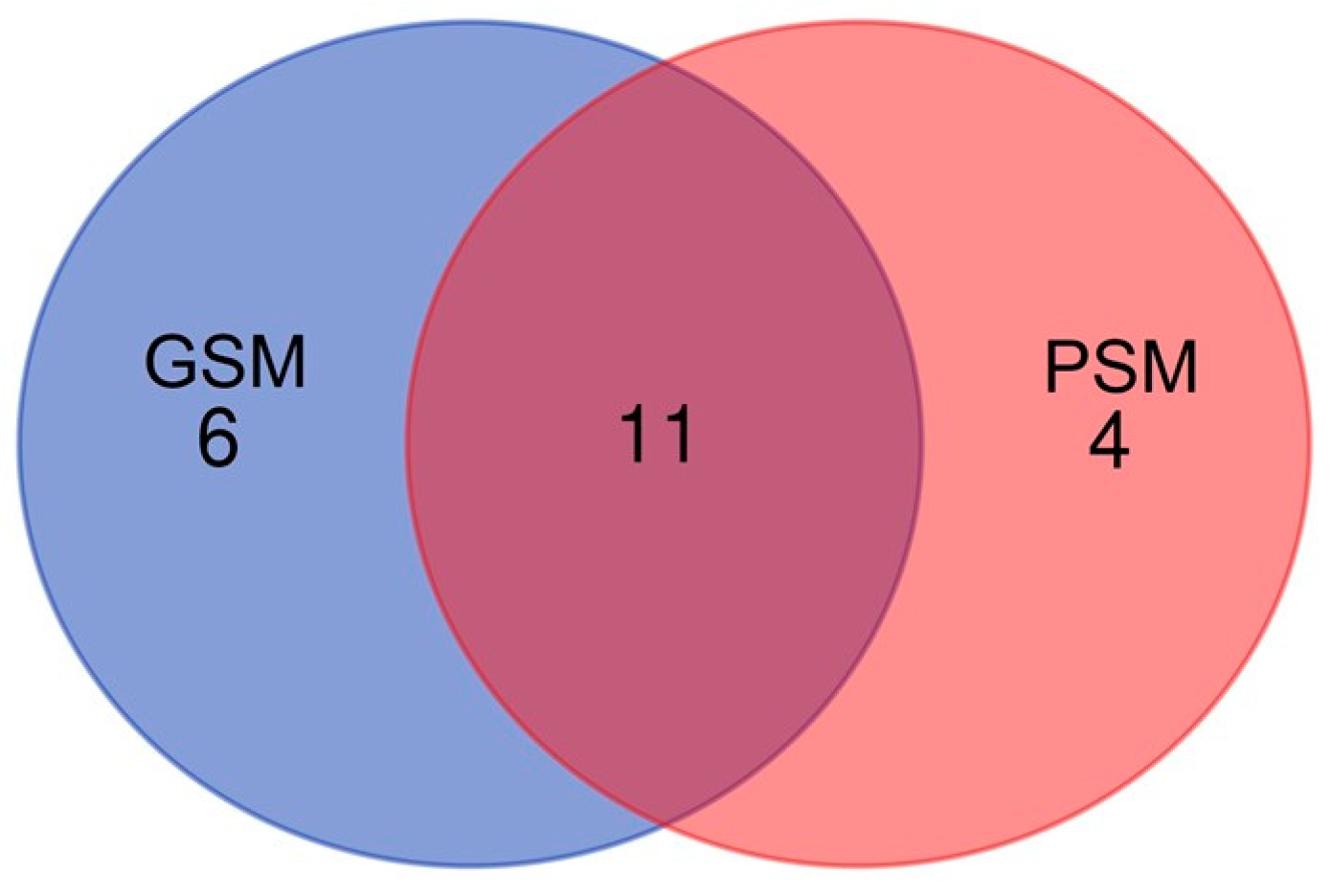
2.2.1. Qualitative Analysis
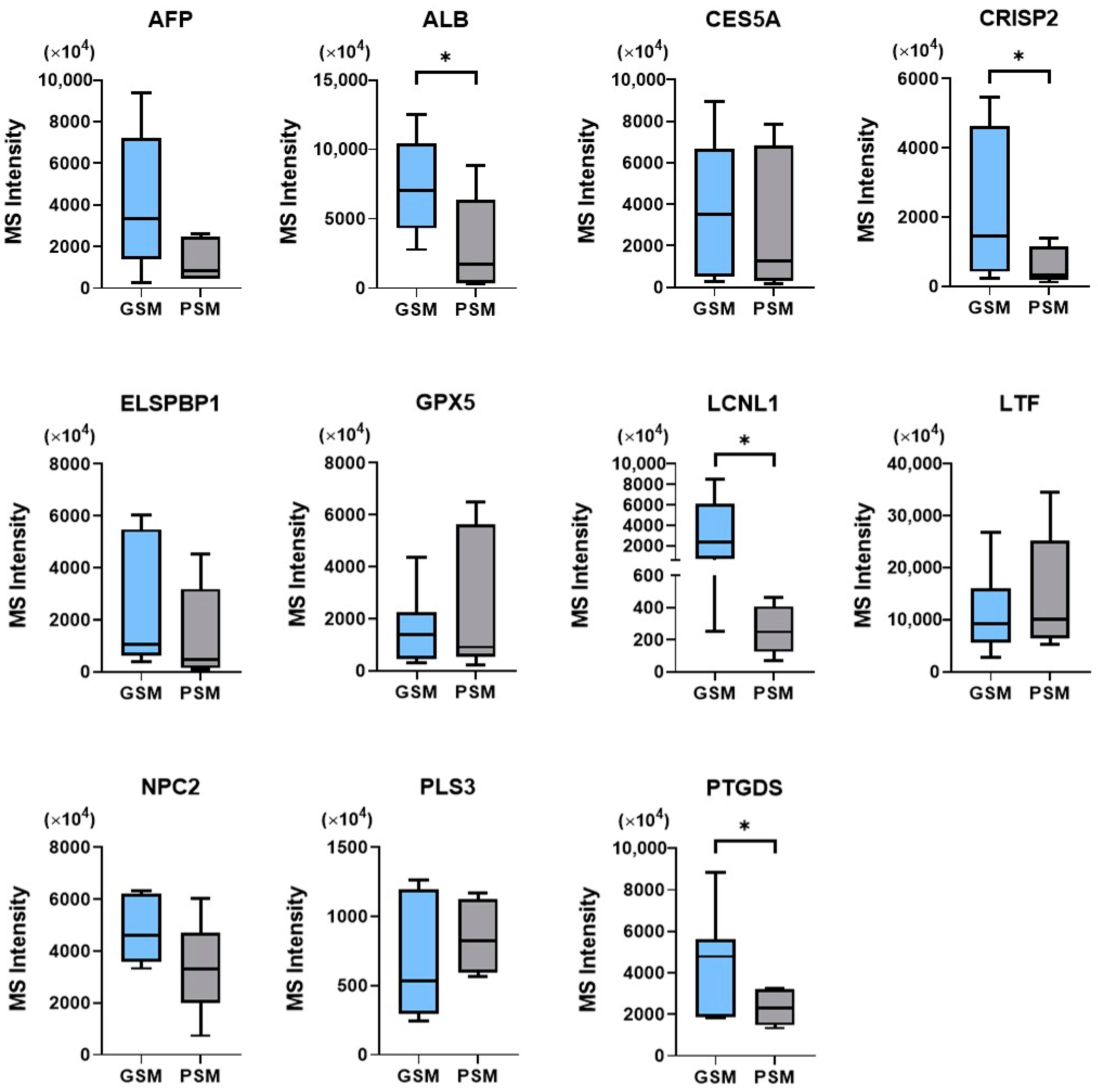
2.2.2. Semi-Quantitative Analysis
2.3. Bioinformatic Analysis
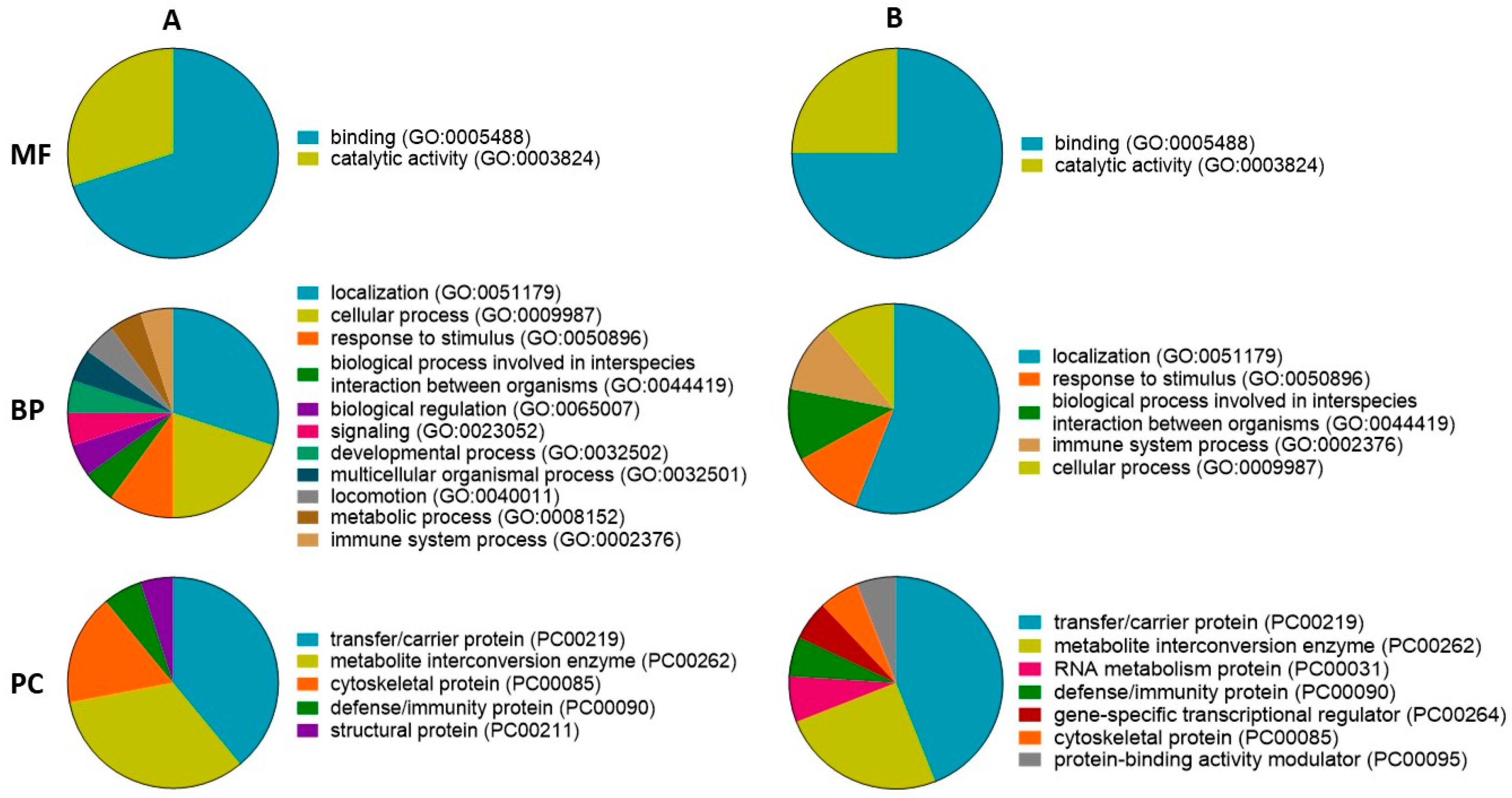
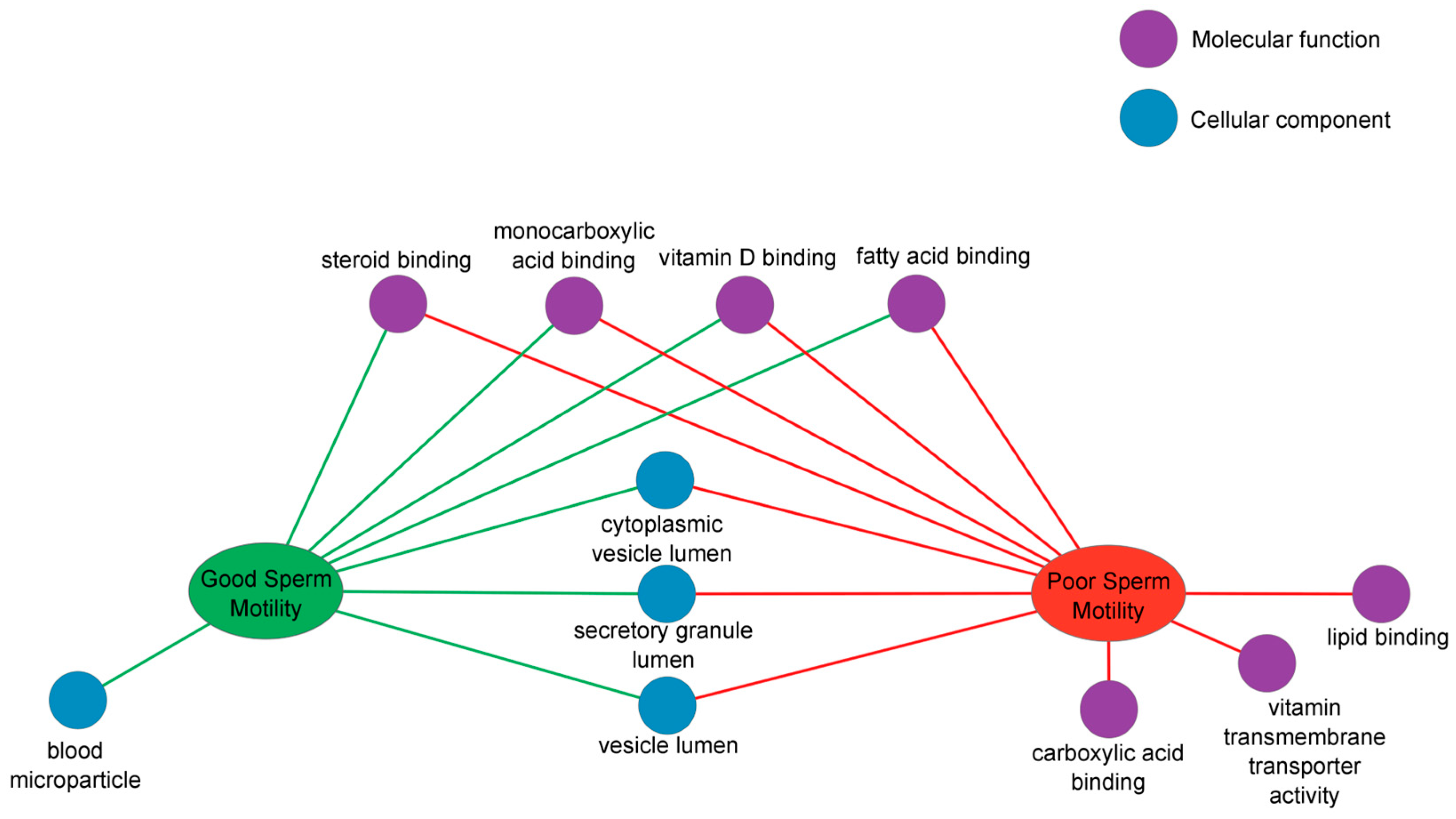
2.3.1. Gene Ontology and ToppCluster Analysis
2.3.2. The Kyoto Encyclopedia of Genes and Genomes Pathway Analysis
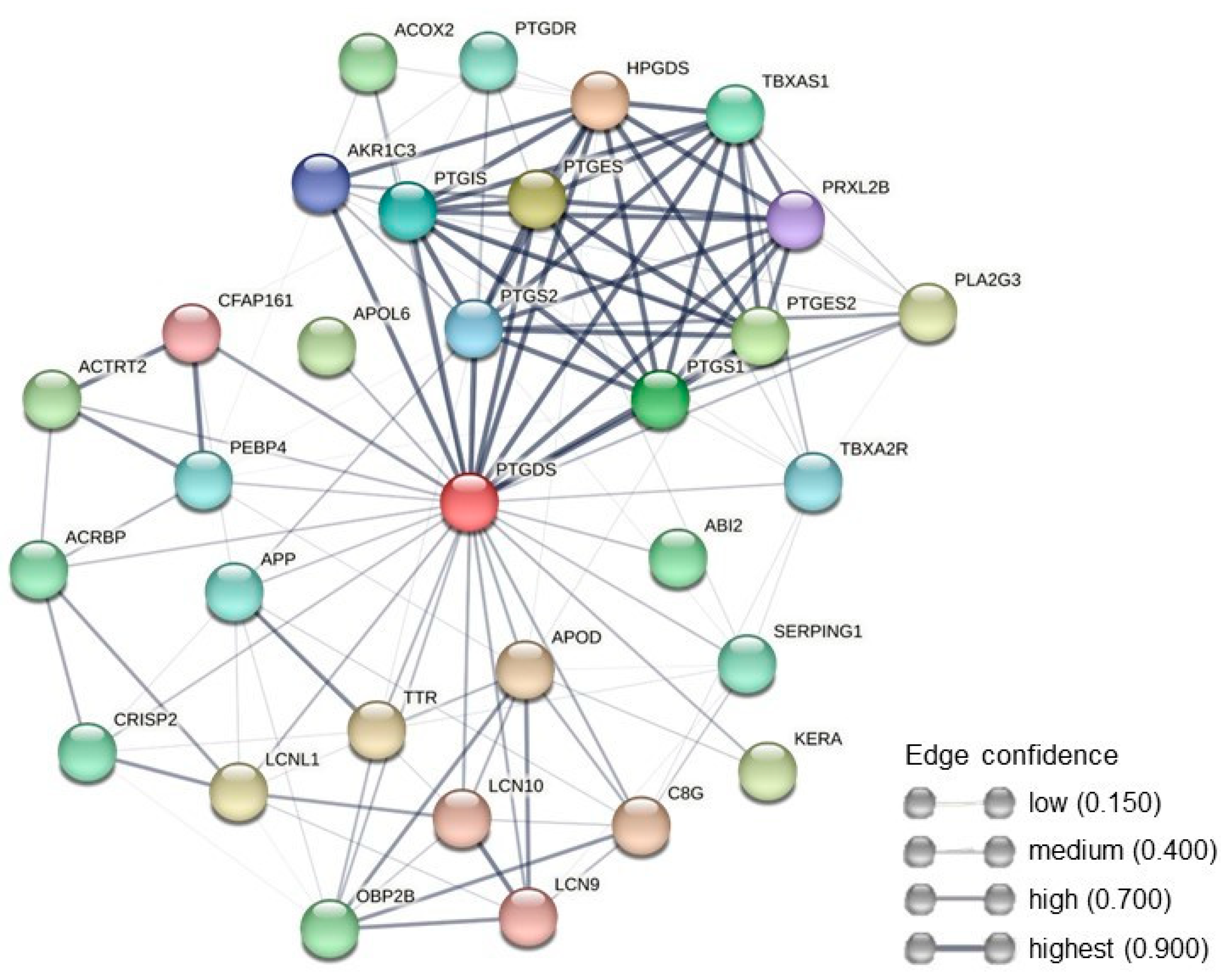
2.3.3. Protein–Protein Interaction Network Analysis
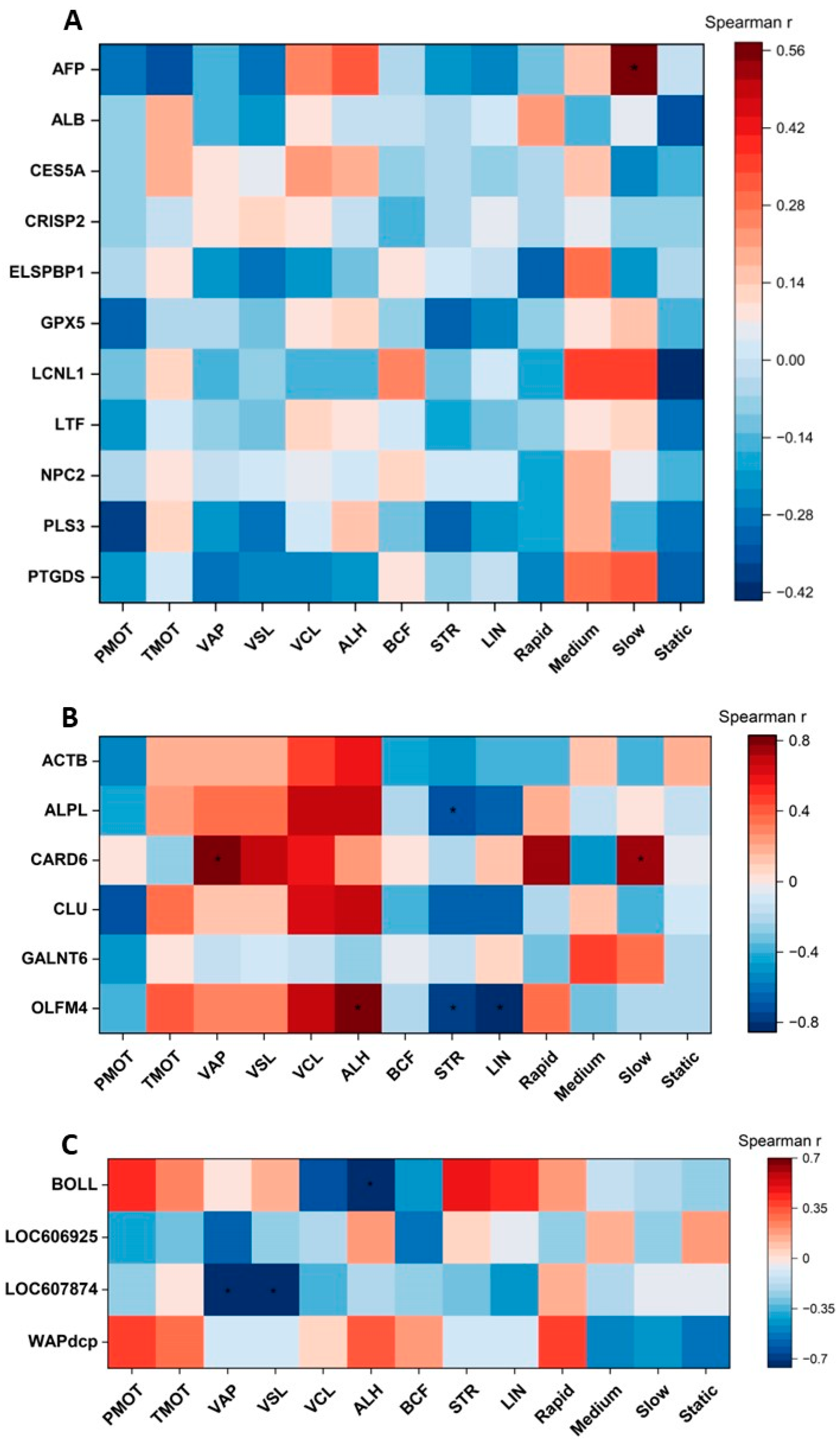
2.4. Correlations between Selected Cauda Epididymal Fluid Proteins and Epididymal Spermatozoa Motion Parameters
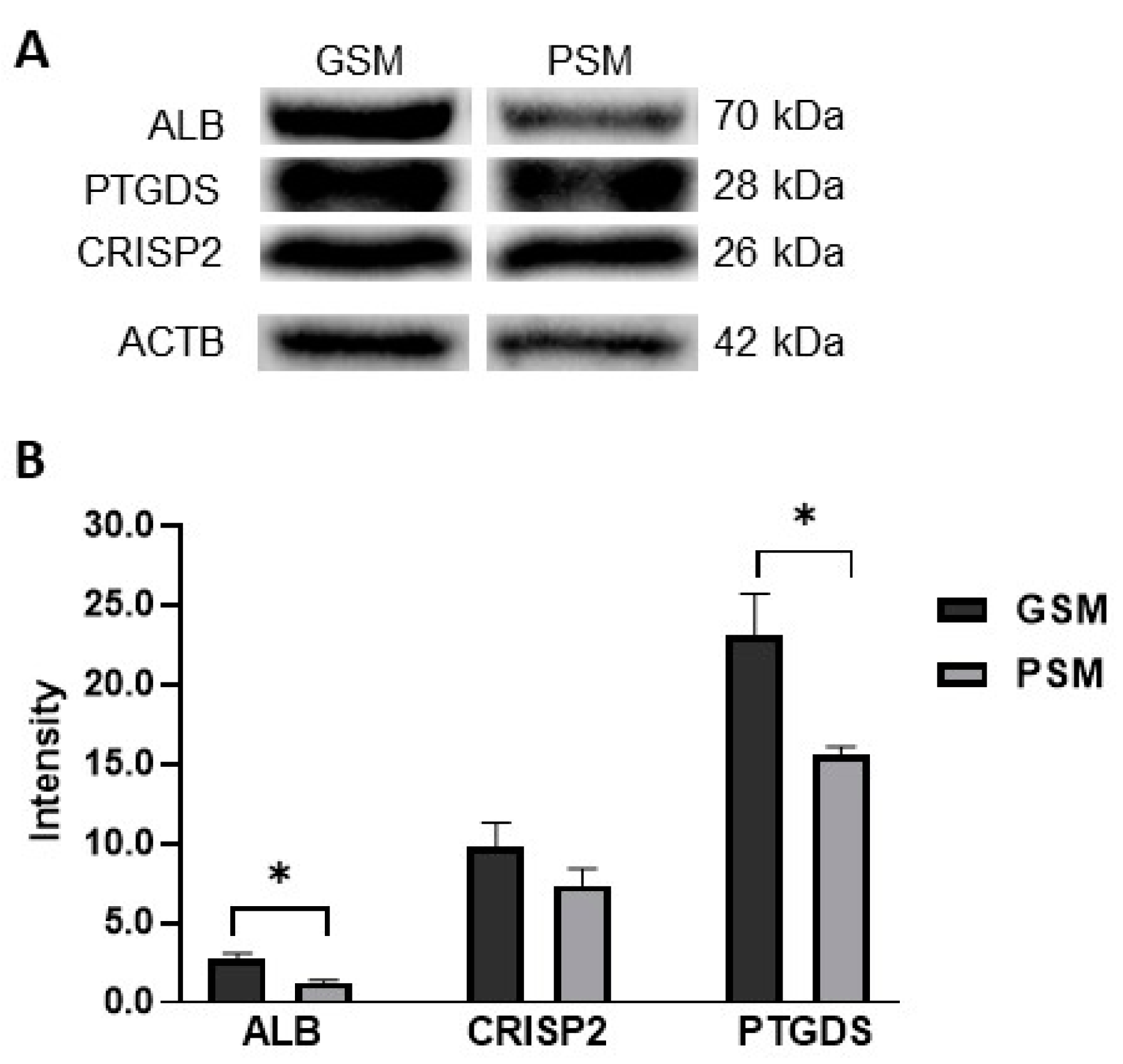
2.5. Validation of Results by Western Blot
3. Discussion
4. Materials and Methods
4.1. Chemicals and Media
4.2. Animals
4.3. Cauda Epididymal Semen Collection
4.4. Cauda Epididymal Spermatozoa Quality Assessment
4.5. Cauda Epididymal Fluid Isolation
4.6. Preliminary Sample Preparation for Proteomic Analysis
4.6.1. Total Protein Content Measurement
4.6.2. Sample Preparation for NanoUPLC-Q-TOF/MS Analysis
4.6.3. In-Solution Trypsin Digestion
4.7. NanoUPLC-Q-TOF/MS Analysis
4.8. Bioinformatic Analyses and Imaging of Data
4.9. Western Blot Analysis
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Björkgren, I.; Sipilä, P. The impact of epididymal proteins on sperm function. Reproduction 2019, 158, R155–R167. [Google Scholar] [CrossRef]
- de Souza, A.P.B.; Schorr-Lenz, A.M.; Lucca, F.; Bustamante-Filho, I.C. The epididymis and its role on sperm quality and male fertility. Anim. Reprod. 2017, 14, 1234–1244. [Google Scholar] [CrossRef]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The role of the epididymis and the contribution of epididymosomes to mammalian reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, J.L.; Dacheux, F. New insights into epididymal functions in relation to sperm maturation. Reproduction 2014, 147, R27–R42. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, M.G.; Visconti, P.E. Molecular changes and signaling events occurring in spermatozoa during epididymal maturation. Andrology 2017, 5, 204–218. [Google Scholar] [CrossRef]
- Dacheux, J.L.; Gatti, J.L.; Dacheux, F. Contribution of epididymal secretory proteins for spermatozoa maturation. Microsc. Res. Tech. 2003, 61, 7–17. [Google Scholar] [CrossRef]
- Bertol, M.A.F. Cryopreservation of Epididymal Sperm. In Cryopreservation in Eukaryotes; Marco-Jiménez, F., Akdemir, H., Eds.; IntechOpen Limited: London, UK, 2016; pp. 121–132. ISBN 978–953-51-2780-2. [Google Scholar]
- Zhao, C.; Huo, R.; Wang, F.Q.; Lin, M.; Zhou, Z.M.; Sha, J.H. Identification of several proteins involved in regulation of sperm motility by proteomic analysis. Fertil. Steril. 2007, 87, 436–438. [Google Scholar] [CrossRef]
- Maňásková-Postlerová, P.; Davidová, N.; Jonáková, V. Biochemical and binding characteristics of boar epididymal fluid proteins. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2011, 879, 100–106. [Google Scholar] [CrossRef]
- Dcunha, R.; Hussein, R.S.; Ananda, H.; Kumari, S.; Adiga, S.K.; Kannan, N.; Zhao, Y.; Kalthur, G. Current insights and latest updates in sperm motility and associated applications in assisted reproduction. Reprod. Sci. 2020, 29, 7–25. [Google Scholar] [CrossRef]
- World Health Organization. Available online: https://www.who.int/news/item/04-04-2023-1-in-6-people-globally-affected-by-infertility (accessed on 7 July 2023).
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2021, 53, e13711. [Google Scholar] [CrossRef]
- Chakraborty, S.; Saha, S. Understanding sperm motility mechanisms and the implication of sperm surface molecules in promoting motility. Middle East Fertil. Soc. J. 2022, 27, 4. [Google Scholar] [CrossRef]
- Das, S.; Saha, S.; Majumder, G.C.; Dungdung, S.R. Purification and characterization of a sperm motility inhibiting factor from caprine epididymal plasma. PLoS ONE 2010, 5, e12039. [Google Scholar] [CrossRef] [PubMed]
- Plante, G.; Manjunath, P. Murine binder of sperm protein homolog 1: A new player in HDL-induced capacitation. Reproduction 2015, 149, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Eskandari-Shahraki, M.; Prud’homme, B.; Bergeron, F.; Manjunath, P. Epididymal proteins Binder of Sperm Homologs 1 and 2 (BSPH1/2) are dispensable for male fertility and sperm motility in mice. Sci. Rep. 2020, 10, 8982. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Ahmed, S.; Liu, J.; Ahmed, S.; Quansah, E.; Solangi, T.H.; Wu, Y.; Yangliu, Y.; Wang, H.; Zhu, J.; et al. Comparative iTRAQ proteomics identified proteins associated with sperm maturation between yak and cattleyak epididymis. BMC Vet. Res. 2021, 17, 255. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, G.A. New insights into epididymal biology and function. Hum. Reprod. Update 2009, 15, 213–227. [Google Scholar] [CrossRef]
- Dacheux, J.L.; Belghazi, M.; Lanson, Y.; Dacheux, F. Human epididymal secretome and proteome. Mol. Cell Endocrinol. 2006, 250, 36–42. [Google Scholar] [CrossRef]
- Weber, A.; Argenti, L.E.; de Souza, A.P.B.; Santi, L.; Beys-da-Silva, W.O.; Yates, J.R., 3rd; Bustamante-Filho, I.C. Ready for the journey: A comparative proteome profiling of porcine cauda epididymal fluid and spermatozoa. Cell Tissue Res. 2020, 379, 389–405. [Google Scholar] [CrossRef]
- Moura, A.A.; Souza, C.E.; Stanley, B.A.; Chapman, D.A.; Killian, G.J. Proteomics of cauda epididymal fluid from mature Holstein bulls. J. Proteomics 2010, 73, 2006–2020. [Google Scholar] [CrossRef]
- Belleannée, C.; Labas, V.; Teixeira-Gomes, A.P.; Gatti, J.L.; Dacheux, J.L.; Dacheux, F. Identification of luminal and secreted proteins in bull epididymis. J. Proteomics 2011, 74, 59–78. [Google Scholar] [CrossRef]
- Westfalewicz, B.; Dietrich, M.A.; Mostek, A.; Partyka, A.; Bielas, W.; Niżański, W.; Ciereszko, A. Identification and functional analysis of bull (Bos taurus) cauda epididymal fluid proteome. J. Dairy Sci. 2017, 100, 6707–6719. [Google Scholar] [CrossRef] [PubMed]
- Zoca, S.M.; Northrop-Albrecht, E.J.; Walker, J.A.; Cushman, R.A.; Perry, G.A. Proteomics dataset of epididymal fluid, seminal plasma, and proteins loosely attached to epididymal and ejaculated sperm from Angus bulls. Data Brief 2022, 42, 108150. [Google Scholar] [CrossRef]
- Gatti, J.L.; Métayer, S.; Belghazi, M.; Dacheux, F.; Dacheux, J.L. Identification, proteomic profiling, and origin of ram epididymal fluid exosome-like vesicles. Biol. Reprod. 2005, 72, 1452–1465. [Google Scholar] [CrossRef] [PubMed]
- van Tilburg, M.; Sousa, S.; Lobo, M.D.P.; Monteiro-Azevedo, A.C.O.M.; Azevedo, R.A.; Araújo, A.A.; Moura, A.A. Mapping the major proteome of reproductive fluids and sperm membranes of rams: From the cauda epididymis to ejaculation. Theriogenology 2021, 159, 98–107. [Google Scholar] [CrossRef]
- Liu, X.; Jin, S.H.; Liu, X.X.; Wang, W.J.; Liu, F.J. Proteome profiling of the sperm maturation milieu in the rhesus monkey (Macaca mulatta) epididymis. Reprod. Fertil. Dev. 2014, 28, 732–741. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.F.; Hoffmann, E.K.; Mills, J.W. The cytoskeleton and cell volume regulation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 130, 385–399. [Google Scholar] [CrossRef]
- Cabello-Agüeros, J.F.; Hernández-González, E.O.; Mújica, A. The role of F-actin cytoskeleton-associated gelsolin in the guinea pig capacitation and acrosome reaction. Cell Motil. Cytoskelet. 2003, 56, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Naresh, S. Effect of cooling (4 °C) and cryopreservation on cytoskeleton actin and protein tyrosine phosphorylation in buffalo spermatozoa. Cryobiology 2016, 72, 7–13. [Google Scholar] [CrossRef]
- Zmudzinska, A.; Wisniewski, J.; Mlynarz, P.; Olejnik, B.; Mogielnicka-Brzozowska, M. Age-dependent variations in functional quality and proteomic characteristics of canine (Canis lupus familiaris) epididymal spermatozoa. Int. J. Mol. Sci. 2022, 23, 9143. [Google Scholar] [CrossRef]
- Frenette, G.; Dubé, J.Y.; Tremblay, R.R. Origin of alkaline phosphatase of canine semen. Arch. Androl. 1986, 16, 235–241. [Google Scholar] [CrossRef]
- Johnston, S.D. Performing a complete canine semen evaluation in a small animal hospital. Vet. Clin. North Am. Small Anim. Pract. 1991, 21, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.S. Localization of alkaline phosphatase and NADH diaphorase in the principal cells of the guinea pig epididymis. Acta Anat. 1983, 116, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Bouchier-Hayes, L.; Martin, S.J. CARD games in apoptosis and immunity. EMBO Rep. 2002, 3, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Dufner, A.; Pownall, S.; Mak, T.W. Caspase recruitment domain protein 6 is a microtubule-interacting protein that positively modulates NF-κB activation. Proc. Natl. Acad. Sci. USA 2006, 103, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Dacheux, J.L.; Belleannée, C.; Jones, R.; Labas, V.; Belghazi, M.; Guyonnet, B.; Druart, X.; Gatti, J.L.; Dacheux, F. Mammalian epididymal proteome. Mol. Cell Endocrinol. 2009, 306, 45–50. [Google Scholar] [CrossRef]
- Fouchécourt, S.; Métayer, S.; Locatelli, A.; Dacheux, F.; Dacheux, J.L. Stallion epididymal fluid proteome: Qualitative and quantitative characterization; secretion and dynamic changes of major proteins. Biol. Reprod. 2000, 62, 1790–1803. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, J.K.; Edwards, C.A.; Xu, Z.; Taichman, R.; Wang, C.Y. Clusterin inhibits apoptosis by interacting with activated Bax. Nat. Cell Biol. 2005, 7, 909–915. [Google Scholar] [CrossRef]
- Kierszenbaum, A.L.; Rivkin, E.; Tres, L.L. Acroplaxome, an F-actin-keratin-containing plate, anchors the acrosome to the nucleus during shaping of the spermatid head. Mol. Biol. Cell 2003, 14, 4628–4640. [Google Scholar] [CrossRef]
- Raman, J.; Guan, Y.; Perrine, C.L.; Gerken, T.A.; Tabak, L.A. UDP-N-acetyl-α-d galactosamine:polypeptide N-acetylgalactosaminyltransferases: Completion of the family tree. Glycobiology 2012, 22, 768–777. [Google Scholar] [CrossRef]
- Takasaki, N.; Tachibana, K.; Ogasawara, S.; Matsuzaki, H.; Hagiuda, J.; Ishikawa, H.; Mochida, K.; Inoue, K.; Ogonuki, N.; Ogura, A.; et al. A heterozygous mutation of GALNTL5 affects male infertility with impairment of sperm motility. Proc. Natl. Acad. Sci. USA 2014, 111, 1120–1125. [Google Scholar] [CrossRef]
- Li, H.; Jiang, Q.; Ju, Z.H.; Huang, J.M.; Wang, X.G.; Yang, C.H.; Sun, Y.; Yang, G.W.; Zhong, J.F.; Wang, C.F. A novel splice variant of the bovine GALNTL5 gene identified in Chinese Holstein bull testis tissue and its mRNA expression. Genet. Mol. Res. 2016, 15, gmr.15027723. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Yang, H.; Zhang, Y.; Ren, C.; Nie, H.; Fan, Y.; Zhou, W.; Wang, S.; Feng, X.; Wang, F. Characterization of GALNTL5 gene sequence and expression in ovine testes and sperm. Theriogenology 2017, 95, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, W.; Chen, W.; Zhu, J.; Deng, C.X.; Rodgers, G.P. Olfactomedin 4 deficiency promotes prostate neoplastic progression and is associated with upregulation of the hedgehog-signaling pathway. Sci. Rep. 2015, 5, 16974. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Rodgers, G.P. Olfactomedin 4 expression and functions in innate immunity, inflammation, and cancer. Cancer Metastasis Rev. 2016, 35, 201–212. [Google Scholar] [CrossRef]
- Anholt, R.R.H. Olfactomedin proteins: Central players in development and disease. Front. Cell Dev. Biol. 2014, 2, 6. [Google Scholar] [CrossRef]
- Thimon, V.; Frenette, G.; Saez, F.; Thabet, M.; Sullivan, R. Protein composition of human epididymosomes collected during surgical vasectomy reversal: A proteomic and genomic approach. Hum. Reprod. 2008, 23, 1698–1707. [Google Scholar] [CrossRef]
- Peng, F.; Muhuitijiang, B.; Zhou, J.; Liang, H.; Zhang, Y.; Zhou, R. An artificial neural network model to diagnose non-obstructive azoospermia based on RNA-binding protein-related genes. Aging Albany NY 2023, 15, 3120–3140. [Google Scholar] [CrossRef]
- Lin, Y.M.; Kuo, P.L.; Lin, Y.H.; Teng, Y.N.; Nan Lin, J.S. Messenger RNA transcripts of the meiotic regulator BOULE in the testis of azoospermic men and their application in predicting the success of sperm retrieval. Hum. Reprod. 2005, 20, 782–788. [Google Scholar] [CrossRef]
- Kostova, E.; Yeung, C.H.; Luetjens, C.M.; Brune, M.; Nieschlag, E.; Gromoll, J. Association of three isoforms of the meiotic BOULE gene with spermatogenic failure in infertile men. Mol. Hum. Reprod. 2007, 13, 85–93. [Google Scholar] [CrossRef]
- Urrutia, R. KRAB-containing zinc-finger repressor proteins. Genome Biol. 2003, 4, 231. [Google Scholar] [CrossRef]
- Lorenz, P.D.S.; Dietmann, S.; Wilhelm, T.; Koczan, D.; Autran, S.; Gad, S.; Wen, G.; Ding, G.; Li, Y.; Rousseau-Merck, M.F.; et al. The ancient mammalian KRAB zinc finger gene cluster on human chromosome 8q24.3 illustrates principles of C2H2 zinc finger evolution associated with unique expression profiles in human tissues. BMC Genom. 2010, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Choi, H.; Kwon, J.T.; Kim, J.; Jeong, J.; Kim, J.; Ham, S.; Cho, B.N.; Yoo, Y.J.; Cho, C. Identification and characterization of reproductive KRAB-ZF genes in mice. Gene 2015, 565, 45–55. [Google Scholar] [CrossRef]
- Sutton, H.G.; Fusco, A.; Cornwall, G.A. Cystatin-related epididymal spermatogenic protein colocalizes with luteinizing hormone-β protein in mouse anterior pituitary gonadotropes. Endocrinology 1999, 140, 2721–2732. [Google Scholar] [CrossRef]
- Pera, I.; Ivell, R.; Kirchhoff, C. Regional variation of gene expression in the dog epididymis as revealed by in-situ transcript hybridization. Int. J. Androl. 1994, 17, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., 3rd; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Zmudzinska, A.; Bromke, M.A.; Strzezek, R.; Zielinska, M.; Olejnik, B.; Mogielnicka-Brzozowska, M. Proteomic Analysis of Intracellular and Membrane-Associated Fractions of Canine (Canis lupus familiaris) Epididymal Spermatozoa and Sperm Structure Separation. Animals 2022, 12, 772. [Google Scholar] [CrossRef] [PubMed]
- Hardy, D.M.; Huang, T.T., Jr.; Driscoll, W.J.; Tung, K.S.K.; Wild, G.C. Purification and characterization of the primary acrosomal autoantigen of guinea pig epididymal spermatozoa. Biol. Reprod. 1988, 38, 423–437. [Google Scholar] [CrossRef]
- Kasahara, M.; Gutknecht, J.; Brew, K.; Spurr, N.; Goodfellow, P.N. Cloning and mapping of a testis-specific gene with sequence similarity to a sperm-coating glycoprotein gene. Genomics 1989, 5, 527–534. [Google Scholar] [CrossRef]
- Foster, J.A.; Gerton, G.L. Autoantigen 1 of the guinea pig sperm acrosome is the homologue of mouse Tpx-1 and human TPX1 and is a member of the cysteine-rich secretory protein (CRISP) family. Mol. Reprod. Dev. 1996, 44, 221–229. [Google Scholar] [CrossRef]
- O’Bryan, M.K.; Loveland, K.L.; Herszfeld, D.; McFarlane, J.R.; Hearn, M.T.; de Kretser, D.M. Identification of a rat testis-specific gene encoding a potential rat outer dense fibre protein. Mol. Reprod. Dev. 1998, 50, 313–322. [Google Scholar] [CrossRef]
- Reddy, T.; Gibbs, G.M.; Merriner, D.J.; Kerr, J.B.; O’Bryan, M.K. Cysteine-rich secretory proteins are not exclusively expressed in the male reproductive tract. Dev. Dyn. 2008, 237, 3313–3323. [Google Scholar] [CrossRef]
- Lim, S.; Kierzek, M.; O’Connor, A.E.; Brenker, C.; Merriner, D.J.; Okuda, H.; Volpert, M.; Gaikwad, A.; Bianco, D.; Potter, D.; et al. CRISP2 is a regulator of multiple aspects of sperm function and male fertility. Endocrinology 2019, 160, 915–924. [Google Scholar] [CrossRef]
- Jamsai, D.; Reilly, A.; Smith, S.J.; Gibbs, G.M.; Baker, H.W.; McLachlan, R.I.; de Kretser, D.M.; O’Bryan, M.K. Polymorphisms in the human cysteine-rich secretory protein 2 (CRISP2) gene in Australian men. Hum. Reprod. 2008, 23, 2151–2159. [Google Scholar] [CrossRef]
- Zhou, J.H.; Zhou, Q.Z.; Lyu, X.M.; Zhu, T.; Chen, Z.J.; Chen, M.K.; Xia, H.; Wang, C.Y.; Qi, T.; Li, X.; et al. The expression of cysteine-rich secretory protein 2 (CRISP2) and its specific regulator miR-27b in the spermatozoa of patients with asthenozoospermia. Biol. Reprod. 2015, 92, 28. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Metzger, J.; Martinsson, G.; Sieme, H.; Distl, O. Genome-wide association study for semen quality traits in German Warmblood stallions. Anim. Reprod. Sci. 2016, 171, 81–86. [Google Scholar] [CrossRef]
- Salier, J.P. Chromosomal location, exon/intron organization and evolution of lipocalin genes. Biochim. Biophys. Acta 2000, 1482, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Hamil, K.G.; Liu, Q.; Sivashanmugam, P.; Anbalagan, M.; Yenugu, S.; Soundararajan, R.; Grossman, G.; Rao, A.J.; Birse, C.E.; Ruben, S.M.; et al. LCN6, a novel human epididymal lipocalin. Reprod. Biol. Endocrinol. 2003, 1, 112. [Google Scholar] [CrossRef] [PubMed]
- Thimon, V.; Koukoui, O.; Calvo, E.; Sullivan, R. Region-specific gene expression profiling along the human epididymis. Mol. Hum. Reprod. 2007, 13, 691–704. [Google Scholar] [CrossRef]
- Elzanaty, S.; Malm, J. Effects of ejaculation-to-analysis delay on levels of markers of epididymal and accessory sex gland functions and sperm motility. J. Androl. 2007, 28, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Macıas-Garcıa, B.; Gonzalez-Fernández, L.; Loux, S.C.; Rocha, A.M.; Guimarães, T.; Pena, F.J.; Varner, D.D.; Hinrichs, K. Effect of calcium, bicarbonate, and albumin on capacitation-related events in equine sperm. Reproduction 2015, 149, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Arroteia, K.F.; Barbieri, M.F.; Souza, G.H.; Tanaka, H.; Eberlin, M.N.; Hyslop, S.; Alvares, L.E.; Pereira, L.A. Albumin is synthesized in epididymis and aggregates in a high molecular mass glycoprotein complex involved in sperm-egg fertilization. PLoS ONE 2014, 9, e103566. [Google Scholar] [CrossRef] [PubMed]
- Mogielnicka-Brzozowska, M.; Fraser, L.; Dziekońska, A.; Gackowska, K.; Sobiewska, M.; Kuzborska, A.; Majewska, A.M.; Filipowicz, K.; Kordan, W. Identification of proteoforms of albumin and kallikrein in stallion seminal plasma and their correlations with sperm motility. Pol. J. Vet. Sci. 2019, 22, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Stewart, A.J.; Sadler, P.J.; Pinheiro, T.J.; Blindauer, C.A. Albumin as a zinc carrier: Properties of its high-affinity zinc-binding site. Biochem. Soc. Trans. 2008, 36, 1317–1321. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett 2008, 582, 1783–1787. [Google Scholar] [CrossRef]
- Candenas, L.; Chianese, R. Exosome composition and seminal plasma proteome: A promising source of biomarkers of male infertility. Int. J. Mol. Sci. 2020, 21, 7022. [Google Scholar] [CrossRef] [PubMed]
- Mogielnicka-Brzozowska, M.; Prochowska, S.; Niżański, W.; Bromke, M.A.; Wiśniewski, J.; Olejnik, B.; Kuzborska, A.; Fraser, L.; Młynarz, P.; Kordan, W. Proteome of cat semen obtained after urethral catheterization. Theriogenology 2020, 141, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Shaw, N.; Li, Y.; Zhao, Y.; Zhang, R.; Liu, Z.J. Structure-function analysis of human l-prostaglandin D synthase bound with fatty acid molecules. FASEB J. 2010, 24, 4668–4677. [Google Scholar] [CrossRef]
- Stillwell, W.; Wassall, S.R. Interactions of retinoids with phospholipid membranes: Optical spectroscopy. Methods Enzymol. 1990, 189, 373–382. [Google Scholar] [CrossRef]
- Breitbart, H. Intracellular calcium regulation in sperm capacitation and acrosomal reaction. Mol. Cell. Endocrinol. 2002, 187, 139–144. [Google Scholar] [CrossRef]
- Kao, S.H.; Chao, H.T.; Chen, H.W.; Hwang, T.I.S.; Liao, T.L.; Wei, Y.H. Increase of oxidative stress in human sperm with lower motility. Fertil. Steril. 2008, 89, 1183–1190. [Google Scholar] [CrossRef]
- Chen, D.Y.; Zhu, M.Y.; Cui, Y.D.; Huang, T.H. Relationship between contents of lipocalin-type prostaglandin D synthase on the surface of infertility sperm and in seminal plasma. Biochemistry 2007, 72, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Ramos Angrimani, D.S.; Nichi, M.; Losano, J.D.A.; Lucio, C.F.; Lima Veiga, G.A.; Franco, M.V.M.J.; Vannucchi, C.I. Fatty acid content in epididymal fluid and spermatozoa during sperm maturation in dogs. J. Anim. Sci. Biotechnol. 2017, 8, 18. [Google Scholar] [CrossRef]
- Morelle, W.; Michalski, J.C. Analysis of protein glycosylation by mass spectrometry. Nat. Protoc. 2007, 2, 1585–1602. [Google Scholar] [CrossRef] [PubMed]
- Kaimal, V.; Bardes, E.E.; Tabar, S.C.; Jegga, A.G.; Aronow, B.J. ToppCluster: A multiple gene list feature analyzer for comparative enrichment clustering and network-based dissection of biological systems. Nucleic Acids Res. 2010, 38, W96–W102. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Sionovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sperm Parameters | Good Sperm Motility (n = 13) | Poor Sperm Motility (n = 10) | p-Value |
|---|---|---|---|
| Total motility (TMOT, %) | 92.46 ± 0.4 | 87.9 ± 2.3 | 0.031 |
| Progressive motility (PMOT, %) | 62.08 ± 1.5 | 37.5 ± 5.2 | 0.001 |
| Average path velocity (VAP, µm/s) | 135.9 ± 3.5 | 127.8 ± 7.5 | 0.784 |
| Straight line velocity (VSL, µm/s) | 116.1 ± 3.3 | 99.06 ± 6.3 | 0.030 |
| Curvilinear velocity (VCL, µm/s) | 194.5 ± 5.2 | 217.3 ± 8.8 | 0.011 |
| Amplitude of lateral head displacement (ALH, µm) | 6.48 ± 0.2 | 7.68 ± 0.3 | 0.003 |
| Beat cross frequency (BCF, Hz) | 18.51 ± 1.0 | 14.0 ± 0.5 | 0.001 |
| Straightness (STR, %) | 84.46 ± 0.7 | 76.3 ± 0.8 | 0.001 |
| Linearity (LIN, %) | 61.92 ± 1.6 | 49.1 ± 1.4 | 0.001 |
| Rapid (%) | 82.23 ± 1.6 | 81.38 ± 1.94 | 0.744 |
| Medium (%) | 10.31 ± 1.8 | 8.37 ± 0.88 | 0.405 |
| Slow (%) | 4.31 ± 0.4 | 5.63 ± 0.82 | 0.112 |
| Static (%) | 3.15 ± 0.4 | 4.62 ± 0.73 | 0.080 |
| GSM | ||||
|---|---|---|---|---|
| ID | Pathway Name | Protein Counts | −Log10 (Corrected p-Value) | Protein Names |
| cfa00590 | Arachidonic acid metabolism | 2 | 4.69 × 10−4 | PTGDS|GPX5 |
| cfa04918 | Thyroid hormone synthesis | 2 | 6.47 × 10−4 | GPX5|ALB |
| cfa04390 | Hippo signaling pathway | 2 | 2.79 × 10−3 | ACTB|AFP |
| cfa01100 | Metabolic pathways | 4 | 4.48 × 10−3 | PTGDS|GALNT6|GPX5|ALPL |
| PSM | ||||
| ID | Pathway Name | Protein Counts | −Log10 (Corrected p-Value) | Protein Names |
| cfa00590 | Arachidonic acid metabolism | 2 | 2.16 × 10−4 | PTGDS|GPX5 |
| cfa04918 | Thyroid hormone synthesis | 2 | 2.99 × 10−4 | GPX5|ALB |
| cfa04979 | Cholesterol metabolism | 1 | 1.71 × 10−2 | NPC2 |
| cfa00480 | Glutathione metabolism | 1 | 1.85 × 10−2 | GPX5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cichowska, A.W.; Wisniewski, J.; Bromke, M.A.; Olejnik, B.; Mogielnicka-Brzozowska, M. Proteome Profiling of Canine Epididymal Fluid: In Search of Protein Markers of Epididymal Sperm Motility. Int. J. Mol. Sci. 2023, 24, 14790. https://doi.org/10.3390/ijms241914790
Cichowska AW, Wisniewski J, Bromke MA, Olejnik B, Mogielnicka-Brzozowska M. Proteome Profiling of Canine Epididymal Fluid: In Search of Protein Markers of Epididymal Sperm Motility. International Journal of Molecular Sciences. 2023; 24(19):14790. https://doi.org/10.3390/ijms241914790
Chicago/Turabian StyleCichowska, Aleksandra W., Jerzy Wisniewski, Mariusz A. Bromke, Beata Olejnik, and Marzena Mogielnicka-Brzozowska. 2023. "Proteome Profiling of Canine Epididymal Fluid: In Search of Protein Markers of Epididymal Sperm Motility" International Journal of Molecular Sciences 24, no. 19: 14790. https://doi.org/10.3390/ijms241914790
APA StyleCichowska, A. W., Wisniewski, J., Bromke, M. A., Olejnik, B., & Mogielnicka-Brzozowska, M. (2023). Proteome Profiling of Canine Epididymal Fluid: In Search of Protein Markers of Epididymal Sperm Motility. International Journal of Molecular Sciences, 24(19), 14790. https://doi.org/10.3390/ijms241914790